Diversity and Antimicrobial Resistance in the Streptococcus bovis/Streptococcus equinus Complex (SBSEC) Isolated from Korean Domestic Ruminants

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care
2.2. Collection of Rumen Fluids
2.3. Bacterial Isolation and Identification
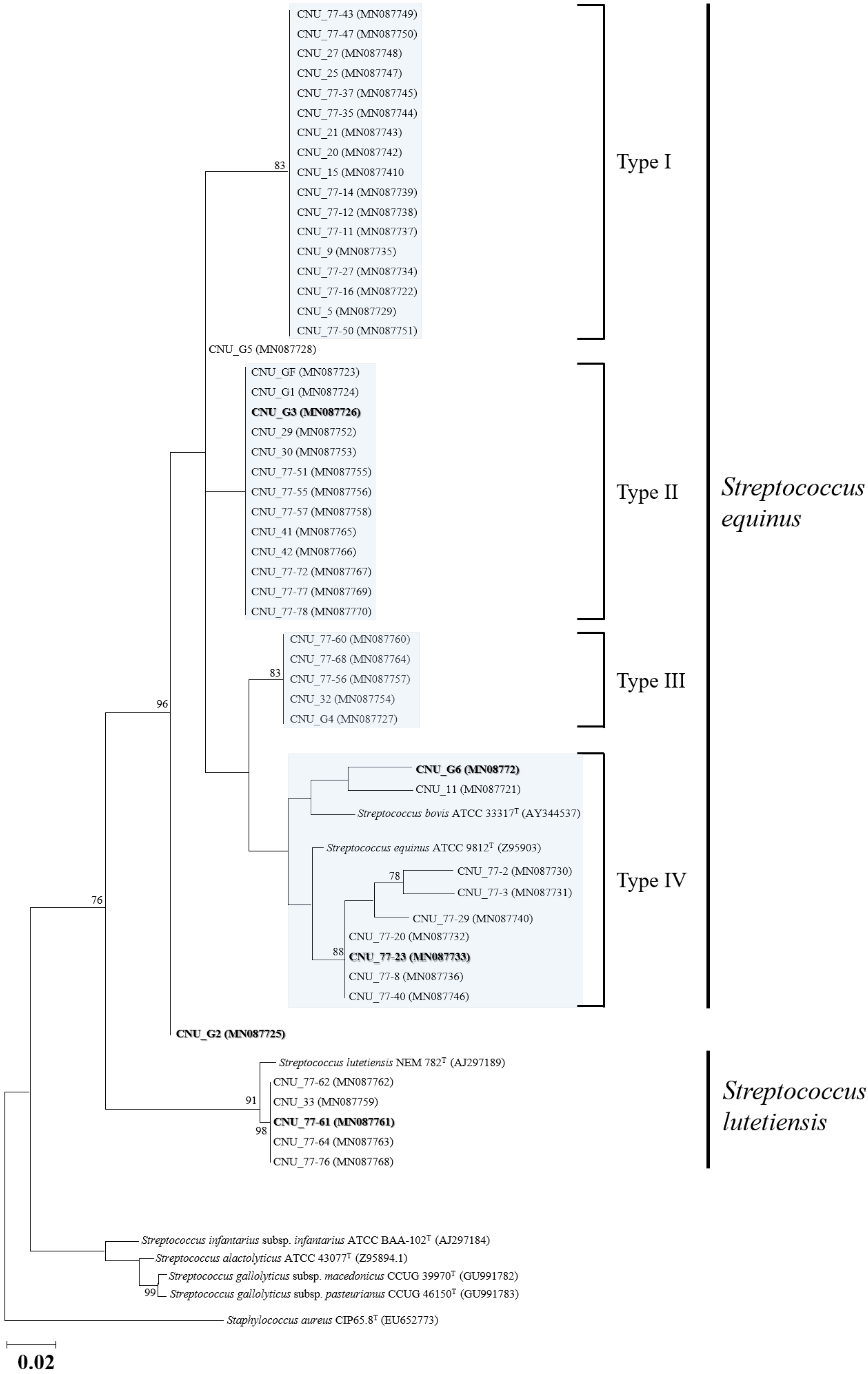
2.4. Species Discrimination and Phylogenetic Analysis
2.5. Antimicrobial Susceptibility Test
2.6. Genome Sequencing of the SBSEC Isolates
2.7. Determination of Antimicrobial-resistant Genes and L-lactate Dehydrogenase Genes in the SBSEC Isolates
2.8. Accession Numbers of Nucleotide Sequences and Strain Deposition
3. Results
3.1. SBSEC Isolation and Identification
3.2. Antimicrobial Susceptibility of the SBSEC Isolates
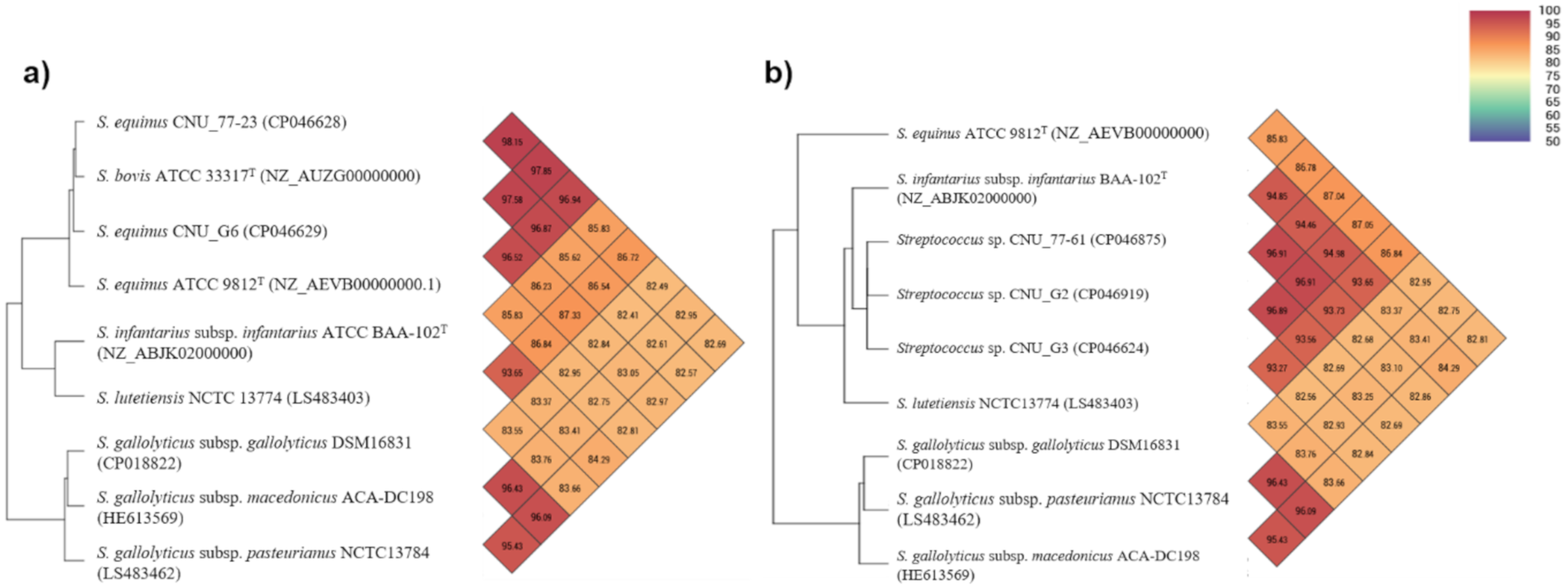
3.3. Genomic Features of the SBSEC Isolates
3.4. Antimicrobial-Resistant Genes and L-lactate Dehydrogenase Genes in the SBSEC Isolates
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Jouany, J.; Michalet-Doreau, B.; Doreau, M. Manipulation of the rumen ecosystem to support high-performance beef cattle-Review. Asian-Australas. J. Anim. Sci. 2000, 13, 96–114. [Google Scholar]
- Tajima, K.; Arai, S.; Ogata, K.; Nagamine, T.; Matsui, H.; Nakamura, M.; Aminov, R.I.; Benno, Y. Rumen bacterial community transition during adaptation to high-grain diet. Anaerobe 2000, 6, 273–284. [Google Scholar] [CrossRef]
- Chen, L.; Shen, Y.; Wang, C.; Ding, L.; Zhao, F.; Wang, M.; Fu, J.; Wang, H. Megasphaera elsdenii lactate degradation pattern shifts in rumen acidosis models. Front. Microbiol. 2019, 10, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hungate, R.E. The Rumen and Its Microbes; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Russell, J.B. Rumen Microbiology and Its Role in Ruminant Nutrition; Department of Microbiology, Cornell University: Ithaca, NY, USA, 2002. [Google Scholar]
- Freer, S.N. Purification and characterization of the extracellular alpha-amylase from Streptococcus bovis JB1. Appl. Environ. Microbiol. 1993, 59, 1398–1402. [Google Scholar] [CrossRef] [Green Version]
- Asanuma, N.; Hino, T. Regulation of fermentation in a ruminal bacterium, Streptococcus bovis, with special reference to rumen acidosis. Anim. Sci. J. 2002, 73, 313–325. [Google Scholar] [CrossRef]
- Enemark, J.M. The monitoring, prevention and treatment of sub-acute ruminal acidosis (SARA): A review. Vet. J. 2008, 176, 32–43. [Google Scholar] [CrossRef]
- Herrera, P.; Kwon, Y.M.; Ricke, S.C. Ecology and pathogenicity of gastrointestinal Streptococcus bovis. Anaerobe 2009, 15, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Lean, I.; Wade, L.; Curtis, M.; Porter, J. New approaches to control of ruminal acidosis in dairy cattle. Asian Australas. J. Anim. Sci. 2000, 13, 266–269. [Google Scholar]
- Jans, C.; Meile, L.; Lacroix, C.; Stevens, M.J. Genomics, evolution, and molecular epidemiology of the Streptococcus bovis/Streptococcus equinus complex (SBSEC). Infect. Genet. Evol. 2015, 33, 419–436. [Google Scholar] [CrossRef]
- Jans, C.; Meile, L.; Kaindi, D.W.M.; Kogi-Makau, W.; Lamuka, P.; Renault, P.; Kreikemeyer, B.; Lacroix, C.; Hattendorf, J.; Zinsstag, J. African fermented dairy products—Overview of predominant technologically important microorganisms focusing on African Streptococcus infantarius variants and potential future applications for enhanced food safety and security. Int. J. Food Microbiol. 2017, 250, 27–36. [Google Scholar] [CrossRef]
- Parker, M.; Ball, L.C. Streptococci and aerococci associated with systemic infection in man. J. Med. Microbiol. 1976, 9, 275–302. [Google Scholar] [CrossRef] [PubMed]
- Dekker, J.P.; Lau, A.F. An update on the Streptococcus bovis group: Classification, identification, and disease associations. J. Clin. Microbiol. 2016, 54, 1694–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pompilio, A.; Di Bonaventura, G.; Gherardi, G. An overview on Streptococcus bovis/Streptococcus equinus complex isolates: Identification to the species/subspecies level and antibiotic resistance. Int. J. Mol. Sci. 2019, 20, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jans, C.; Boleij, A. The road to infection: Host-microbe interactions defining the pathogenicity of Streptococcus bovis/Streptococcus equinus complex members. Front. Microbiol. 2018, 9, 603. [Google Scholar] [CrossRef] [Green Version]
