1. Introduction
A large variety of wet by-products are produced by the food and beverage industries, with annual global amounts as high as one billion tons. Animal husbandry successfully makes use of these by-products as ingredients in animal feed [
1]. The by-products are often stored under anaerobic fermentation in silos. Thus, ensiling is regarded as the most efficient way to preserve wet by-products. Furthermore, ensiled non-food biomass is also regarded as a renewable energy source, being used to produce biofuels.
A significant amount of soybean curd residue (SCR) is generated as a by-product from the manufacturing of tofu, a traditional Asian food prepared by coagulating soy milk. The sum of SCR in China, Korea, and Japan is approximately 3.9 million tons per year, equivalent to the annual amount of tomato pomace generated globally [
2,
3]. Although the low dry matter (DM) content (about 200 g/kg) of wet SCR poses a challenge to its handling and transportation, its high crude protein content (about 250 g/kg DM) makes it an attractive ingredient for use in animal feed. In addition, the storability of SCR can vary greatly between factories and seasons because of large differences in the handling procedures executed by vendors.
During tofu manufacturing, soybeans are milled and mashed. As a result, SCR is more easily compacted than other forage crops. A density of >900 kg/m
3 is difficult to obtain in forage ensiling, even with fine chopping and intensive compaction. However, this can be easily achieved when using SCR, without the help of special devices [
4]. Moreover, the physical properties of SCR may accelerate the establishment of an anaerobic environment, allowing for lactic acid fermentation without the need for additives. As a result of the heating step, during which the soy milk and SCR are separated, the resulting SCR can reach temperatures as high as 80 °C, which may restrict the general bacterial population at the 10
3–5 CFU/g level and can change its bacterial composition under non-farm environmental conditions [
4]. Meanwhile, the major components of soybean sugars are removed from soymilk during processing; hence, the addition of soluble sugars would help increase lactic acid fermentation of SCR silage. Because the moisture content of raw SCR is relatively high, the addition of dried by-products, such as wheat bran and beet pulp (BP), would also help improve the storability of SCR silage.
The industrial scale of tofu manufacturing varies widely across factories. In addition, their locations are scattered, and are separate from the livestock sector. Daily on-site ensiling is difficult to perform, thus sealing is often delayed in practice. This lowers the opportunity for the stable production of a high-quality feed and increases the risk of spoilage due to rancidity, especially in hot summers.
In this study, wet SCR was obtained from two tofu factories (F1 and F2) and ensiled with and without BP on the day of tofu production (prompt sealing (PS)) and 2 days after being piled and unprocessed (delayed sealing (DS)). Ensiling was performed for 6 months since the effects of DS could lead to changes in the microbiota under conditions of long-term storage. The objective was to characterize the microbiota associated with SCR silage stored under PS and DS conditions. Both bacterial and fungal microbiota were assessed using high-throughput amplicon sequencing.
2. Materials and Methods
2.1. Ensiling
Two sets of SCRs were obtained from F1 and F2, and each was further divided into two subsets. The first and second subsets of SCR were subjected to PS and DS ensiling, respectively. Briefly, 300 g of wet SCR was sealed in a plastic bag (Hiryu BN-12; Asahi Kasei Pax, Tokyo, Japan) in triplicate, with or without dried BP (60 g). Air was removed using a vacuum sealer (SQ-303; Asahi Kasei Pax, Tokyo, Japan), and the bags were stored at room temperature. The silos containing the samples were opened after 2 weeks and 3 and 6 months to examine the fermentation products and evaluate the bacterial and fungal microbiota.
2.2. Chemical Composition Analyses and Epiphytic Microbiota Counts
The DM content was determined after oven drying at 60 °C for 48 h. The pH values and fermentation products were determined using water extracts. The lactic acid, acetic acid, and ethanol contents were measured by ion-exclusion polymeric high-performance liquid chromatography with refractive index detection [
5]. For the determination of soluble sugars, freeze-dried samples were extracted using an ethanol/water (80:20,
v/
v) solution. The fructose, glucose, sucrose, maltose, raffinose, and stachyose contents were determined by hydrophilic interaction chromatography with refractive index detection [
6].
The levels of lactic acid bacteria, and yeast and molds were determined using de Man, Rogosa, Sharpe agar and Yeast Malt agar, respectively. The serially diluted plates were incubated at 30 °C for 2 days.
2.3. Microbiota Analyses
Silage samples were added to a 20× volume of sterilized phosphate-buffered saline (pH 7.4). DNA extraction was performed as described by Yu et al. [
7]. DNA purification was performed using a commercial kit (DNeasy Blood & Tissue Kit; Qiagen, Germantown, MD, USA) according to the manufacturer’s instructions.
