Candida albicans as an Essential “Keystone” Component within Polymicrobial Oral Biofilm Models?

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth and Standardization of Bacteria
2.2. Development of Multi-Species Biofilms
2.3. Hard Tissue (HT)—Caries Associated Biofilm
2.4. Soft Tissue (ST)—Periodontitis/Denture Stomatitis Associated Biofilm
2.5. Testing of Oral Models
2.6. Metabolic Analysis of Oral Biofilms
2.7. Assessment of Oral Biofilms Biomass
2.8. Quantitative Analysis of Biofilm Composition
2.9. Visualization of Oral Biofilms
2.10. Assays to Assess Pathogenicity of the Oral Biofilms
2.11. Treatment of Oral Biofilms
2.12. Statistical Analysis and Data Presentation
3. Results
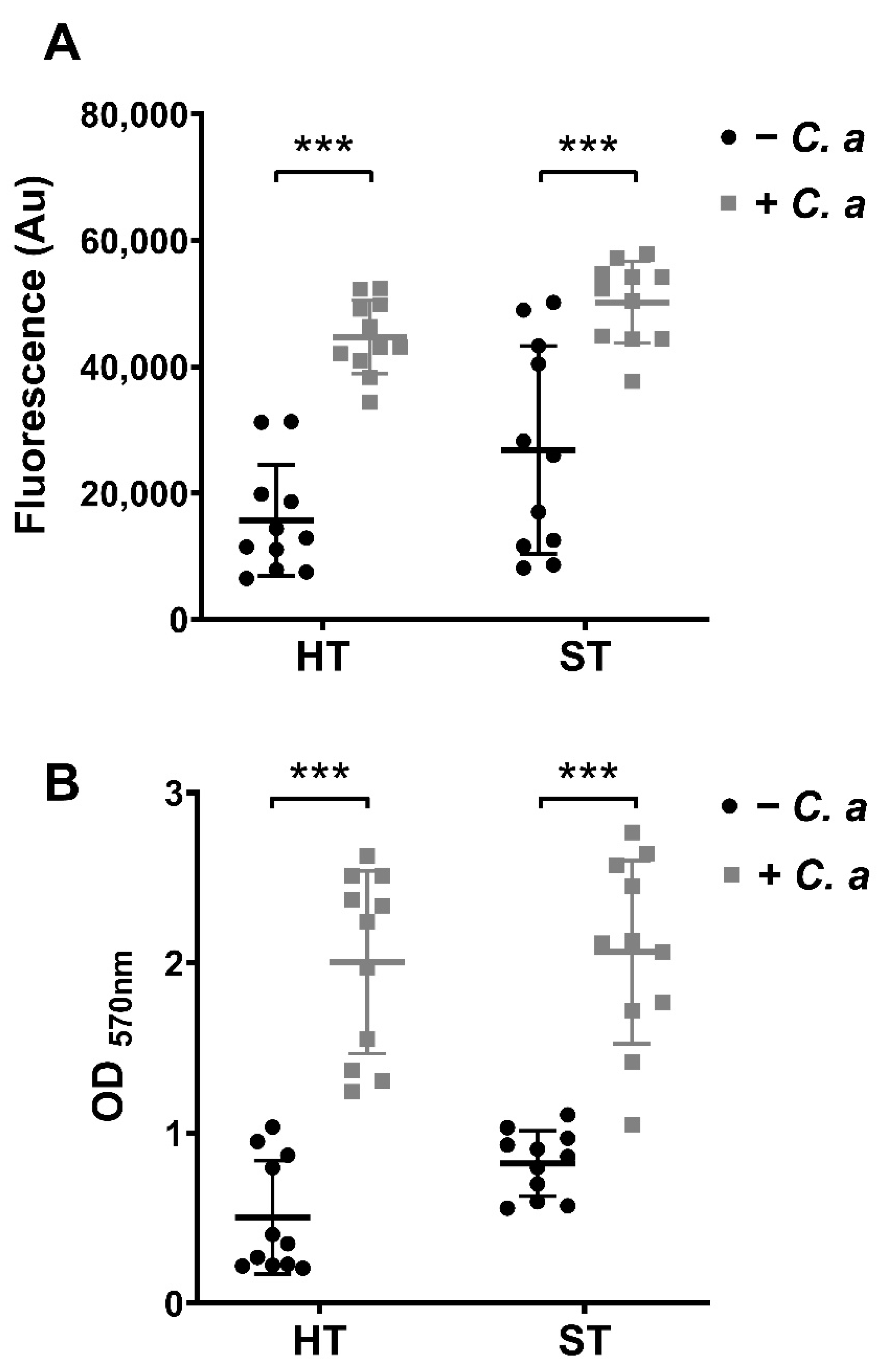
3.1. Metabolic, Biomass and Compositional Profiles of Oral Biofilm Models with or without C. albicans
3.2. Scanning Electron Microscopy Visualization of Oral Biofilm Models with or without C. albicans
3.3. Change in Pathogenic Biomarkers in Oral Biofilm Models in the Presence and Absence of C. albicans
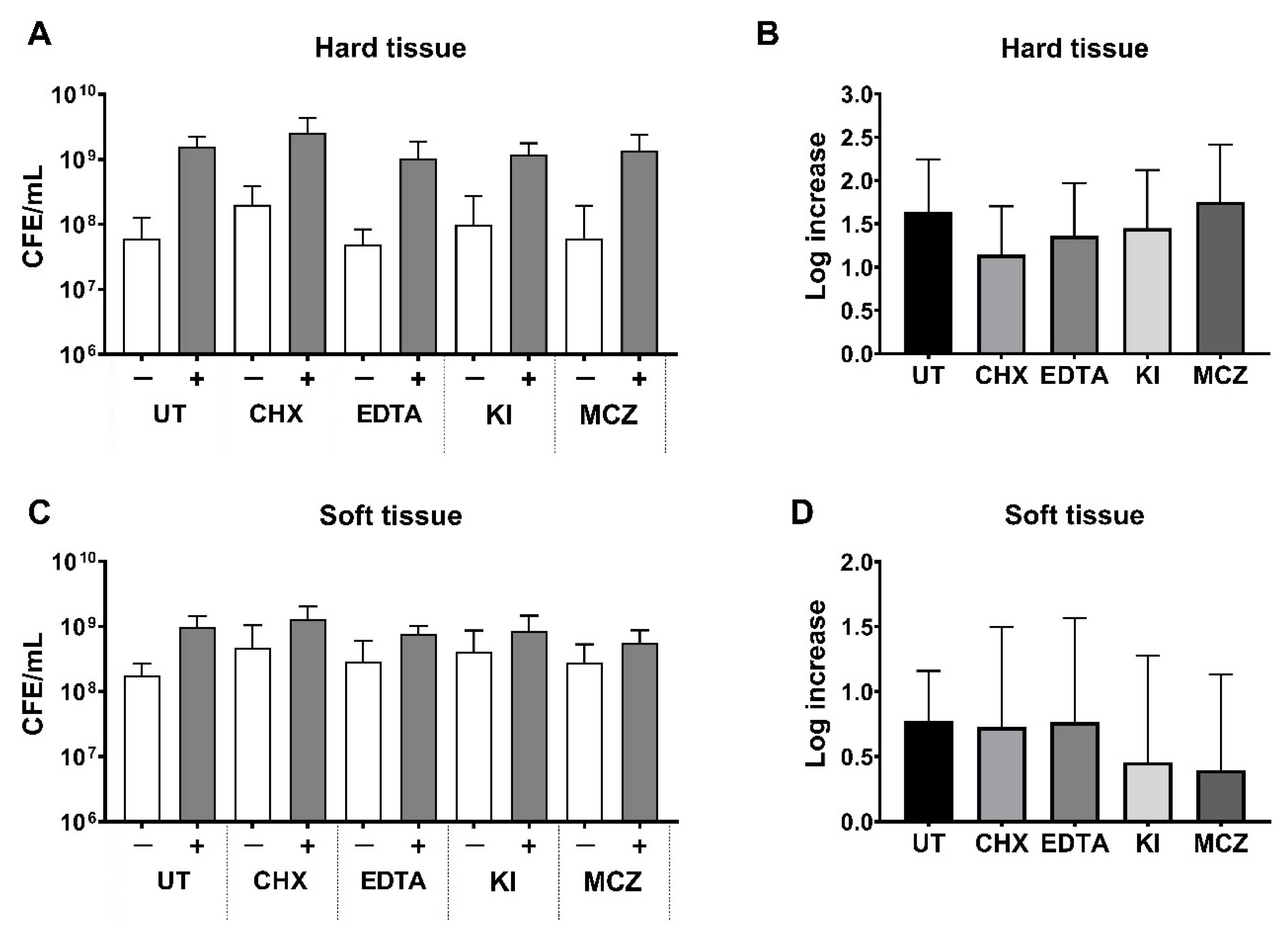
3.4. Bacterial Load Following Treatment in Oral Biofilm Models with or without C. albicans
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.R.; Yu, W.-H.; Lakshmanan, A.; Wade, W.G. The Human Oral Microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef] [Green Version]
- Marsh, P.D. The commensal microbiota and the development of human disease—An introduction. J. Oral Microbiol. 2015, 7. [Google Scholar] [CrossRef]
- Jin, L.J.; Lamster, I.B.; Greenspan, J.S.; Pitts, N.B.; Scully, C.; Warnakulasuriya, S. Global burden of oral diseases: Emerging concepts, management and interplay with systemic health. Oral Dis. 2016, 22, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Vos, T.; Abajobir, A.A.; Abate, K.H.; Abbafati, C.; Abbas, K.M.; Abd-Allah, F.; Abdulkader, R.S.; Abdulle, A.M.; Abebo, T.A.; Abera, S.F.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1211–1259. [Google Scholar] [CrossRef] [Green Version]
- W.H.O. What is the Burden of Oral Disease. Available online: https://www.who.int/oral_health/disease_burden/global/en/ (accessed on 30 January 2020).
