
Effects of Radiation Intensity, Mineral Matrix, and Pre-Irradiation on the Bacterial Resistance to Gamma Irradiation under Low Temperature Conditions

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
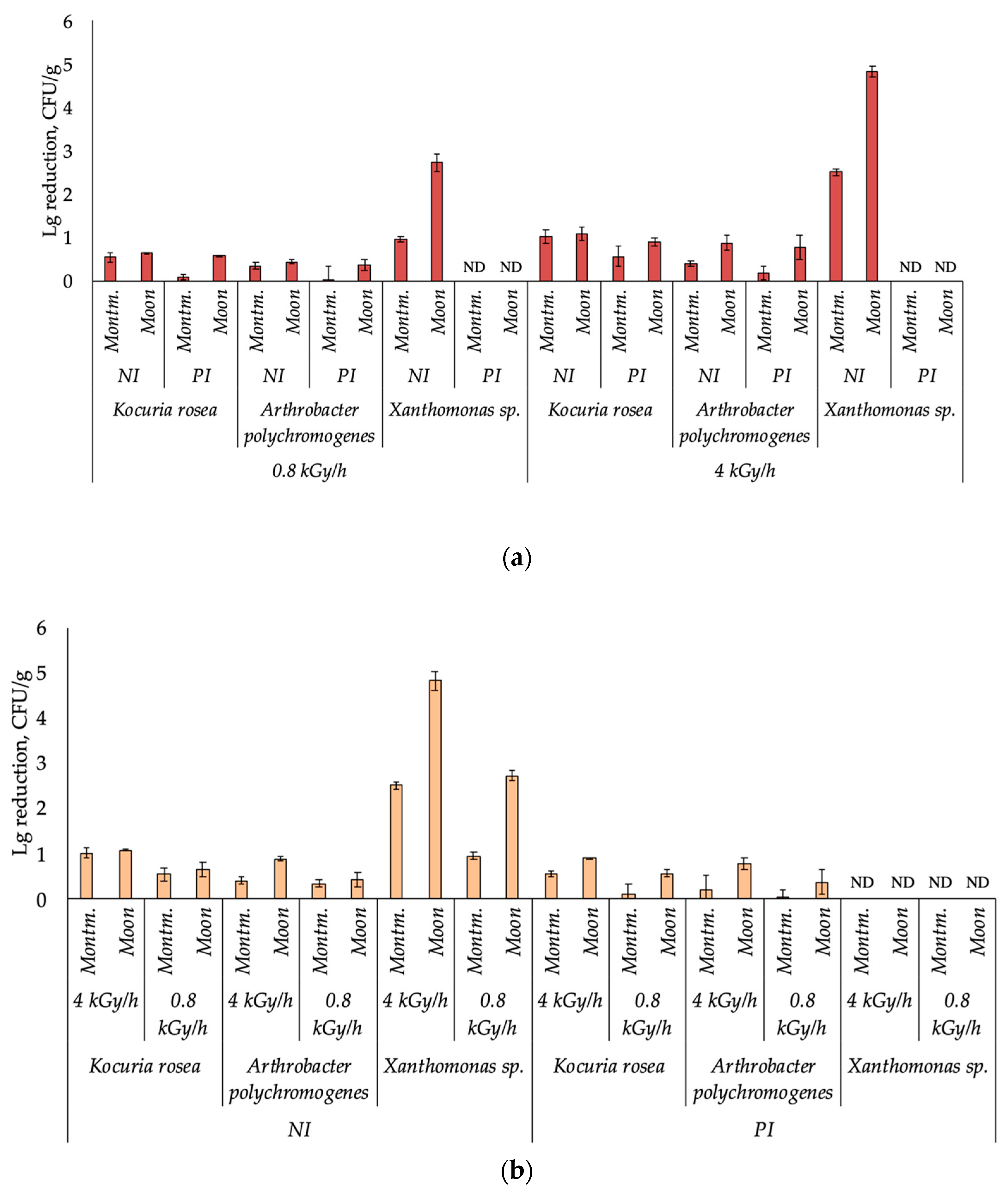
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Strain | Irradiation Intensity, kGy/h | Mineral Matrix | Pre-Irradiation | Number of CFU Relative to Control |
|---|---|---|---|---|
| Kocuria rosea SN_T60 | 4 | Montmorillonite | Non pre-irradiated | 9.40 ± 0.36% |
| 4 | Lunar dust analog | Non pre-irradiated | 8.09 ± 0.24% | |
| 4 | Montmorillonite | Pre-irradiated | 27.01 ± 1.03% | |
| 4 | Lunar dust analog | Pre-irradiated | 12.54 ± 0.67% | |
| 0.8 | Montmorillonite | Non pre-irradiated | 28.08 ± 1.4% | |
| 0.8 | Lunar dust analog | Non pre-irradiated | 21.92 ± 1.44% | |
| 0.8 | Montmorillonite | Pre-irradiated | 80.12 ± 5.68% | |
| 0.8 | Lunar dust analog | Pre-irradiated | 26.50 ± 1.62% | |
| Arthrobacter polychromogenes SN_T61 | 4 | Montmorillonite | Non pre-irradiated | 39.14 ± 1.48% |
| 4 | Lunar dust analog | Non pre-irradiated | 12.96 ± 0.31% | |
| 4 | Montmorillonite | Pre-irradiated | 63.93 ± 6.2% | |
| 4 | Lunar dust analog | Pre-irradiated | 16.63 ± 1.35% | |
| 0.8 | Montmorillonite | Non pre-irradiated | 44.76 ± 1.54% | |
| 0.8 | Lunar dust analog | Non pre-irradiated | 36.50 ± 1.83% | |
| 0.8 | Montmorillonite | Pre-irradiated | 92.62 ± 7.37% | |
| 0.8 | Lunar dust analog | Pre-irradiated | 42.40 ± 4.2% | |
| Xanthomonas sp. DP3 | 4 | Montmorillonite | Non pre-irradiated | 0.31 ± 0.02% |
| 4 | Lunar dust analog | Non pre-irradiated | 0.002 ± 0.0001% | |
| 0.8 | Montmorillonite | Non pre-irradiated | 10.99 ± 0.87% | |
| 0.8 | Lunar dust analog | Non pre-irradiated | 0.19 ± 0.01% |
References
- Gargaud, M.; Amils, R.; Cleaves, H.J. (Eds.) Encyclopedia of Astrobiology; Springer Science & Business Media: Berlin, Germany, 2011; Volume 1, ISBN 978-3-662-44184-8. [Google Scholar]
- Cockell, C.S. Astrobiology: Understanding Life in the Universe; John Wiley & Sons: Hoboken, NJ, USA, 2020; ISBN 978-1-119-55035-8. [Google Scholar]
- Cottin, H.; Kotler, J.M.; Bartik, K.; Cleaves, H.J.; Cockell, C.S.; De Vera, P.; Ehrenfreund, P.; Leuko, S.; Kate, T.I.L.; Martins, Z.; et al. Astrobiology and the possibility of life on earth and elsewhere…. Space Sci. Rev. 2017, 209, 1–42. [Google Scholar] [CrossRef] [Green Version]
- Röling, W.F.; Aerts, J.W.; Patty, C.L.; Ten Kate, I.L.; Ehrenfreund, P.; Direito, S.O. The significance of microbe-mineral-biomarker interactions in the detection of life on Mars and beyond. Astrobiology 2015, 15, 492–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze-Makuch, D.; Dohm, J.M.; Fairén, A.G.; Baker, V.R.; Fink, W.; Strom, R.G. Venus, Mars, and the ices on Mercury and the Moon: Astrobiological implications and proposed mission designs. Astrobiology 2005, 5, 