- Nagaraja, T.; Titgemeyer, E. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook. J. Dairy Sci. 2007, 90, E17–E38. [Google Scholar] [CrossRef] [Green Version]
- Fernando, S.C.; Purvis, H.T.; Najar, F.Z.; Sukharnikov, L.O.; Krehbiel, C.R.; Nagaraja, T.G.; Roe, B.A.; DeSilva, U. Rumen microbial population dynamics during adaptation to a high-grain diet. Appl. Environ. Microbiol. 2010, 76, 7482–7490. [Google Scholar] [CrossRef] [Green Version]
- Petri, R.M.; Schwaiger, T.; Penner, G.B.; Beauchemin, K.A.; Forster, R.J.; McKinnon, J.J.; McAllister, T.A. Characterization of the core rumen microbiome in cattle during transition from forage to concentrate as well as during and after an acidotic challenge. PLoS ONE 2013, 8, e83424. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, G.N.; Kurtovic, A.; Hay, A.G.; Russell, J.B. The physiological and genetic diversity of bovine Streptococcus bovis strains. FEMS Microbiol. Ecol. 2001, 35, 49–56. [Google Scholar] [CrossRef]
- Klieve, A.V.; Heck, G.L.; Prance, M.A.; Shu, Q. Genetic homogeneity and phage susceptibility of ruminal strains of Streptococcus bovis isolated in Australia. Lett. Appl. Microbiol. 1999, 29, 108–112. [Google Scholar] [CrossRef]
- Jans, C.; de Wouters, T.; Bonfoh, B.; Lacroix, C.; Kaindi, D.W.M.; Anderegg, J.; Böck, D.; Vitali, S.; Schmid, T.; Isenring, J. Phylogenetic, epidemiological and functional analyses of the Streptococcus bovis/Streptococcus equinus complex through an overarching MLST scheme. BMC Microbiol. 2016, 16, 117. [Google Scholar] [CrossRef] [Green Version]
- Marshall, B.M.; Levy, S.B. Food animals and antimicrobials: Impacts on human health. Clin. Microbiol. Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Yang, Q.E.; Zhou, X.; Wang, F.-H.; Muurinen, J.; Virta, M.P.; Brandt, K.K.; Zhu, Y.-G. Antibiotic resistome in the livestock and aquaculture industries: Status and solutions. Crit. Rev. Environ. Sci. Technol. 2020, 1–38. [Google Scholar] [CrossRef]
- Economou, V.; Gousia, P. Agriculture and food animals as a source of antimicrobial-resistant bacteria. Infect. Drug Resist. 2015, 8, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Guo, X.; Yan, Z.; Wang, W.; Chen, B.; Ge, F.; Ye, B. A comprehensive analysis on spread and distribution characteristic of antibiotic resistance genes in livestock farms of Southeastern China. PLoS ONE 2016, 11, e0156889. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.-M.; Lim, S.-K.; Kang, H.-M.; Kim, J.-M.; Moon, J.-S.; Jang, K.-C.; Joo, Y.-S.; Kang, M.-I.; Jung, S.-C. Antimicrobial resistance of streptococci isolated from mastitic bovine milk samples in Korea. J. Vet. Diagn. Investig. 2009, 21, 698–701. [Google Scholar] [CrossRef]
- Nomoto, R.; Le, H.T.T.; Sekizaki, T.; Osawa, R. Antimicrobial susceptibility of Streptococcus gallolyticus isolated from humans and animals. Jpn. J. Infect Dis. 2013, 66, 334–336. [Google Scholar] [CrossRef] [Green Version]
- Romero-Hernández, B.; Tedim, A.P.; Sánchez-Herrero, J.F.; Librado, P.; Rozas, J.; Muñoz, G.; Baquero, F.; Cantón, R.; Del Campo, R. Streptococcus gallolyticus subsp. gallolyticus from human and animal origins: Genetic diversity, antimicrobial susceptibility, and characterization of a vancomycin-resistant calf isolate carrying a vanA-Tn1546-like element. Antimicrob. Agents Chemother. 2015, 59, 2006–2015. [Google Scholar]
- Lee, M. Diversity of Rumen Bacterial Community and Streptococcus bovis Strains in Korean Domestic Ruminants Fed a High-Concentrate Diet. Master’s Thesis, Chungnam National University, Daejeon, Korea, 2016. [Google Scholar]
- Lee, M.; Jeong, S.; Seo, J.; Seo, S. Changes in the ruminal fermentation and bacterial community structure by a sudden change to a high-concentrate diet in Korean domestic ruminants. Asian-Australas. J. Anim. Sci. 2019, 32, 92. [Google Scholar] [CrossRef] [Green Version]