- Zaura, E.; Keijser, B.J.F.; Huse, S.M.; Crielaard, W. Defining the healthy "core microbiome" of oral microbial communities. BMC Microbiol. 2009, 9, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bik, E.M.; Long, C.D.; Armitage, G.C.; Loomer, P.; Emerson, J.; Mongodin, E.F.; Nelson, K.E.; Gill, S.R.; Fraser-Liggett, C.M.; Relman, D.A. Bacterial diversity in the oral cavity of 10 healthy individuals. ISME J 2010, 4, 962–974. [Google Scholar] [CrossRef] [PubMed]
- Zarco, M.F.; Vess, T.J.; Ginsburg, G.S. The oral microbiome in health and disease and the potential impact on personalized dental medicine. Oral Dis. 2012, 18, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.A.; Jurevic, R.J.; Mukherjee, P.K.; Cui, F.; Sikaroodi, M.; Naqvi, A.; Gillevet, P.M. Characterization of the oral fungal microbiome (mycobiome) in healthy individuals. PLoS Pathog. 2010, 6, e1000713. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, A.K.; David, M.S.; Li, L.; Heider, T.N.; Peterson, J.D.; Montano, E.A.; Dongari-Bagtzoglou, A.; Diaz, P.I.; Strausbaugh, L.D. Redefining the Human Oral Mycobiome with Improved Practices in Amplicon-based Taxonomy: Discovery of Malassezia as a Prominent Commensal. PLoS ONE 2014, 9, e90899. [Google Scholar] [CrossRef] [Green Version]
- Delaney, C.; O’Donnell, L.E.; Kean, R.; Sherry, L.; Brown, J.L.; Calvert, G.; Nile, C.J.; Cross, L.; Bradshaw, D.J.; Brandt, B.W.; et al. Interkingdom interactions on the denture surface: Implications for oral hygiene. Biofilm 2019, 1, 100002. [Google Scholar] [CrossRef]
- Witherden, E.A.; Shoaie, S.; Hall, R.A.; Moyes, D.L. The Human Mucosal Mycobiome and Fungal Community Interactions. J. Fungi 2017, 3, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iliev, I.D.; Leonardi, I. Fungal dysbiosis: Immunity and interactions at mucosal barriers. Nat. Rev. Immunol. 2017, 17, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.A.; Noverr, M.C. Fungal interactions with the human host: Exploring the spectrum of symbiosis. Curr. Opin. Microbiol. 2017, 40, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Janus, M.M.; Willems, H.M.; Krom, B.P. Candida albicans in Multispecies Oral Communities; A Keystone Commensal? Adv. Exp. Med. Biol. 2016, 931, 13–20. [Google Scholar] [CrossRef]
- O’Donnell, L.E.; Millhouse, E.; Sherry, L.; Kean, R.; Malcolm, J.; Nile, C.J.; Ramage, G. Polymicrobial Candida biofilms: Friends and foe in the oral cavity. FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Delaney, C.; Kean, R.; Short, B.; Tumelty, M.; McLean, W.; Nile, C.J.; Ramage, G. Fungi at the Scene of the Crime: Innocent Bystanders or Accomplices in Oral Infections? Curr. Clin. Microbiol. Rep. 2018, 5, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Huang, X.; Alkhers, N.; Alzamil, H.; Alzoubi, S.; Wu, T.T.; Castillo, D.A.; Campbell, F.; Davis, J.; Herzog, K.; et al. Candida albicans and Early Childhood Caries: A Systematic Review and Meta-Analysis. Caries Res. 2018, 52, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Negrini, T.d.C.; Koo, H.; Arthur, R.A. Candida–Bacterial Biofilms and Host–Microbe Interactions in Oral Diseases. In Proceedings of the Oral Mucosal Immunity and Microbiome. Adv. Exp. Med. Biol. 2019, 1197, 119–141. [Google Scholar]
- Thurnheer, T.; Karygianni, L.; Flury, M.; Belibasakis, G.N. Fusobacterium Species and Subspecies Differentially Affect the Composition and Architecture of Supra- and Subgingival Biofilms Models. Front. Microbiol. 2019, 10, 1716. [Google Scholar] [CrossRef] [Green Version]
- Sztukowska, M.N.; Dutton, L.C.; Delaney, C.; Ramsdale, M.; Ramage, G.; Jenkinson, H.F.; Nobbs, A.H.; Lamont, R.J. Community Development between Porphyromonas gingivalis and Candida albicans Mediated by InlJ and Als3. MBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Chu Chun, H.; Mei, L.E.I.; Seneviratne Chaminda, J.; Lo Edward Chin, M.A.N. Effects of silver diamine fluoride on dentine carious lesions induced by Streptococcus mutans and Actinomyces naeslundii biofilms. Int. J. Paediatr. Dent. 2011, 22, 2–10. [Google Scholar] [CrossRef]
- Peyyala, R.; Kirakodu, S.S.; Novak, K.F.; Ebersole, J.L. Oral Epithelial Cell Responses to Multispecies Microbial Biofilms. J. Dent. Res. 2013, 92, 235–240. [Google Scholar] [CrossRef]
