
Immunity Profiling of COVID-19 Infection, Dynamic Variations of Lymphocyte Subsets, a Comparative Analysis on Four Different Groups


 , , , ,
, , , ,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. RT-PCR and Nucleic Acid Detection
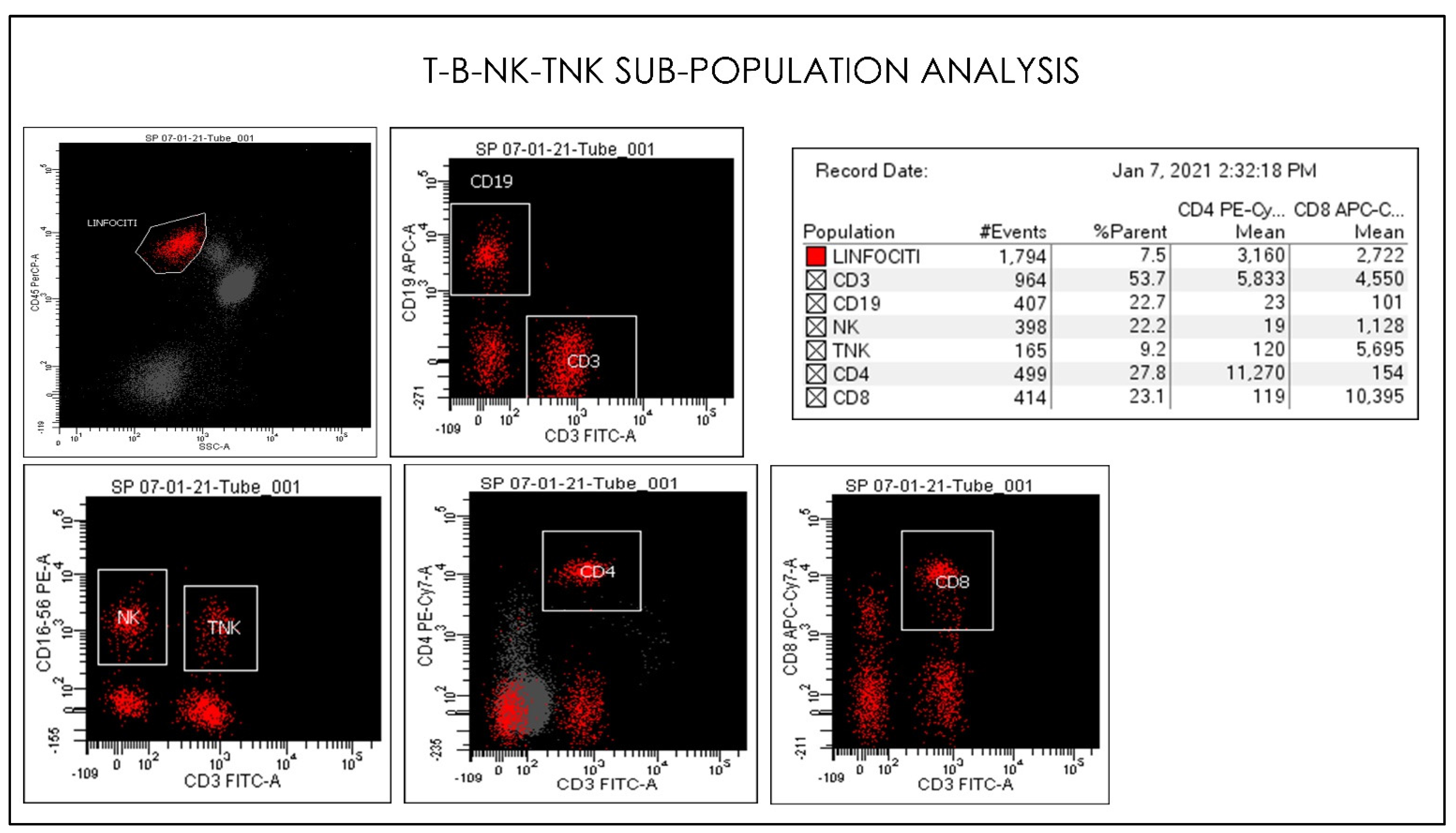
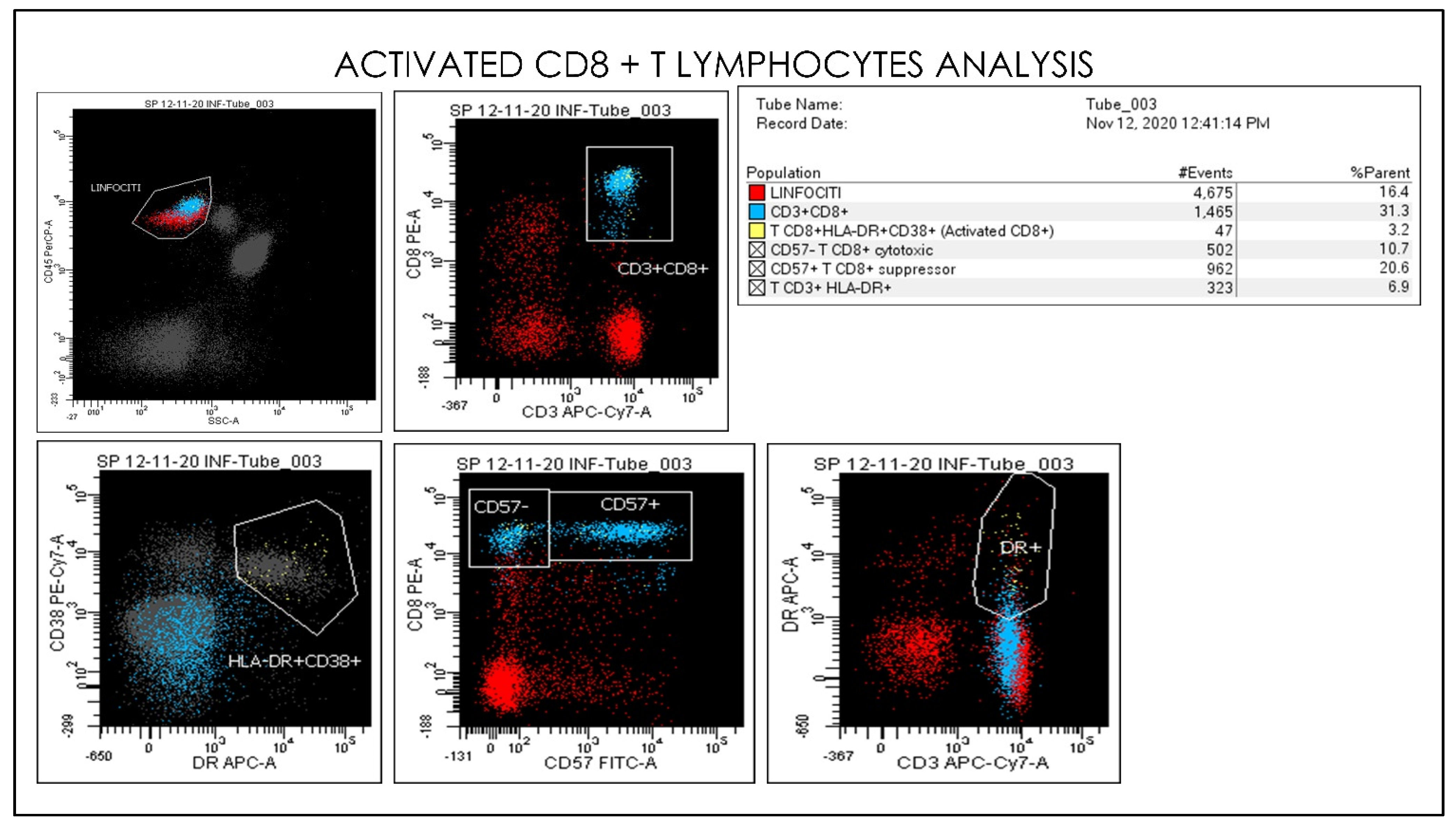
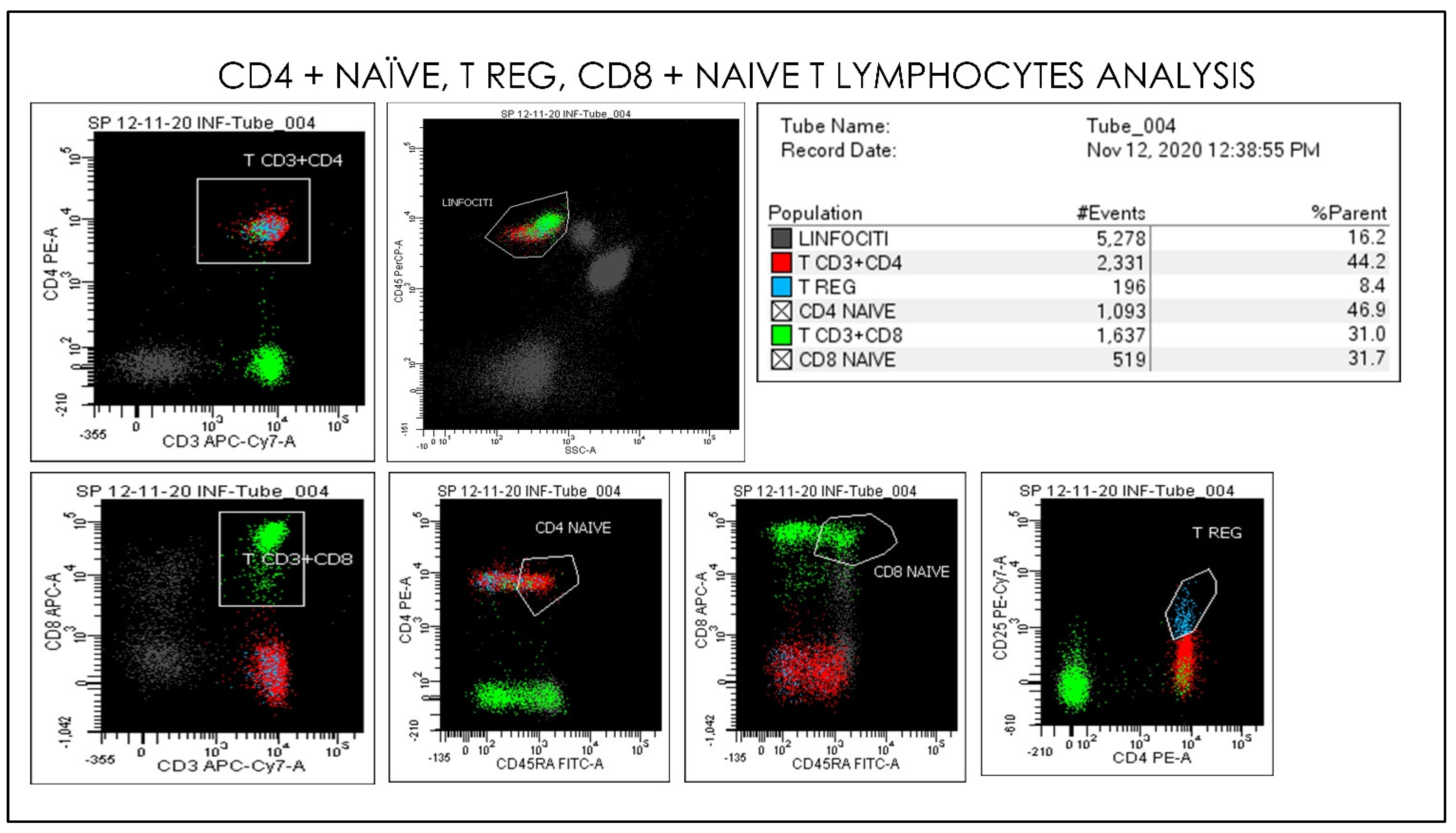
2.3. Flow Cytometry Hematology Parameters
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pham, V.H.; Gargiulo Isacco, C.; Nguyen, K.C.D.; Le, S.H.; Tran, D.K.; Nguyen, Q.V.; Pham, H.T.; Aityan, S.; Pham, S.T.; Cantore, S.; et al. Rapid and Sensitive Diagnostic Procedure for Multiple Detection of Pandemic Coronaviridae Family Members SARS-CoV-2, SARS-CoV, MERS-CoV and HCoV: A Translational Research and Cooperation between the Phan Chau Trinh University in Vietnam and University of Bari “Aldo Moro” in Italy. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7173–7191. [Google Scholar] [CrossRef]
- Wang, F.; Nie, J.; Wang, H.; Zhao, Q.; Xiong, Y.; Deng, L.; Song, S.; Ma, Z.; Mo, P.; Zhang, Y. Characteristics of Peripheral Lymphocyte Subset Alteration in COVID-19 Pneumonia. J. Infect. Dis. 2020, 221, 1762–1769. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.M.; Lau, E.H.Y.; Wong, J.Y.; et al. Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus-Infected Pneumonia. N. Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef]
- Kalicińska, E.; Szymczak, D.; Zińczuk, A.; Adamik, B.; Smiechowicz, J.; Skalec, T.; Nowicka-Suszko, D.; Biernat, M.; Bogucka-Fedorczuk, A.; Rybka, J.; et al. Immunosuppression as a Hallmark of Critical COVID-19: Prospective Study. Cells 2021, 10, 1293. [Google Scholar] [CrossRef]
- Gallais, F.; Velay, A.; Wendling, M.-J.; Nazon, C.; Partisani, M.; Sibilia, J.; Candon, S.; Fafi-Kremer, S. Intrafamilial Exposure to SARS-CoV-2 Induces Cellular Immune Response without Seroconversion. Emerg. Infect. Dis. 2020, 27, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, F.; Inchingolo, F.; Scarano, A. The Impact Of The Novel Covid-19 On The Scientific Production Spread: A Five-Month Bibliometric Report Of The Worldwide Research Community. Acta Medica. Mediterr. 2020, 36, 1–4. [Google Scholar]
- Cossarizza, A.; Ortolani, C.; Paganelli, R.; Barbieri, D.; Monti, D.; Sansoni, P.; Fagiolo, U.; Castellani, G.; Bersani, F.; Londei, M.; et al. CD45 isoforms expression on CD4+ and CD8+ T cells throughout life, from newborns to centenarians: Implications for T cell memory. Mech. Ageing Dev. 1996, 86, 173–195. [Google Scholar] [CrossRef]
- Chaplin, D.D. Overview of the Immune Response. J. Allergy Clin. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501. [Google Scholar] [CrossRef]
- Wherry, E.J.; Kurachi, M. Molecular and Cellular Insights into T Cell Exhaustion. Nat. Rev. Immunol. 2015, 15, 486–499. [Google Scholar] [CrossRef]
- Jiao, Y.; Fu, J.; Xing, S.; Fu, B.; Zhang, Z.; Shi, M.; Wang, X.; Zhang, J.; Jin, L.; Kang, F.; et al. The Decrease of Regulatory T Cells Correlates with Excessive Activation and Apoptosis of CD8+ T Cells in HIV-1-Infected Typical Progressors, but Not in Long-Term Non-Progressors. Immunology 2009, 128, e366–e375. [Google Scholar] [CrossRef]
