
Active Surveillance for Highly Pathogenic Avian Influenza Viruses in Wintering Waterbirds in Northeast Italy, 2020–2021

 , , ,
, , ,
Abstract
:1. Introduction
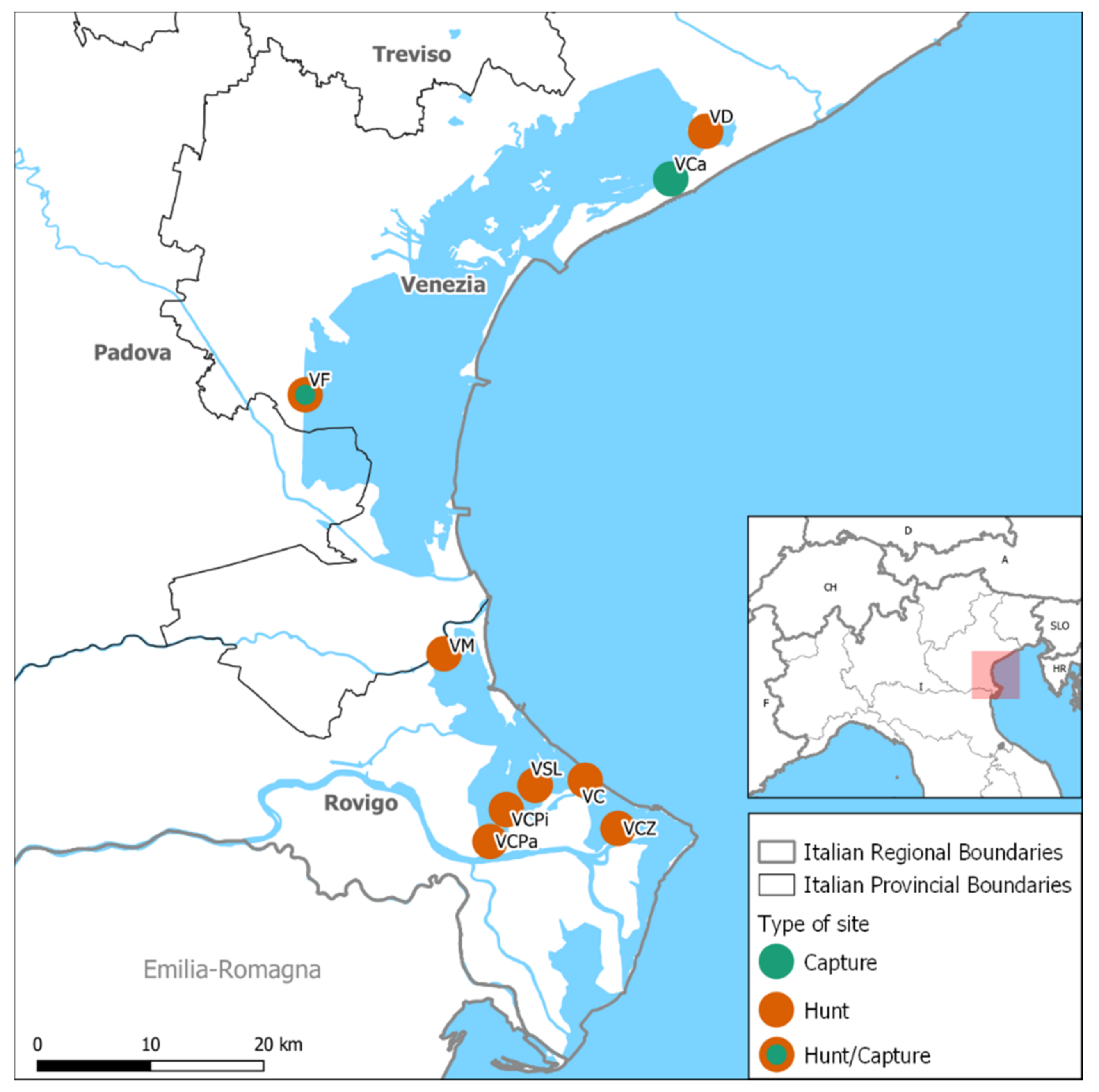
2. Materials and Methods
3. Results
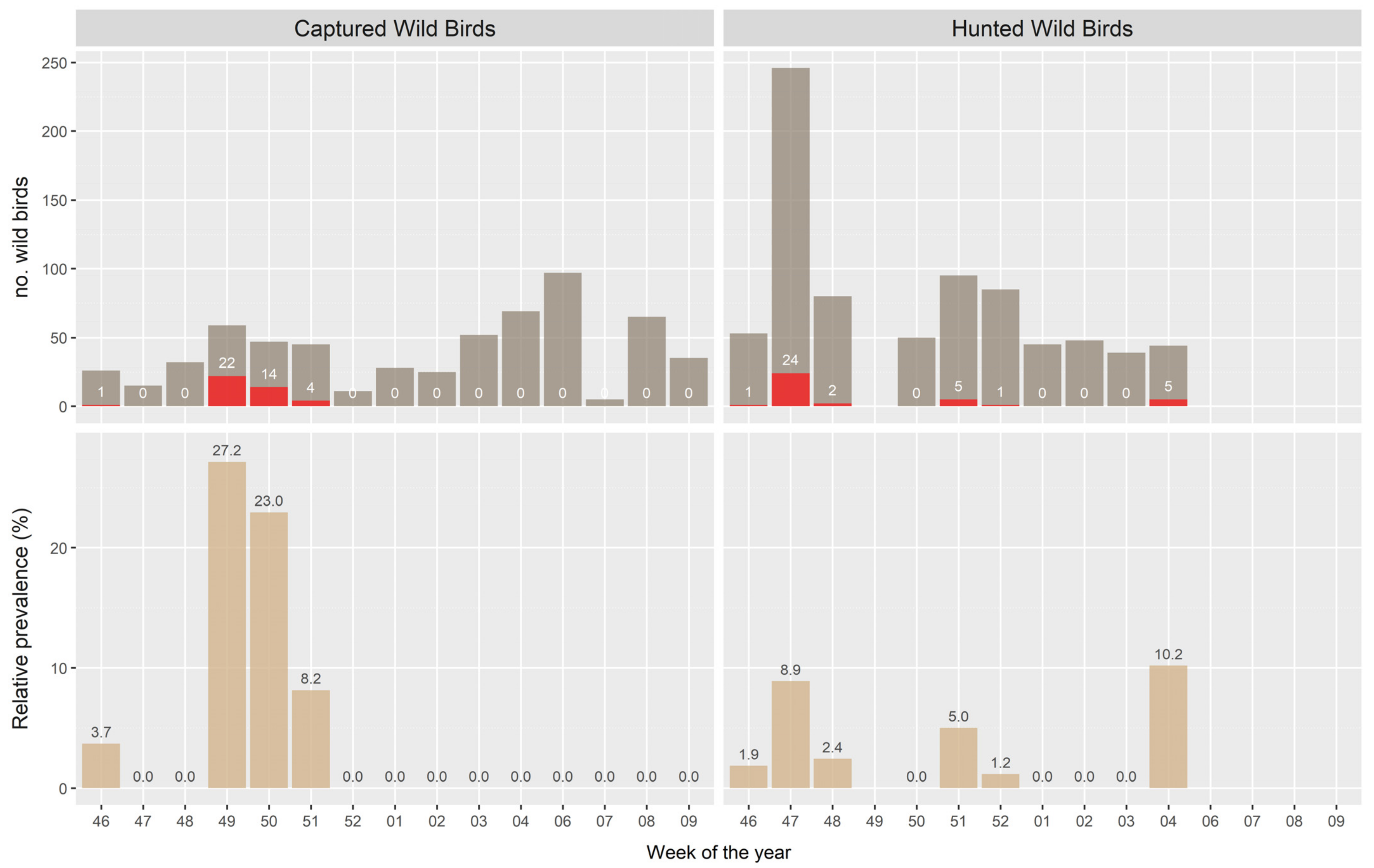
3.1. Surveillance Activity 1 (Hunted Ducks)
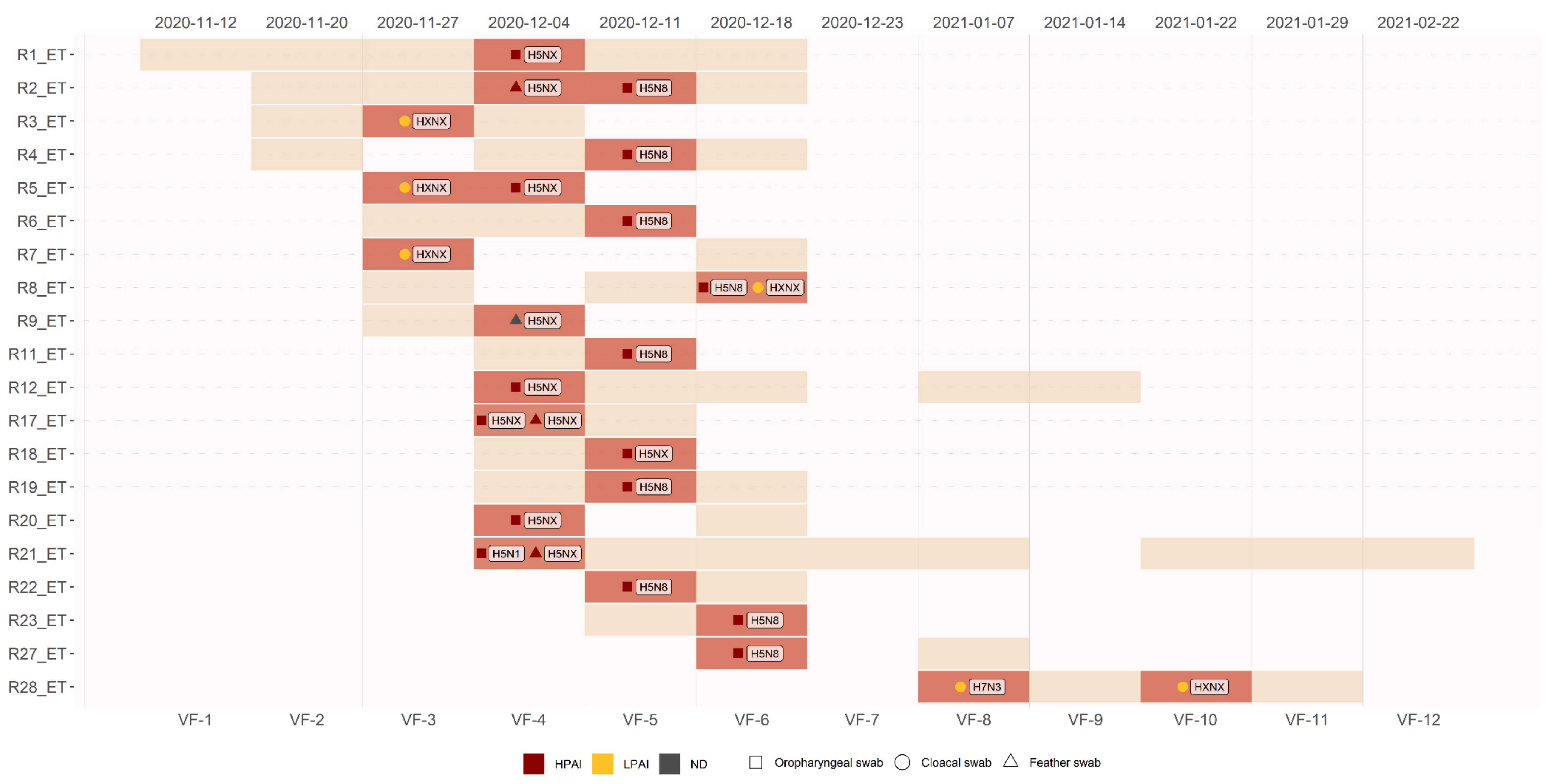
3.2. Survaillance Activity 2 (Captured Ducks)
3.3. Probability of HPAIV Detection
3.4. Genetic Analysis
3.5. Serological Analyses Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Verhagen, J.H.; Fouchier, R.A.M.; Lewis, N. Highly Pathogenic Avian Influenza Viruses at the Wild–Domestic Bird Interface in Europe: Future directions for Research and Surveillance. Viruses 2021, 13, 212. [Google Scholar] [CrossRef]
- Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Niqueux, É.; Staubach, C.; Terregino, C.; Lima, E.; Muñoz Guajardo, I.; et al. Scientific report: Avian influenza overview December 2020–February 2021. EFSA J. 2021, 19, 6497. [Google Scholar] [CrossRef]
- Dugan, V.G.; Chen, R.; Spiro, D.J.; Sengamalay, N.; Zaborsky, J.; Ghedin, E.; Nolting, J.; Swayne, D.E.; Runstadler, J.A.; Happ, G.M.; et al. The evolutionary genetics and emergence of avian influenza viruses in wild birds. PLoS Pathog. 2008, 4, e1000076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delogu, M.; De Marco, M.A.; Donatelli, I.; Campitelli, L.; Catelli, E. Ecological aspects of influenza A virus circulation in wild birds of the Western Palearctic. Vet. Res. Commun. 2003, 27 (Suppl. 1), 101–106. [Google Scholar] [CrossRef] [PubMed]
