
Multi-Omics Reveals the Inhibition of Lactiplantibacillus plantarum CCFM8724 in Streptococcus mutans-Candida albicans Mixed-Species Biofilms

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Bacterial Strains and Growth Conditions
2.3. Sample Preparation for Metabolomics and Transcriptomics Analysis
2.4. Untargeted Metabolomics by Gas Chromatography–Mass Spectrometry
2.5. RNA Extraction and Illumine Sequencing
2.6. DNA Extraction and RT-qPCR
2.7. Comparative Genomics and Enzyme Analysis
2.8. MultiStatistical Analysis and Visualization
3. Results
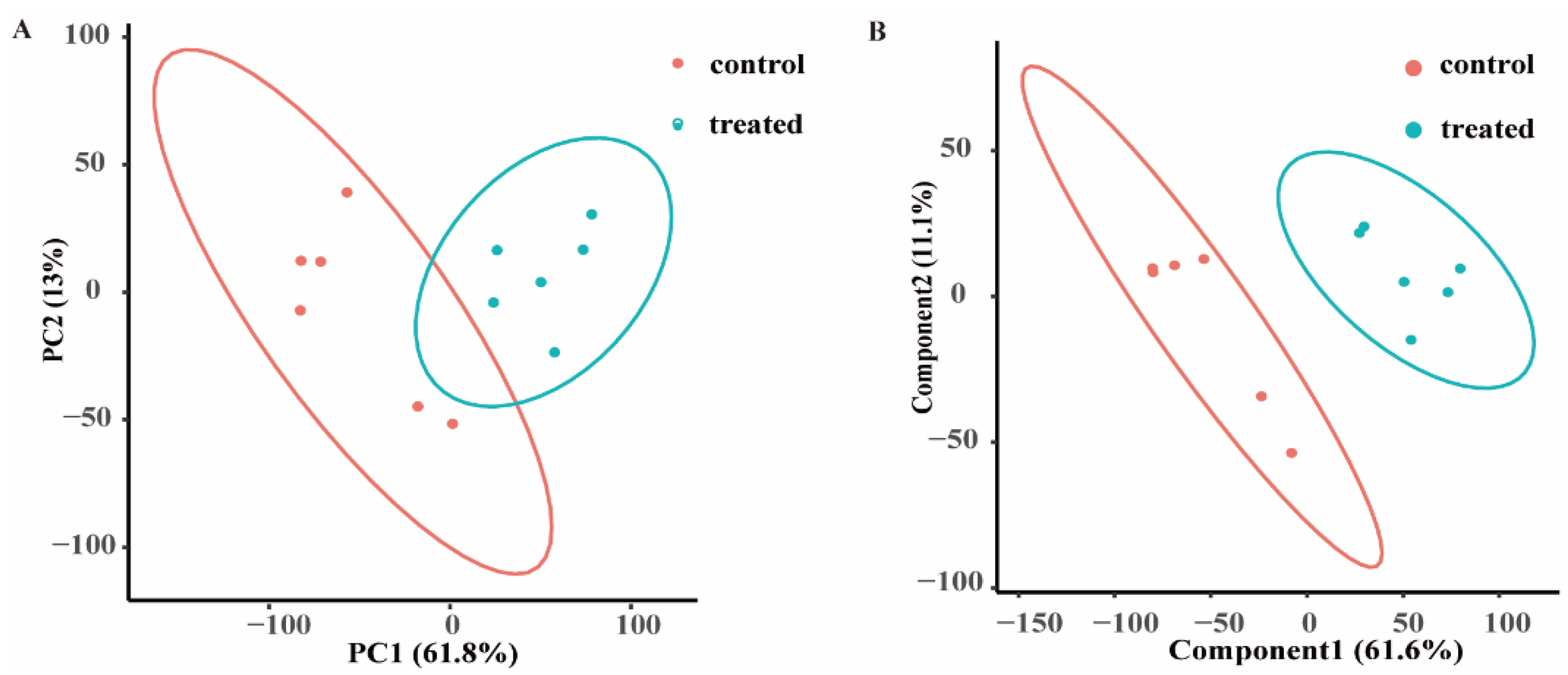
3.1. Analysis of Untargeted Metabolomic PCA and PLS-DA Data
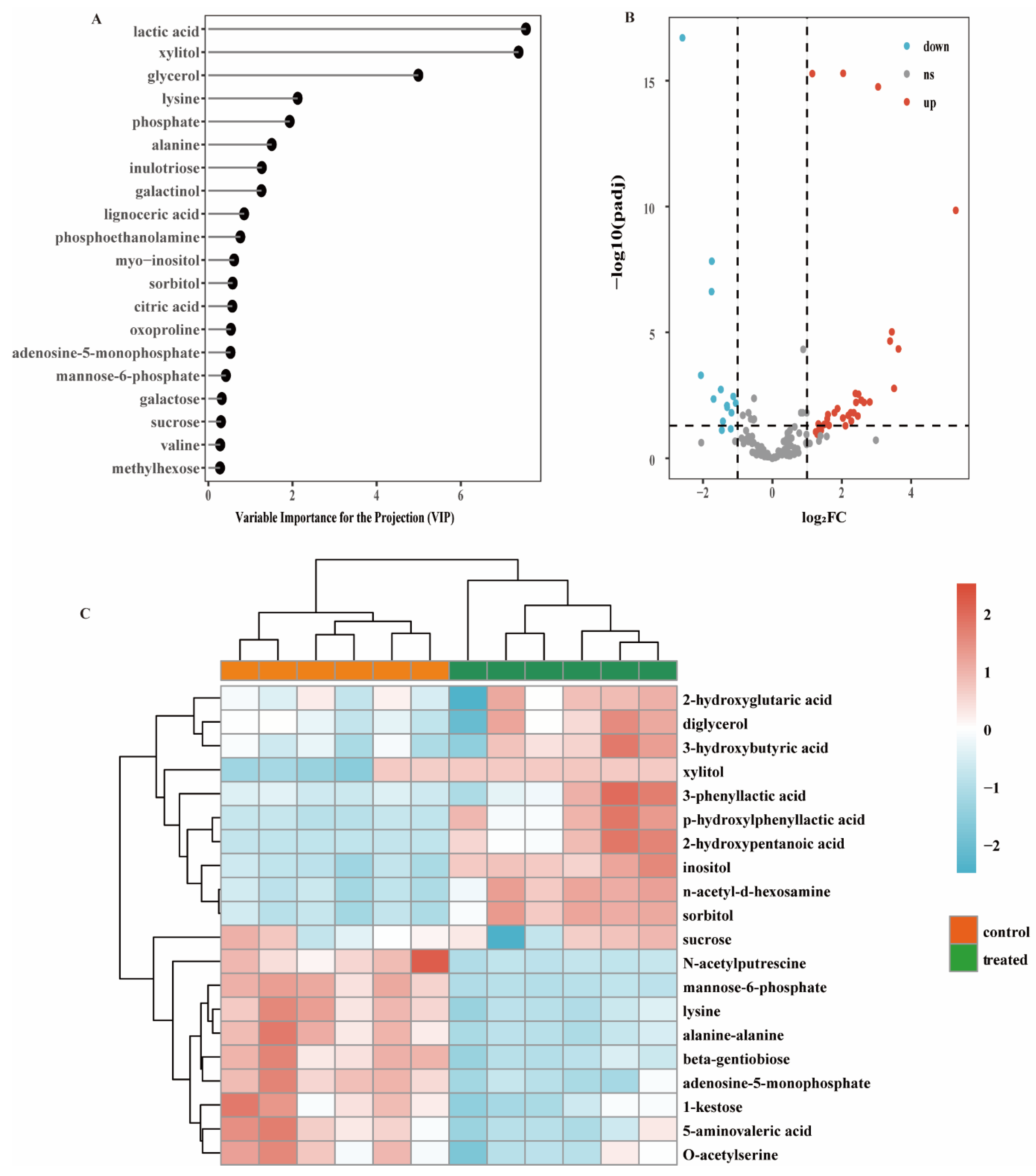
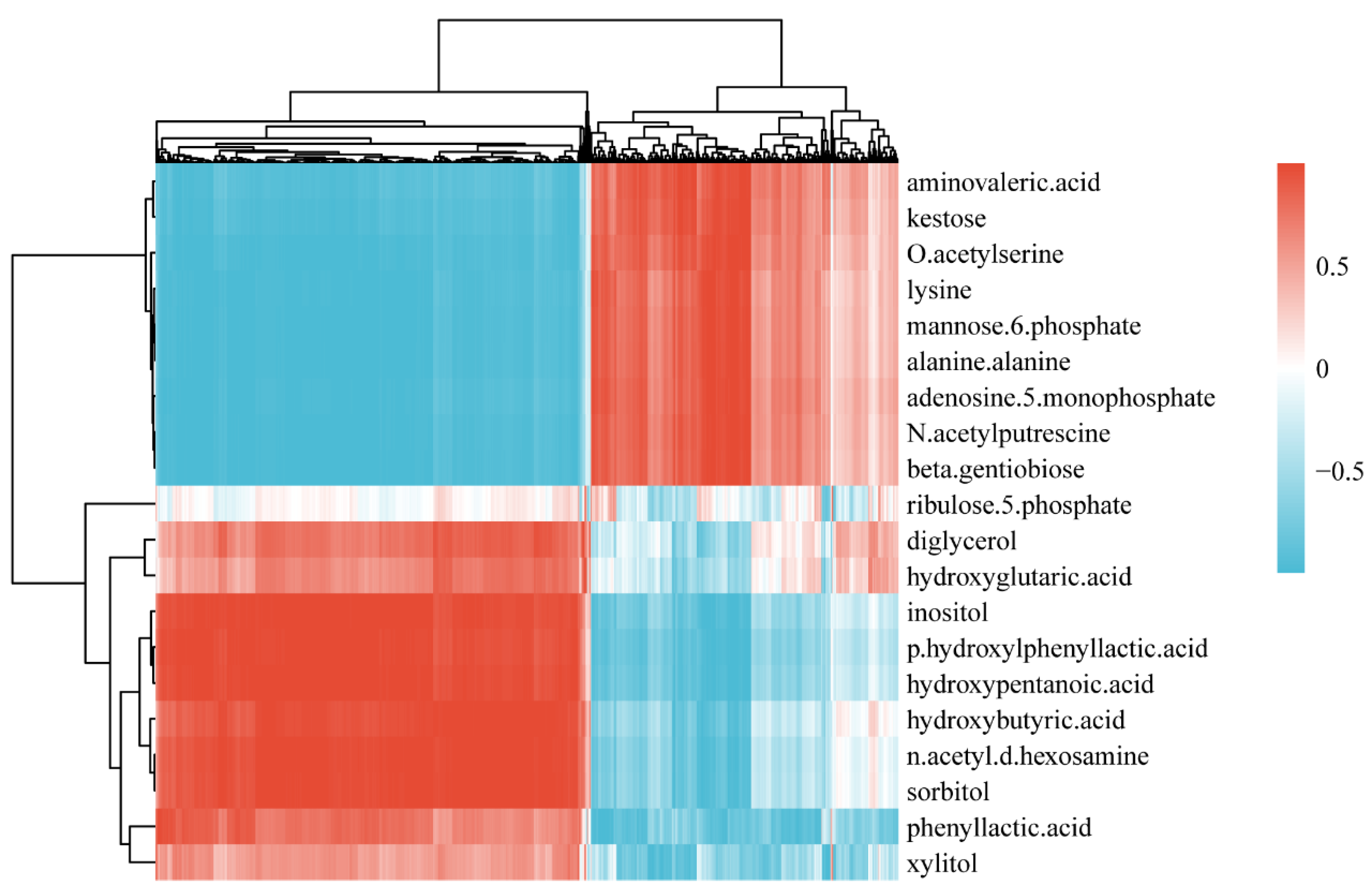
3.2. Differential Metabolites via Biofilm Profiling Analysis
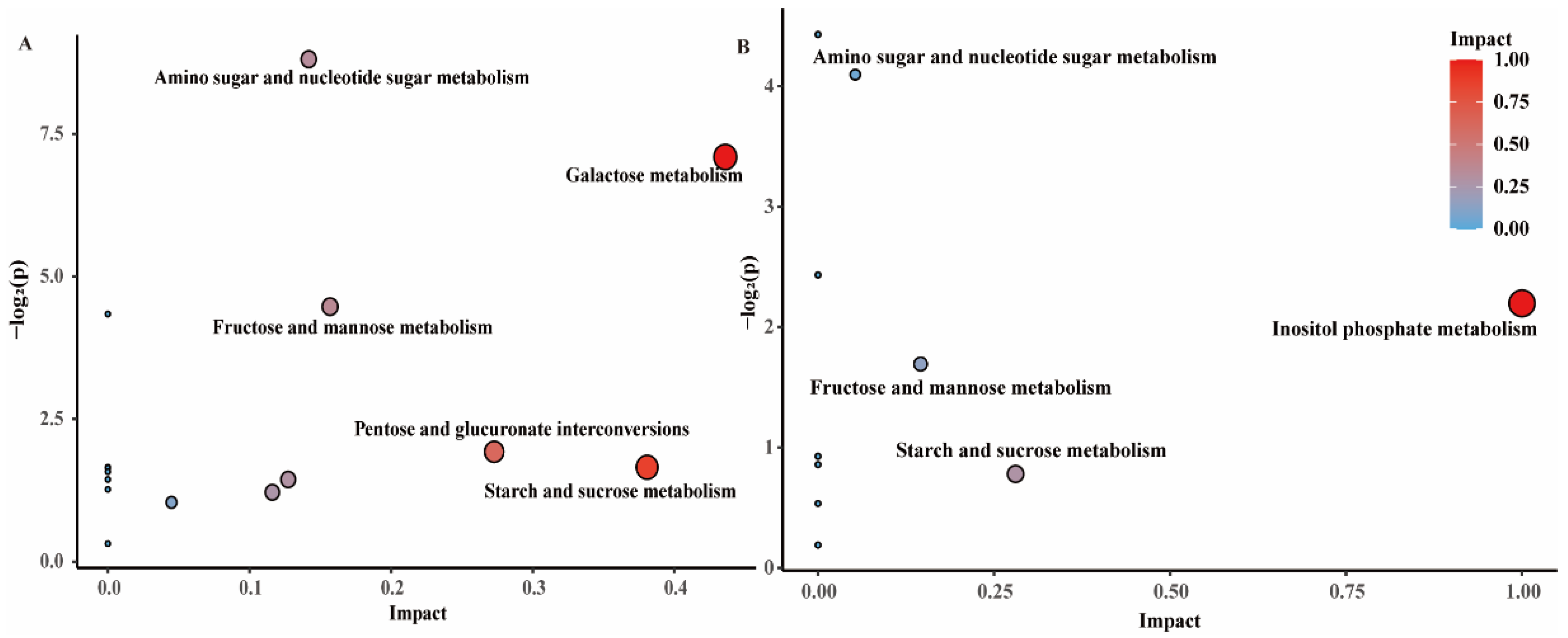
3.3. Metabolic Pathways
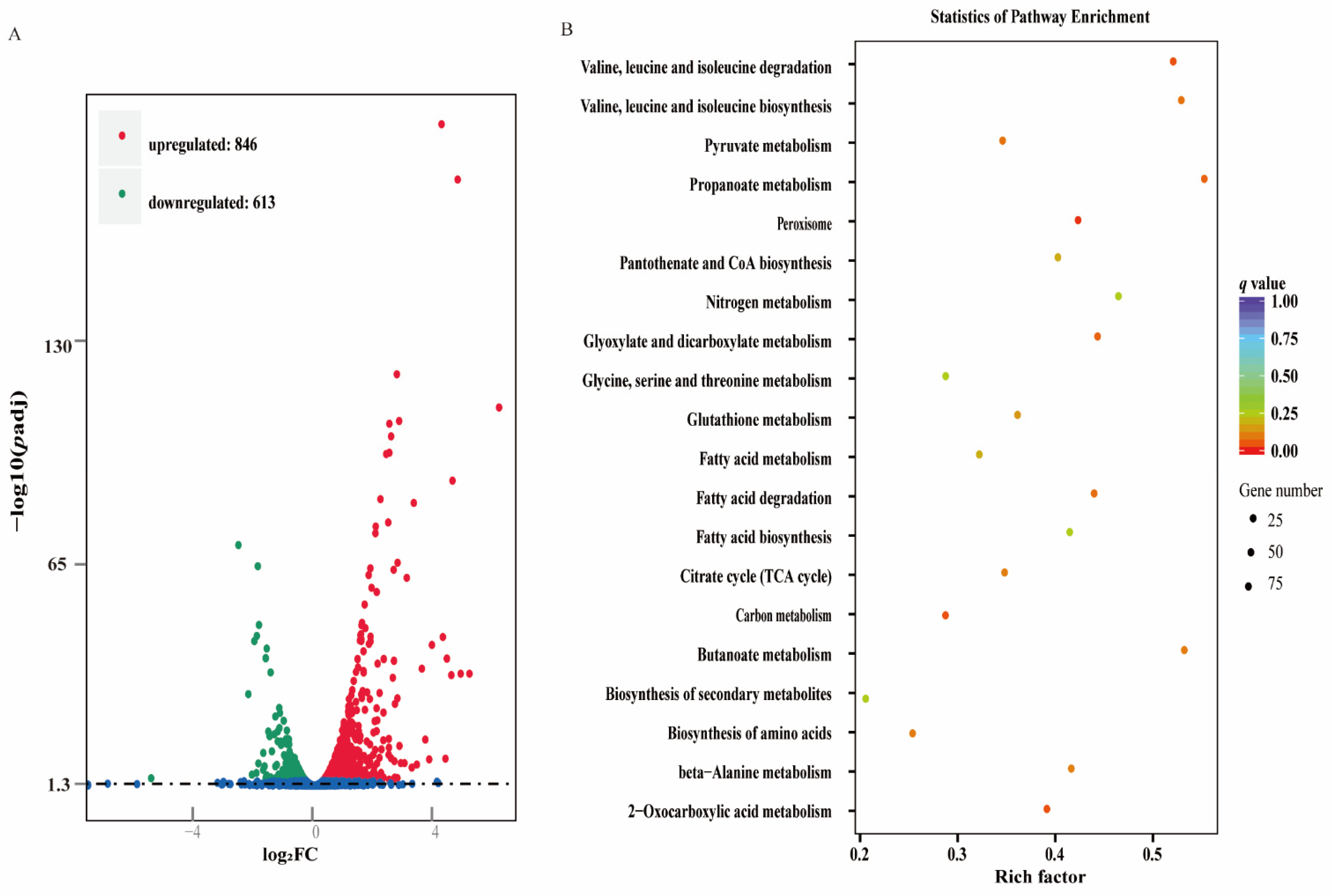
3.4. Global Changes at the Transcriptome Level
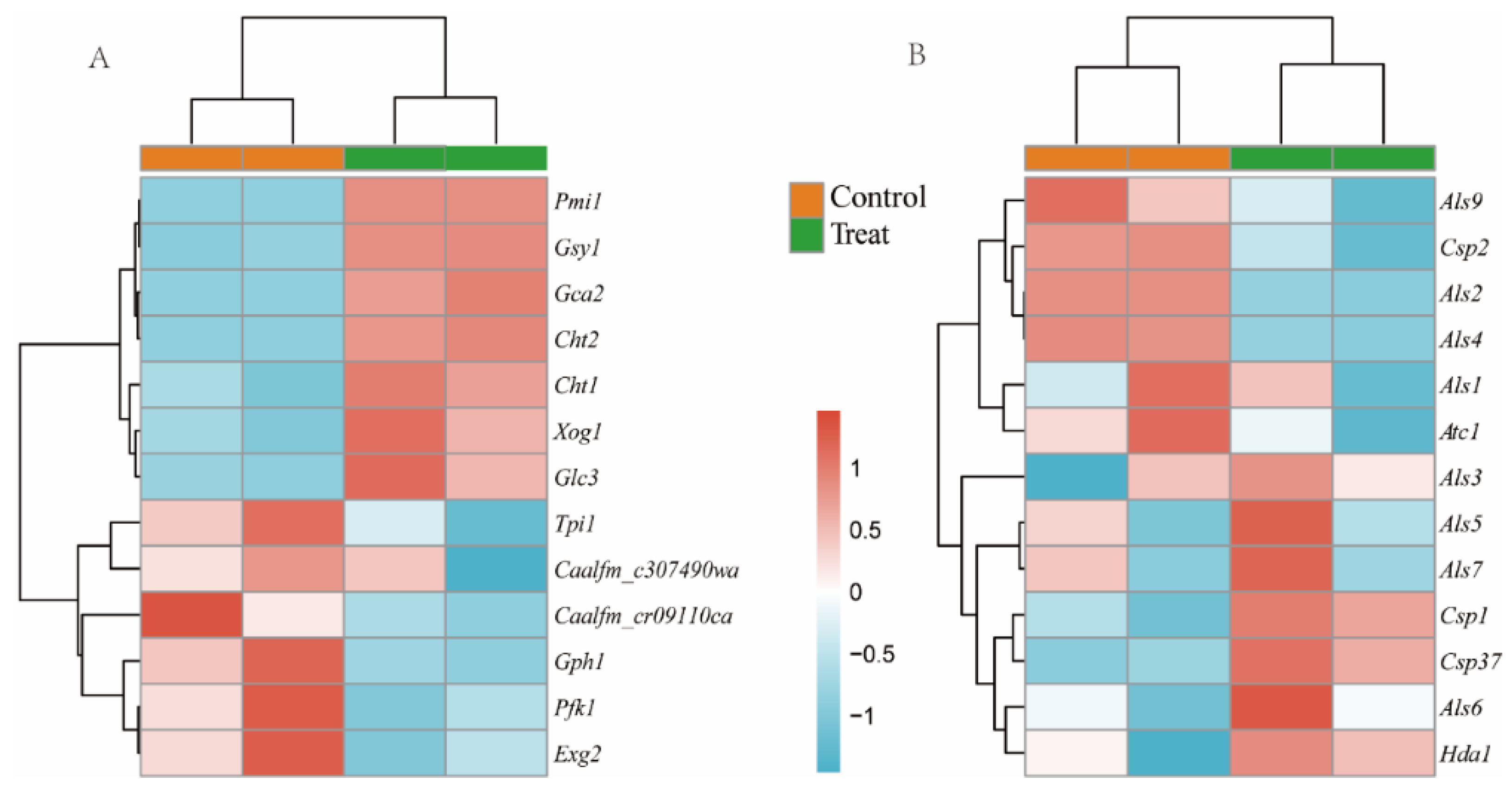
3.5. Differentially Expressed Genes (DEGs) between the Biofilms of the Treated and Control Groups