- Tumedei, M.; Piattelli, A.; Degidi, M.; Mangano, C.; Iezzi, G. A 30-Year (1988–2018) Retrospective Microscopical Evaluation of Dental Implants Retrieved for Different Causes: A Narrative Review. Int J. Periodontics Restor. Dent. 2020, 40, e211–e227. [Google Scholar] [CrossRef]
- Fujiwara, S.; Kato, S.; Bengazi, F.; Urbizo Velez, J.; Tumedei, M.; Kotsu, M.; Botticelli, D. Healing at Implants Installed in Osteotomies Prepared Either with a Piezoelectric Device or Drills: An Experimental Study in Dogs. Oral Maxillofac. Surg. 2021, 25, 65–73. [Google Scholar] [CrossRef]
- Kotsu, M.; Urbizo Velez, J.; Bengazi, F.; Tumedei, M.; Fujiwara, S.; Kato, S.; Botticelli, D. Healing at Implants Installed from ~ 70- to < 10-Ncm Insertion Torques: An Experimental Study in Dogs. Oral Maxillofac. Surg. 2021, 25, 55–64. [Google Scholar] [CrossRef]
- Balzanelli, M.G.; Distratis, P.; Aityan, K.S.; Amatulli, F.; Catucci, O.; Cefalo, A.; De Michele, A.; Dipalma, G.; Inchingolo, F.; Lazzaro, R.; et al. An Alternative “Trojan Horse” Hypothesis for COVID-19: Immune Deficiency of IL-10 and SARS-CoV-2 Biology. Endocr. Metab. Immune. Disord. Drug Targets 2021. [Google Scholar] [CrossRef]
- Cox, R.J.; Brokstad, K.A. Not just antibodies: B cells and T cells mediate immunity to COVID-19. Nat. Rev. Immunol. 2020, 20, 581–582. [Google Scholar] [CrossRef] [PubMed]
- Vermesan, D.; Prejbeanu, R.; Poenaru, D.V.; Petrescu, H.; Apostol, E.; Inchingolo, F.; Dipalma, G.; Abbinante, A.; Caprio, M.; Potenza, M.A.; et al. Do Intramedullary Implants Improve Survival in Elderly Patients with Trochanteric Fractures? A Retrospective Study. Clin. Ter. 2015, 166, e140–e145. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.X.; Tang, X.J.; Shi, Q.L. Clinical and immunological assessment of asymptomatic SARS-CoV-2 infections. Nat. Med. 2020, 26, 1200–1204. [Google Scholar] [CrossRef]
- Campanella, V.; Syed, J.; Santacroce, L.; Saini, R.; Ballini, A.; Inchingolo, F. Oral Probiotics Influence Oral and Respiratory Tract Infections in Pediatric Population: A Randomized Double-Blinded Placebo-Controlled Pilot Study. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8034–8041. [Google Scholar] [CrossRef]
- Jiang, M.; Guo, Y.; Luo, Q.; Huang, Z.; Zhao, R.; Liu, S.; Le, A.; Li, J.; Wan, L. T-Cell Subset Counts in Peripheral Blood Can Be Used as Discriminatory Biomarkers for Diagnosis and Severity Prediction of Coronavirus Disease 2019. J. Infect. Dis. 2020, 222, 198–202. [Google Scholar] [CrossRef]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of Immune Response in Patients With Coronavirus 2019 (COVID-19) in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Charitos, I.A.; Ballini, A.; Bottalico, L.; Cantore, S.; Passarelli, P.C.; Inchingolo, F.; D’Addona, A.; Santacroce, L. Special features of SARS-CoV-2 in daily practice. World J. Clin. Cases 2020, 8, 3920–3933. [Google Scholar] [CrossRef] [PubMed]
- Charitos, I.A.; Del Prete, R.; Inchingolo, F.; Mosca, A.; Carretta, D.; Ballini, A.; Santacroce, L. What we have learned for the future about COVID-19 and healthcare management of it? Acta. Biomed. 2020, 91, e2020126. [Google Scholar] [PubMed]