- Makkar, H.P.; McSweeney, C.S. Methods in Gut Microbial Ecology for Ruminants; Springer: Dordrecht, The Netherlands, 2005. [Google Scholar]
- Poyart, C.; Quesne, G.; Trieu-Cuot, P. Taxonomic dissection of the Streptococcus bovis group by analysis of manganese-dependent superoxide dismutase gene (sodA) sequences: Reclassification of ‘Streptococcus infantarius subsp. coli’ as Streptococcus lutetiensis sp. nov. and of Streptococcus bovis biotype 11.2 as Streptococcus pasteurianus sp. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 1247–1255. [Google Scholar]
- Romero, B.; Morosini, M.-I.; Loza, E.; Rodríguez-Baños, M.; Navas, E.; Cantón, R.; del Campo, R. Reidentification of Streptococcus bovis isolates causing bacteremia according to the new taxonomy criteria: Still an issue? J. Clin. Microbiol. 2011, 49, 3228–3233. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals M31-A2, 2nd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2002. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals VET01S, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, NJ, USA, 2015. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing M100S, 29th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- Jun, L. Characterization of Antibiotic Resistant Genes Carried by Fish Pathogens in Korea. Ph.D. Thesis, Pukyong National University, Busan, Korea, 2010. [Google Scholar]
- Lüthje, P.; Schwarz, S. Molecular basis of resistance to macrolides and lincosamides among staphylococci and streptococci from various animal sources collected in the resistance monitoring program BfT-GermVet. Int. J. Antimicrob. Agents 2007, 29, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Olsen, R.H.; Shi, L.; Ye, L.; He, J.; Meng, H. Characterization of a plasmid carrying cat, ermB and tetS genes in a foodborne Listeria monocytogenes strain and uptake of the plasmid by cariogenic Streptococcus mutans. Int. J. Food Microbiol. 2016, 238, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Volokhov, D.; Chizhikov, V.; Chumakov, K.; Rasooly, A. Microarray analysis of erythromycin resistance determinants. J. Appl. Microbiol. 2003, 95, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, R.; Huet, C.; Picherot, M.; Trieu-Cuot, P.; Poyart, C. Genetic basis of antibiotic resistance in clinical isolates of Streptococcus gallolyticus (Streptococcus bovis). Antimicrob. Agents Chemother. 2005, 49, 1646–1648. [Google Scholar] [CrossRef] [Green Version]
- De Vries, L.E.; Christensen, H.; Skov, R.L.; Aarestrup, F.M.; Agersø, Y. Diversity of the tetracycline resistance gene tet(M) and identification of Tn916-and Tn5801-like (Tn 6014) transposons in Staphylococcus aureus from humans and animals. J. Antimicrob. Chemother. 2009, 64, 490–500. [Google Scholar] [CrossRef] [Green Version]
- Hinse, D.; Vollmer, T.; Erhard, M.; Welker, M.; Moore, E.; Kleesiek, K.; Dreier, J. Differentiation of species of the Streptococcus bovis/equinus-complex by MALDI-TOF mass spectrometry in comparison to sodA sequence analyses. Syst. Appl. Microbiol. 2011, 34, 52–57. [Google Scholar] [CrossRef]
- Wyckoff, H.; Chow, J.; Whitehead, T.; Cotta, M. Cloning, sequence, and expression of the L-(+) lactate dehydrogenase of Streptococcus bovis. Curr. Microbiol. 1997, 34, 367–373. [Google Scholar] [CrossRef]
- Yun, M.-J.; Yoon, S.; Lee, Y.J. Monitoring and characteristics of major mastitis pathogens from bulk tank milk in Korea. Animals 2020, 10, 1562. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, H.G.; Kwon, I.; Seo, J. Characterization of endolysin LyJH307 with antimicrobial activity against Streptococcus bovis. Animals 2020, 10, 963. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Cheng, L.; Kang, K.; Tian, G.; Al-Mamun, M.; Xue, B.; Wang, L.; Zou, H.; Gicheha, M.G.; Wang, Z. Effects of yeast and yeast cell wall polysaccharides supplementation on beef cattle growth performance, rumen microbial populations and lipopolysaccharides production. J. Integr. Agric. 2020, 19, 810–819. [Google Scholar] [CrossRef]