3.6. Integrated Metabolome and Transcriptome Analysis
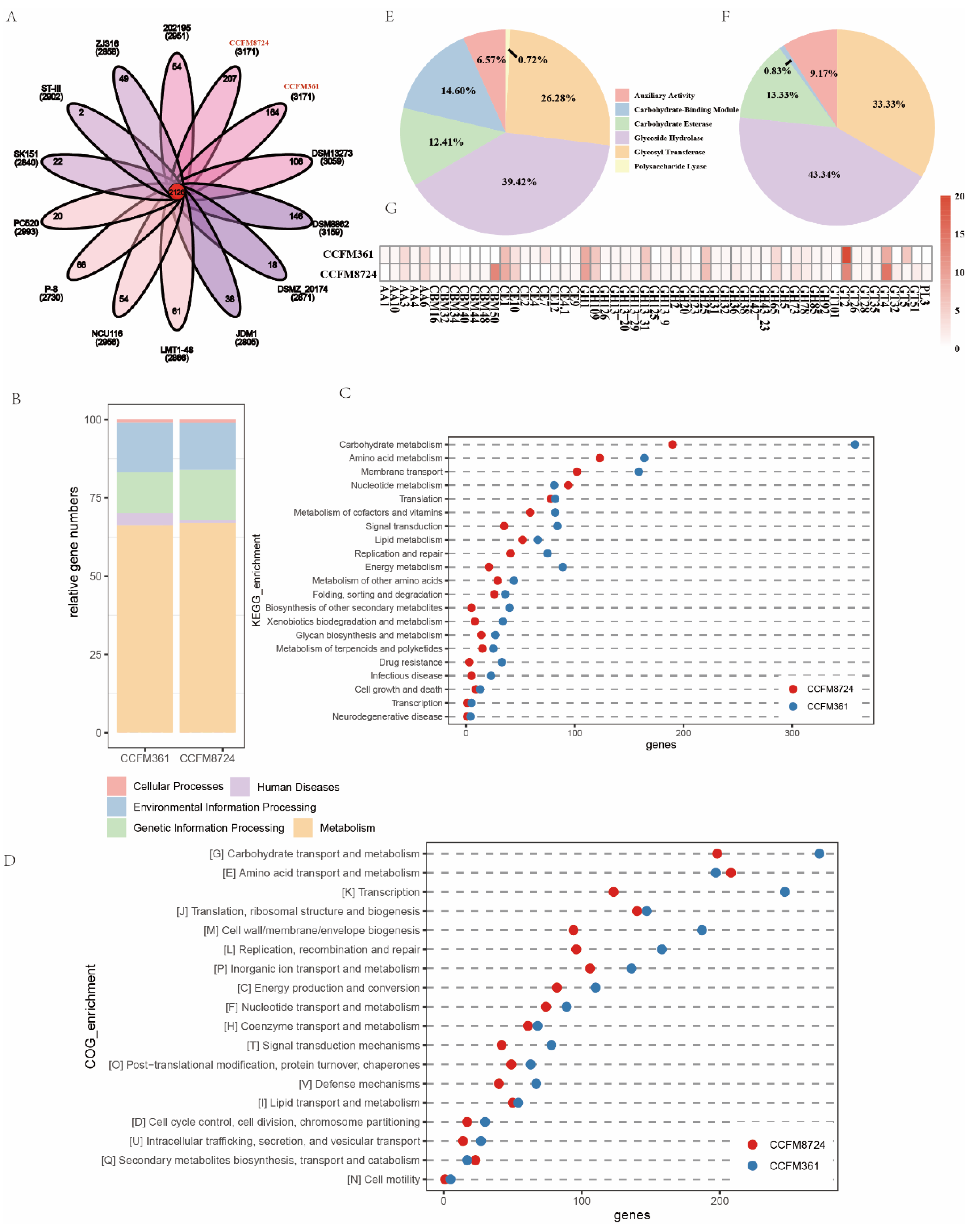
3.7. Genomics Analysis of L. plantarum CCFM8724
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mitrakul, K.; Srisatjaluk, R.; Srisukh, V.; Lomarat, P.; Vongsawan, K.; Kosanwat, T. Cymbopogon Citratus (Lemongrass Oil) Oral Sprays as Inhibitors of Mutans Streptococci Biofilm Formation. J. Clin. Diagn. Res. 2018, 12, 6–12. [Google Scholar] [CrossRef]
- Xiao, J.; Moon, Y.; Li, L.; Rustchenko, E.; Wakabayashi, H.; Zhao, X.; Feng, C.; Gill, S.R.; McLaren, S.; Malmstrom, H.; et al. Candida albicans Carriage in Children with Severe Early Childhood Caries (S-ECC) and Maternal Relatedness. PLoS ONE 2016, 11, e0164242. [Google Scholar] [CrossRef] [Green Version]
- Cugini, C.; Shanmugam, M.; Landge, N.; Ramasubbu, N. The Role of Exopolysaccharides in Oral Biofilms. J. Dent. Res. 2019, 98, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Ren, B.; He, J.; Peng, X.; Guo, Q.; Zheng, L.; Li, J.; Dai, H.; Chen, V.; Zhang, L.; et al. Candida albicans promotes tooth decay by inducing oral microbial dysbiosis. ISME J. 2021, 15, 894–908. [Google Scholar] [CrossRef]