- Spina, F.; Volponi, S. Atlante della Migrazione Degli Uccelli in Italia. 1. Non-Passeriformi, 1st ed.; Ministero dell’Ambiente e della Tutela del Territorio e del Mare, Istituto Superiore per la Protezione e la Ricerca Ambientale (ISPRA); Tipografia CSR: Roma, Italy, 2008; pp. 113–189. ISBN 978-88-448-0378-0. [Google Scholar]
- Delogu, M.; De Marco, M.A.; Di Trani, L.; Raffini, E.; Cotti, C.; Puzelli, S.; Ostanello, F.; Webster, R.G.; Cassone, A.; Donatelli, I. Can Preening Contribute to Influenza A Virus Infection in Wild Waterbirds? PLoS ONE 2010, 5, e11315. [Google Scholar] [CrossRef] [Green Version]
- Fair, J.; Paul, E.; Jones, J. (Eds.) Guidelines to the Use of Wild Birds in Research; Ornithological Council: Washington, DC, USA, 2010; Available online: https://birdnet.org/info-for-ornithologists/guidelines-english-3rd-edition-2010/ (accessed on 24 August 2021).
- Spackman, E.; Senne, D.A.; Myers, T.J.; Bulaga, L.L.; Garber, L.P.; Perdue, M.L.; Lohman, K.; Daum, L.T.; Suarez, D.L. Development of a Real-Time Reverse Transcriptase PCR Assay for Type A Influenza Virus and the Avian H5 and H7 Hemagglutinin Subtypes. J. Clin. Microbiol. 2020, 40, 3256–3260. [Google Scholar] [CrossRef] [Green Version]
- Slomka, M.J.; Pavlidis, T.; Banks, J.; Shell, W.; McNally, A.; Essen, S.; Brown, I.H. Validated H5 Eurasian realtime reverse transcriptase-polymerase chain reaction and its application in H5N1 outbreaks in 2005–2006. Avian Dis. 2007, 51 (Suppl. 1), 373–377. [Google Scholar] [CrossRef] [PubMed]
- Slomka, M.J.; Pavlidis, T.; Coward, V.J.; Voermans, J.; Koch, G.; Hanna, A.; Banks, J.; Brown, I.H. Validated RealTime reverse transcriptase PCR methods for the diagnosis and pathotyping of Eurasian H7 avian influenza viruses. Influenza Other Respir. Viruses 2009, 3, 151–164. [Google Scholar] [CrossRef]