- Chuard, C.; Reller, L. Bile-esculin test for presumptive identification of enterococci and streptococci: Effects of bile concentration, inoculation technique, and incubation time. J. Clin. Microbiol. 1998, 36, 1135–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devriese, L.; Hommez, J.; Laevens, H.; Pot, B.; Vandamme, P.; Haesebrouck, F. Identification of aesculin-hydrolyzing streptococci, lactococci, aerococci and enterococci from subclinical intramammary infections in dairy cows. Vet. Microbiol. 1999, 70, 87–94. [Google Scholar] [CrossRef]
- Sasaki, E.; Osawa, R.; Nishitani, Y.; Whiley, R.A. Development of a diagnostic PCR assay targeting the Mn-dependent superoxide dismutase gene (sodA) for identification of Streptococcus gallolyticus. J. Clin. Microbiol. 2004, 42, 1360–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, T.; Fujiwara, T.; Kilian, M. Use of phylogenetic and phenotypic analyses to identify nonhemolytic streptococci isolated from bacteremic patients. J. Clin. Microbiol. 2005, 43, 6073–6085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teles, C.; Smith, A.; Ramage, G.; Lang, S. Identification of clinically relevant viridans group streptococci by phenotypic and genotypic analysis. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Aphale, D.; Kulkarni, A. Modifications and optimization of manual methods for polymerase chain reaction and 16S rRNA gene sequencing quality community DNA extraction from goat rumen digesta. Vet. World 2018, 11, 990. [Google Scholar] [CrossRef]
- Rosa, N.M.; Agnoletti, F.; Lollai, S.; Tola, S. Comparison of PCR-RFLP, API® 20 Strep and MALDI-TOF MS for identification of Streptococcus spp. collected from sheep and goat milk samples. Small Rumin. Res. 2019, 180, 35–40. [Google Scholar] [CrossRef]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Sys. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auch, A.F.; von Jan, M.; Klenk, H.-P.; Göker, M. Digital DNA-DNA hybridization for microbial species delineation by means of genome-to-genome sequence comparison. Stand. Genom. Sci. 2010, 2, 117–134. [Google Scholar] [CrossRef] [Green Version]
- Gherardi, G.; Palmieri, C.; Marini, E.; Pompilio, A.; Crocetta, V.; Di Bonaventura, G.; Creti, R.; Facinelli, B. Identification, antimicrobial resistance and molecular characterization of the human emerging pathogen Streptococcus gallolyticus subsp. pasteurianus. Diagn. Microbiol. Infec. Dis. 2016, 86, 329–335. [Google Scholar] [CrossRef]
- Streit, J.M.; Steenbergen, J.N.; Thorne, G.M.; Alder, J.; Jones, R.N. Daptomycin tested against 915 bloodstream isolates of viridans group streptococci (eight species) and Streptococcus bovis. J. Antimicrob. Chemother. 2005, 55, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Teng, L.-J.; Hsueh, P.-R.; Ho, S.-W.; Luh, K.-T. High prevalence of inducible erythromycin resistance among Streptococcus bovis isolates in Taiwan. Antimicrob. Agents Chemother. 2001, 45, 3362–3365. [Google Scholar] [CrossRef] [Green Version]
- Rato, M.G.; Bexiga, R.; Florindo, C.; Cavaco, L.M.; Vilela, C.L.; Santos-Sanches, I. Antimicrobial resistance and molecular epidemiology of streptococci from bovine mastitis. Vet. Microbiol. 2013, 161, 286–294. [Google Scholar] [CrossRef]
- Kaczorek, E.; Małaczewska, J.; Wójcik, R.; Rękawek, W.; Siwicki, A. Phenotypic and genotypic antimicrobial susceptibility pattern of Streptococcus spp. isolated from cases of clinical mastitis in dairy cattle in Poland. J. Dairy Sci. 2017, 100, 6442–6453. [Google Scholar] [CrossRef]
- Rodríguez-Avial, I.; Rodríguez-Avial, C.; Culebras, E.; Picazo, J.J. In vitro activity of telithromycin against viridans group streptococci and Streptococcus bovis isolated from blood: Antimicrobial susceptibility patterns in different groups of species. Antimicrob. Agents Chemother. 2005, 49, 820–823. [Google Scholar] [CrossRef] [Green Version]
- Haenni, M.; Lupo, A.; Madec, J.Y. Antimicrobial resistance in Streptococcus spp. Antimicrob. Resist. Bact. Livest. Companion Anim. 2018, 159–184. [Google Scholar] [CrossRef]