- Krzyściak, W.; Kościelniak, D.; Papież, M.; Vyhouskaya, P.; Zagórska-Świeży, K.; Kołodziej, I.; Bystrowska, B.; Jurczak, A. Effect of a Lactobacillus Salivarius Probiotic on a Double-Species Streptococcus mutans and Candida albicans Caries Biofilm. Nutrients 2017, 9, 1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.-H.; Pan, T.-M. Inhibitory effect of Lactobacillus paracasei subsp. paracasei NTU 101 on rat dental caries. J. Funct. Foods 2014, 10, 223–231. [Google Scholar] [CrossRef]
- Hu, X.; Huang, Z.; Zhang, Y.; Hong, Y.; Zheng, Y. Effects of a probiotic drink containing Lactobacillus casei strain Shirota on dental plaque microbiota. J. Int. Med. Res. 2019, 47, 3190–3202. [Google Scholar] [CrossRef] [Green Version]
- Meto, A.; Colombari, B.; Odorici, A.; Giva, L.B.; Pericolini, E.; Regina, A.L. Antibacterial Effects of MicroRepair® BIOMA-Based Toothpaste and Chewing Gum on Orthodontic Elastics Contaminated In Vitro with Saliva from Healthy Donors: A Pilot Study. Appl. Sci. 2020, 10, 6721. [Google Scholar] [CrossRef]