778–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rummel, J.D.; Beaty, D.W.; Jones, M.A.; Bakermans, C.; Barlow, N.G.; Boston, P.J.; Chevrier, V.F.; Clark, B.C.; de Vera, J.P.; Gough, R.V.; et al. A new analysis of Mars “special regions”: Findings of the second MEPAG Special Regions Science Analysis Group (SR-SAG2). Astrobiology 2014, 14, 887–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, D.; Margonelli, A.; Pace, E.; Giardi, M.T.; Faraloni, C.; Torzillo, G.; Zanini, A. The effect of ionising radiation on photosynthetic oxygenic microorganisms for survival in space flight revealed by automatic photosystem II-based biosensors. Microgravity Sci. Technol. 2006, 18, 215. [Google Scholar] [CrossRef]
- Moissl-Eichinger, C.; Cockell, C.; Rettberg, P. Venturing into new realms? Microorganisms in space. FEMS Microbiol. Rev. 2016, 40, 722–737. [Google Scholar] [CrossRef]
- Preston, L.J.; Dartnell, L.R. Planetary habitability: Lessons learned from terrestrial analogues. Int. J. Astrobiol. 2014, 13, 81–98. [Google Scholar] [CrossRef] [Green Version]
- Szocik, K.; Wójtowicz, T.; Rappaport, M.B.; Corbally, C. Ethical issues of human enhancements for space missions to Mars and beyond. Futures 2020, 115, 102489. [Google Scholar] [CrossRef]
- Voorhies, A.A.; Ott, C.M.; Mehta, S.; Pierson, D.L.; Crucian, B.E.; Feiveson, A.; Oubre, C.M.; Torralba, M.; Moncera, K.; Zhang, Y.; et al. Study of the impact of long-duration space missions at the International Space Station on the astronaut microbiome. Sci. Rep. 2019, 9, 1–17. [Google Scholar] [CrossRef]
- Levchenko, I.; Xu, S.; Mazouffre, S.; Keidar, M.; Bazaka, K. Mars colonization: Beyond getting there. Glob. Chall. 2019, 3, 1800062. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.V.; Peixoto, R.S.; Rosado, A.S. Inevitable future: Space colonization beyond Earth with microbes first. FEMS Microbiol. Ecol. 2019, 95, fiz127. [Google Scholar] [CrossRef]
- Dartnell, L.R.; Hunter, S.J.; Lovell, K.V.; Coates, A.J.; Ward, J.M. Low-temperature ionizing radiation resistance of Deinococcus radiodurans and Antarctic Dry Valley bacteria. Astrobiology 2010, 10, 717–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheptsov, V.S.; Vorobyova, E.A.; Manucharova, N.A.; Gorlenko, M.V.; Pavlov, A.K.; Vdovina, M.A.; Lomasov, V.N.; Bulat, S.A. 100 kGy gamma-affected microbial communities within the ancient Arctic permafrost under simulated Martian conditions. Extremophiles 2017, 21, 1057–1067. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cheptsov, V.; Vorobyova, E.; Belov, A.; Pavlov, A.; Tsurkov, D.; Lomasov, V.; Bulat, S. Survivability of soil and permafrost microbial communities after irradiation with accelerated electrons under simulated Martian and open space conditions. Geosciences 2018, 8, 298. [Google Scholar] [CrossRef] [Green Version]
- Shuryak, I.; Matrosova, V.Y.; Gaidamakova, E.K.; Tkavc, R.; Grichenko, O.; Klimenkova, P.; Volpe, R.P.; Daly, M.J. Microbial cells can cooperate to resist high-level chronic ionizing radiation. PLoS ONE 2017, 12, e0189261. [Google Scholar] [CrossRef] [PubMed]