The resulting DNA was subjected to 2-step polymerase chain reaction (PCR) to generate amplicon libraries. For bacterial MiSeq analysis, primers targeting the V4 region of 16S rRNA genes (forward: 5′-ACACTCTTTCCCTACACGACGCTCTTCCGATCTGTGCCAGCMGCCGCGGTAA-3′; reverse: 5′-GTGACTGGAGTTCAGACGTGTGCTCTTCCGATCTGGACTACHVGGGTWTCTAAT-3′) were used in the first round PCR [
8]. The PCR protocol was as follows: initiation at 94 °C for 2 min, followed by 25 cycles of 94 °C for 30 s, 50 °C for 30 s, and 72 °C for 30 s, and a final elongation at 72 °C for 5 min. For fungal MiSeq analysis, primers targeting the internal transcribed spacers (ITS2) separated by 5.8S (forward: 5′-ACACTCTTTCCCTACACGACGCTCTTCCGATCTGTGARTCATCGARTCTTTG-3′; reverse: 5′-GTGACTGGAGTTCAGACGTGTGCTCTTCCGATCTTCCTCCGCTTATTGATATGC-3′) were employed in the first round PCR [
9]. The PCR protocol was as follows: initiation at 95 °C for 5 min, followed by 30 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 1 min, and a final elongation at 72 °C for 10 min. The products were purified using the Fast Gene Gel/PCR Extraction Kit (NIPPON Genetics Co., Ltd., Tokyo, Japan) and used in the second round PCR with the adapter-attached primers. The second PCR protocol was as follows: initiation at 94 °C for 2 min followed by 10 cycles of 94 °C for 30 s, 59 °C for 30 s, and 72 °C for 30 s, and a final elongation at 72 °C for 5 min. The PCR products were purified as described above.
2.4. Illumina MiSeq Sequencing
The purified amplicons were pair-end sequenced (2 × 250 bp) on an Illumina MiSeq platform at FASMAC Co., Ltd. (Kanagawa, Japan). The raw sequence data were analyzed using Quantitative Insights into Microbial Ecology (version 1.9.0). The 250-bp reads were truncated at any site, with an average quality score under 20. The truncated reads that were shorter than 225 bp were discarded. For primer matching, sequences with overlaps longer than 200 bp were assembled. The final reads obtained after pair-end joining were grouped into operational taxonomic units using a 97% similarity threshold. The sequence data were analyzed and categorized from the phylum to the family level using the default settings of the Ribosomal Database Project classifier. The results of the sequence analysis are available in the DDBJ Sequence Read Archive under project identification number PRJDB10470.
2.5. Data Analyses
The fermentation product data were subjected to two-way analysis of variance (ANOVA), with sealing time and BP addition as the main factors. The microbiota data were subjected to principal coordinate analysis (PCoA) to define the assignment and clustering accounting for the variations in the microbiota. Discriminant vectors with a Pearson correlation >0.7 were considered significant. Two-way ANOVA was performed using JMP software (version 11; SAS Institute, Tokyo, Japan), and PCoA was carried out using Primer (version 7) with the Permanova+ add-on (Primer-E; Plymouth Marine Laboratory, Plymouth, UK).
3. Results
3.1. Chemical Composition and Fermentation Products
The DM contents of raw SCR were 182 and 239 g/kg for the F1 and F2 products, respectively, indicating that the contents did not change much during piling and unprocessing for 2 days (
Table 1). A small amount of lactic acid was observed in raw SCR, while acetic acid and NH
3–N were found in DS pre-ensiled SCR.
When F1 SCR was ensiled, lactic acid fermentation was observed regardless of PS and DS (
Table 2). The lactic acid levels were 37.5 and 55.5 g/kg DM at 2 weeks in PS and DS silage, respectively. After storage for 6 months, the lactic acid content increased by about 40% compared to that at 2 weeks. BP addition suppressed lactic acid production, and this effect was manifested in DS silage. The acetic acid content was 6–20% of the lactic acid content, with levels higher than PS silage. The addition of BP increased the acetic acid content in PS silage stored for 3 and 6 months. The ethanol content was lower than the acetic acid content and greater in DS than in PS silage. Although the NH
3–N content was as low as 0.13 g/kg DM even in long-stored PS silage, the level was substantially high (as much as 2.0 g/kg DM) in DS silage. The addition of BP lowered the NH
3–N level in DS silage but not in PS silage.
When F2 SCR was ensiled, lactic acid fermentation was predominant, and DS enhanced the lactic and acetic acid contents (
Table 3). Similar to F1 SCR silages, BP addition suppressed the lactic acid content in DS silage and increased the acetic acid content in PS silage. Likewise, the NH
3–N content attained at 2.0 g/kg DM in DS silage and BP addition only lowered the NH
3–N levels in PS silage. Unlike F1 SCR ensiling, however, substantial amounts of propionic and butyric acid were produced, and the levels of 20 g/kg DM at 6 months were greater than those of acetic acid in PS silage.