- Chen, L.; Ren, Z.; Zhou, X.; Zeng, J.; Zou, J.; Li, Y. Inhibition of Streptococcus mutans biofilm formation, extracellular polysaccharide production, and virulence by an oxazole derivative. Appl. Microbiol. Biotechnol. 2016, 100, 857–867. [Google Scholar] [CrossRef]
- Abinaya, M.; Gayathri, M. Inhibition of biofilm formation, quorum sensing activity and molecular docking study of isolated 3, 5, 7-Trihydroxyflavone from Alstonia scholaris leaf against P.aeruginosa. Bioorganic Chem. 2019, 87, 291–301. [Google Scholar] [CrossRef]
- Srivastava, N.; Ellepola, K.; Venkiteswaran, N.; Chai, L.Y.A.; Ohshima, T.; Seneviratne, C.J. Lactobacillus Plantarum 108 Inhibits Streptococcus mutans and Candida albicans Mixed-Species Biofilm Formation. Antibiotics 2020, 9, 478. [Google Scholar] [CrossRef]
- Qin, S.; Xu, W.; Zhang, Q.; Zhao, J.; Zhang, H.; Chen, W. Inhibitory effect of Lactobacillus plantarum CCFM8724 on caries-causing dual biofilms. Food Ferment. Ind. 2020, 46, 127–132. [Google Scholar] [CrossRef]