- Deo, N.; Natarajan, K.A.; Somasundaran, P. Mechanisms of adhesion of Paenibacillus polymyxa onto hematite, corundum and quartz. Int. J. Miner. Process. 2001, 62, 27–39. [Google Scholar] [CrossRef]
- Jiang, D.; Huang, Q.; Cai, P.; Rong, X.; Chen, W. Adsorption of Pseudomonas putida on clay minerals and iron oxide. Colloid Surf. B 2007, 54, 217–221. [Google Scholar] [CrossRef]
- Fletcher, M. The physiological activity of bacteria attached to solid surfaces. In Advances in Microbial Physiology; Academic Press: Cambridge, MA, USA, 1991; pp. 53–85. [Google Scholar] [CrossRef]
- Fletcher, M. Comparative physiology of attached and free-living bacteria. In Microbial Adhesion and Aggregation; Springer: Berlin/Heidelberg, Germany, 1984; pp. 223–232. [Google Scholar] [CrossRef]
- Zakharov, Y.A.; Nevostruev, V.A. Radiolysis of solid inorganic salts with oxygen-containing anions. Russ. Chem. Rev. 1968, 37, 61. Available online: https://0-iopscience-iop-org.brum.beds.ac.uk/article/10.1070/RC1968v037n01ABEH001608/pdf (accessed on 15 November 2020). [CrossRef]
- Wang, Z.; Walter, E.D.; Sassi, M.; Zhang, X.; Zhang, H.; Li, X.S.; Chen, Y.; Cui, W.; Tuladhar, A.; Chase, Z.; et al. The role of surface hydroxyls on the radiolysis of gibbsite and boehmite nanoplatelets. J. Hazard. Mater. 2020, 122853. [Google Scholar] [CrossRef]
- Gournis, D.; Mantaka-Marketou, A.E.; Karakassides, M.A.; Petridis, D. Ionizing radiation-induced defects in smectite clays. Phys. Chem. Miner. 2001, 28, 285–290. [Google Scholar] [CrossRef]
- Brown, A.R.; Wincott, P.L.; LaVerne, J.A.; Small, J.S.; Vaughan, D.J.; Pimblott, S.M.; Lloyd, J.R. The impact of γ radiation on the bioavailability of Fe (III) minerals for microbial respiration. Environ. Sci. Technol. 2014, 48, 10672–10680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaddissy, J.A.; Esnouf, S.; Durand, D.; Saffre, D.; Foy, E.; Renault, J.P. Radiolytic events in nanostructured aluminum hydroxides. J. Phys. Chem. C 2017, 121, 6365–6373. [Google Scholar] [CrossRef]
- Kaučić, S.; Maddock, A.G. Effect of cation vacancies on radiolysis of sodium nitrate. J. Chem. Soc. Faraday Trans. 1969, 65, 1083–1090. [Google Scholar] [CrossRef]
- Onofri, S.; Selbmann, L.; Pacelli, C.; Zucconi, L.; Rabbow, E.; de Vera, J.P. Survival, DNA, and ultrastructural integrity of a cryptoendolithic Antarctic fungus in Mars and Lunar rock analogs exposed outside the International Space Station. Astrobiology 2019, 19, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Aureli, L.; Pacelli, C.; Cassar, A.; Fujimori, A.; Moeller, R.; Onofri, S. Iron ion particle radiation resistance of dried colonies of Cryomyces antarcticus embedded in Martian regolith analogues. Life 2020, 10, 306. [Google Scholar] [CrossRef]