- Slomka, M.J.; Coward, V.J.; Banks, J.; Lӧndt, B.Z.; Brown, I.H.; Voermans, J.; Koch, G.; Handberg, K.J.; Jørgensen, P.H.; Cherbonnel-Pansart, M.; et al. Identification of sensitive and specific avian influenza polymerase chain reaction methods through blind ring trials organized in the European Union. Avian Dis. 2007, 51, 227–234. [Google Scholar] [CrossRef]
- Hoffmann, B.; Hoffmann, D.; Henritzi, D.; Beer, M.; Harder, T.C. Riems influenza A typing array (RITA): An RTqPCR-based low density array for subtyping avian and mammalian influenza A viruses. Sci. Rep. 2016, 6, 27211. [Google Scholar] [CrossRef] [Green Version]
- James, J.; Slomka, M.J.; Reid, S.M.; Thomas, S.S.; Mahmood, S.; Byrne, A.M.P.; Cooper, J.; Russel, C.; Mollett, B.C.; Agyeman-Dua, E.; et al. Development and application of real-time PCR assays for specific detection of contemporary avian influenza virus subtypes N5, N6, N7, N8 and N9. Avian Dis. 2018, 63 (Suppl. 1), 209–218. [Google Scholar] [CrossRef]
- Monne, I.; Ormelli, S.; Salviato, A.; De Battisti, C.; Bettini, F.; Salomoni, A.; Drago, A.; Zecchin, B.; Capua, I.; Cattoli, G. Development and validation of a one-step real-time PCR assay for simultaneous detection of subtype H5, H7, and H9 avian influenza viruses. J. Clin. Microbiol. 2008, 46, 1769–1773. [Google Scholar] [CrossRef] [Green Version]
- Diagnostic Protocols. Available online: https://www.izsvenezie.com/reference-laboratories/avian-influenza-newcastle-disease/diagnostic-protocols/ (accessed on 24 August 2021).