3.2. Bacterial Microbiota
The MiSeq sequencing resulted in non-chimeric sequence reads with an average of 68,820 and 70,626 for F1 and F2 samples, respectively.
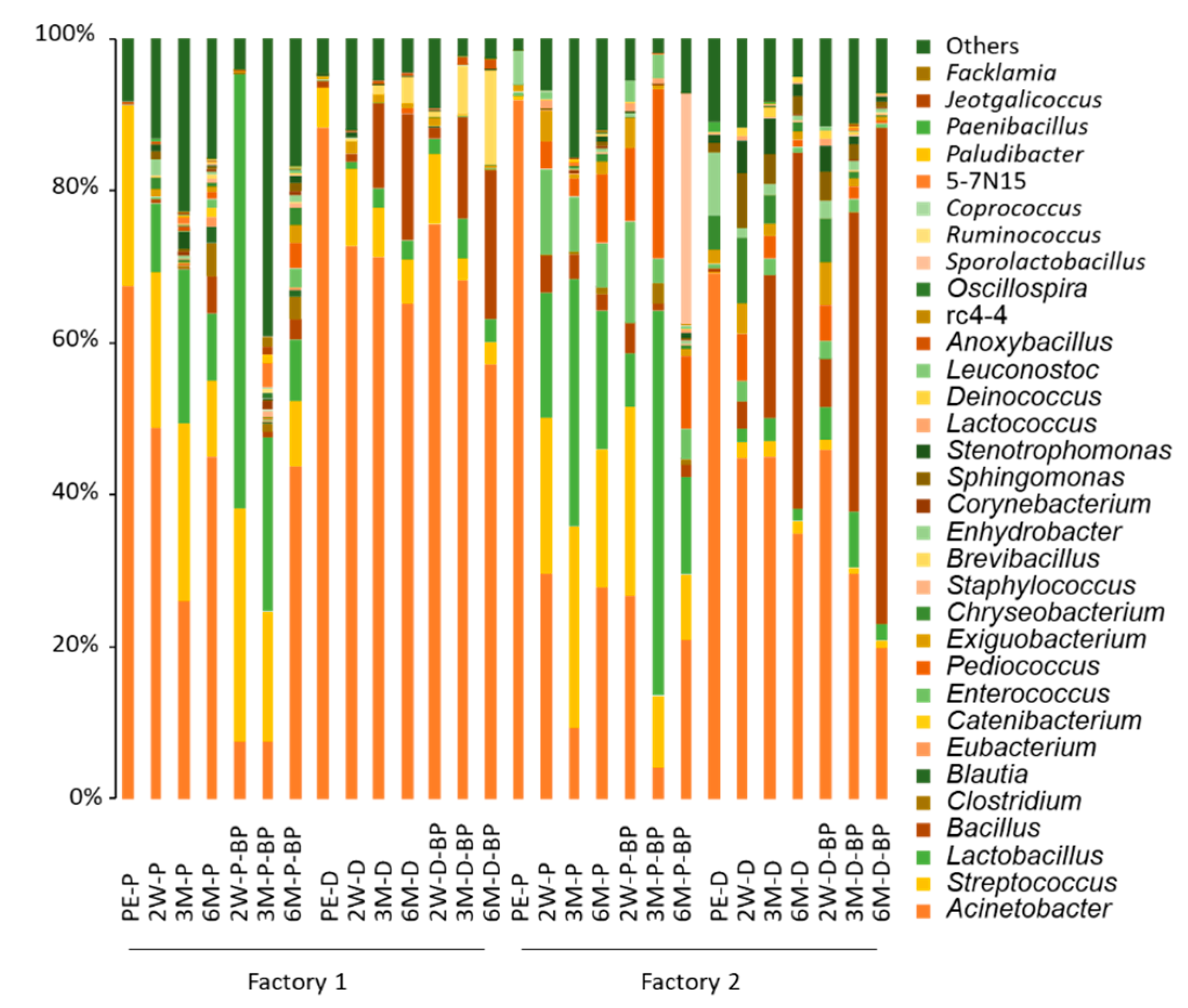
Acinetobacter spp. were the most abundant bacteria (67.4%) in pre-ensiled F1 SCR (
Figure 1). In F1 PS silages, the abundance of
Acinetobacter spp. was 7.6–26.1% at 3 months, growing to >40% at 6 months. Although
Lactobacillus spp. were undetectable in pre-ensiled F1 SCR, the abundance increased to 8.99% and >30% at 2 weeks and 3 months, respectively, in F1 PS silage.