- Norheim, O.F. Protecting the population with immune individuals. Nat. Med. 2020, 26, 823–824. [Google Scholar] [CrossRef]
- Klimpel, G.R. Immune Defenses. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Cao, W.C.; Liu, W.; Zhang, P.H.; Zhang, F.; Richardus, J.H. Disappearance of antibodies to SARS-associated coronavirus after recovery. N. Engl. J. Med. 2007, 357, 1162–1163. [Google Scholar] [CrossRef]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.I.; Haupt, S.; Frazier, A.; et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef]
- Suthar, M.S.; Zimmerman, M.G.; Kauffman, R.C.; Mantus, G.; Linderman, S.L.; Hudson, W.H.; Vanderheiden, A.; Nyhoff, L.; Davis, C.W.; Adekunle, O.; et al. Rapid Generation of Neutralizing Antibody Responses in COVID-19 Patients. Cell Rep. Med. 2020, 1, 100040. [Google Scholar] [CrossRef]
- Pušnik, J.; Richter, E.; Schulte, B.; Dolscheid-Pommerich, R.; Bode, C.; Putensen, C.; Hartmann, G.; Alter, G.; Streeck, H. Memory B cells targeting SARS-CoV-2 spike protein and their dependence on CD4+ T cell help. Cell Rep. 2021, 35, 109320. [Google Scholar] [CrossRef]
- Okba, N.M.A.; Müller, M.A.; Li, W.; Wang, C.; GeurtsvanKessel, C.H.; Corman, V.M.; Lamers, M.M.; Sikkema, R.S.; de Bruin, E.; Chandler, F.D.; et al. Severe Acute Respiratory Syndrome Coronavirus 2−Specific Antibody Responses in Coronavirus Disease Patients. Emerg. Infect. Dis. 2020, 26, 1478–1488. [Google Scholar] [CrossRef]
- Oguntuyo, K.Y.; Stevens, C.S.; Hung, C.-T.; Ikegame, S.; Acklin, J.A.; Kowdle, S.S.; Carmichael, J.C.; Chiu, H.-P.; Azarm, K.D.; Haas, G.D.; et al. Quantifying Absolute Neutralization Titers against SARS-CoV-2 by a Standardized Virus Neutralization Assay Allows for Cross-Cohort Comparisons of COVID-19 Sera. mBio 2021, 12, e02492-20. [Google Scholar] [CrossRef]
- Balzanelli, M.G.; Distratis, P.; Lazzaro, R.; Cefalo, A.; D’angela, G.; Catucci, O.; Palazzo, D.; Amatulli, F.; Tomassone, D.; Pham, H.V.; et al. Would the End of COVID-19 Infection as a Chronic Disease? J. Stem. Cell Res. Dev. Ther. 2020, 6, 057. [Google Scholar]
- Balzanelli, G.M.; Distratis, P.; Aityan, S.K.; Amatulli, F.; Catucci, O.; Cefalo, A.; Dipalma, G.; Inchingolo, F.; Lazzaro, R.; Nguyen, K.C.D.; et al. COVID-19 and COVID-like Patients: A Brief Analysis and Findings of Two Deceased Cases. Open Access Maced. J. Med. Sci. 2020, 8, 490–495. [Google Scholar] [CrossRef]
- Balzanelli, M.G.; Distratis, P.; Nguyen, K.C.D.; Lazzaro, R.; Catucci, O.; Cefalo, A.; Palazzo, D.; D’Angela, G.; Amatulli, F.; Gargiulo-Isacco, C.; et al. Could immunity boosting therapy help Covid-19 patients? Biomed. J. Sci. Tech. Res. 2020, 30, 23149–23150. [Google Scholar] [CrossRef]
- Balzanelli, M.G.; Distratis, P.; Catucci, O.; Cefalo, A.; Lazzaro, R.; Inchingolo, F.; Tomassone, D.; Aityan, S.K.; Ballini, A.; Nguyen, K.C.D.; et al. Mesenchymal Stem Cells: The Secret Children’s Weapons against the SARS-CoV-2 Lethal Infection. Appl. Sci. 2021, 11, 1696. [Google Scholar] [CrossRef]