- Zhou, Y.; Millhouse, E.; Shaw, T.; Lappin, D.F.; Rajendran, R.; Bagg, J.; Lin, H.; Ramage, G. Evaluating Streptococcus mutans Strain Dependent Characteristics in a Polymicrobial Biofilm Community. Front. Microbiol. 2018, 9, 1498. [Google Scholar] [CrossRef] [Green Version]
- Diaz, P.I.; Xie, Z.; Sobue, T.; Thompson, A.; Biyikoglu, B.; Ricker, A.; Ikonomou, L.; Dongari-Bagtzoglou, A. Synergistic interaction between Candida albicans and commensal oral streptococci in a novel in vitro mucosal model. Infect. Immun. 2012, 80, 620–632. [Google Scholar] [CrossRef] [Green Version]
- Cavalcanti, I.M.; Nobbs, A.H.; Ricomini-Filho, A.P.; Jenkinson, H.F.; Del Bel Cury, A.A. Interkingdom cooperation between Candida albicans, Streptococcus oralis and Actinomyces oris modulates early biofilm development on denture material. Pathog. Dis. 2016, 74. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.X.; Moyes, D.L.; Richardson, J.P.; Blagojevic, M.; Naglik, J.R. Epithelial discrimination of commensal and pathogenic Candida albicans. Oral Dis. 2016, 22 (Suppl. 1), 114–119. [Google Scholar] [CrossRef]
- Desai, J.V. Candida albicans Hyphae: From Growth Initiation to Invasion. J. Fungi 2018, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Bor, B.; Cen, L.; Agnello, M.; Shi, W.; He, X. Morphological and physiological changes induced by contact-dependent interaction between Candida albicans and Fusobacterium nucleatum. Sci. Rep. 2016, 6, 27956. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Darveau, R.P.; Curtis, M.A. The keystone-pathogen hypothesis. Nat. Rev. Microbiol. 2012, 10, 717–725. [Google Scholar] [CrossRef]
- Fakhruddin, K.S.; Perera Samaranayake, L.; Egusa, H.; Chi Ngo, H.; Panduwawala, C.; Venkatachalam, T.; Kumarappan, A.; Pesee, S. Candida biome of severe early childhood caries (S-ECC) and its cariogenic virulence traits. J. Oral Microbiol. 2020, 12, 1724484. [Google Scholar] [CrossRef] [Green Version]
- Sardi, J.C.; Duque, C.; Mariano, F.S.; Peixoto, I.T.; Hofling, J.F.; Goncalves, R.B. Candida spp. in periodontal disease: A brief review. J. Oral Sci. 2010, 52, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Puryer, J. Denture Stomatitis—A Clinical Update. Dent. Update 2016, 43, 529–530, 533–525. [Google Scholar] [CrossRef] [Green Version]
- Marsh, P.D. Microbial ecology of dental plaque and its significance in health and disease. Adv. Dent. Res. 1994, 8, 263–271. [Google Scholar] [CrossRef]
- Marsh, P.D. Are dental diseases examples of ecological catastrophes? Microbiology 2003, 149, 279–294. [Google Scholar] [CrossRef]
- Allkja, J.; Bjarnsholt, T.; Coenye, T.; Cos, P.; Fallarero, A.; Harrison, J.J.; Lopes, S.P.; Oliver, A.; Pereira, M.O.; Ramage, G.; et al. Minimum information guideline for spectrophotometric and fluorometric methods to assess biofilm formation in microplates. Biofilm 2019. [Google Scholar] [CrossRef]
- Millhouse, E.; Jose, A.; Sherry, L.; Lappin, D.F.; Patel, N.; Middleton, A.M.; Pratten, J.; Culshaw, S.; Ramage, G. Development of an in vitro periodontal biofilm model for assessing antimicrobial and host modulatory effects of bioactive molecules. BMC Oral Health 2014, 14, 80. [Google Scholar] [CrossRef] [Green Version]
- Sherry, L.; Lappin, G.; O’Donnell, L.E.; Millhouse, E.; Millington, O.R.; Bradshaw, D.J.; Axe, A.S.; Williams, C.; Nile, C.J.; Ramage, G. Viable Compositional Analysis of an Eleven Species Oral Polymicrobial Biofilm. Front. Microbiol. 2016, 7, 912. [Google Scholar] [CrossRef] [Green Version]
- Pitts, N.B.; Zero, D.T.; Marsh, P.D.; Ekstrand, K.; Weintraub, J.A.; Ramos-Gomez, F.; Tagami, J.; Twetman, S.; Tsakos, G.; Ismail, A. Dental caries. Nat. Rev. Dis. Primers 2017, 3, 17030. [Google Scholar] [CrossRef] [Green Version]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent Jr, R.L. Microbial complexes in subgingival plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef]
- Brown, J.L.; Johnston, W.; Delaney, C.; Rajendran, R.; Butcher, J.; Khan, S.; Bradshaw, D.; Ramage, G.; Culshaw, S. Biofilm-stimulated epithelium modulates the inflammatory responses in co-cultured immune cells. Sci. Rep. 2019, 9, 15779. [Google Scholar] [CrossRef] [Green Version]