- Pacelli, C.; Cassaro, A.; Aureli, L.; Moeller, R.; Fujimori, A.; Onofri, S. The responses of the black fungus Cryomyces antarcticus to high doses of accelerated helium ions radiation within martian regolith simulants and their relevance for Mars. Life 2020, 10, 130. [Google Scholar] [CrossRef]
- Paulino-Lima, I.G.; Janot-Pacheco, E.; Galante, D.; Cockell, C.; Olsson-Francis, K.; Brucato, J.R.; Baratta, G.A.; Strazzulla, G.; Merrigan, T.; McCullough, R.; et al. Survival of Deinococcus radiodurans against laboratory-simulated solar wind charged particles. Astrobiology 2011, 11, 875–882. [Google Scholar] [CrossRef]
- Bauermeister, A.; Moeller, R.; Reitz, G.; Sommer, S.; Rettberg, P. Effect of relative humidity on Deinococcus radiodurans’ resistance to prolonged desiccation, heat, ionizing, germicidal, and environmentally relevant UV radiation. Microb. Ecol. 2011, 61, 715–722. [Google Scholar] [CrossRef]
- Cheptsov, V.S.; Vorobyova, E.A.; Osipov, G.A.; Manucharova, N.A.; Polyanskaya, L.M.; Gorlenko, M.V.; Pavlov, A.K.; Rosanova, M.S.; Lomasov, V.N. Microbial activity in Martian analog soils after ionizing radiation: Implications for the preservation of subsurface life on Mars. AIMS Microbiol. 2018, 4, 541. [Google Scholar] [CrossRef]
- Minton, K.W. DNA repair in the extremely radioresistant bacterium Deinococcus radiodurans. Mol. Microbiol. 1994, 13, 9–15. [Google Scholar] [CrossRef]
- Cox, M.M.; Battista, J.R. Deinococcus radiodurans—The consummate survivor. Nat. Rev. Microbiol. 2005, 3, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Musilova, M.; Wright, G.; Ward, J.M.; Dartnell, L.R. Isolation of radiation-resistant bacteria from Mars analog Antarctic Dry Valleys by preselection, and the correlation between radiation and desiccation resistance. Astrobiology 2015, 15, 1076–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kottemann, M.; Kish, A.; Iloanusi, C.; Bjork, S.; DiRuggiero, J. Physiological responses of the halophilic archaeon Halobacterium sp. strain NRC1 to desiccation and gamma irradiation. Extremophiles 2005, 9, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Homeck, G.; Mileikowsky, C.; Melosh, H.J.; Wilson, J.W.; Cucinotta, F.A.; Gladman, B. Viable transfer of microorganisms in the solar system and beyond. In Astrobiology; Springer: Berlin/Heidelberg, Germany, 2002; pp. 57–76. [Google Scholar] [CrossRef]
- Westall, F.; Loizeau, D.; Foucher, F.; Bost, N.; Betrand, M.; Vago, J.; Kminek, G. Habitability on Mars from a microbial point of view. Astrobiology 2013, 13, 887–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, D.R.; Pollock, S.V.; Wood, E.A.; Goiffon, R.J.; Klingele, A.J.; Cabot, E.L.; Schackwitz, W.; Martin, J.; Eggington, J.; Durfee, T.J.; et al. Directed evolution of ionizing radiation resistance in Escherichia coli. J. Bacteriol. 2009, 191, 5240–5252. [Google Scholar] [CrossRef] [Green Version]