- Ackermann, M.; Anders, H.J.; Bilyy, R.; Bowlin, G.L.; Daniel, C.; De Lorenzo, R.; Egeblad, M.; Henneck, T.; Hidalgo, A.; Hoffmann, M.; et al. Patients with COVID-19: In the dark-NETs of neutrophils. Cell Death Differ. 2021. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.R.; Ma, A.C.; Tavener, S.A.; McDonald, B.; Goodarzi, Z.; Kelly, M.M.; Patel, K.D.; Chakrabarti, S.; McAvoy, E.; Sinclair, G.D.; et al. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat. Med. 2007, 13, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Veras, F.P.; Pontelli, M.C.; Silva, C.M.; Toller-Kawahisa, J.E.; de Lima, M.; Nascimento, D.C.; Schneider, A.H.; Caetité, D.; Tavares, L.A.; Paiva, I.M.; et al. SARS-CoV-2-triggered neutrophil extracellular traps mediate COVID-19 pathology. J. Exp. Med. 2020, 217, e20201129. [Google Scholar] [CrossRef]
- Balzanelli, G.M.; Distratis, P.; Aityan, K.S.; Amatulli, F.; Catucci, O.; Cefalo, A.; D’Angela, G.; Dipalma, G.; Inchingolo, F.; Lazzaro, R.; et al. Clinical features in predicting COVID-19. Biome. J. Sci Tech. Res. 2020, 29, 22921–22926. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef]
- Pan, Y.; Jiang, X.; Yang, L.; Chen, L.; Zeng, X.; Liu, G.; Tang, Y.; Qian, C.; Wang, X.; Cheng, F.; et al. SARS-CoV-2-specific immune response in COVID-19 convalescent individuals. Sig. Transduct. Target. Ther. 2021, 6, 256. [Google Scholar] [CrossRef]
- Ahn, J.J.; Abu-Rub, M.; Miller, R.H. B Cells in Neuroinflammation: New Perspectives and Mechanistic Insights. Cells 2021, 10, 1605. [Google Scholar] [CrossRef] [PubMed]
- Den Haan, J.M.M.; Arens, R.; van Zelm, M.C. The Activation of the Adaptive Immune System: Cross-Talk between Antigen-Presenting Cells, T Cells and B Cells. Immunol. Lett. 2014, 162, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Fan, Y.; Lai, Y.; Han, T.; Li, Z.; Zhou, P.; Pan, P.; Wang, W.; Hu, D.; Liu, X.; et al. Coronavirus Infections and Immune Responses. J. Med. Virol. 2020, 92, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Sarvaria, A.; Madrigal, J.A.; Saudemont, A. B Cell Regulation in Cancer and Anti-Tumor Immunity. Cell Mol. Immunol. 2017, 14, 662–674. [Google Scholar] [CrossRef] [Green Version]
- Atri, C.; Guerfali, F.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. IJMS 2018, 19, 1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edridge, A.W.D.; Kaczorowska, J.; Hoste, A.C.R.; Bakker, M.; Klein, M.; Loens, K.; Jebbink, M.F.; Matser, A.; Kinsella, C.M.; Rueda, P. Seasonal coronavirus protective immunity is short-lasting. Nat. Med. 2020, 26, 1691–1693. [Google Scholar] [CrossRef]
- Peghin, M.; Bouza, E.; Fabris, M.; De Martino, M.; Palese, A.; Bontempo, G.; Graziano, E.; Gerussi, V.; Bressan, V.; Sartor, A.; et al. Low risk of reinfections and relation with serological response after recovery from the first wave of COVID-19. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 11, 1–8. [Google Scholar] [CrossRef]
- Hanrath, A.T.; Payne, B.A.I.; Duncan, C.J.A. Prior SARS-CoV-2 infection is associated with protection against symptomatic reinfection. J. Infect. 2021, 82, e29–e30. [Google Scholar] [CrossRef]
- Berger, A. Th1 and Th2 Responses: What Are They? BMJ 2000, 321, 424. [Google Scholar] [CrossRef] [Green Version]