- OIE Terrestrial Manual. Chapter 3.3.4. Avian Influenza (Including Infection with High Pathogenicity Avian Influenza Viruses); OIE: Paris, France, 2021. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 24 August 2021).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R. emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.4.7. 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 24 August 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 1st ed.; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Van Der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; Del Angel, G.; Levy-Moonshine, A.; DePristo, M.A. From FastQ data to high confidence variant calls: The Genome Analysis ToolNeg best practices pipeline. Curr. Protoc. Bioinfor. 2014, 43, 11.10.1–11.10.33. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; DePristo, M.A. The Genome Analysis ToolNeg: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depristo, M.A.; Banks, E.; Poplin, R.E.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Wilm, A.; Aw, P.P.K.; Bertrand, D.; Yeo, G.H.T.; Ong, S.H.; Wong, C.H.; Khor, C.C.; Petric, R.; Hibberd, M.L.; Nagarajan, N. LoFreq: A sequence-quality aware, ultra-sensitive variant caller for uncovering cell-population heterogeneity from high-throughput sequencing datasets. Nucleic Acids Res. 2012, 40, 11189–11201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Delogu, M.; De Marco, M.A.; Cotti, C.; Di Trani, L.; Raffini, E.; Puzelli, S.; Webster, R.G.; Cassone, A.; Donatelli, I. Hu-man and animal integrated influenza surveillance Human and animal integrated influenza surveillance: A novel sampling approach for an additional transmission way in the aquatic bird reservoir. Ital. J. Public Health 2012, 9, 29–36. [Google Scholar]
- Harvey, W.T.; Mulatti, P.; Fusaro, A.; Scolamacchia, F.; Zecchin, B.; Monne, I.; Marangon, S. Spatiotemporal reconstruction and transmission dynamics during the 2016–17 H5N8 highly pathogenic avian influenza epidemic in Italy. Transbound. Emerg. Dis. 2021, 68, 37–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giunchi, D.; Baldaccini, N.E.; Lenzoni, A.; Luschi, P.; Sorrenti, M.; Cerritelli, G.; Vanni, L. Spring migratory routes and stopver duration of satellite-tracked Eurasian teal Anas crecca wintering in Italy. Ibis 2018, 161, 117–130. [Google Scholar] [CrossRef] [Green Version]
- Van den Brand, J.M.A.; Verhagen, J.H.; Veldhuis Kroeze, E.J.B.; Van de Bildt, M.W.G.; Bodewes, R.; Herfst, S.; Richard, M.; Lexmond, P.; Bestebroer, T.M.; Fouchier, R.A.M.; et al. Wild ducks excrete highly pathogenic avian influenza virus H5N8 (2014-2015) without clinical or pathological evidence of disease. Emerg. Microbes Infect. 2018, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Verhagen, J.H.; Van der Jeugd, H.P.; Nolet, B.A.; Slaterus, R.; Kharitonov, S.P.; De Vries, P.P.; Vuong, O.; Majoor, F.; Kuiken, T.; Fouchier, R.A. Wild bird surveillance around outbreaks of highly pathogenic avian influenza A (H5N8) virus in the Netherlands, 2014, within the context of global flyways. Eurosurveillance 2015, 20, 21069. [Google Scholar] [CrossRef]
- Jeong, J.; Kang, H.M.; Lee, E.K.; Song, B.M.; Kwon, Y.K.; Kim, H.R.; Choi, K.S.; Kim, J.Y.; Lee, H.J.; Moon, O.K.; et al. Highly pathogenic avian influenza (H5N8) in domestic poultry and its relationship with migratory birds in South Korea during 2014. Vet. Microbiol. 2014, 173, 249–257. [Google Scholar] [CrossRef]
- Marchenko, V.Y.; Susloparov, I.M.; Kolosova, N.P.; Goncharova, N.I.; Shipovalov, A.V.; Durymanov, A.G.; Ilyicheva, T.N.; Budatsirenova, L.V.; Ivanova, V.K.; Ignatyev, G.A.; et al. Influenza A(H5N8) virus isolation in Russia, 2014. Arch. Virol. 2015, 160, 2857–2860. [Google Scholar] [CrossRef]
- Poen, M.J.; Verhagen, J.H.; Manvell, R.J.; Brown, I.; Bestebroer, T.M.; Van der Vliet, S.; Vuong, O.; Scheuer, R.D.; Van der Jeugd, H.P.; Nolet, B.A.; et al. Lack of virological and serological evidence for continued circulation of higly pathogenic avian influenza H5N8 virus in wild birds in the Netherlands, 14 November to 31 January 2016. Eurosurveillance 2016, 38, 30349. [Google Scholar] [CrossRef] [Green Version]
- Kida, H.; Yanagawa, R.; Matsuoka, Y. Duck influenza lacking evidence of disease signs and immune response. Infect. Immun. 1980, 30, 547–553. [Google Scholar] [CrossRef]