Streptococcus spp. were found at >20% in pre-ensiled F1 SCR. The abundance was retained until 3 months before decreasing to <10% at 6 months. After 6 months of storage,
Bacillus (2.7–4.9%),
Enterococcus (1.2–2.5%), and
Clostridium spp. (3.0–4.4%) in F1 PS silage became non-negligible.
The abundance of Acinetobacter spp. in SCR increased to 88% during piling and unprocessing for 2 days. Although the abundance decreased during prolonged ensiling, levels of 57.2–65.2% were observed in F1 DS silage even after 6 months. BP addition did not affect the abundance of Acinetobacter spp. The abundance of Lactobacillus spp. was maintained at <6% throughout the 6 months in F1 DS silage, regardless of BP addition. Likewise, Streptococcus spp. were found at 5.3% after ensiling, however, the abundance was almost unchanged throughout ensiling. Meanwhile, the abundance of Bacillus spp. increased to 16.7–19.7% at 6 months from <1% at the time of ensiling in F1 DS silage.
Acinetobacter spp. were by far the most abundant bacteria (91.9%) in pre-ensiled F2 SCR, followed by Enhydrobacter spp. (4.47%). In F2 PS silage, the abundance of Acinetobacter spp. was <10% at 3 months, and increased to >20% at 6 months, regardless of BP addition. Similar to pre-ensiled F1 SCR, although Lactobacillus spp. were initially undetectable at ensiling in F2 PS silage, the abundance increased to 7.19–16.6% at 2 weeks and 32.5–50.7% at 3 months. Streptococcus spp. were observed at <0.5% at ensiling, >20% at 2 weeks, and 8.5–18.0% at 6 months in F2 PS silage. As a result of long-term storage, Bacillus (1.7–2.1%) and Enterococcus spp. (4.0–5.6%) were also observed in F2 PS silage. Unlike the F1 SCR silages, Clostridium spp. were found at around <1.0% in F2 PS silage stored for 6 months.