- Janeway, C.A., Jr.; Travers, P.; Walport, M. Immunological memory. In Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Dohan Ehrenfest, D.M.; Del Corso, M.; Inchingolo, F.; Charrier, J.-B. Selecting a Relevant in Vitro Cell Model for Testing and Comparing the Effects of a Choukroun’s Platelet-Rich Fibrin (PRF) Membrane and a Platelet-Rich Plasma (PRP) Gel: Tricks and Traps. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2010, 110, 409–413. [Google Scholar] [CrossRef]
- Anichini, G.; Terrosi, C.; Gandolfo, C.; Gori Savellini, G.; Fabrizi, S.; Miceli, G.B.; Cusi, M.G. SARS-CoV-2 antibody response in persons with past natural infection. N. Engl. J. Med. 2021, 385, 90–92. [Google Scholar] [CrossRef]
- Prendergast, A.J.; Klenerman, P.; Goulder, P.J. The impact of differential antiviral immunity in children and adults. Nat. Rev. Immunol. 2012, 12, 636–648. [Google Scholar] [CrossRef] [Green Version]
- Boonnak, K.; Vogel, L.; Feldmann, F.; Feldmann, H.; Legge, K.L.; Subbarao, K. Lymphopenia Associated with Highly Virulent H5N1 Virus Infection Due to Plasmacytoid Dendritic Cell–Mediated Apoptosis of T Cells. J. Immunol. 2014, 192, 5906–5912. [Google Scholar] [CrossRef]
- Maeurer, M.J.; Martin, D.; Walter, W.; Liu, K.; Zitvogel, L.; Halusczcak, K.; Rabinowich, H.; Duquesnoy, R.; Storkus, W.; Lotze, M.T. Human Intestinal Vdelta1+ Lymphocytes Recognize Tumor Cells of Epithelial Origin. J. Exp. Med. 1996, 183, 1681–1696. [Google Scholar] [CrossRef] [PubMed]
- Balzanelli, M.; Distratis, P.; Catucci, O.; Amatulli, F.; Cefalo, A.; Lazzaro, R.; Aytian, S.K.; Dalagni, G.; Nico, A.; De Michele, A.; et al. Clinical and Diagnostic Findings in COVID-19 Patients: An Original Research from SG Moscati Hospital in Taranto Italy. J. Biol. Regul. Homeost Agents 2021, 35, 171–183. [Google Scholar] [CrossRef]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic Characterisation and Epidemiology of 2019 Novel Coronavirus: Implications for Virus Origins and Receptor Binding. The Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Mathew, D.; Giles, J.R.; Baxter, A.E.; Oldridge, D.A.; Greenplate, A.R.; Wu, J.E.; Alanio, C.; Kuri-Cervantes, L.; Pampena, M.B.; D’Andrea, K.; et al. Deep Immune Profiling of COVID-19 Patients Reveals Distinct Immunotypes with Therapeutic Implications. Science 2020, 369, eabc8511. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, N.M.; Hasegawa, M.; Ishijima, M.; Nakhaei, K.; Okubo, T.; Taniyama, T.; Ghassemi, A.; Tahsili, T.; Park, W.; Hirota, M.; et al. Biological and Osseointegration Capabilities of Hierarchically (Meso-/Micro-/Nano-Scale) Roughened Zirconia. Int J. Nanomed. 2018, 13, 3381–3395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Han, J.; Cheng, X.; Yu, L.; Zhang, L.; Wang, W.; Ni, L.; Wei, C.; Huang, Y.; Cheng, Z. Reduced Numbers of T Cells and B Cells Correlates with Persistent SARS-CoV-2 Presence in Non-Severe COVID-19 Patients. Sci. Rep. 2020, 10, 17718. [Google Scholar] [CrossRef]
- Bordea, I.R.; Xhajanka, E.; Candrea, S.; Bran, S.; Onișor, F.; Inchingolo, A.D.; Malcangi, G.; Pham, V.H.; Inchingolo, A.M.; Scarano, A.; et al. Coronavirus (SARS-CoV-2) Pandemic: Future Challenges for Dental Practitioners. Microorganisms 2020, 8, 1704. [Google Scholar] [CrossRef]