- Tesfai, A.T.; Beamer, S.K.; Matak, K.E.; Jaczynski, J. Radioresistance development of DNA repair deficient Escherichia coli DH5α in ground beef subjected to electron beam at sub-lethal doses. Int. J. Radiat. Biol. 2011, 87, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, A.K.; Kalinin, V.L.; Konstantinov, A.N.; Shelegedin, V.N.; Pavlov, A.A. Was Earth ever infected by martian biota? Clues from radioresistant bacteria. Astrobiology 2006, 6, 911–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheptsov, V.; Belov, A.; Vorobyova, E.; Osipov, G.; Bulat, S. Viability of the soddy–podzolic soil microbial community after 148–1250 kGy gamma irradiation. Planet. Space Sci. 2019, 172, 8–13. [Google Scholar] [CrossRef]
- Cheptsov, V.S.; Vorobyova, E.A.; Tambiev, A.N. Influence of gamma irradiation in simulated martian conditions on catalase activity and reactivity of exometabolites of Kocuria rosea and Arthrobacter polychromogenes. In Proceedings of the Seventh Moscow Solar System Symposium (7M-S3), Moscow, Russia, 10–14 October 2016; Russian Academy of Science: Moscow, Russia, 2016; pp. 262–264. [Google Scholar]
- Cheptsov, V.S.; Vorobyova, E.A.; Gorlenko, M.V.; Manucharova, N.A.; Pavlov, A.K.; Vdovina, M.A.; Zvyagintsev, D.G. The impact of gamma radiation, low pressure and low temperature on the viability of the microbial community of serozem as an analytical model of the Martian regolith. Mod. Probl. Sci. Educ. 2015, 3, 619. Available online: http://www.science-education.ru/pdf/2015/3/515.pdf (accessed on 15 November 2020).
- Craig, P.I.; Mickol, R.L.; Archer, P.D.; Kral, T.A. Nontronite and montmorillonite as nutrient sources for life on Mars. In Proceedings of the Lunar and Planetary Science XLVIII, The Woodlands, TX, USA, 20–24 March 2017; p. 1997. Available online: https://ntrs.nasa.gov/citations/20170002223 (accessed on 15 November 2020).
- Clark, B.C., III; Arvidson, R.E.; Gellert, R.; Morris, R.V.; Ming, D.W.; Richter, L.; Ruff, S.W.; Michalski, J.R.; Farrand, W.H.; Yen, A.; et al. Evidence for montmorillonite or its compositional equivalent in Columbia Hills, Mars. J. Geophys. Res. Planet 2007, 112. [Google Scholar] [CrossRef] [Green Version]
- Frinak, E.K.; Mashburn, C.D.; Tolbert, M.A.; Toon, O.B. Infrared characterization of water uptake by low-temperature Na-montmorillonite: Implications for Earth and Mars. J. Geophys. Res. Atmos. 2005, 110. [Google Scholar] [CrossRef]
- Bishop, J.L.; Murad, E. Characterization of minerals and biogeochemical markers on Mars: A Raman and IR spectroscopic study of montmorillonite. J. Raman. Spectrosc. 2004, 35, 480–486. [Google Scholar] [CrossRef]
- Poroykov, A.Y.; Bednyakov, S.A.; Zaharov, A.V.; Dolnikov, G.G.; Lyash, A.N.; Shashkova, I.A.; Kuznetsov, I.A. Application of the particle trajectory imaging for modelling dusty plasma levitation on the Moon. In Journal of Physics: Conference Series; IOP Publishing: Bristol, UK; p. 2037. Available online: https://0-iopscience-iop-org.brum.beds.ac.uk/article/10.1088/1742-6596/1421/1/012037/meta (accessed on 15 November 2020).