- Tang, Y.; Jiang, Q.; Tang, H.; Wang, Z.; Yin, Y.; Ren, F.; Kong, L.; Jiao, X.; Huang, J. Characterization and prevalence of Campylobacter spp. from broiler chicken rearing period to the slaughtering process in eastern China. Front. Vet. Sci. 2020, 7, 227. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yu, F.-Q.; Luo, L.-P.; He, J.-Z.; Hou, R.-G.; Zhang, H.-Q.; Li, S.-M.; Su, J.-L.; Han, B. Antibiotic resistance of Streptococcus agalactiae from cows with mastitis. Vet. J. 2012, 194, 423–424. [Google Scholar] [CrossRef] [PubMed]
- McDougall, S.; Hussein, H.; Petrovski, K. Antimicrobial resistance in Staphylococcus aureus, Streptococcus uberis and Streptococcus dysgalactiae from dairy cows with mastitis. N. Z. Vet. J. 2014, 62, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Ruegg, P.; Oliveira, L.; Jin, W.; Okwumabua, O. Phenotypic antimicrobial susceptibility and occurrence of selected resistance genes in gram-positive mastitis pathogens isolated from Wisconsin dairy cows. J. Dairy Sci. 2015, 98, 4521–4534. [Google Scholar] [CrossRef]
- Rice, L.B. Tn916 family conjugative transposons and dissemination of antimicrobial resistance determinants. Antimicrob. Agents Chemother. 1998, 42, 1871–1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dogan, B.; Schukken, Y.; Santisteban, C.; Boor, K.J. Distribution of serotypes and antimicrobial resistance genes among Streptococcus agalactiae isolates from bovine and human hosts. J. Clin. Microbiol. 2005, 43, 5899–5906. [Google Scholar] [CrossRef] [Green Version]
- Khaldi, T.E.M.; Kebouchi, M.; Soligot, C.; Gomri, M.A.; Kharroub, K.; Le Roux, Y.; Roux, E. Streptococcus macedonicus strains isolated from traditional fermented milks: Resistance to gastrointestinal environment and adhesion ability. Appl. Microbiol. Biotechnol. 2019, 103, 2759–2771. [Google Scholar] [CrossRef]
- Kmet, V.; Flint, H.; Wallace, R. Probiotics and manipulation of rumen development and function. Arch. Anim. Nutr. 1993, 44, 1–10. [Google Scholar] [CrossRef]
- Kumar, K.; Chaudhary, L.; Agarwal, N.; Kamra, D. Effect of supplementing Streptococcus gallolyticus strain TDGB 406 on rumen fermentation and microbial profile of goats fed on oak leaves. Anim. Nutr. Feed Technol. 2016, 16, 495–503. [Google Scholar] [CrossRef]
- Tarrah, A.; da Silva Duarte, V.; Pakroo, S.; Corich, V.; Giacomini, A. Genomic and phenotypic assessments of safety and probiotic properties of Streptococcus macedonicus strains of dairy origin. Food Res. Int. 2020, 130, 108931. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No. | Bacterial Strains | Host | Isolate Year | 16S rRNA Nos. * | sodA Gene Nos. * | Deposition No. ** |
|---|---|---|---|---|---|---|
| 1 | Streptococcus equinus CNU_5 | Bos taurus | 2014 | MN075416 | MN087729 | KCCM 90360 |
| 2 | Streptococcus equinus CNU_77-2 | Bos taurus | 2014 | MN075447 | MN087730 | KCCM 90361 |
| 3 | Streptococcus equinus CNU_77-3 | Bos taurus | 2014 | MN075448 | MN087731 | KCCM 90362 |
| 4 | Streptococcus equinus CNU_11 | Bos taurus | 2014 | MN075419 | MN087721 | KCCM 90363 |
| 5 | Streptococcus equinus CNU_77-16 | Bos taurus | 2014 | MN075450 | MN087722 | KCCM 90364 |
| 6 | Streptococcus equinus CNU_77-20 | Bos taurus | 2014 | MN075451 | MN087732 | KCCM 90365 |
| 7 | Streptococcus equinus CNU_77-23 | Bos taurus | 2014 | MN075454 | MN087733 | KCCM 90366 |
| 8 | Streptococcus equinus CNU_77-27 | Bos taurus | 2014 | MN075455 | MN087734 | KCCM 90367 |
| 9 | Streptococcus equinus CNU_GF | Bos taurus | 2018 | MN075408 | MN087723 | KCCM 90354 |
| 10 | Streptococcus equinus CNU_G1 | Bos taurus | 2019 | MN075409 | MN087724 | KCCM 90381 |
| 11 | Streptococcus equinus CNU_G2 | Bos taurus | 2019 | MN075410 | MN087725 | KCCM 90355 |
| 12 | Streptococcus equinus CNU_G3 | Bos taurus | 2019 | MN075411 | MN087726 | KCCM 90356 |
| 13 | Streptococcus equinus CNU_G4 | Bos taurus | 2019 | MN075412 | MN087727 | KCCM 90357 |
| 14 | Streptococcus equinus CNU_G5 | Bos taurus | 2019 | MN075413 | MN087728 | KCCM 90358 |
| 15 | Streptococcus equinus CNU_G6 | Bos taurus | 2019 | MN075414 | MN087720 | KCCM 90359 |
| 16 | Streptococcus equinus CNU_9 | Bos taurus coreanae | 2014 | MN075423 | MN087735 | KCCM 90368 |