- Maucourant, C.; Filipovic, I.; Ponzetta, A.; Aleman, S.; Cornillet, M.; Hertwig, L.; Strunz, B.; Lentini, A.; Reinius, B.; Brownlie, D.; et al. Natural Killer Cell Immunotypes Related to COVID-19 Disease Severity. Sci. Immunol. 2020, 5, eabd6832. [Google Scholar] [CrossRef] [PubMed]
- Varchetta, S.; Mele, D.; Oliviero, B.; Mantovani, S.; Ludovisi, S.; Cerino, A.; Bruno, R.; Castelli, A.; Mosconi, M.; Vecchia, M.; et al. Unique Immunological Profile in Patients with COVID-19. Cell Mol. Immunol. 2021, 18, 604–612. [Google Scholar] [CrossRef]
- Costela-Ruiz, V.J.; Illescas-Montes, R.; Puerta-Puerta, J.M.; Ruiz, C.; Melguizo-Rodríguez, L. SARS-CoV-2 Infection: The Role of Cytokines in COVID-19 Disease. Cytokine Growth Factor Rev. 2020, 54, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Coperchini, F.; Chiovato, L.; Croce, L.; Magri, F.; Rotondi, M. The Cytokine Storm in COVID-19: An Overview of the Involvement of the Chemokine/Chemokine-Receptor System. Cytokine Growth Factor Rev. 2020, 53, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Hojyo, S.; Uchida, M.; Tanaka, K.; Hasebe, R.; Tanaka, Y.; Murakami, M.; Hirano, T. How COVID-19 Induces Cytokine Storm with High Mortality. Inflamm. Regen. 2020, 40, 37. [Google Scholar] [CrossRef]
- Inchingolo, A.D.; Inchingolo, A.M.; Bordea, I.R.; Malcangi, G.; Xhajanka, E.; Scarano, A.; Lorusso, F.; Farronato, M.; Tartaglia, G.M.; Isacco, C.G.; et al. SARS-CoV-2 Disease through Viral Genomic and Receptor Implications: An Overview of Diagnostic and Immunology Breakthroughs. Microorganisms 2021, 9, 793. [Google Scholar] [CrossRef] [PubMed]
- Inchingolo, F.; Martelli, F.S.; Gargiulo Isacco, C.; Borsani, E.; Cantore, S.; Corcioli, F.; Boddi, A.; Nguyễn, K.C.D.; De Vito, D.; Aityan, S.K.; et al. Chronic Periodontitis and Immunity, Towards the Implementation of a Personalized Medicine: A Translational Research on Gene Single Nucleotide Polymorphisms (SNPs) Linked to Chronic Oral Dysbiosis in 96 Caucasian Patients. Biomedicines 2020, 8, 115. [Google Scholar] [CrossRef]
- Cantore, S.; Mirgaldi, R.; Ballini, A.; Coscia, M.F.; Scacco, S.; Papa, F.; Inchingolo, F.; Dipalma, G.; De Vito, D. Cytokine Gene Polymorphisms Associate with Microbiogical Agents in Periodontal Disease: Our Experience. Int. J. Med. Sci. 2014, 11, 674–679. [Google Scholar] [CrossRef] [Green Version]
- Badawi, A. Hypercytokinemia and Pathogen–Host Interaction in COVID-19. J. Inflamm. Res. 2020, 13, 255–261. [Google Scholar] [CrossRef]
- Hou, Y.; Zhao, J.; Martin, W.; Kallianpur, A.; Chung, M.K.; Jehi, L.; Sharifi, N.; Erzurum, S.; Eng, C.; Cheng, F. New Insights into Genetic Susceptibility of COVID-19: An ACE2 and TMPRSS2 Polymorphism Analysis. BMC Med. 2020, 18, 216. [Google Scholar] [CrossRef]
- McElvaney, O.J.; Hobbs, B.D.; Qiao, D.; McElvaney, O.F.; Moll, M.; McEvoy, N.L.; Clarke, J.; O’Connor, E.; Walsh, S.; Cho, M.H.; et al. A Linear Prognostic Score Based on the Ratio of Interleukin-6 to Interleukin-10 Predicts Outcomes in COVID-19. EBioMedicine 2020, 61, 103026. [Google Scholar] [CrossRef]
- Odak, I.; Barros-Martins, J.; Bošnjak, B.; Stahl, K.; David, S.; Wiesner, O.; Busch, M.; Hoeper, M.M.; Pink, I.; Welte, T.; et al. Reappearance of Effector T Cells Is Associated with Recovery from COVID-19. EBioMedicine 2020, 57, 102885. [Google Scholar] [CrossRef] [PubMed]