- Fereidouni, S.R.; Starick, E.; Beer, M.; Wilking, H.; Kalthoff, D.; Grund, C.; Häuslaigner, R.; Breithaupt, A.; Lange, E.; Harder, T.C. Highly pathogenic avian influenza virus infection of mallards with homo- and heterosubtypic immunity induced by low pathogenic avian influenza viruses. PLoS ONE 2009, 20, e6706. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| HVSS ID | Samplingyy-mm-dd | Eurasian Teal | Gadwall | Northern Pintail | Eurasian Wigeon | Mallard | Northern Shoveler | Dabbling Ducks | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ET | Pos. N. (%) | Ga | Pos. N. (%) | NP | Pos. N. (%) | EW | Pos. N. (%) | Ma | Pos. N. (%) | NS | Pos. N. (%) | DD | Pos. N. (%) | ||||||||||||||||
| OS | CS | FS | OS | CS | FS | OS | CS | FS | OS | CS | FS | OS | CS | FS | OS | CS | FS | OS | CS | FS | |||||||||
| VCPa-1st | 20-11-09 | 7 | 0 | 1 (14.3) | 0 | 19 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 6 | 0 | 2 (33.3) | 0 | 8 | 0 | 0 | 0 | 13 | 0 | 0 | 0 | 54 | 0 | 3 (5.6) | 0 |
| VCZ-1st | 20-11-16 | 1 | 0 | 0 | 0 | 5 | 0 | 0 | 1 (20) | 4 | 0 | 0 | 0 | 10 | 1 (10) | 2 (20) | 0 | 19 | 1 (5.3) | 1 (5.3) | 2 (10.5) | 1 | 0 | 0 | 0 | 40 | 2 (5) | 3 (7.5) | 3 (7.5) |
| VD-1st | 20-11-21 | 0 | — | — | — | 4 | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 100 | 10 (10) | 9 (9) | 5 (5) | 0 | — | — | — | 11 | 0 | 1 (9.1) | 1 (9.1) | 120 | 10 (8.3) | 10 (8.3) | 6 (5) |
| VSL-1st | 20-11-21 | 8 | 0 | 0 | 0 | 0 | — | — | — | 0 | — | — | — | 54 | 2 (3.7) | 3 (5.6) | 1 (1.9) | 8 | 0 | 0 | 0 | 40 | 0 | 0 | 0 | 110 | 2 (1.8) | 3 (2.7) | 1 (0.9) |
| VC-1st | 20-11-26 | 32 | 2 (6.3) | 0 | 2 (6.3) | 2 | 0 | 0 | 0 | 0 | — | — | — | 16 | 0 | 0 | 2 (12.5) | 8 | 0 | 0 | 1 (12.5) | 4 | 0 | 0 | 0 | 62 | 2 (3.2) | 0 | 5 (8.1) |
| VD-2nd | 20-11-28 | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 20 | 0 | 0 | — | 0 | — | — | — | 0 | — | — | — | 20 | 0 | 0 | — |
| VCPi | 20-12-07 | 46 | 1 (2.2) | 0 | 0 | 0 | — | — | — | 0 | — | — | — | 2 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | — | — | — | 50 | 1 (2) | 0 | 0 |
| VCPa-2nd | 20-12-15 | 1 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | — | — | — | 89 | 3 (3.4) | 4 (4.5) | 1 (1.1) | 5 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 100 | 3 (3) | 4 (4) | 1 (1) |
| VM-1st | 20-12-22 | 60 | 1 (1.7) | 0 | 0 | 2 | 0 | 0 | 0 | 0 | — | — | — | 0 | — | — | — | 23 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 86 | 1 (1.2) | 0 | 0 |
| VC-2nd | 21-01-09 | 20 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | — | — | — | 16 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 45 | 0 | 0 | 0 |
| VD-3rd | 21-01-11 | 15 | 0 | 1 (6.7) | 0 | 0 | — | — | — | 0 | — | — | — | 15 | 0 | 0 | 0 | 0 | — | — | — | 0 | — | — | — | 30 | 0 | 1 (3.3) | 0 |
| VSL-2nd | 21-01-16 | 1 | 0 | 0 | 0 | 0 | — | — | — | 0 | — | — | — | 4 | 0 | 0 | 0 | 6 | 0 | 0 | 0 | 7 | 0 | 1 (14.3) | 0 | 18 | 0 | 1 (5.6) | 0 |
| VF | 21-01-24 | 1 | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 15 | 0 | 0 | 0 | 8 | 0 | 1 (12.5) | 0 | 39 | 0 | 1 (2.6) | 0 |
| VCZ-2nd | 21-01-25 | 3 | 1 (33.3) | 0 | 0 | 0 | — | — | — | 0 | — | — | — | 3 | 1 (33.3) | 0 | 2 (66.7) | 5 | 1 (20) | 0 | 1 (20) | 2 | 1 (50) | 0 | 0 | 13 | 4 (30.8) | 0 | 3 (23.1) |
| VM-2nd | 21-01-30 | 25 | 0 | 1 (4) | 0 | 2 | 0 | 0 | 0 | 0 | — | — | — | 0 | — | — | — | 8 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 36 | 0 | 1 (2.8) | 0 |
| Total duck & Pos. (%) | 220 | 5 (2.3) | 3 (1.4) | 2 (0.9) | 49 | 0 | 0 | 1 (2) | 12 | 0 | 0 | 0 | 337 | 17 (5) | 20 (5.9) | 11 (3.3) | 111 | 2 (1.8) | 1 (0.9) | 4 (3.6) | 94 | 1 (1.1) | 3 (3.2) | 1 (1.1) | 823 | 25 (3) | 27 (3.3) | 19 (2.3) | |
| TSS ID | Samplingyy-mm-dd | Eurasian Teal | Gadwall | Northern Pintail | Eurasian Wigeon | Mallard | Common Shelduck | Dabbling Ducks | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ET | Pos. N. (%) | Ga | Pos. N. (%) | NP | Pos. N. (%) | EW | Pos. N. (%) | Ma | Pos. N. (%) | CSh | Pos. N. (%) | DD | Pos. N. (%) | ||||||||||||||||
| OS | CS | FS | OS | CS | FS | OS | CS | FS | OS | CS | FS | OS | CS | FS | OS | CS | FS | OS | CS | FS | |||||||||
| VF-1 | 20-11-12 | 24 | 1 (4.2) | 0 | 0 | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 3 | 0 | 0 | 0 | 0 | — | — | — | 27 | 1 (3.7) | 0 | 0 |
| VF-2 | 20-11-12 | 10 | 0 | 1 (10) | 0 | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 3 | 0 | 0 | 0 | 0 | — | — | — | 13 | 0 | 1 (7.7) | 0 |
| VF-3 | 20-11-27 | 24 | 0 | 2 (8.3) | 0 | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 2 | 0 | 0 | 0 | 0 | — | — | — | 26 | 0 | 2 (7.7) | 0 |