| 17 | Streptococcus equinus CNU_77-8 | Bos taurus coreanae | 2014 | MN075456 | MN087736 | KCCM 90369 |
| 18 | Streptococcus equinus CNU_77-11 | Bos taurus coreanae | 2014 | MN075459 | MN087737 | KCCM 90370 |
| 19 | Streptococcus equinus CNU_77-12 | Bos taurus coreanae | 2014 | MN075460 | MN087738 | KCCM 90371 |
| 20 | Streptococcus equinus CNU_77-14 | Bos taurus coreanae | 2014 | MN075461 | MN087739 | KCCM 90372 |
| 21 | Streptococcus equinus CNU_77-29 | Bos taurus coreanae | 2014 | MN075462 | MN087740 | KCCM 90373 |
| 22 | Streptococcus equinus CNU_15 | Bos taurus coreanae | 2014 | MN075424 | MN087741 | KCCM 90374 |
| 23 | Streptococcus equinus CNU_20 | Bos taurus coreanae | 2014 | MN075427 | MN087742 | KCCM 90375 |
| 24 | Streptococcus equinus CNU_21 | Bos taurus coreanae | 2014 | MN075428 | MN087743 | KCCM 90376 |
| 25 | Streptococcus equinus CNU_77-35 | Bos taurus coreanae | 2014 | MN075465 | MN087744 | KCCM 90377 |
| 26 | Streptococcus equinus CNU_77-37 | Bos taurus coreanae | 2014 | MN075467 | MN087745 | KCCM 90378 |
| 27 | Streptococcus equinus CNU_77-40 | Bos taurus coreanae | 2014 | MN075468 | MN087746 | KCCM 90379 |
| 28 | Streptococcus equinus CNU_25 | Bos taurus coreanae | 2014 | MN075431 | MN087747 | KCCM 90380 |
| 29 | Streptococcus equinus CNU_27 | Bos taurus coreanae | 2014 | MN075433 | MN087748 | KCCM 90382 |
| 30 | Streptococcus equinus CNU_77-43 | Bos taurus coreanae | 2014 | MN075470 | MN087749 | KCCM 90383 |
| 31 | Streptococcus equinus CNU_77-47 | Bos taurus coreanae | 2014 | MN075473 | MN087750 | KCCM 90384 |
| 32 | Streptococcus equinus CNU_77-50 | Bos taurus coreanae | 2014 | MN075475 | MN087751 | KCCM 90385 |
| 33 | Streptococcus equinus CNU_29 | Capra aegagrus hircus | 2014 | MN075435 | MN087752 | KCCM 90386 |
| 34 | Streptococcus equinus CNU_30 | Capra aegagrus hircus | 2014 | MN075436 | MN087753 | KCCM 90387 |
| 35 | Streptococcus equinus CNU_32 | Capra aegagrus hircus | 2014 | MN075438 | MN087754 | KCCM 90388 |
| 36 | Streptococcus equinus CNU_77-51 | Capra aegagrus hircus | 2014 | MN075476 | MN087755 | KCCM 90389 |
| 37 | Streptococcus equinus CNU_77-55 | Capra aegagrus hircus | 2014 | MN075478 | MN087756 | KCCM 90390 |
| 38 | Streptococcus equinus CNU_77-56 | Capra aegagrus hircus | 2014 | MN075479 | MN087757 | KCCM 90391 |
| 39 | Streptococcus equinus CNU_77-57 | Capra aegagrus hircus | 2014 | MN075480 | MN087758 | KCCM 90392 |
| 40 | Streptococcus equinus CNU_77-60 | Capra aegagrus hircus | 2014 | MN075481 | MN087760 | KCCM 90394 |
| 41 | Streptococcus equinus CNU_77-68 | Capra aegagrus hircus | 2014 | MN075488 | MN087764 | KCCM 90398 |
| 42 | Streptococcus equinus CNU_41 | Capra aegagrus hircus | 2014 | MN075445 | MN087765 | KCCM 90399 |
| 43 | Streptococcus equinus CNU_42 | Capra aegagrus hircus | 2014 | MN075446 | MN087766 | KCCM 90400 |
| 44 | Streptococcus equinus CNU_77-72 | Capra aegagrus hircus | 2014 | MN075490 | MN087767 | KCCM 90401 |
| 45 | Streptococcus equinus CNU_77-77 | Capra aegagrus hircus | 2014 | MN075492 | MN087769 | KCCM 90403 |
| 46 | Streptococcus equinus CNU_77-78 | Capra aegagrus hircus | 2014 | MN075493 | MN087770 | KCCM 90404 |
| 47 | Streptococcus lutetiensis CNU_33 | Capra aegagrus hircus | 2014 | MN075439 | MN087759 | KCCM 90393 |
| 48 | Streptococcus lutetiensis CNU_77-61 | Capra aegagrus hircus | 2014 | MN075482 | MN087761 | KCCM 90395 |
| 49 | Streptococcus lutetiensis CNU_77-62 | Capra aegagrus hircus | 2014 | MN075483 | MN087762 | KCCM 90396 |
| 50 | Streptococcus lutetiensis CNU_77-64 | Capra aegagrus hircus | 2014 | MN075485 | MN087763 | KCCM 90397 |
| 51 | Streptococcus lutetiensis CNU_77-76 | Capra aegagrus hircus | 2014 | MN075491 | MN087768 | KCCM 90402 |
| Bacterial Strain | Tetracycline | Lincosamides | MLSb ** | Ldh *** | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| tet(M) | tet(O) | tet(Q) | tet(S) | Tn916-like transposase | lnu(C) | erm(A) | erm(C) | erm(B) | mef(A) | ||
| Streptococcus equinus | |||||||||||
| CNU_5 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-2 | + (MT949160) | - | - | - | + (MT949172) | - | - | - | - | - | + |