Keeping SCR piled and unprocessed for 2 days decreased the abundance of Acinetobacter spp. (69.2%) in pre-ensiled F2 SCR. Although Acinetobacter spp. remained the most abundant taxon at 2 weeks, Bacillus spp. became apparent after prolonged ensiling and were the most abundant bacteria (46.8–65.3%) at 6 months in F2 DS silage. The abundance of Lactobacillus spp. in F2 DS silage remained low (<10%), regardless of BP addition.
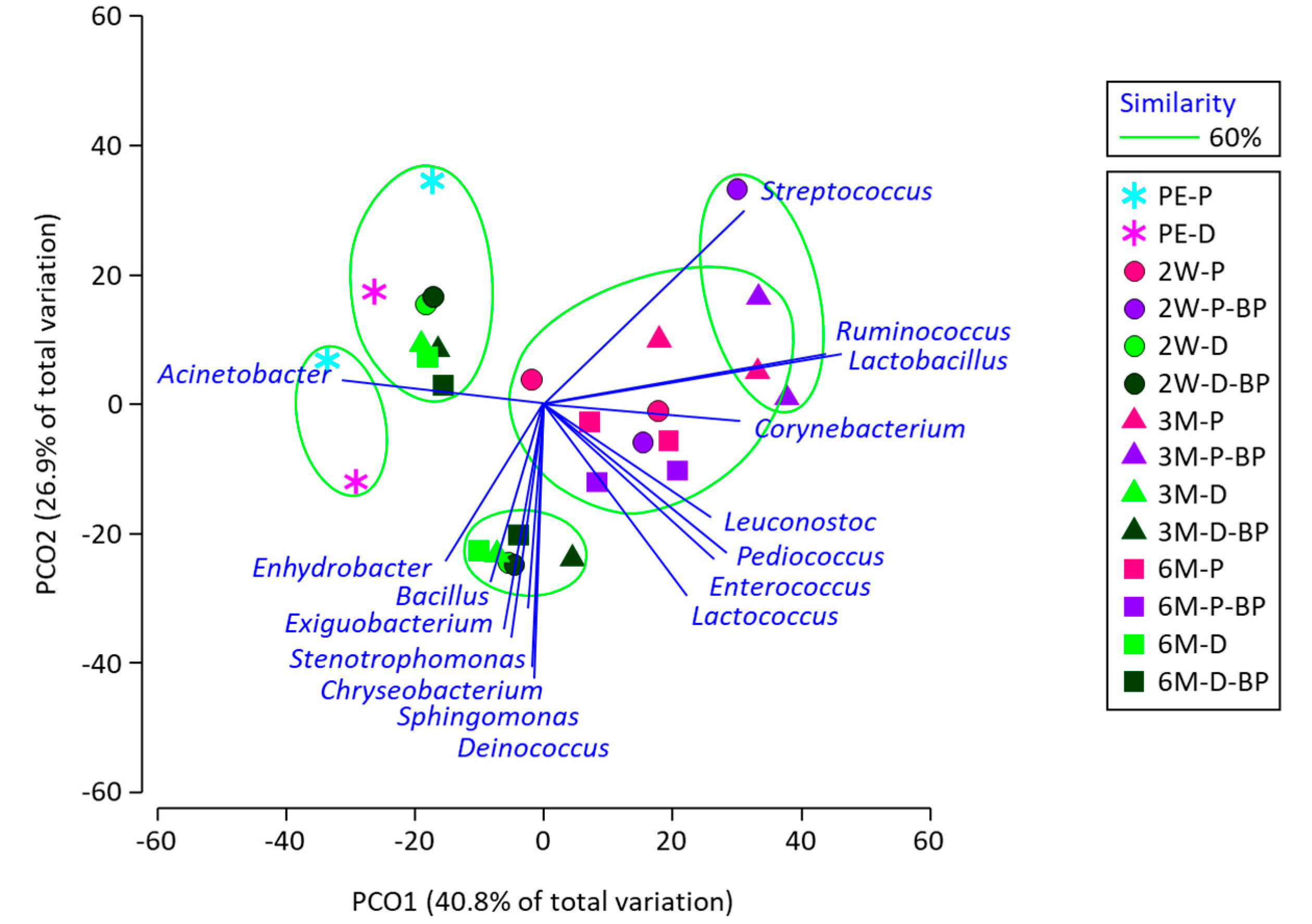
The PCoA results demonstrated that the microbiota of SCR silage was clearly separated by sealing time (
Figure 2). Although the difference between F1 and F2 was not observed for PS silage, the difference was clear for DS silage. The effect of BP addition on silage microbiota was not distinctive in either the PS or DS silages.
3.3. Fungal Microbiota
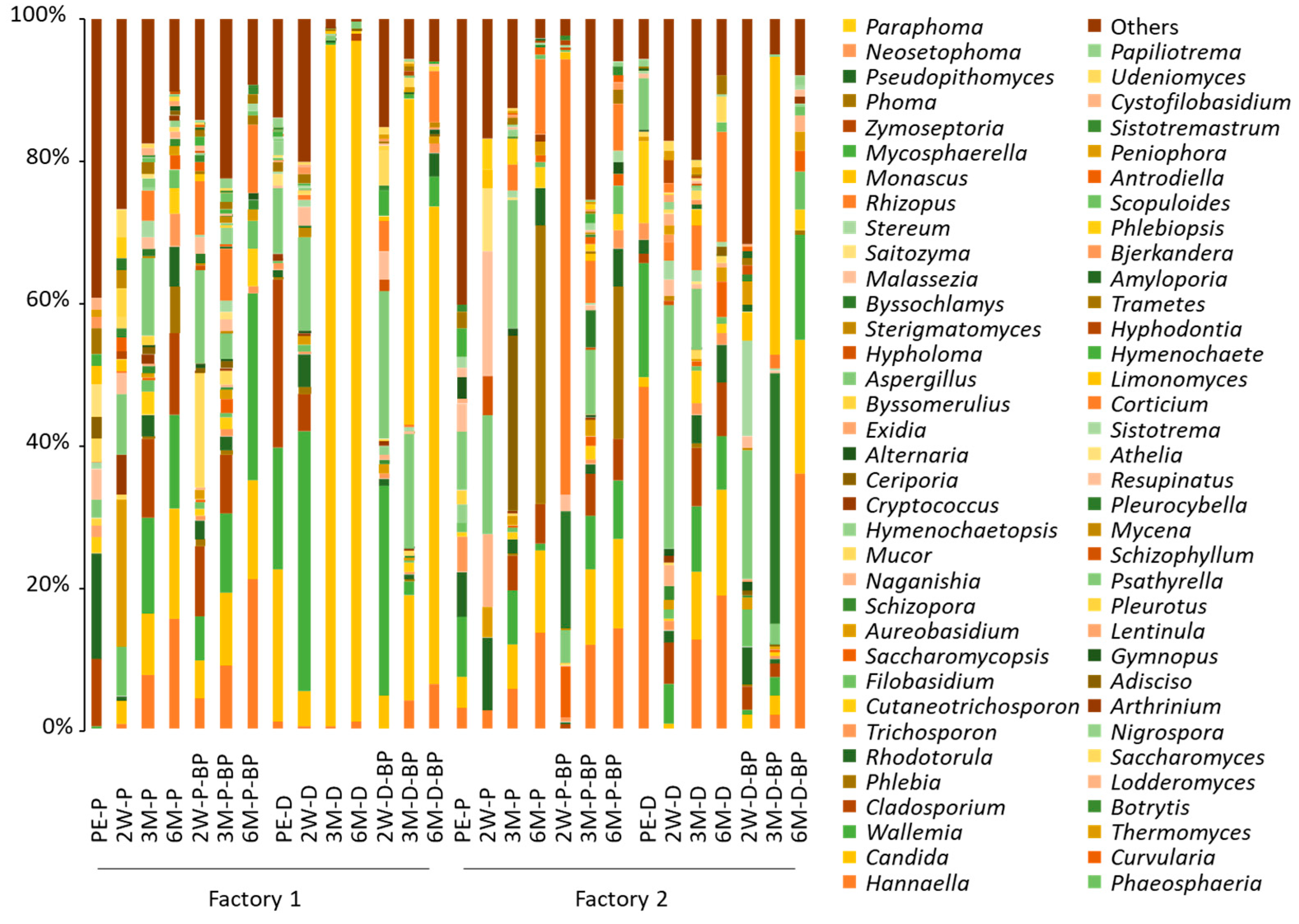
The number of non-chimeric sequence reads was low for the analysis of fungal microbiota, with an average of 7649 and 5854 for F1 and F2 samples, respectively. However, diverse fungal species with relative abundances of <10%, except for
Rhodotorula spp. (14.8%), were found in pre-ensiled F1 SCR (
Figure 3). In F1 PS silage,
Aspergillus,
Candida,
Cladosporium,
Aureobasidium,
Hannaella,
Wallemia, and
Mucor spp. were detected at >10% at 2 weeks. The major fungi detected at 3 months were
Aspergillus (3.6–11.0%),
Cladosporium (8.3–11.1%), and
Wallemia spp. (11.1–13.6%) at 3 months, and
Candida (14.0–15.4%),
Hannaella (15.6–21.2%), and
Wallemia spp. (13.4–26.3%) at 6 months in F1 PS silage.