| VF-4 | 20-12-04 | 68 | 15 (22.1) | 4 (5.9) | 9 (13.2) | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 1 | 0 | 0 | 0 | 0 | — | — | — | 69 | 15 (21.7) | 4 (5.8) | 9 (13) |
| VF-5 | 20-12-11 | 36 | 7 (19.4) | 0 | 1 (2.8) | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 2 | 0 | 0 | 0 | 0 | — | — | — | 38 | 7 (18.4) | 0 | 1 (2.6) |
| VF-6 | 20-12-18 | 24 | 2 (8.3) | 1 (4.2) | 0 | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 2 | 0 | 0 | 0 | 0 | — | — | — | 26 | 2 (7.7) | 1 (3.8) | 0 |
| VF-7 | 20-12-23 | 6 | 0 | 0 | 0 | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 1 | 0 | 0 | 0 | 0 | — | — | — | 7 | 0 | 0 | 0 |
| VF-8 | 21-01-07 | 10 | 0 | 1(10) | 0 | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 1 * | 0 | 0 | 0 | 0 | — | — | — | 11 | 0 | 1 (9.1) | 0 |
| VF-9 | 21-01-14 | 6 | 0 | 0 | 0 | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 10 | 0 | 0 | 1 (10) | 0 | — | — | — | 16 | 0 | 0 | 1 (6.3) |
| VF-10 | 21-01-22 | 40 | 0 | 2 (5) | 0 | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 6 ** | 0 | 0 | 0 | 0 | — | — | — | 46 | 0 | 2 (4.3) | 0 |
| VF-11 | 21-01-29 | 51 | 1 (2) | 3 (5.9) | 0 | 2 | 0 | 0 | 0 | 0 | — | — | — | 0 | — | — | — | 9 | 1 (11.1) | 0 | 0 | 0 | — | — | — | 62 | 2 (3.2) | 3 (4.8) | 0 |
| VF-12 | 21-02-22 | 19 | 0 | 0 | 0 | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 2 | 0 | 0 | 0 | 0 | — | — | — | 21 | 0 | 0 | 0 |
| VF-13 | 21-02-25 | 29 | 0 | 2 (6.9) | 0 | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 5 | 0 | 0 | 0 | 0 | — | — | — | 34 | 0 | 2 (5.9) | 0 |
| VF-14 | 21-03-05 | 9 | 0 | 0 | 0 | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 14 | 0 | 0 | 0 | 0 | — | — | — | 23 | 0 | 0 | 0 |
| VCa-1 | 21-02-12 | 18 | 0 | 0 | 0 | 0 | — | — | — | 74 | 0 | 3 (4.1) | 0 | 4 | 0 | 0 | 0 | 0 | — | — | — | 1 | 0 | 1 (100) | 0 | 97 | 0 | 4 (4.1) | 0 |
| VCa-2 | 21-02-15 | 0 | — | — | — | 0 | — | — | — | 5 | 0 | 1 (20) | 0 | 0 | — | — | — | 0 | — | — | — | 0 | — | — | — | 5 | 0 | 1 (20) | 0 |
| Total duck & Pos. (%) | 374 | 26 (7) | 16 (4.3) | 10 (2.7) | 2 | 0 | 0 | 0 | 79 | 0 | 4 (5.1) | 0 | 4 | 0 | 0 | 0 | 61 | 1 (1.6) | 0 | 1 (1.6) | 1 | 0 | 1 (100) | 0 | 521 | 27 (5.2) | 21 (4) | 11 (2.1) | |
| AIV Pos | OS Virological Results | CS Virological Results | FS Virological Results | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HVSS ID (yy-mm-dd) | H-Duck ID | Sex | Age | AIV M Gene | AIV P-T | AIV S-T | AIV M Gene | AIV P-T | AIV S-T | AIV M Gene | AIV P-T | AIV S-T |
| VCPa-1st (20-11-09) | H1_ET | n.a. | n.a. | - | n.d. | n.d. | Pos. | LPAI | H5Nx | - | n.d. | n.d. |
| H2_EW | n.a. | n.a. | - | n.d. | n.d. | Pos. | HPAI | H5Nx | - | n.d. | n.d. | |
| H3_EW | n.a. | n.a. | - | n.d. | n.d. | Pos. | LPAI | H9Nx | - | n.d. | n.d. | |
| VCZ-1st (20-11-16) | H4_EW | n.a. | n.a. | - | n.d. | n.d. | Pos. | HPAI | H5Nx | - | n.d. | n.d. |
| H5_EW | n.a. | n.a. | Pos. | HPAI | H5N8 | Pos. | HPAI | H5Nx | - | n.d. | n.d. | |
| H6_Ga | n.a. | n.a. | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | LPAI | HxNx | |
| H7_Ma | n.a. | n.a. | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | LPAI | HxNx | |
| H8_Ma | n.a. | n.a. | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5N8 | |
| H9_Ma | n.a. | n.a. | Pos. | HPAI | H5N8 | Pos. | HPAI | H5N8 | - | n.d. | n.d. | |
| VD-1st (20-11-21) | H10_EW | F | J | Pos. | HPAI | H5N8 | Pos. | HPAI | H5Nx | - | n.d. | n.d. |
| H11_EW | F | J | Pos. | HPAI | H5Nx | Pos. | HPAI | H5Nx | - | n.d. | n.d. | |
| H12_EW | F | A | Pos. | n.a. | H5Nx | - | n.d. | n.d. | - | n.d. | n.d. | |
| H13_EW | F | J | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| H14_EW | M | A | - | n.d. | n.d. | Pos. | n.a. | H5Nx | - | n.d. | n.d. | |
| H15_EW | F | J | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. | |
| H16_EW | F | J | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. | |
| H17_EW | F | J | - | n.d. | n.d. | Pos. | HPAI | H5Nx | - | n.d. | n.d. | |
| H18_EW | M | A | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| H19_EW | F | J | Pos. | HPAI | H5Nx | Pos. | n.a. | H5Nx | - | n.d. | n.d. | |
| H20_EW | M | J | Pos. | n.a. | H5Nx | - | n.d. | n.d. | - | n.d. | n.d. | |
| H21_EW | M | J | - | n.d. | n.d. | Pos. | HPAI | H5Nx | - | n.d. | n.d. | |
| H22_EW | F | J | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. | |
| H23_EW | M | A | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| H24_EW | M | J | Pos. | HPAI | H5Nx | Pos. | HPAI | H5Nx | Pos. | HPAI | H5Nx | |
| H25_EW | F | A | Pos. | HPAI | H5Nx | - | n.d. | n.d. | - | n.d. | n.d. | |