- Di Micco, P.; Camporese, G.; Russo, V.; Cardillo, G.; Imbalzano, E.; Tufano, A.; Bernardi, E.; Fontanella, A. Clinical Differences between COVID-19 and a COVID-Like Syndrome. J. Clin. Med. 2021, 10, 2519. [Google Scholar] [CrossRef] [PubMed]
- Halpin, S.; O’Connor, R.; Sivan, M. Long COVID and chronic COVID syndromes. J. Med. Virol. 2021, 93, 1242–1243. [Google Scholar] [CrossRef]
- Raveendran, A.V.; Jayadevan, R.; Sashidharan, S. Long COVID: An overview. Diabetes Metab. Syndr. 2021, 15, 869–875. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
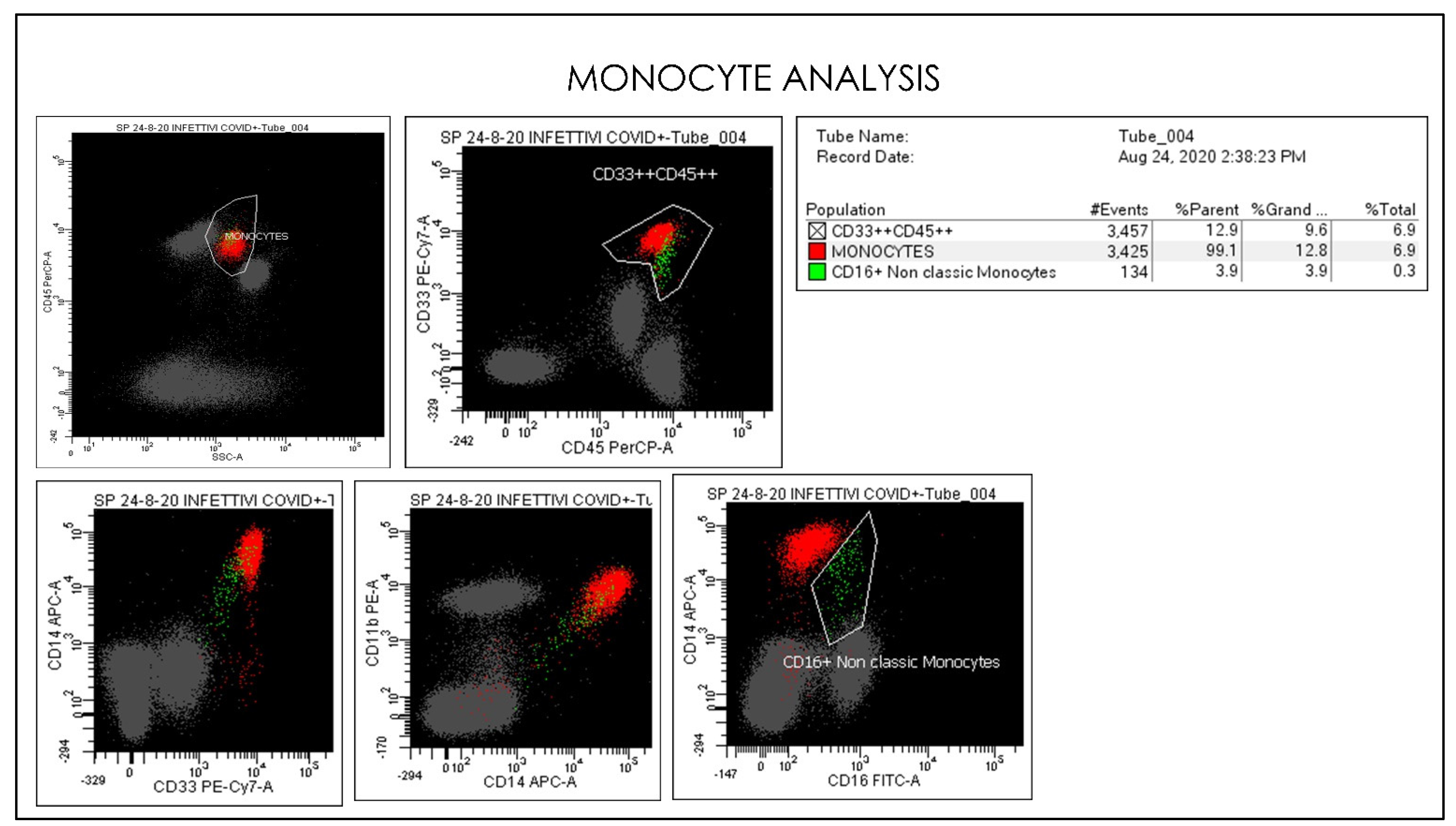{kind=link}
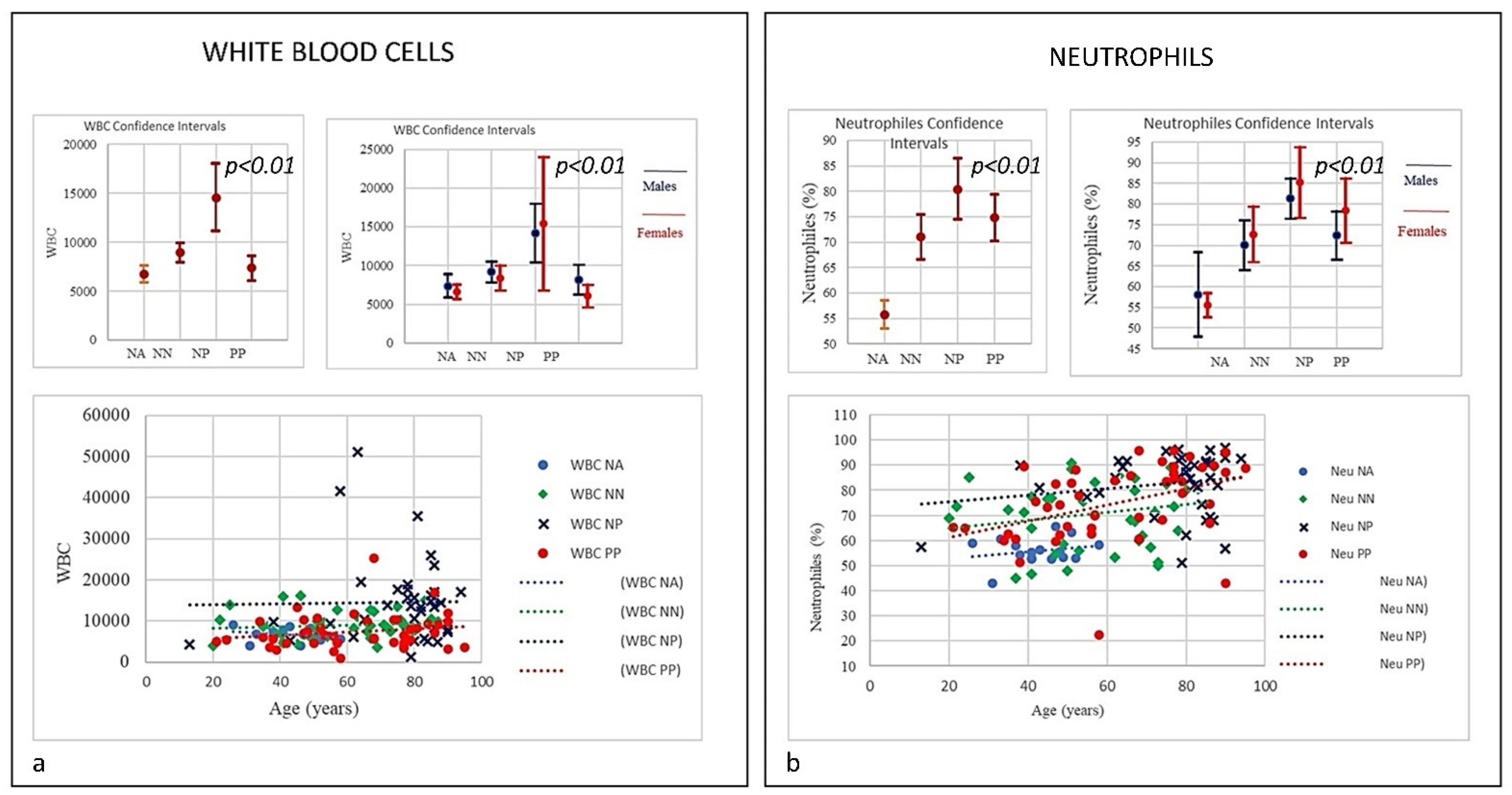
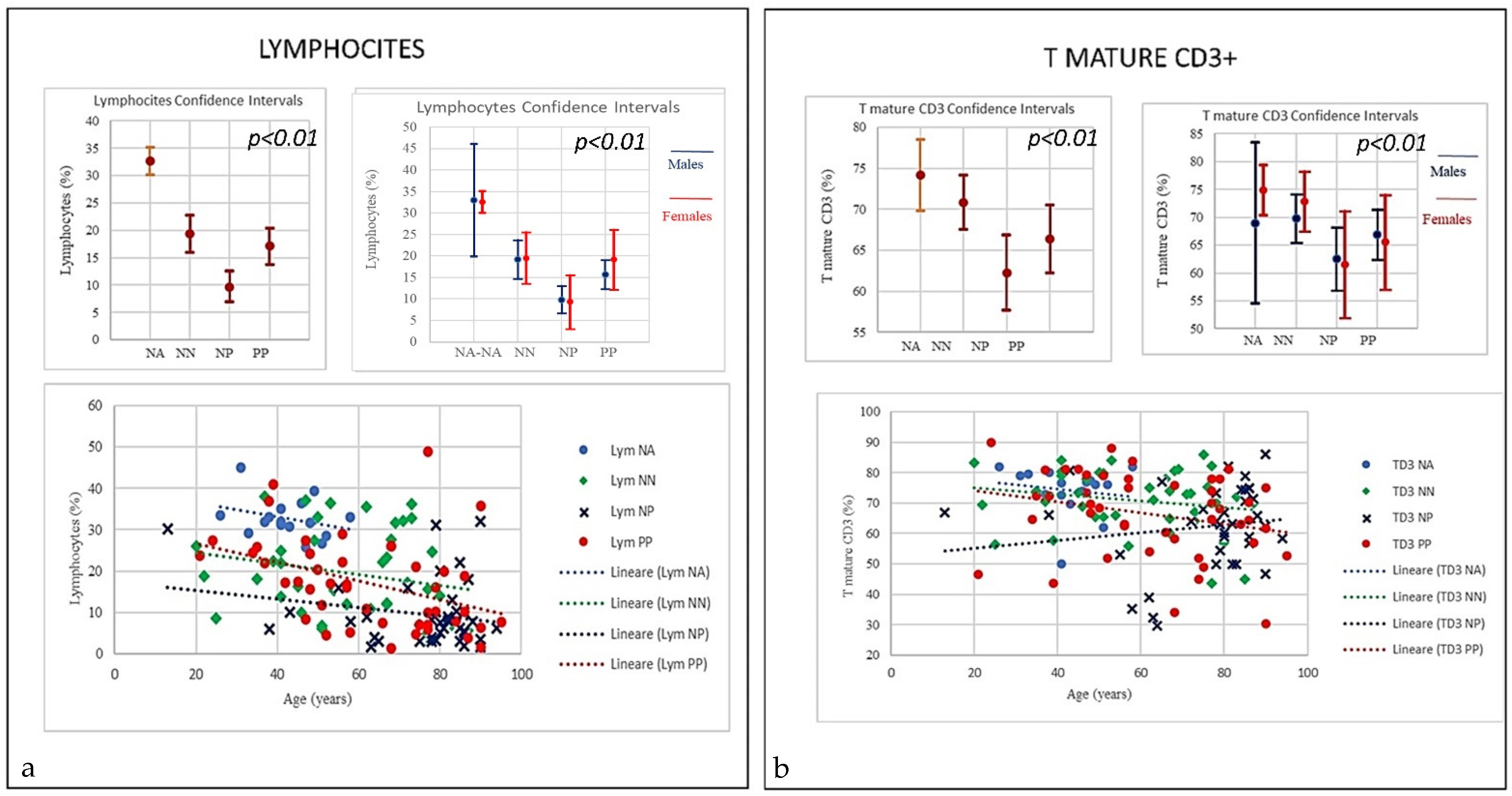
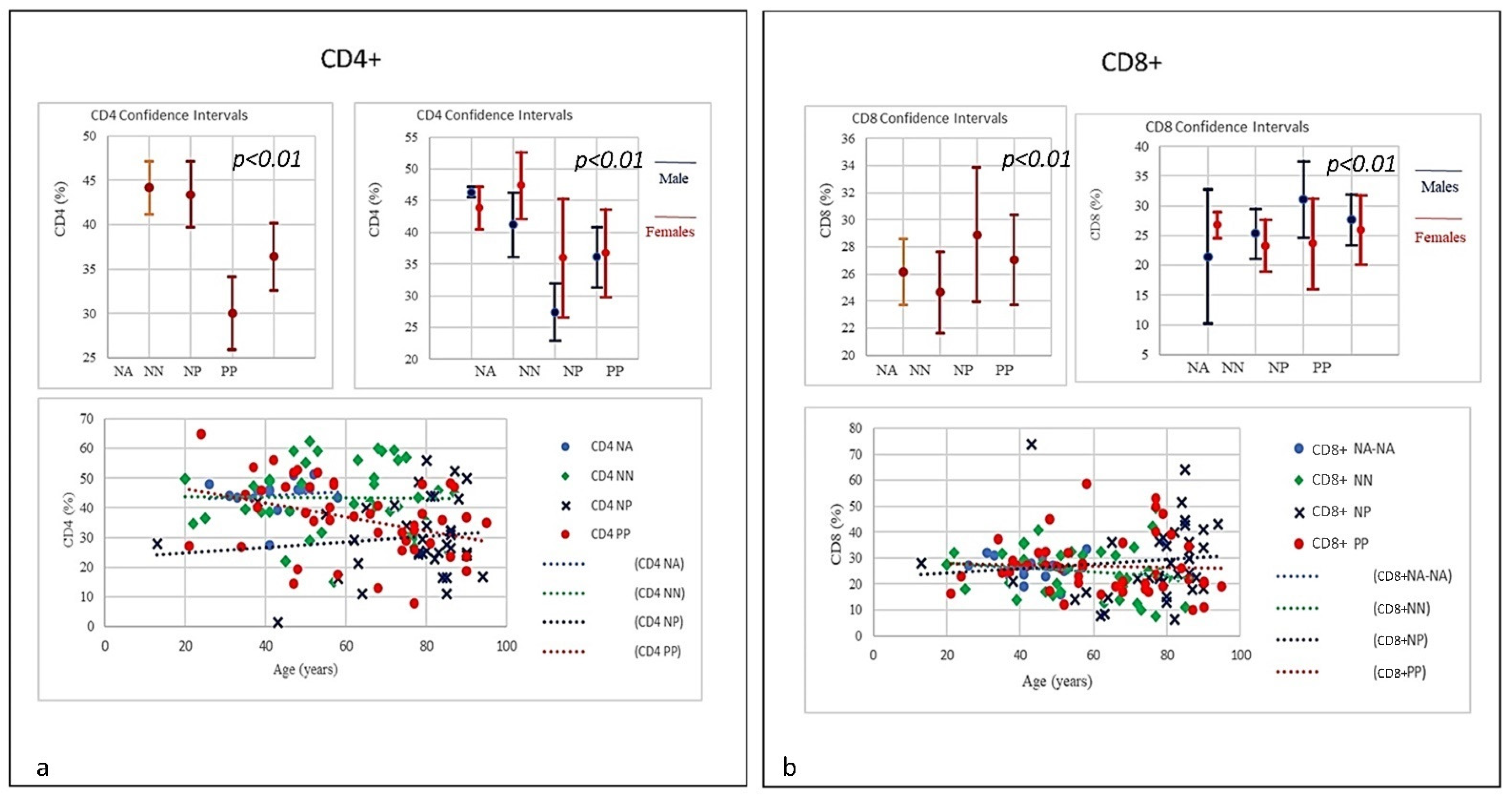
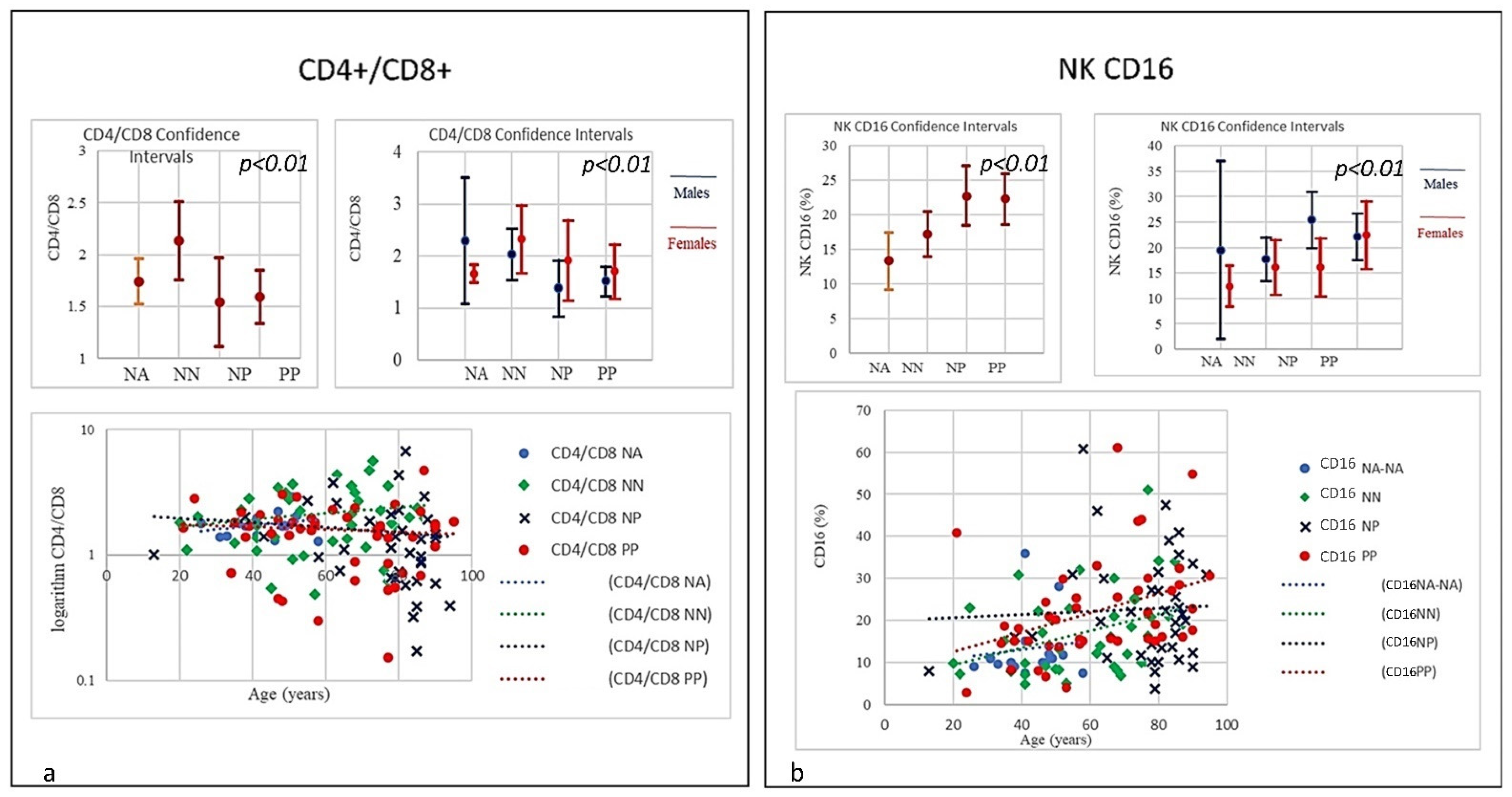
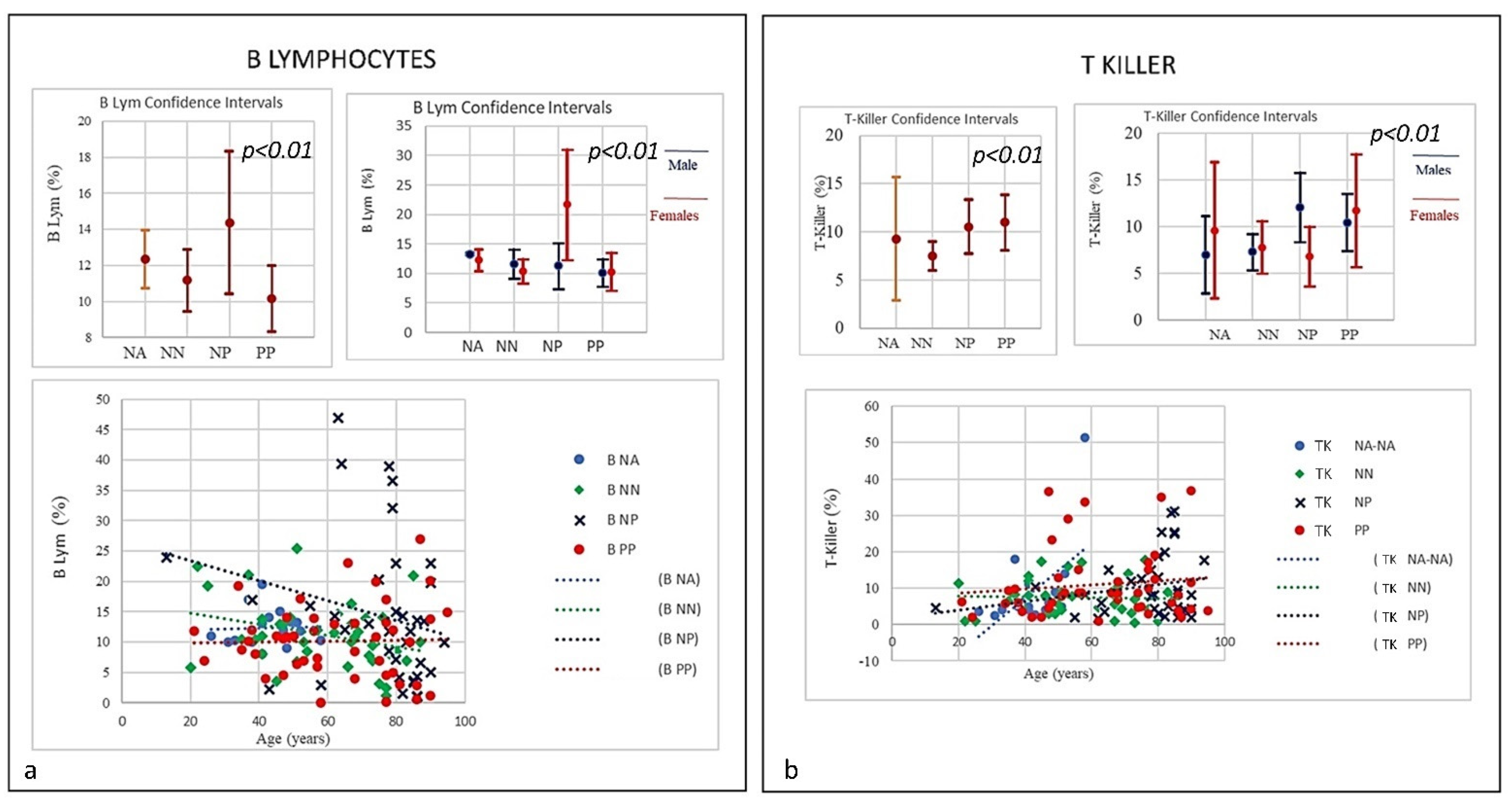
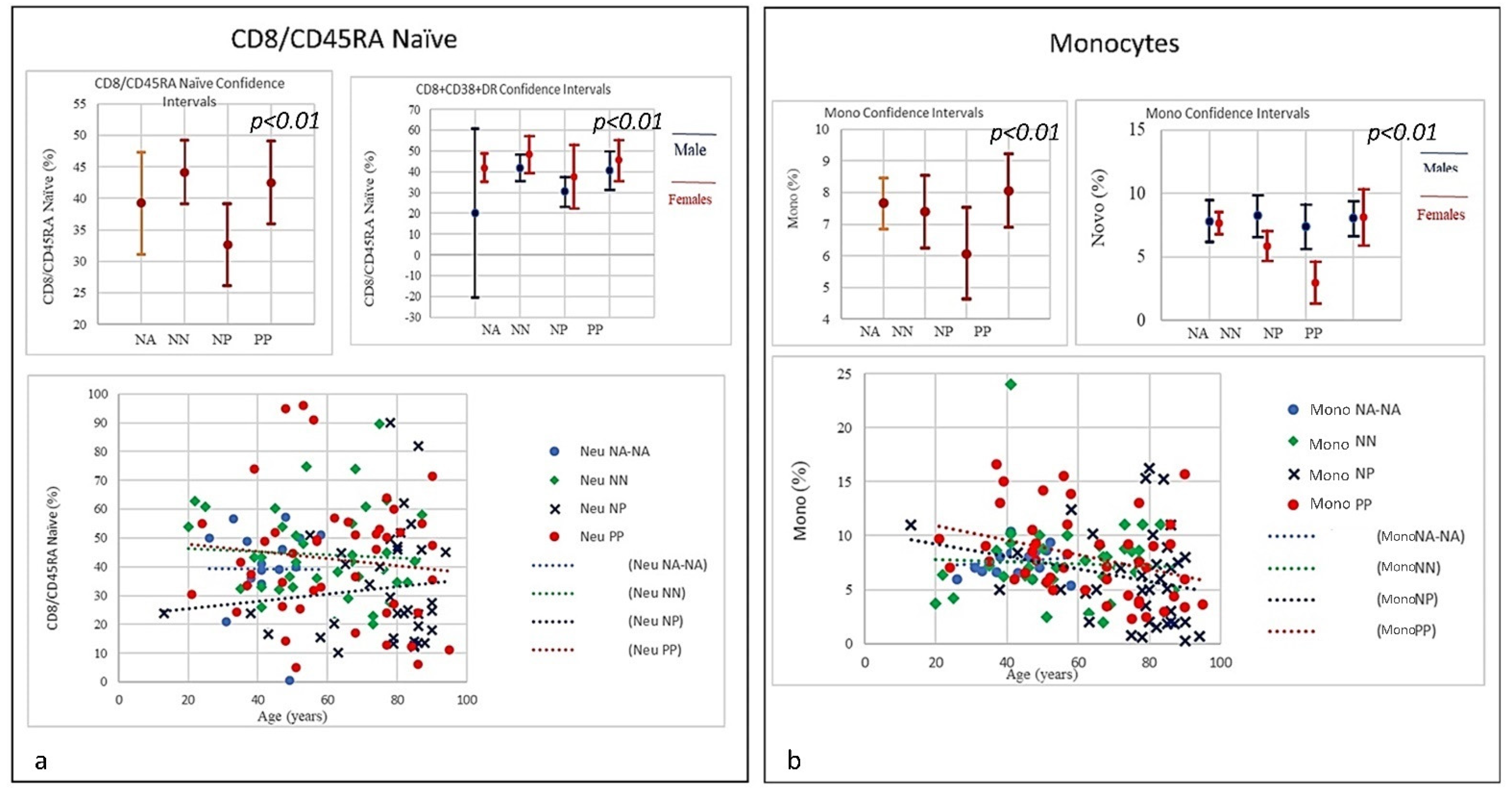
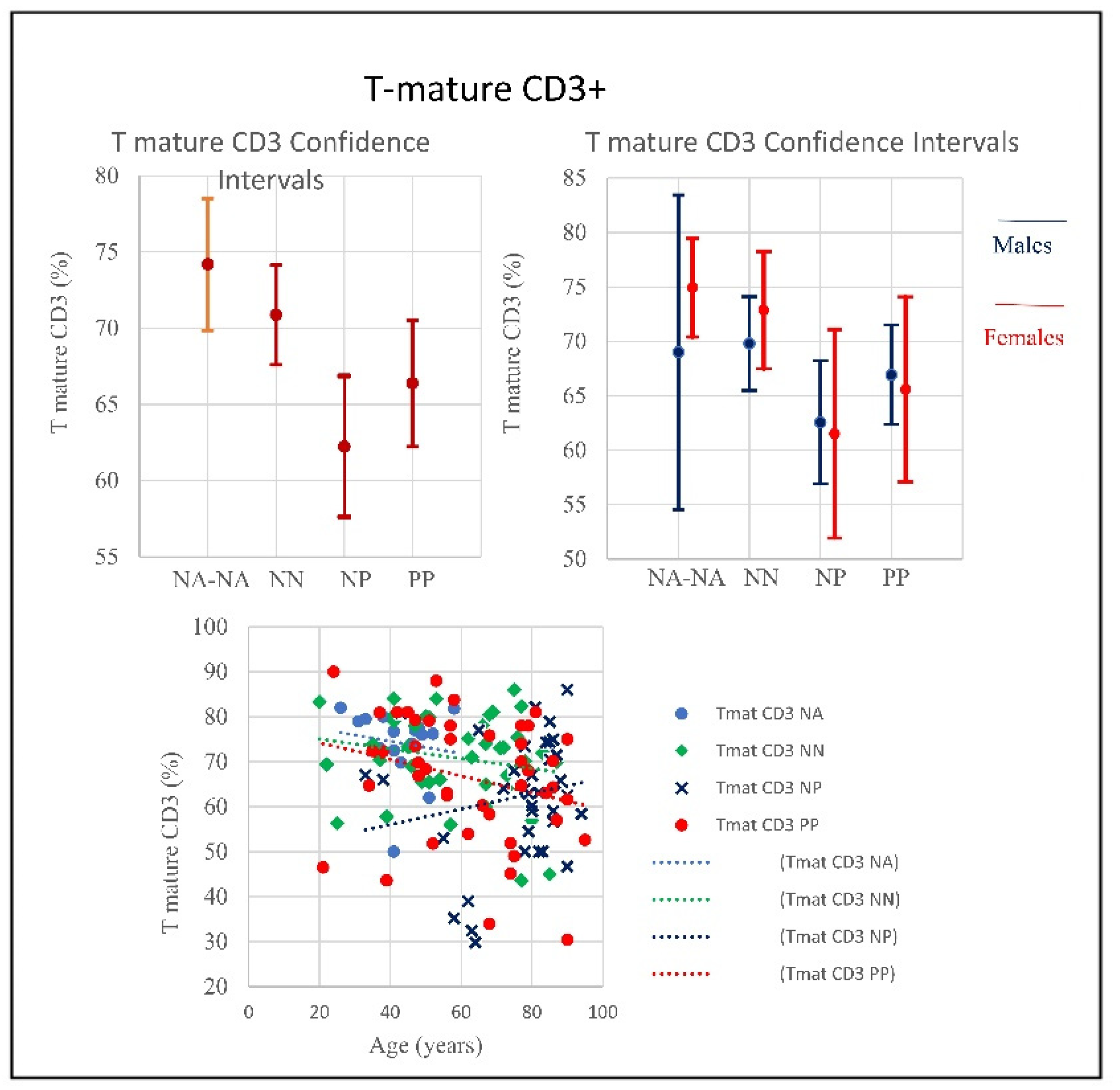
| SubSet NV% | Neut 45–75 | Lym 20–45 | Tmat CD3 61–84 | CD4 25–65 | CD8 15–50 | CD4/CD8 Ratio 1.0–4.0 | NK CD16+/CD56+ 4–28 | B Lym 10–40 | T-NK CD3+CD56+ <5 | T-CD3+DR+ Active 1–18 | CD8 Cyt. 1–28 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| PP | 49% | 64% | 31% | 20% | 7% | 26.67% | 26.67% | 60% | 73.3% | 13.3% | 0% |
| N. 45 | 22 ≥ 75 | 27 ≤ 20 | 14 ≤ 61 | 9 ≤ 25 | 3 ≤ 15 | 12 ≤ 1 | 12 ≥ 28 | 27 ≤ 10 | 33 ≥ 5 | 6 ≥ 18 | Norm |
| St.Dev | 15.20 | 10.95 | 13.82 | 12.68 | 11.10 | 0.85 | 12.14 | 6.12 | 9.57 | 11.41 | 8.17 |
| 19.42 | |||||||||||
| Mean | 74.85 | 17.07 | 66.38 | 36.38 | 27.03 | 1.59 | 22.26 | 10.16 | 10.98 | 11.70 | 15.10 |
| 62.33 | |||||||||||
| CI 95% | 4.56 | 3.29 | 4.15 | 3.81 | 3.33 | 0.25 | 3.65 | 1.84 | 2.87 | 3.43 | 2.45 |
| Conf. δ | |||||||||||
| NP | 78.4% | 91.9% | 40.5% | 43.2% | 18.9% | 43.2% | 35.1% | 43.2% | 67.6% | 45.9% | 0% |
| N.37 | 29 ≥ 5 | 34 ≤ 20 | 15 ≤ 61 | 16 ≤ 25 | 7 ≤ 15 | 16 ≤ 1 | 13 ≥ 28 | 16 ≤ 10 | 25 ≥ 5 | 17 ≥ 18 | Norm |
| St.Dev | 17.93 | 8.22 | 13.87 | 12.45 | 14.89 | 1.28 | 12.93 | 11.87 | 8.43 | 10.13 | 7.29 |
| 16.67 | |||||||||||
| Mean | 80.41 | 9.68 | 62.24 | 29.98 | 28.90 | 1.54 | 22.68 | 14.36 | 10.52 | 17.89 | 15.15 |
| 75.41 | |||||||||||
| CI 95% | 5.97 | 2.73 | 4.61 | 4.14 | 4.95 | 0.42 | 4.30 | 3.95 | 2.80 | 3.37 | 2.42 |
| Conf. δ | |||||||||||
| NN | 45% | 52.5% | 17.5% | 5% | 17.5% | 17.5% | 10% | 42.5% | 65% | 17.5% | 0% |
| N.40 | 18 ≥ 75 | 21 ≤ 20 | 7 ≤ 61 | 2 ≤ 25 | 7 ≤ 15 | 7 ≤ 15 | 4 ≥ 28 | 17 ≤ 10 | 26 ≥ 5 | 7 ≥ 18 | Norm |
| St.Dev | 13.75 | 10.76 | 10.21 | 11.67 | 9.38 | 1.18 | 10.03 | 5.39 | 4.76 | 7.19 | 6.27 |
| 17.93 | |||||||||||
| Mean | 70.99 | 19.34 | 70.86 | 43.42 | 24.66 | 2.1 | 17.19 | 11.18 | 7.49 | 11.58 | 14.76 |
| 58.5 | |||||||||||
| CI 95% | 4.39 | 3.43 | 3.26 | 3.73 | 2.99 | 0.37 | 3.20 | 1.72 | 1.52 | 2.29 | 2.00 |
| Conf. δ | |||||||||||
| NA | 0% | 0% | 6% | 0% | 0% | 0% | 12.5% | 12.5% | 62.5% | 0% | 0% |
| N. 16 | Norm | Norm | Norm | Norm | Norm | Norm | 2 ≥ 28 | 2 ≤ 10 | 10 ≥ 5 | Norm | Norm |
| St.Dev | 5.17 | 4.75 | 8.13 | 5.58 | 4.56 | 0.41 | 7.75 | 3.01 | 12.03 | 3.46 | 5.37 |
| 8.41 | |||||||||||