After SCR was piled and unprocessed for 2 days, Candida (21.7%), Cladosporium (23.6%), and Wallemia spp. (17.2%) were found to be major fungi, while the abundance of Rhodotorula spp. was substantially low (1.1%) in pre-ensiled F1 SCR. Wallemia (17.2–29.7%) and Aspergillus spp. (13.1–20.9%) were found in F1 DS silage at 2 weeks, while Candida spp. became predominant (>95%) at 3 months. The abundance of Candida spp. (67%) was also high at 6 months in F2 DS silage stored with BP.
Fungal microbiota were also highly diverse in pre-ensiled F2 SCR. Although Aspergillus, Rhodotorula, and Wallemia spp. were the major species, their abundance was only <10%. Candida (6.1–12.8%) and Hannaella spp. (5.9–14.2%) became apparent at 3 and 6 months in F2 PS silage, respectively. Phlebia spp. was detected at <0.2% at 3 months, but its abundance increased abruptly (21.4–39.4%) in F2 PS silage at 6 months.
Hannaella (48.1%), Wallemia (16.3%), and Cutaneotrichosporon spp. (11.5%) were the major fungi in pre-ensiled F2 SCR after being piled and unprocessed for 2 days. The abundances of Hannaella and Wallemia spp. decreased at 2 weeks and 3 months but returned to high levels (18.8–36.2%) at 6 months in F2 DS silage. The abundance of Cutaneotrichosporon spp. declined to <3% at 6 months. Although Aspergillus spp. were found at high levels (18.3–34.0%) at 2 weeks, the species was not observed in F2 DS silage at 6 months. The abundance of Candida spp. gradually increased during ensiling, culminating in an abundance of 15.0–18.7% at 6 months in F2 DS silage. Byssochlamys (35.2%) and Monascus spp. (42.0%) were the major fungi in F2 DS silage at 3 months. However, these fungi became undetectable at 6 months in F2 DS silage stored with BP.