| CNU_77-3 | + (MT949161) | - | - | - | + (MT949173) | - | - | - | - | - | + |
| CNU_11 | + (MT949162) | - | - | - | + (MT949174) | - | - | - | - | - | + |
| CNU_77-16 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-20 | + (MT949163) | - | - | - | + (MT949175) | - | - | - | - | - | + |
| CNU_77-23 | + (MT949164) | - | - | - | + (MT949176) | + (MT949150) | - | - | - | - | + |
| CNU_77-27 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_GF | + (MT949165) | - | - | - | + (MT949177) | + (MT949151) | - | - | - | - | + |
| CNU_G1 | + (MT949166) | - | - | - | + (MT949178) | + (MT949152) | - | - | - | - | + |
| CNU_G2 | + (MT949167) | - | - | - | + (MT949179) | - | - | - | - | - | + |
| CNU_G3 | - | - | - | - | - | + (MT949153) | - | - | - | - | + |
| CNU_G4 | - | - | - | - | - | + (MT949154) | - | - | - | - | + |
| CNU_G5 | + (MT949168) | - | - | - | + (MT949180) | - | - | - | - | - | + |
| CNU_G6 | + (MT949169) | - | - | - | + (MT949181) | - | - | - | - | - | + |
| CNU_9 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-8 | - | - | - | - | - | + (MT949155) | - | - | - | - | + |
| CNU_77-11 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-12 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-14 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-29 | + (MT949170) | - | - | - | + (MT949182) | - | - | - | - | - | + |
| CNU_15 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_20 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_21 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-35 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-37 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-40 | + (MT949171) | - | - | - | + (MT949183) | - | - | - | - | - | + |
| CNU_25 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_27 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-43 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-47 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-50 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_29 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_30 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_32 | - | - | - | - | - | + (MT949156) | - | - | - | - | + |
| CNU_77-51 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-55 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-56 | - | - | - | - | - | + (MT949157) | - | - | - | - | + |
| CNU_77-57 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-60 | - | - | - | - | - | + (MT949158) | - | - | - | - | + |
| CNU_77-68 | - | - | - | - | - | + (MT949159) | - | - | - | - | + |
| CNU_41 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_42 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-72 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-77 | - | - | - | - | - | - | - | - | - | - | + |
| CNU_77-78 | - | - | - | - | - | - | - | - | - | - | + |
| Streptococcus lutetiensis | |||||||||||
| CNU_33 | - | - | - | - | + (MT949184) | - | - | - | - | - | + |
| CNU_77-61 | - | - | - | - | + (MT949185) | - | - | - | - | - | + |
| CNU_77-62 | - | - | - | - | + (MT949186) | - | - | - | - | - | + |
| CNU_77-64 | - | - | - | - | + (MT949187) | - | - | - | - | - | + |
| CNU_77-76 | - | - | - | - | + (MT949188) | - | - | - | - | - | + |
| Strains | Antimicrobial Agents [μg (Disks) or mg/L (MIC)] | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Am * | Carb | Cep | Fq | Lin * | Ma * | Penicillins | Phe | Tet * | Gly | Oxa | ||
| CN (0.016–256) | IPM (10) | KF (30) | LEV (5) | CD (0.016–256) | E (0.016–256) | OX (1) | P (10) | C (30) | TE (0.016–256) | VA (30) | LZD (30) | |
| Streptococcus equinus | ||||||||||||
| CNU_5 | 3 | 0.094 | 0.19 | 3 | ||||||||
| CNU_77-2 | 2 | 0.064 | 0.047 | 12 | ||||||||
| CNU_77-3 | 1.5 | 0.047 | 0.50 | 16 | ||||||||
| CNU_11 | 5 | 0.064 | 0.19 | 4 | ||||||||
| CNU_77-16 | 12 | 0.064 | 0.19 | 4 | ||||||||
| CNU_77-20 | 4 | 0.064 | 0.50 | 32 | ||||||||
| CNU_77-23 | 16 | 1 | 0.25 | 32 | ||||||||
| CNU_77-27 | 12 | 0.094 | 0.25 | 3 | ||||||||
| CNU_GF | 12 | 0.125 | 0.19 | 6 | ||||||||
| CNU_G1 | 12 | 0.125 | 0.094 | 32 | ||||||||