| H26_EW | M | J | - | n.d. | n.d. | Pos. | n.a. | H5Nx | - | n.d. | n.d. | |
| H27_EW | M | J | - | n.d. | n.d. | Pos. | HPAI | H5N1 | - | n.d. | n.d. | |
| H28_EW | M | J | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| H29_NS | F | J | - | n.d. | n.d. | Pos. | HPAI | H5Nx | - | n.d. | n.d. | |
| H30_NS | F | J | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| VSL-1st (20-11-21) | H31_EW | n.a. | n.a. | Pos. | HPAI | H5N8 | Pos. | HPAI | H5N8 | - | n.d. | n.d. |
| H32_EW | n.a. | n.a. | - | n.d. | n.d. | Pos. | n.a. | H5Nx | Pos. | HPAI | H5N8 | |
| H33_EW | n.a. | n.a. | Pos. | HPAI | H5N8 | Pos. | HPAI | H5N8 | - | n.d. | n.d. | |
| VC-1st (20-11-26) | H34_Ma | n.a. | n.a. | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | n.a. | H5Nx |
| H35_ET | n.a. | n.a. | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. | |
| H36_ET | n.a. | n.a. | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | n.a. | H5Nx | |
| H37_ET | n.a. | n.a. | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | n.a. | H5Nx | |
| H38_ET | n.a. | n.a. | Pos. | HPAI | H5N5 | - | n.d. | n.d. | - | n.d. | n.d. | |
| H39_EW | n.a. | n.a. | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | n.a. | H5Nx | |
| H40_EW | n.a. | n.a. | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | n.a. | H5Nx | |
| VCPi (20-12-07) | H41_ET | n.a. | n.a. | Pos. | LPAI | HxNx | - | n.d. | n.d. | - | n.d. | n.d. |
| VCPa-2nd (20-12-15) | H42_EW | n.a. | n.a. | Pos. | HPAI | H5N8 | Pos. | HPAI | H5N8 | - | n.d. | n.d. |
| H43_EW | n.a. | n.a. | Pos. | HPAI | H5N8 | Pos. | HPAI | H5N8 | Pos. | HPAI | H5N8 | |
| H44_EW | n.a. | n.a. | Pos. | HPAI | H5N1 | - | n.d. | n.d. | - | n.d. | n.d. | |
| H45_EW | n.a. | n.a. | - | n.d. | n.d. | Pos. | HPAI | H5N1 | - | n.d. | n.d. | |
| H46_EW | n.a. | n.a. | - | n.d. | n.d. | Pos. | HPAI | H5N1 | - | n.d. | n.d. | |
| VM-1st (20-12-22) | H47_ET | n.a. | n.a. | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. |
| VD-3rd (21-01-11) | H48_ET | M | J | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. |
| VSL-2nd (21-01-16) | H49_NS | n.a. | n.a. | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. |
| VF (21-01-24) | H50_NS | M | J | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. |
| VCZ-2nd (21-01-25) | H51_Ma | n.a. | n.a. | Pos. | HPAI | H5N8 | - | n.d. | n.d. | Pos. | HPAI | H5N8 |
| H52_EW | n.a. | n.a. | Pos. | HPAI | H5N8 | - | n.d. | n.d. | Pos. | HPAI | H5N8 | |
| H53_NS | n.a. | n.a. | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. | |
| H54_ET | n.a. | n.a. | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. | |
| H55_EW | n.a. | n.a. | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5N8 | |
| VM-2nd (21-01-30) | H56_ET | n.a. | n.a. | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. |
| AIV Pos | OS Virological Results | CS Virological Results | FS Virological Results | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TSS ID (yy-mm-dd) | C-Duck ID | Sex | Age | AIV M Gene | AIV P-T | AIV S-T | AIV M Gene | AIV P-T | AIV S-T | AIV M Gene | AIV P-T | AIV S-T |
| VF-1 (20-11-12) | C1_ET | M | J | Pos. | HPAI | H5Nx | - | n.d. | n.d. | - | n.d. | n.d. |
| VF-2 (20-11-20) | C2_ET | F | A | - | n.d. | n.d. | Pos. | LPAI | H5Nx | - | n.d. | n.d. |
| VF-3 (20-11-27) | C3_ET | M | J | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. |
| C4_ET | M | J | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. | |
| VF-4 (20-12-04) | C5_ET | F | J | Pos. | HPAI | H5Nx | - | n.d. | n.d. | - | n.d. | n.d. |
| C6_ET | M | J | Pos. | HPAI | H5Nx | - | n.d. | n.d. | - | n.d. | n.d. | |
| C7_ET | F | J | Pos. | HPAI | H5Nx | - | n.d. | n.d. | - | n.d. | n.d. | |
| C8_ET | F | J | Pos. | HPAI | H5Nx | Pos. | HPAI | H5Nx | - | n.d. | n.d. | |
| C9_ET | F | J | Pos. | HPAI | H5Nx | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| C10_ET | F | A | Pos. | LPAI | HxNx | - | n.d. | n.d. | - | n.d. | n.d. | |
| C11_ET | F | A | Pos. | HPAI | H5Nx | - | n.d. | n.d. | - | n.d. | n.d. | |
| C12_ET | M | J | Pos. | HPAI | H5N8 | Pos. | HPAI | H5Nx | - | n.d. | n.d. | |
| C13_ET | M | A | Pos. | HPAI | H5Nx | - | n.d. | n.d. | - | n.d. | n.d. | |
| C14_ET | F | J | Pos. | HPAI | H5Nx | - | n.d. | n.d. | - | n.d. | n.d. | |
| C15_ET | M | J | Pos. | HPAI | H5N8 | Pos. | HPAI | H5Nx | - | n.d. | n.d. | |
| C16_ET | F | J | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. | |
| C17_ET | F | J | Pos. | HPAI | H5Nx | - | n.d. | n.d. | - | n.d. | n.d. | |