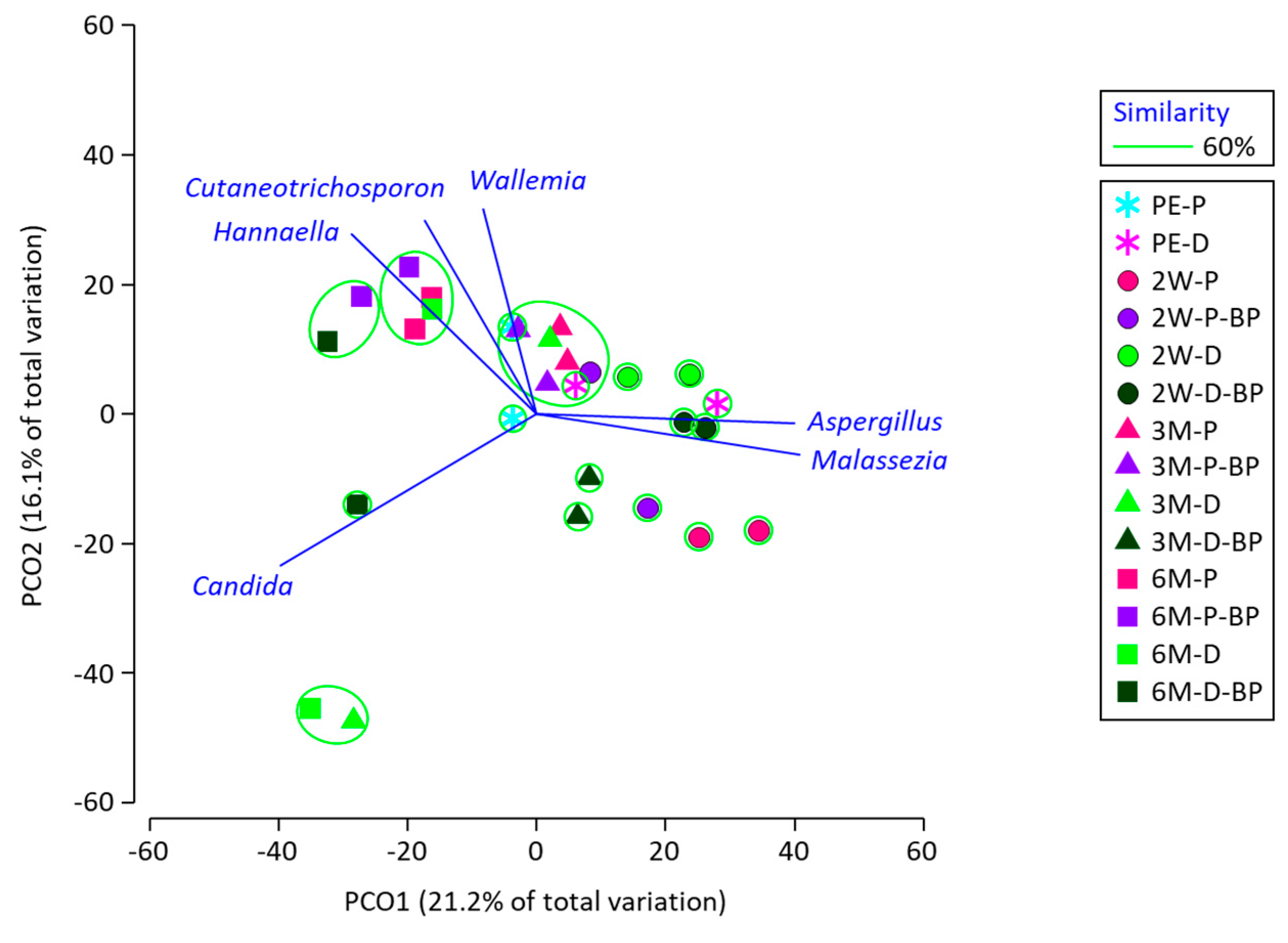
The PCoA results illustrate the diversity of fungal microbiota in SCR (
Figure 4). All data for silages at 2 weeks were grouped separately. Silages at 6 months were characterized by
Cutaneotrichosporon and
Hannaella spp., but no fungi showed differences between F1 and F2 and the effect of BP addition.
4. Discussion
Tofu is a traditional Asian food, and many studies have been conducted to examine the process of soybean fermentation and identify additives for SCR ensiling [
4]. Because of its milled and mashed physical properties, SCR is regarded as an easy material to ensile. As a result, the fermentation of this legume can be performed by lactic acid without the need for additives. The microbiota associated with SCR ensiling have also been examined in a number of studies [
10]. However, except for the total mixed ration silage, containing SCR as an ingredient, most of the previous findings have been derived from plate culture, which enumerated limited taxa [
11,
12]. The rancidity arising from piling and unprocessing is an important obstacle to the widespread use of SCR. Although this study examined two types of wet SCRs derived from two different factories (F1 and F2), the microbiota associated with SCR ensiling were found to be different from those associated with forage ensiling.
After separation from soy milk, wet SCR (pre-ensiled PS material) had small amounts (16.7–22.1 g/kg DM in total) of soluble sugars, with sucrose and stachyose being the major sugars. In pre-ensiled DS materials, the sucrose content decreased, stachyose disappeared, and maltose appeared, indicating that the microbiota of wet SCR exerted starch hydrolyzing activity before ensiling.