- Orger, N.C.; Toyoda, K.; Masui, H.; Cho, M. Experimental investigation on silica dust lofting due to charging within micro-cavities and surface electric field in the vacuum chamber. Adv. Space Res. 2019, 63, 3270–3288. [Google Scholar] [CrossRef]
- Pavlov, A.K.; Shelegedin, V.N.; Vdovina, M.A.; Pavlov, A.A. Growth of microorganisms in martian-like shallow subsurface conditions: Laboratory modelling. Int. J. Astrobiol. 2010, 9, 51–58. [Google Scholar] [CrossRef]
- Vorobyova, E.A.; Belov, A.A.; Cheptsov, V.S. Resistance of microorganisms of extreme xerophytic ecotopes to the effects of inactivating factors. Izv. Timiryazevskaya Agric. Acad. 2018, 4, 111–127. [Google Scholar] [CrossRef]
- Belov, A.A.; Cheptsov, V.S.; Vorobyova, E.A.; Manucharova, N.A.; Ezhelev, Z.S. Stress-tolerance and taxonomy of culturable bacterial communities isolated from a central Mojave Desert soil sample. Geosciences 2019, 9, 166. [Google Scholar] [CrossRef] [Green Version]
- Goodfellow, M.; Nouioui, I.; Sanderson, R.; Xie, F.; Bull, A.T. Rare taxa and dark microbial matter: Novel bioactive actinobacteria abound in Atacama Desert soils. Antonie Leeuwenhoek 2018, 111, 1315–1332. [Google Scholar] [CrossRef]
- Idris, H.; Goodfellow, M.; Sanderson, R.; Asenjo, J.A.; Bull, A.T. Actinobacterial rare biospheres and dark matter revealed in habitats of the Chilean Atacama Desert. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mohammadipanah, F.; Wink, J. Actinobacteria from arid and desert habitats: Diversity and biological activity. Front. Microbiol. 2016, 6, 1541. [Google Scholar] [CrossRef] [Green Version]
- Belov, A.A.; Cheptsov, V.S.; Manucharova, N.A.; Ezhelev, Z.S. Bacterial communities of Novaya Zemlya archipelago ice and permafrost. Geosciences 2020, 10, 67. [Google Scholar] [CrossRef] [Green Version]
- Wassmann, M.; Moeller, R.; Reitz, G.; Rettberg, P. Adaptation of Bacillus subtilis cells to archean-like UV climate: Relevant hints of microbial evolution to remarkably increased radiation resistance. Astrobiology 2010, 10, 605–615. [Google Scholar] [CrossRef] [Green Version]
- Romanovskaya, V.A.; Rokitko, P.V.; Malashenko, Y.R.; Krishtab, T.P.; Chernaya, N.A. Sensitivity of soil bacteria isolated from the alienated zone around the chernobyl nuclear power plant to various stress factors. Microbiology 1999, 68, 465–469. Available online: https://europepmc.org/article/med/10576090 (accessed on 15 November 2020).