| C18_ET | F | J | Pos. | HPAI | H5Nx | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| C19_ET | F | J | Pos. | HPAI | H5Nx | - | n.d. | n.d. | - | n.d. | n.d. | |
| C20_ET | M | A | Pos. | HPAI | H5N1 | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| C21_ET | M | J | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| C22_ET | F | J | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| C23_ET | M | J | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| C24_ET | F | A | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| C25_ET | F | J | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| C26_ET | F | J | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5Nx | |
| VF-5 (20-12-11) | C27_ET | M | J | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. |
| C28_ET | F | J | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. | |
| C29_ET | F | J | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. | |
| C30_ET | F | J | Pos. | HPAI | H5N1-N8 | - | n.d. | n.d. | - | n.d. | n.d. | |
| C31_ET | F | A | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. | |
| C32_ET | F | J | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. | |
| C33_ET | F | J | Pos. | n.a. | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. | |
| C34_ET | F | J | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | HPAI | H5N8 | |
| VF-6 (20-12-18) | C35_ET | F | J | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. |
| C36_ET | M | J | Pos. | HPAI | H5N8 | - | n.d. | n.d. | - | n.d. | n.d. | |
| C37_ET | F | J | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. | |
| VF-8 (21-01-07) | C38_ET | F | J | - | n.d. | n.d. | Pos. | LPAI | H7N3 | - | n.d. | n.d. |
| VF-9 (21-01-14) | C39_Ma | F | J | - | n.d. | n.d. | - | n.d. | n.d. | Pos. | LPAI | H9Nx |
| VF-10 (21-01-22) | C40_ET | F | J | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. |
| C41_ET | F | J | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. | |
| VF-11 (21-01-29) | C42_ET | M | J | Pos. | LPAI | HxNx | - | n.d. | n.d. | - | n.d. | n.d. |
| C43_ET | F | J | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. | |
| C44_ET | M | J | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. | |
| C45_ET | F | J | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. | |
| C46_Ma | M | J | Pos. | LPAI | HxNx | - | n.d. | n.d. | - | n.d. | n.d. | |
| VF-13 (21-02-25) | C47_ET | F | J | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. |
| C48_ET | M | J | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. | |
| VCa-1 (21-02-12) | C49_CSh | F | J | - | n.d. | n.d. | Pos. | LPAI | H5Nx | - | n.d. | n.d. |
| C50_NP | A | M | - | n.d. | n.d. | Pos. | LPAI | H5Nx | - | n.d. | n.d. | |
| C51_NP | A | F | - | n.d. | n.d. | Pos. | LPAI | H5Nx | - | n.d. | n.d. | |
| C52_NP | J | F | - | n.d. | n.d. | Pos. | LPAI | H5Nx | - | n.d. | n.d. | |
| VCa-1 (21-02-15) | C53_NP | n.a. | n.a. | - | n.d. | n.d. | Pos. | LPAI | HxNx | - | n.d. | n.d. |
| HPAIV Antigen | HI Titers in ELISA-Positive Ducks | Duck Species (Wetland) | |
|---|---|---|---|
| Eurasian Teal (VF) | Eurasian Wigeon (VD) | ||
| H5N8/2020 (*) Clade 2.3.4.4b | <1:16 | 93 | 11 |
| 1:16 | 0 | 2 | |
| 1:32 | 2 | 1 | |
| 1:64 | 2 | 0 | |
| 1:128 | 0 | 0 | |
| seroprevalence (%) | 4/138 (2.9%) | 3/18 (16.7%) | |
| H5N5/2016 (§) Clade 2.3.4.4b | <1:16 | 93 | 12 |
| 1:16 | 2 | 0 | |
| 1:32 | 1 | 1 | |
| 1:64 | 1 | 1 | |
| 1:128 | 0 | 0 | |
| seroprevalence (%) | 4/138 (2.9%) | 2/18 (11.1%) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gobbo, F.; Fornasiero, D.; De Marco, M.A.; Zecchin, B.; Mulatti, P.; Delogu, M.; Terregino, C. Active Surveillance for Highly Pathogenic Avian Influenza Viruses in Wintering Waterbirds in Northeast Italy, 2020–2021. Microorganisms 2021, 9, 2188. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112188
Gobbo F, Fornasiero D, De Marco MA, Zecchin B, Mulatti P, Delogu M, Terregino C. Active Surveillance for Highly Pathogenic Avian Influenza Viruses in Wintering Waterbirds in Northeast Italy, 2020–2021. Microorganisms. 2021; 9(11):2188. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112188
Chicago/Turabian StyleGobbo, Federica, Diletta Fornasiero, Maria Alessandra De Marco, Bianca Zecchin, Paolo Mulatti, Mauro Delogu, and Calogero Terregino. 2021. "Active Surveillance for Highly Pathogenic Avian Influenza Viruses in Wintering Waterbirds in Northeast Italy, 2020–2021" Microorganisms 9, no. 11: 2188. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112188