| Mean | 55.81 | 32.67 | 74.18 | 44.16 | 26.13 | 1.74 | 13.3 | 12.36 | 9.29 | 7.96 | 18.36 |
| 42.6 | |||||||||||
| CI 95% | 2.75 | 2.53 | 4.33 | 2.97 | 2.43 | 0.21 | 4.12 | 1.60 | 6.41 | 1.84 | 2.86 |
| Conf. δ | |||||||||||
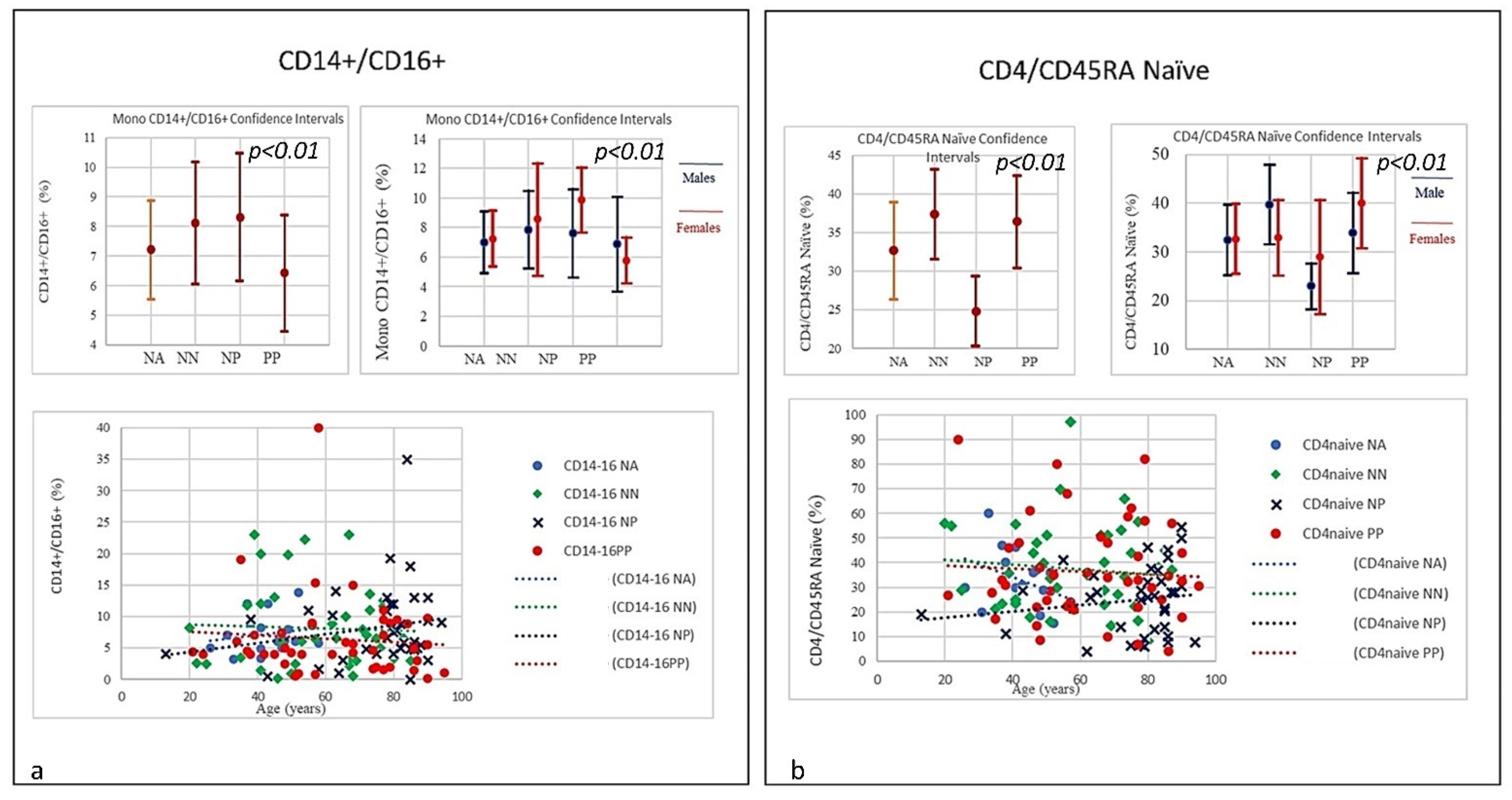
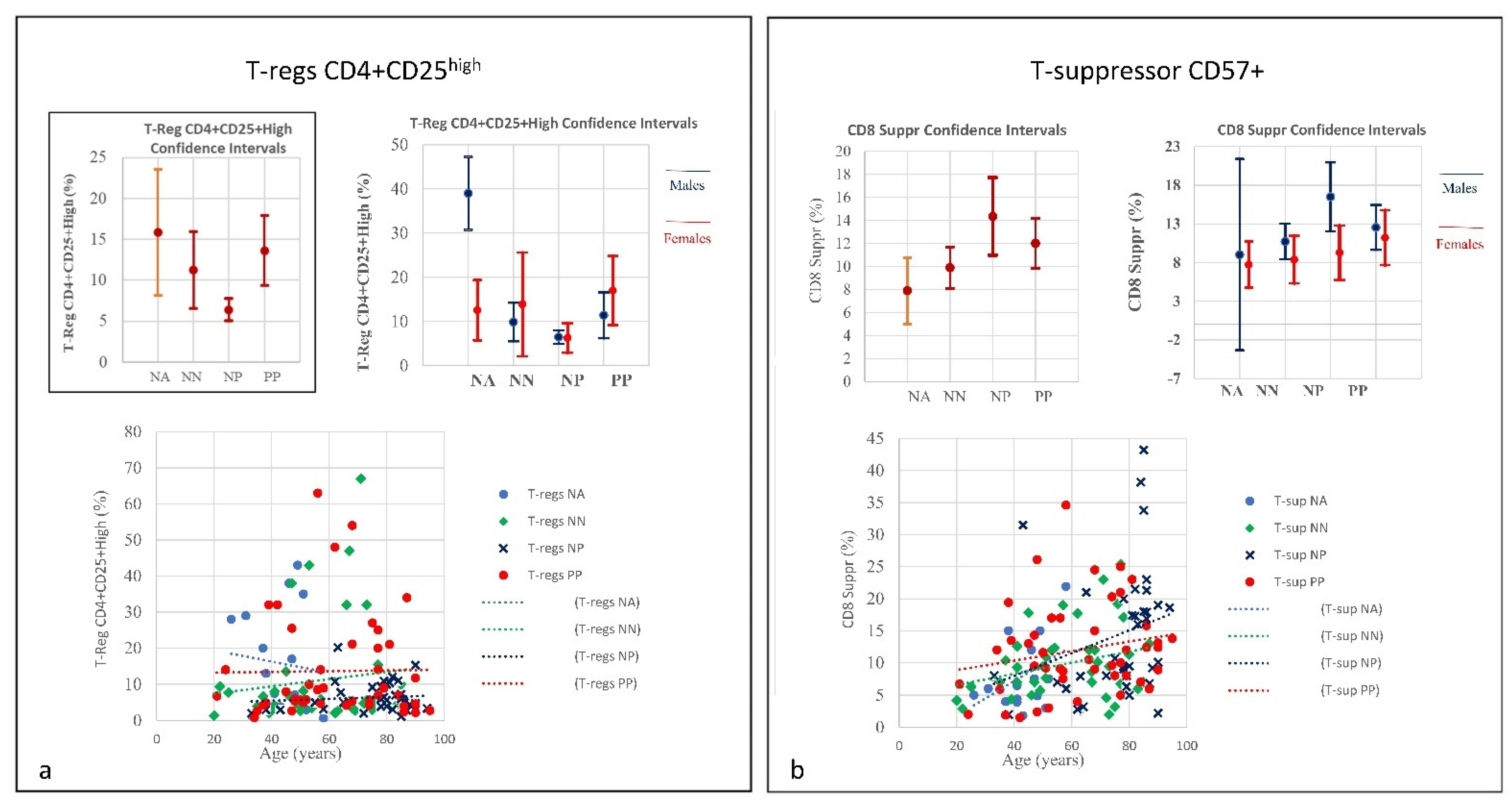
| SubSet NV% | CD8 suppr CD57+ 0–10 | CD8+CD38+ DR+ 0.30–2.30 | T-reg CD4+CD25+high < 10 | CD4/CD45RA Naïve 26–62 | CD8/CD45RA Naïve 16–40 | MONO 16–40 | MONO CD14+/CD16+ 1–10 | ||||
| PP | 64.44% | 80% | 37.78% | 33.33% | 53.33% | 28.89% | 17.78% | ||||
| N. 45 | 29 ≥ 10 | 36 ≥ 2.30 | 17 ≤ 10 | 15 ≤ 26 | 24 ≥ 40 | 13 ≥ 10 | 13 ≥ 10 | ||||
| St.Dev 19.42 | 7.21 | 11.97 | 14.33 | 19.94 | 22.00 | 3.84 | 6.53 | ||||
| Mean 62.33 | 12.00 | 8.52 | 13.64 | 36.42 | 42.52 | 8.06 | 6.43 | ||||
| CI 95% Conf. δ | 2.16 | 3.59 | 4.30 | 5.99 | 6.61 | 1.15 | 1.96 | ||||
| NP | 62.16% | 51.35% | 18.92% | 48.65% | 73.84% | 18.92% | 32.43% | ||||
| N. 37 | 23 ≥ 10 | 19 ≥ 2.30 | 7 ≤ 10 | 18 ≤ 26 | 14 ≥ 40 | 7 ≥ 10 | 12 ≥ 10 | ||||
| St.Dev 16.67 | 10.12 | 16.57 | 4.06 | 13.62 | 19.56 | 4.35 | 6.47 | ||||
| Mean 75.41 | 14.34 | 7.65 | 6.41 | 24.84 | 32.60 | 6.06 | 8.30 | ||||
| CI 95% Conf. δ | 3.37 | 5.51 | 1.35 | 4.53 | 6.51 | 1.45 | 2.15 | ||||
| NN | 37.5% | 45% | 25% | 32.5% | 55% | 17.5% | 13.5% | ||||
| N. 40 | 15 ≥ 10 | 18 ≥ 2.30 | 10 ≤ 10 | 13 ≤ 26 | 22 ≥ 40 | 7 ≥ 10 | 13 ≥ 10 | ||||
| St.Dev 17.93 | 5.62 | 2.43 | 14.66 | 18.32 | 15.66 | 3.62 | 6,45 | ||||
| Mean 58.5 | 9.89 | 2.23 | 11.26 | 37.4 | 44.19 | 7.4 | 8.12 | ||||
| CI 95% Conf. δ | 1.79 | 0.77 | 4.68 | 5.85 | 5.00 | 1.15 | 2.06 | ||||
| NA | 12.5% | 18.75% | 50% | 0% | 56.25% | 12.5% | 18.75% | ||||
| N. 16 | 2 ≥ 10 | 3 ≥ 2.30 | 8 ≤ 10 | Norm | 9 ≥ 40 | 2 ≥ 10 | 3 ≥ 10 | ||||
| St.Dev 8.41 | 5.42 | 2.59 | 14.45 | 11.84 | 15.27 | 1.51 | 3.11 | ||||
| Mean 42.6 | 7.88 | 1.99 | 15.86 | 32.66 | 39.23 | 7.66 | 7.21 | ||||
| CI 95% Conf. δ | 2.89 | 1.38 | 7.70 | 6.31 | 8.13 | 0.80 | 1.66 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balzanelli, M.G.; Distratis, P.; Dipalma, G.; Vimercati, L.; Catucci, O.; Amatulli, F.; Cefalo, A.; Lazzaro, R.; Palazzo, D.; Aityan, S.K.; et al. Immunity Profiling of COVID-19 Infection, Dynamic Variations of Lymphocyte Subsets, a Comparative Analysis on Four Different Groups. Microorganisms 2021, 9, 2036. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102036
Balzanelli MG, Distratis P, Dipalma G, Vimercati L, Catucci O, Amatulli F, Cefalo A, Lazzaro R, Palazzo D, Aityan SK, et al. Immunity Profiling of COVID-19 Infection, Dynamic Variations of Lymphocyte Subsets, a Comparative Analysis on Four Different Groups. Microorganisms. 2021; 9(10):2036. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102036
Chicago/Turabian StyleBalzanelli, Mario Giosuè, Pietro Distratis, Gianna Dipalma, Luigi Vimercati, Orazio Catucci, Felice Amatulli, Angelo Cefalo, Rita Lazzaro, Davide Palazzo, Sergey Khachatur Aityan, and et al. 2021. "Immunity Profiling of COVID-19 Infection, Dynamic Variations of Lymphocyte Subsets, a Comparative Analysis on Four Different Groups" Microorganisms 9, no. 10: 2036. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102036