- Miles, A.A.; Misra, S.S.; Irwin, J.O. The estimation of the bactericidal power of the blood. J. Hyg. 1938, 38, 732–749. [Google Scholar] [CrossRef] [Green Version]
- Montelongo-Jauregui, D.; Srinivasan, A.; Ramasubramanian, A.K.; Lopez-Ribot, J.L. An In Vitro Model for Oral Mixed Biofilms of Candida albicans and Streptococcus gordonii in Synthetic Saliva. Front. Microbiol. 2016, 7, 686. [Google Scholar] [CrossRef]
- Loozen, G.; Boon, N.; Pauwels, M.; Quirynen, M.; Teughels, W. Live/dead real-time polymerase chain reaction to assess new therapies against dental plaque-related pathologies. Mol. Oral. Microbiol. 2011, 26, 253–261. [Google Scholar] [CrossRef]
- Periasamy, S.; Chalmers, N.I.; Du-Thumm, L.; Kolenbrander, P.E. Fusobacterium nucleatum ATCC 10953 requires Actinomyces naeslundii ATCC 43146 for growth on saliva in a three-species community that includes Streptococcus oralis 34. Appl. Enviorn. Microbiol. 2009, 75, 3250–3257. [Google Scholar] [CrossRef] [Green Version]
- Sherry, L.; Millhouse, E.; Lappin, D.F.; Murray, C.; Culshaw, S.; Nile, C.J.; Ramage, G. Investigating the biological properties of carbohydrate derived fulvic acid (CHD-FA) as a potential novel therapy for the management of oral biofilm infections. BMC Oral Health 2013, 13, 47. [Google Scholar] [CrossRef] [Green Version]
- Park, S.N.; Park, J.Y.; Kook, J.K. Development of Porphyromonas gingivalis-specific quantitative real-time PCR primers based on the nucleotide sequence of rpoB. J. Microbiol. 2011, 49, 315–319. [Google Scholar] [CrossRef]
- Periasamy, S.; Kolenbrander, P.E. Mutualistic biofilm communities develop with Porphyromonas gingivalis and initial, early, and late colonizers of enamel. J. Bacteriol. 2009, 191, 6804–6811. [Google Scholar] [CrossRef] [Green Version]
- Desai, A.R.; Shah, N.P.; Powell, I.B. Discrimination of dairy industry isolates of the Lactobacillus casei group. J. Dairy Sci. 2006, 89, 3345–3351. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Yoshida, A.; Saito, T.; Kawada, M.; Nakano, Y. Quantitative microbiological study of subgingival plaque by real-time PCR shows correlation between levels of Tannerella forsythensis and Fusobacterium spp. J. Clin. Microbiol. 2004, 42, 2255–2257. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, R.; Borghi, E.; Falleni, M.; Perdoni, F.; Tosi, D.; Lappin, D.F.; O’Donnell, L.; Greetham, D.; Ramage, G.; Nile, C. Acetylcholine Protects against Candida albicans Infection by Inhibiting Biofilm Formation and Promoting Hemocyte Function in a Galleria mellonella Infection Model. Eukaryot Cell 2015, 14, 834–844. [Google Scholar] [CrossRef] [Green Version]
- Erlandsen, S.L.; Kristich, C.J.; Dunny, G.M.; Wells, C.L. High-resolution visualization of the microbial glycocalyx with low-voltage scanning electron microscopy: Dependence on cationic dyes. J. Histochem. Cytochem. 2004, 52, 1427–1435. [Google Scholar] [CrossRef]
- James, P.; Worthington, H.V.; Parnell, C.; Harding, M.; Lamont, T.; Cheung, A.; Whelton, H.; Riley, P. Chlorhexidine mouthrinse as an adjunctive treatment for gingival health. Cochrane Database Syst. Rev. 2017, 3, Cd008676. [Google Scholar] [CrossRef]
- McComb, D.; Smith, D.C. A preliminary scanning electron microscopic study of root canals after endodontic procedures. J. Endod. 1975, 1, 238–242. [Google Scholar] [CrossRef]
- Safavi, K.E.; Spangberg, L.S.; Langeland, K. Root canal dentinal tubule disinfection. J. Endod. 1990, 16, 207–210. [Google Scholar] [CrossRef]
- Kean, R.; Rajendran, R.; Haggarty, J.; Townsend, E.M.; Short, B.; Burgess, K.E.; Lang, S.; Millington, O.; Mackay, W.G.; Williams, C.; et al. Candida albicans Mycofilms Support Staphylococcus aureus Colonization and Enhances Miconazole Resistance in Dual-Species Interactions. Front. Microbiol. 2017, 8, 258. [Google Scholar] [CrossRef] [Green Version]
- Marsh, P.D. Sugar, fluoride, pH and microbial homeostasis in dental plaque. Proc. Finn. Dent. Soc. 1991, 87, 515–525. [Google Scholar]
- Zaura, E.; Brandt, B.W.; Prodan, A.; Teixeira de Mattos, M.J.; Imangaliyev, S.; Kool, J.; Buijs, M.J.; Jagers, F.L.; Hennequin-Hoenderdos, N.L.; Slot, D.E.; et al. On the ecosystemic network of saliva in healthy young adults. ISME J. 2017, 11, 1218–1231. [Google Scholar] [CrossRef]