| CNU_G2 | 16 | 0.125 | 0.19 | 12 | ||||||||
| CNU_G3 | 6 | 0.5 | 0.19 | 6 | ||||||||
| CNU_G4 | 16 | 0.047 | 0.19 | 0.75 | ||||||||
| CNU_G5 | 8 | 0.064 | 0.064 | 3 | ||||||||
| CNU_G6 | 4 | 0.094 | 0.094 | 32 | ||||||||
| CNU_9 | 24 | 2.5 | 0.50 | 3 | ||||||||
| CNU_77-8 | 4 | 0.25 | 0.125 | 4 | ||||||||
| CNU_77-11 | 24 | 0.094 | 0.38 | 3 | ||||||||
| CNU_77-12 | 8 | 0.094 | 0.38 | 4 | ||||||||
| CNU_77-14 | 8 | 0.19 | 0.38 | 4 | ||||||||
| CNU_77-29 | 3 | 0.064 | 0.094 | 0.50 | ||||||||
| CNU_15 | 12 | 0.094 | 0.50 | 3 | ||||||||
| CNU_20 | 8 | 0.125 | 0.19 | 2 | ||||||||
| CNU_21 | 8 | 0.19 | 0.50 | 3 | ||||||||
| CNU_77-35 | 32 | 0.19 | 0.50 | 3 | ||||||||
| CNU_77-37 | 12 | 0.25 | 0.19 | 4 | ||||||||
| CNU_77-40 | 4 | 0.094 | 0.19 | 48 | ||||||||
| CNU_25 | 8 | 0.125 | 0.75 | 2 | ||||||||
| CNU_27 | 12 | 0.125 | 0.50 | 3 | ||||||||
| CNU_77-43 | 24 | 0.38 | 6 | 6 | ||||||||
| CNU_77-47 | 24 | 0.094 | 0.50 | 3 | ||||||||
| CNU_77-50 | 12 | 0.125 | 0.50 | 3 | ||||||||
| CNU_29 | 6 | 0.094 | 0.19 | 2 | ||||||||
| CNU_30 | 6 | 0.125 | 0.25 | 0.5 | ||||||||
| CNU_32 | 6 | 0.125 | 0.19 | 0.75 | ||||||||
| CNU_77-51 | 6 | 0.094 | 0.19 | 0.5 | ||||||||
| CNU_77-55 | 8 | 0.094 | 0.25 | 0.5 | ||||||||
| CNU_77-56 | 12 | 0.19 | 0.19 | 0.5 | ||||||||
| CNU_77-57 | 16 | 0.125 | 0.0125 | 3 | ||||||||
| CNU_77-60 | 24 | 0.125 | 0.25 | 3 | ||||||||
| CNU_77-68 | 48 | 0.125 | 0.19 | 1 | ||||||||
| CNU_41 | 6 | 0.047 | 0.25 | 3 | ||||||||
| CNU_42 | 8 | 0.125 | 0.125 | 1 | ||||||||
| CNU_77-72 | 8 | 0.094 | 0.125 | 3 | ||||||||
| CNU_77-77 | 6 | 0.094 | 0.032 | 0.50 | ||||||||
| CNU_77-78 | 8 | 0.064 | 0.125 | 0.38 | ||||||||
| Streptococcus lutetiensis | ||||||||||||
| CNU_33 | 12 | 0.125 | 0.032 | 1 | ||||||||
| CNU_77-61 | 8 | 0.19 | 0.25 | 3 | ||||||||
| CNU_77-62 | 12 | 0.064 | 0.19 | 3 | ||||||||
| CNU_77-64 | 8 | 0.094 | 0.25 | 2 | ||||||||
| CNU_77-76 | 5 | 0.19 | 0.25 | 3 | ||||||||
| Susceptible | 15.7% (8/51) | 100% (51/51) | 90.2% (46/51) | 0% (0/51) | 94.1% (48/51) | 72.5% (37/51) | 98.0% (50/51) | 9.8% (5/51) | 3.9% (2/51) | 29.4% (15/51) | 15.7% (8/51) | 5.9% (3/51) |
| Intermediate | 62.7% (32/51) | 0% (0/51) | 9.8% (5/51) | 76.5% (39/51) | 5.9% (3/51) | 25.5% (13/51) | 2.0% (1/51) | 90.2% (46/51) | 47.1% (24/51) | 54.9% (28/51) | 84.3% ** (43/51) | 94.1% ** (48/51) |
| Resistant | 21.6% (11/51) | 0% (0/51) | 0% (0/51) | 23.5% (12/51) | 0% (0/51) | 2.0% (1/51) | 0% (0/51) | 0% (0/51) | 49.0% (25/51) | 15.7% (8/51) | ||
| Feature | Strains | ||||
|---|---|---|---|---|---|
| CNU_77-23 (CP046628) | CNU_77-61 (CP046875) | CNU_G2 (CP046919) | CNU_G3 (CP046624) | CNU_G6 (CP046629) | |
| Size (bp) | 1,911,874 | 1,917,833 | 1,960,491 | 1,864,202 | 1,910,720 |
| G + C content (%) | 37.4 | 37.8 | 37.9 | 37.8 | 37.4 |
| Conitgs | 1 | 1 | 2 | 2 | 2 |
| plasmids | - | - | 1 | 1 | 1 |
| Total genes | 1883 | 1910 | 2056 | 1885 | 1948 |
| tRNAs | 70 | 70 | 70 | 69 | 70 |
| rRNAs | 21 | 21 | 21 | 21 | 21 |
| ncRNAs | 4 | 4 | 4 | 4 | 4 |
| Protein-coding genes | 1771 | 1778 | 1926 | 1759 | 1834 |
| Pseudogenes | 17 | 37 | 35 | 32 | 19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.Y.; Lee, M.; Lim, S.R.; Kwon, H.; Lee, Y.S.; Kim, J.H.; Seo, S. Diversity and Antimicrobial Resistance in the Streptococcus bovis/Streptococcus equinus Complex (SBSEC) Isolated from Korean Domestic Ruminants. Microorganisms 2021, 9, 98. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010098
Park SY, Lee M, Lim SR, Kwon H, Lee YS, Kim JH, Seo S. Diversity and Antimicrobial Resistance in the Streptococcus bovis/Streptococcus equinus Complex (SBSEC) Isolated from Korean Domestic Ruminants. Microorganisms. 2021; 9(1):98. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010098
Chicago/Turabian StylePark, Seon Young, Mingyung Lee, Se Ra Lim, Hyemin Kwon, Ye Seul Lee, Ji Hyung Kim, and Seongwon Seo. 2021. "Diversity and Antimicrobial Resistance in the Streptococcus bovis/Streptococcus equinus Complex (SBSEC) Isolated from Korean Domestic Ruminants" Microorganisms 9, no. 1: 98. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010098