- Shukla, M.; Chaturvedi, R.; Tamhane, D.; Vyas, P.; Archana, G.; Apte, S.; Bandekar, J.; Desai, A. Multiple-stress tolerance of ionizing radiation-resistant bacterial isolates obtained from various habitats: Correlation between stresses. Curr. Microbiol. 2007, 54, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Kryazhevskikh, N.A.; Demkina, E.V.; Loiko, N.G.; Baslerov, R.V.; Kolganova, T.V.; Soina, V.S.; Manucharova, N.A.; Gal’chenko, V.F.; El’-Registan, G.I. Comparison of the adaptive potential of the Arthrobacter oxydans and Acinetobacter lwoffii isolates from permafrost sedimentary rock and the analogous collection strains. Microbiology 2013, 82, 29–42. [Google Scholar] [CrossRef]
- Boston, P.J.; Ivanov, M.V.; McKay, C.P. On the possibility of chemosynthetic ecosystems in subsurface habitats on Mars. Icarus 1992, 95, 300–308. [Google Scholar] [CrossRef]
- Fairén, A.G.; Davila, A.F.; Lim, D.; Bramall, N.; Bonaccorsi, R.; Zavaleta, J.; Uceda, E.R.; Stoker, C.; Wierzchos, J.; Dohm, J.M.; et al. Astrobiology through the ages of Mars: The study of terrestrial analogues to understand the habitability of Mars. Astrobiology 2010, 10, 821–843. [Google Scholar] [CrossRef]
- Davila, A.F.; Schulze-Makuch, D. The last possible outposts for life on Mars. Astrobiology 2016, 16, 159–168. [Google Scholar] [CrossRef]
- Ramirez, R.M. A warmer and wetter solution for early Mars and the challenges with transient warming. Icarus 2017, 297, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Panikov, N.S.; Flanagan, P.W.; Oechel, W.C.; Mastepanov, M.A.; Christensen, T.R. Microbial activity in soils frozen to below −39 °C. Soil Biol. Biochem. 2006, 38, 785–794. [Google Scholar] [CrossRef]
- Schaefer, K.; Jafarov, E. A parameterization of respiration in frozen soils based on substrate availability. Biogeosci. Discuss. 2015, 12. [Google Scholar] [CrossRef]
- Rivkina, E.M.; Friedmann, E.I.; McKay, C.P.; Gilichinsky, D.A. Metabolic activity of permafrost bacteria below the freezing point. Appl. Environ. Microbiol. 2000, 66, 3230–3233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlov, A.; Cheptsov, V.; Tsurkov, D.; Lomasov, V.; Frolov, D.; Vasiliev, G. Survival of radioresistant bacteria on Europa’s surface after pulse ejection of subsurface ocean water. Geosciences 2019, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Hassler, D.M.; Zeitlin, C.; Wimmer-Schweingruber, R.F.; Ehresmann, B.; Rafkin, S.; Eigenbrode, J.L.; Brinza, D.E.; Weigle, G.; Böttcher, S.; MSL Science Team; et al. Mars’ surface radiation environment measured with the Mars Science Laboratory’s Curiosity rover. Science 2014, 343. [Google Scholar] [CrossRef] [Green Version]
- Dartnell, L.R.; Desorgher, L.; Ward, J.M.; Coates, A.J. Martian sub-surface ionising radiation: Biosignatures and geology. Biogeosci. Discuss. 2007, 4, 455–492. Available online: https://hal.archives-ouvertes.fr/hal-00297871/ (accessed on 15 November 2020). [CrossRef] [Green Version]
- Ehlmann, B.L.; Mustard, J.F.; Murchie, S.L.; Bibring, J.P.; Meunier, A.; Fraeman, A.A.; Langevin, Y. Subsurface water and clay mineral formation during the early history of Mars. Nature 2011, 479, 53–60. [Google Scholar] [CrossRef]
- Sun, V.Z.; Milliken, R.E. Ancient and recent clay formation on Mars as revealed from a global survey of hydrous minerals in crater central peaks. J. Geophys. Res. Planet. 2015, 120, 2293–2332. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheptsov, V.S.; Belov, A.A.; Vorobyova, E.A.; Pavlov, A.K.; Lomasov, V.N. Effects of Radiation Intensity, Mineral Matrix, and Pre-Irradiation on the Bacterial Resistance to Gamma Irradiation under Low Temperature Conditions. Microorganisms 2021, 9, 198. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010198
Cheptsov VS, Belov AA, Vorobyova EA, Pavlov AK, Lomasov VN. Effects of Radiation Intensity, Mineral Matrix, and Pre-Irradiation on the Bacterial Resistance to Gamma Irradiation under Low Temperature Conditions. Microorganisms. 2021; 9(1):198. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010198
Chicago/Turabian StyleCheptsov, Vladimir S., Andrey A. Belov, Elena A. Vorobyova, Anatoli K. Pavlov, and Vladimir N. Lomasov. 2021. "Effects of Radiation Intensity, Mineral Matrix, and Pre-Irradiation on the Bacterial Resistance to Gamma Irradiation under Low Temperature Conditions" Microorganisms 9, no. 1: 198. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010198