- Tomofuji, T.; Irie, K.; Sanbe, T.; Azuma, T.; Ekuni, D.; Tamaki, N.; Yamamoto, T.; Morita, M. Periodontitis and increase in circulating oxidative stress. Jpn. Dent. Sci. Rev. 2009, 45, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Culp, E.; Wright, G.D. Bacterial proteases, untapped antimicrobial drug targets. J. Antibiot. 2017, 70, 366–377. [Google Scholar] [CrossRef] [Green Version]
- Vylkova, S.; Carman, A.J.; Danhof, H.A.; Collette, J.R.; Zhou, H.; Lorenz, M.C. The fungal pathogen Candida albicans autoinduces hyphal morphogenesis by raising extracellular pH. mBio 2011, 2, e00055-11. [Google Scholar] [CrossRef] [Green Version]
- Dahiya, P.; Kamal, R.; Gupta, R.; Bhardwaj, R.; Chaudhary, K.; Kaur, S. Reactive oxygen species in periodontitis. J. Indian Soc. Periodontol. 2013, 17, 411–416. [Google Scholar] [CrossRef]
- Sonesson, A.; Przybyszewska, K.; Eriksson, S.; Mörgelin, M.; Kjellström, S.; Davies, J.; Potempa, J.; Schmidtchen, A. Identification of bacterial biofilm and the Staphylococcus aureus derived protease, staphopain, on the skin surface of patients with atopic dermatitis. Sci. Rep. 2017, 7, 8689. [Google Scholar] [CrossRef]
- Huffnagle, G.B.; Noverr, M.C. The emerging world of the fungal microbiome. Trends Microbiol. 2013, 21, 334–341. [Google Scholar] [CrossRef] [Green Version]
- Ramage, G.; Martínez, J.P.; López-Ribot, J.L. Candida biofilms on implanted biomaterials: A clinically significant problem. FEMS Yeast Res. 2006, 6, 979–986. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Verma, R.; Murari, A.; Agrawal, A. Oral candidiasis: An overview. J. Oral Maxillofac. Pathol. 2014, 18, S81–S85. [Google Scholar] [CrossRef]
- Kirchner, F.R.; Littringer, K.; Altmeier, S.; Tran, V.D.T.; Schonherr, F.; Lemberg, C.; Pagni, M.; Sanglard, D.; Joller, N.; LeibundGut-Landmann, S. Persistence of Candida albicans in the Oral Mucosa Induces a Curbed Inflammatory Host Response That Is Independent of Immunosuppression. Front. Immunol. 2019, 10, 330. [Google Scholar] [CrossRef] [Green Version]
- Enoch, D.A.; Yang, H.; Aliyu, S.H.; Micallef, C. The Changing Epidemiology of Invasive Fungal Infections. Methods Mol. Biol. 2017, 1508, 17–65. [Google Scholar] [CrossRef]
- Bassetti, M.; Righi, E.; Montravers, P.; Cornely, O.A. What has changed in the treatment of invasive candidiasis? A look at the past 10 years and ahead. J. Antimicrob. Chemother. 2018, 73, i14–i25. [Google Scholar] [CrossRef] [Green Version]
- Costa-Orlandi, C.B.; Sardi, J.C.O.; Pitangui, N.S.; de Oliveira, H.C.; Scorzoni, L.; Galeane, M.C.; Medina-Alarcon, K.P.; Melo, W.; Marcelino, M.Y.; Braz, J.D.; et al. Fungal Biofilms and Polymicrobial Diseases. J. Fungi 2017, 3, 22. [Google Scholar] [CrossRef]
- Rodrigues, M.E.; Gomes, F.; Rodrigues, C.F. Candida spp./Bacteria Mixed Biofilms. J. Fungi 2019, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Rampersad, S.N. Multiple Applications of Alamar Blue as an Indicator of Metabolic Function and Cellular Health in Cell Viability Bioassays. Sensors 2012, 12, 12347–12360. [Google Scholar] [CrossRef]
- Haney, E.F.; Trimble, M.J.; Cheng, J.T.; Vallé, Q.; Hancock, R.E.W. Critical Assessment of Methods to Quantify Biofilm Growth and Evaluate Antibiofilm Activity of Host Defence Peptides. Biomolecules 2018, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Ramage, G. Comparing apples and oranges: Considerations for quantifying candidal biofilms with XTT [2,3-bis(2-methoxy-4-nitro-5-sulfo-phenyl)-2H-tetrazolium-5-carboxanilide] and the need for standardized testing. J. Med. Microbiol. 2016, 65, 259–260. [Google Scholar] [CrossRef]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Di Bonaventura, G.; Hébraud, M.; Jaglic, Z.; et al. Critical review on biofilm methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef] [Green Version]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef]
- Falsetta, M.L.; Klein, M.I.; Colonne, P.M.; Scott-Anne, K.; Gregoire, S.; Pai, C.-H.; Gonzalez-Begne, M.; Watson, G.; Krysan, D.J.; Bowen, W.H.; et al. Symbiotic Relationship between Streptococcus mutans and Candida albicans Synergizes Virulence of Plaque Biofilms In Vivo. Infect. Immun. 2014, 82, 1968. [Google Scholar] [CrossRef] [Green Version]