Acinetobacter spp. were the most abundant bacteria in pre-ensiled SCR. The fact that
Acinetobacter spp. possess adequate levels of amylase activity [
13] supports this. Of note, stachyose and raffinose are soy oligosaccharides that can promote the growth of lactic acid bacteria [
14]; hence, the difference in the soy oligosaccharides content between PS and DS pre-ensiled SCR could account for a higher abundance of
Lactobacillus spp. in PS than that in DS silages.
A diverse range of genera, including
Agrobacterium,
Erwinia,
Methylobacterium,
Microbacterium,
Pedobacter,
Pseudomonas, and
Sphingomonas spp., are the most abundant species in forage ensiling [
15].
Acinetobacter spp. (
Moraxellaceae) are regarded as minor bacterial species in pre-ensiled forages. However, in a previous study,
Acinetobacter spp. was detected at a high abundance in pre-ensiled napier grass and sudan grass [
16].
Acinetobacter spp. are aerobic, non-fermentative, and ubiquitous in nature [
17]; hence, because of the preceding manufacturing processes, including heating, pre-ensiled SCR was likely to be vulnerable to contamination by factory-associated microbiota. In F2 SCR, the levels of
Enhydrobacter spp. numerically increased during piling and unprocessing for 2 days. However, starch hydrolysis activity has not yet been observed in
Enhydrobacter spp.
Our finding that lactic acid dominated the fermentation of SCR silage with PS is in agreement with other studies [
4,
10]. Although the soluble sugar content was low, the lactic acid content reached >30 g/kg DM after 2 weeks. Although the initial abundances of lactic acid-producing bacteria were quite small,
Lactobacillus and
Streptococcus spp. were the main bacteria in F1 PS silage, while
Lactobacillus,
Streptococcus,
Enterococcus, and
Pediococcus spp. were the main bacteria in F2 PS silage. Based on the PCoA results, the bacterial microbiota of PS silage was found to set up and remain separated from that of DS silage. Interestingly, although
Acinetobacter and
Bacillus spp. remained the most abundant bacteria during ensiling, lactic acid was prevalent in DS silage. Since
Acinetobacter spp. lack the ability to produce lactic acid,
Bacillus spp. could have been involved in lactic acid fermentation in DS silage. Although
Bacillus spp. are generally regarded as strictly aerobic bacteria, facultative anaerobic species are also present, and their ability to produce lactic acid has been acknowledged [
18,
19,
20].
Bacillus spp. are known to grow well in soybean products, exhibiting various enzyme activities, including protease, amylase, cellulase, and pectinase [
21,
22]. In the present study, when SCR was unprocessed and unsealed for 2 days, the abundance of
Bacillus spp. increased from 0.03–0.05% to 0.49–0.90%, while maltose became detectable, and a substantial amount of NH
3–N (0.64–1.37 g/kg DM) was produced.
Paenibacillus,
Exiguobacterium,
Enhydrobacter,
Stenotrophomonas, and
Deinococcus spp. also increased their abundance during piling and unprocessing for 2 days. Further,
Acinetobacter spp., the most abundant bacteria in pre-ensiled SCR, were also shown to exert both proteinase and deaminase activities [
23]. Hence, the intensive production of NH
3–N observed in the DS materials could be attributed to the collective activities of bacteria, including
Bacillus and
Acinetobacter spp. The NH
3–N content of PS silage without added BP was as low as 0.31 g/kg DM, indicating that the proteolysis and deamination activities were efficiently suppressed under PS conditions.
Although the cow gut microbiota has been demonstrated to be robust and not easily affected by silage-derived microbiota, several lactic acid bacteria species were shown to inhabit both silage and feces [
24]. Moreover,
Bacillus spp., isolated from fermented soybean, were proven to improve milk production and promote the growth of total, proteolytic, and amylolytic bacteria in the rumen [
25]. Therefore, the potential of SCR silage as a vehicle for probiotics delivery deserves to be explored.
The fungal microbiota in SCR was found to be diverse, with no distinctive changes after the fermentation of the PS and DS silages. These results suggest that bacterial microbiota play a key role in the fermentation of SCR silage. Although
Candida,
Aspergillus,
Cladosporium,
Hannaella, and
Wallemia spp. were the major fungal species in the PS and DS silages, information on
Cladosporium,
Hannaella, and
Wallemia spp. is scarce in relation to their use in feed preservation. Although Pascal et al. [
26] found
Hannaella spp. in whole crop corn silage at a relative abundance of around 20%, their roles in anaerobic storage and spoilage are not known.
Wallemia and
Cladosporium spp. are considered spoilage fungi [
27,
28]; however, no studies have been conducted on the detection and isolation of these species from silage. Furthermore, microbiota assessment has so far been performed mainly on bacteria in silage research. Thus, further studies will be needed to improve our understanding of microbiota control.








{kind=link}
{kind=link}
{kind=link}
{kind=link}