- Gross, E.L.; Beall, C.J.; Kutsch, S.R.; Firestone, N.D.; Leys, E.J.; Griffen, A.L. Beyond Streptococcus mutans: Dental caries onset linked to multiple species by 16S rRNA community analysis. PLoS ONE 2012, 7, e47722. [Google Scholar] [CrossRef]
- Fox, E.P.; Cowley, E.S.; Nobile, C.J.; Hartooni, N.; Newman, D.K.; Johnson, A.D. Anaerobic bacteria grow within Candida albicans biofilms and induce biofilm formation in suspension cultures. Curr. Biol. 2014, 24, 2411–2416. [Google Scholar] [CrossRef] [Green Version]
- Baliga, S.; Muglikar, S.; Kale, R. Salivary pH: A diagnostic biomarker. J. Indian Soc. Periodontol. 2013, 17, 461–465. [Google Scholar] [CrossRef]
- Rane, H.S.; Hayek, S.R.; Frye, J.E.; Abeyta, E.L.; Bernardo, S.M.; Parra, K.J.; Lee, S.A. Candida albicans Pma1p Contributes to Growth, pH Homeostasis, and Hyphal Formation. Front. Microbiol. 2019, 10, 1012. [Google Scholar] [CrossRef] [Green Version]
- Periasamy, S.; Kolenbrander, P.E. Central Role of the Early Colonizer Veillonella sp. in Establishing Multispecies Biofilm Communities with Initial, Middle, and Late Colonizers of Enamel. J. Bacteriol. 2010, 192, 2965. [Google Scholar] [CrossRef] [Green Version]
- Ene, I.V.; Adya, A.K.; Wehmeier, S.; Brand, A.C.; MacCallum, D.M.; Gow, N.A.; Brown, A.J. Host carbon sources modulate cell wall architecture, drug resistance and virulence in a fungal pathogen. Cell Microbiol. 2012, 14, 1319–1335. [Google Scholar] [CrossRef] [Green Version]
- Gambino, M.; Cappitelli, F. Mini-review: Biofilm responses to oxidative stress. Biofouling 2016, 32, 167–178. [Google Scholar] [CrossRef]
- Tally, F.P.; Goldin, B.R.; Jacobus, N.V.; Gorbach, S.L. Superoxide dismutase in anaerobic bacteria of clinical significance. Infect. Immun. 1977, 16, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Fotedar, R.; Al-Hedaithy, S.S.A. Comparison of phospholipase and proteinase activity in Candida albicans and C. dubliniensis. Mycoses 2005, 48, 62–67. [Google Scholar] [CrossRef]
- Cassone, A.; De Bernardis, F.; Mondello, F.; Ceddia, T.; Agatensi, L. Evidence for a correlation between proteinase secretion and vulvovaginal candidosis. J. Infect. Dis. 1987, 156, 777–783. [Google Scholar] [CrossRef]
- Shen, Y.; Zhao, J.; de la Fuente-Nunez, C.; Wang, Z.; Hancock, R.E.; Roberts, C.R.; Ma, J.; Li, J.; Haapasalo, M.; Wang, Q. Experimental and Theoretical Investigation of Multispecies Oral Biofilm Resistance to Chlorhexidine Treatment. Sci. Rep. 2016, 6, 27537. [Google Scholar] [CrossRef]
- Hope, C.K.; Wilson, M. Analysis of the effects of chlorhexidine on oral biofilm vitality and structure based on viability profiling and an indicator of membrane integrity. Antimicrob. Agents Chemother. 2004, 48, 1461–1468. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Wang, S.; Zhou, X.; Zou, Y.; Li, M.; Peng, X.; Ren, B.; Xu, H.H.K.; Weir, M.D.; Cheng, L.; et al. Short-Time Antibacterial Effects of Dimethylaminododecyl Methacrylate on Oral Multispecies Biofilm In Vitro. Biomed. Res. Int. 2019, 2019, 6393470. [Google Scholar] [CrossRef]
- Cavaliere, R.; Ball, J.L.; Turnbull, L.; Whitchurch, C.B. The biofilm matrix destabilizers, EDTA and DNaseI, enhance the susceptibility of nontypeable Hemophilus influenzae biofilms to treatment with ampicillin and ciprofloxacin. Microbiologyopen 2014, 3, 557–567. [Google Scholar] [CrossRef]
- Ramage, G.; Wickes, B.L.; Lopez-Ribot, J.L. Inhibition on Candida albicans biofilm formation using divalent cation chelators (EDTA). Mycopathologia 2007, 164, 301–306. [Google Scholar] [CrossRef]
- Liu, F.; Hansra, S.; Crockford, G.; Koster, W.; Allan, B.J.; Blondeau, J.M.; Lainesse, C.; White, A.P. Tetrasodium EDTA Is Effective at Eradicating Biofilms Formed by Clinically Relevant Microorganisms from Patients’ Central Venous Catheters. mSphere 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Castaneda, P.; McLaren, A.; Tavaziva, G.; Overstreet, D. Biofilm Antimicrobial Susceptibility Increases With Antimicrobial Exposure Time. Clin. Orthop. Relat. Res. 2016, 474, 1659–1664. [Google Scholar] [CrossRef]
- Arias, L.S.; Brown, J.L.; Butcher, M.C.; Delaney, C.; Monteiro, D.R.; Ramage, G. A nanocarrier system that potentiates the effect of miconazole within different interkingdom biofilms. J. Oral. Microbiol. 2020, 12, 1771071. [Google Scholar] [CrossRef]
- Chatzigiannidou, I.; Teughels, W.; Van de Wiele, T.; Boon, N. Oral biofilms exposure to chlorhexidine results in altered microbial composition and metabolic profile. NPJ Biofilms Microbiomes 2020, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Sahni, K.; Khashai, F.; Forghany, A.; Krasieva, T.; Wilder-Smith, P. Exploring Mechanisms of Biofilm Removal. Dent. (Sunnyvale) 2016, 6, 371. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | References |
|---|---|---|
| A. actinomycetemcomitans | F-GAACCTTACCTACTCTTGACATCCGAA R-TGCAGCACCTGTCTCAAAGC | [44] |
| A. naeslundii | F-GGCTGCGATACCGTGAGG R-TCTGCGATTACTAGCGACTCC | [45] |
| C. albicans | F- CTCGTAGTTGAACCTTGGGC R- GGCCTGCTTTGAACACTCTA | [38] |
| F. nucleatum spp. | F-GGATTTATTGGGCGTAAAGC R-GGCATTCCTACAAATATCTACGAA | [46] |
| P. intermedia | F-CGGTCTGTTAAGCGTGTTGTG R-CACCATGAATTCCGCATACG | [44] |
| P. gingivalis | F-GGAAGAGAAGACCGTAGCACAAGGA R-GAGTAGGCGAAACGTCCATCAGGTC | [47] |
| Streptococcus spp. | F-GATACATAGCCGACCTGAG R-CCATTGCCGAAGATTCC | [46] |
| V. dispar | F-CCGTGATGGGATGGAAACTGC R-CCTTCGCCACTGGTGTTCTTC | [48] |
| L. casei | F-TGCACTGAGATTCGACTTAA R-CCCACTGCTGCCTCCCGTAGGAGT | [49] |
| 16S | F-CGCTAGTAATCGTGGATCAGAATG R-TGTGACGGGCGGTGTGTA | [50] |
| 18S | F-CTCGTAGTTGAACCTTGGGC R-GGCCTGCTTTGAACACTCTA | [51] |
| Genus or Species | Mean CFE/mL and Percentage Composition (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Hard Tissue | Soft Tissue | |||||||
| − C. albicans | + C. albicans | − C. albicans | + C. albicans | |||||
| CFE/mL | % | CFE/mL | % | CFE/mL | % | CFE/mL | % | |
| C. a | n/a | n/a | 4.60 × 107 | 12.81 | n/a | n/a | 6.08 × 106 | 4.00 |
| Strep | 7.12 × 106 | 43.10 | 1.04 × 108 | 29.00 | 2.87 × 106 | 14.40 | 4.96 × 106 | 3.26 |
| A. n | 1.29 × 106 | 7.82 | 4.25 × 106 | 1.18 | 9.50 × 105 | 4.76 | 2.36 × 106 | 1.55 |
| V. d | 8.10 × 106 | 49.05 | 2.03 × 108 | 56.63 | 1.58 × 107 | 79.27 | 1.36 × 108 | 89.87 |
| F. n | 2.91 × 103 | 0.02 | 1.38 × 106 | 0.38 | 2.80 × 103 | 0.014 | 5.12 × 103 | 0.05 |
| L. c | 1.99 × 103 | 0.01 | 4.19 × 103 | 0.001 | n/a | n/a | n/a | n/a |
| P. g | n/a | n/a | n/a | n/a | 1.23 × 102 | 0.001 | 1.74 × 104 | 0.011 |
| P. i | n/a | n/a | n/a | n/a | 5.49 × 102 | 0.003 | 5.12 × 103 | 0.003 |
| A. a | n/a | n/a | n/a | n/a | 3.11 × 105 | 1.56 | 1.90 × 106 | 1.25 |
| Biofilm | Treatment | Average CFE/mL | |||
|---|---|---|---|---|---|
| 16S | 18S | ||||
| − C. a | + C. a | − C. a | + C. a | ||
| ST | UT | 1.77 × 108 | 9.96 × 108 | n/a | 6.08 × 106 |
| CHX | 4.74 × 108 | 1.30 × 109 | n/a | 4.31 × 106 | |
| EDTA | 2.91 × 108 | 7.66 × 108 | n/a | 3.91 × 106 | |
| KI | 4.13 × 108 | 8.46 × 108 | n/a | 3.32 × 106 | |
| MCZ | 2.79 × 108 | 5.60 × 108 | n/a | 3.21 × 106 | |
| HT | UT | 6.00 × 107 | 1.59 × 109 | n/a | 4.60 × 107 |
| CHX | 2.02 × 108 | 2.59 × 109 | n/a | 4.21 × 107 | |
| EDTA | 4.91 × 107 | 1.04 × 109 | n/a | 3.34 × 107 | |
| KI | 9.84 × 107 | 1.18 × 109 | n/a | 3.11 × 107 | |
| MCZ | 6.13 × 107 | 1.36× 109 | n/a | 3.52 × 107 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Young, T.; Alshanta, O.-A.; Kean, R.; Bradshaw, D.; Pratten, J.; Williams, C.; Woodall, C.; Ramage, G.; Brown, J.L. Candida albicans as an Essential “Keystone” Component within Polymicrobial Oral Biofilm Models? Microorganisms 2021, 9, 59. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010059
Young T, Alshanta O-A, Kean R, Bradshaw D, Pratten J, Williams C, Woodall C, Ramage G, Brown JL. Candida albicans as an Essential “Keystone” Component within Polymicrobial Oral Biofilm Models? Microorganisms. 2021; 9(1):59. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010059
Chicago/Turabian StyleYoung, Tracy, Om-Alkhir Alshanta, Ryan Kean, David Bradshaw, Jonathan Pratten, Craig Williams, Chris Woodall, Gordon Ramage, and Jason L. Brown. 2021. "Candida albicans as an Essential “Keystone” Component within Polymicrobial Oral Biofilm Models?" Microorganisms 9, no. 1: 59. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010059