- Zhang, Q.; Qin, S.; Xu, X.; Zhao, J.; Zhang, H.; Liu, Z.; Chen, W. Inhibitory Effect of Lactobacillus plantarum CCFM8724 towards Streptococcus mutans- and Candida albicans-Induced Caries in Rats. Oxidative Med. Cell. Longev. 2020, 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sadiq, F.A.; Yan, B.; Zhao, J.; Zhang, H.; Chen, W. Untargeted metabolomics reveals metabolic state of Bifidobacterium bifidum in the biofilm and planktonic states. LWT 2019, 118, 108772. [Google Scholar] [CrossRef]
- Zhao, N.; Zhang, C.; Yang, Q.; Guo, Z.; Yang, B.; Lu, W.; Li, D.; Tian, F.; Liu, X.; Zhang, H.; et al. Selection of Taste Markers Related to Lactic Acid Bacteria Microflora Metabolism for Chinese Traditional Paocai: A Gas Chromatography–Mass Spectrometry-Based Metabolomics Approach. J. Agric. Food Chem. 2016, 64, 2415–2422. [Google Scholar] [CrossRef]
- Lu, H.; Chen, H.; Tang, X.; Yang, Q.; Zhang, H.; Chen, Y.Q.; Chen, W. Evaluation of metabolome sample preparation and extraction methodologies for oleaginous filamentous fungi Mortierella alpina. Metabolomics 2019, 15, 50. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Wang, Q.; Wu, H.; Xia, S.; Guo, H.; Blaženović, I.; Zhang, Y.; Sun, X. Insights into cellular metabolic pathways of the combined toxicity responses of Caco-2 cells exposed to deoxynivalenol, zearalenone and Aflatoxin B1. Food Chem. Toxicol. 2019, 126, 106–112. [Google Scholar] [CrossRef]
- Zhou, T.; Luo, X.; Yu, C.; Zhang, C.; Zhang, L.; Song, Y.-B.; Dong, M.; Shen, C. Transcriptome analyses provide insights into the expression pattern and sequence similarity of several taxol biosynthesis-related genes in three Taxus species. BMC Plant Biol. 2019, 19, 33. [Google Scholar] [CrossRef] [Green Version]
- Cui, F.-X.; Zhang, R.-M.; Liu, H.-Q.; Wang, Y.-F.; Li, H. Metabolic responses to Lactobacillus plantarum contamination or bacteriophage treatment in Saccharomyces cerevisiae using a GC–MS-based metabolomics approach. World J. Microbiol. Biotechnol. 2015, 31, 2003–2013. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Wang, C.; Li, Y.; Bao, X.; Zhang, Y.; Tang, T. The Impact of SlyA on Cell Metabolism of Salmonella typhimurium: A Joint Study of Transcriptomics and Metabolomics. J. Proteome Res. 2021, 20, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Buu, L.; Lee, F.S.; Chen, Y. Temperature and Glucose Level Are Important for Hypha Development and Saps Expression in Candida albicans and the Sap6 Is an Important Virulent Factor in Co-culture of C. albicans and THP-1 Human Mononuclear Cells. In Molecular Biology of the Cell; Amer Soc Cell Biology: Bethesda, MD, USA, 2006; Volume 17. [Google Scholar]
- Leme, A.P.; Koo, H.; Bellato, C.M.; Bedi, G.; Cury, J.A. The Role of Sucrose in Cariogenic Dental Biofilm Formation—New Insight. J. Dent. Res. 2006, 85, 878–887. [Google Scholar] [CrossRef]
- Cai, J.-N.; Jung, J.-E.; Lee, M.-H.; Choi, H.-M.; Jeon, J.-G. Sucrose challenges to Streptococcus mutans biofilms and the curve fitting for the biofilm changes. FEMS Microbiol. Ecol. 2018, 94, 29788432. [Google Scholar] [CrossRef] [Green Version]
- Falsetta, M.L.; Klein, M.; Colonne, P.M.; Scott-Anne, K.; Gregoire, S.; Pai, C.-H.; Gonzalez-Begne, M.; Watson, G.; Krysan, D.; Bowen, W.H.; et al. Symbiotic Relationship between Streptococcus mutans and Candida albicans Synergizes Virulence of Plaque BiofilmsIn Vivo. Infect. Immun. 2014, 82, 1968–1981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellepola, K.; Truong, T.; Liu, Y.; Lin, Q.; Lim, T.K.; Lee, Y.M.; Cao, T.; Koo, H.; Seneviratne, C.J. Multi-omics Analyses Reveal Synergistic Carbohydrate Metabolism in Streptococcus mutans-Candida albicans Mixed-Species Biofilms. Infect. Immun. 2019, 87, e00339-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowen, W.; Koo, H. Biology of Streptococcus mutans-Derived Glucosyltransferases: Role in Extracellular Matrix Formation of Cariogenic Biofilms. Caries Res. 2011, 45, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef]
- Burt, B.A. The use of sorbitol- and xylitol-sweetened chewing gum in caries control. J. Am. Dent. Assoc. 2006, 137, 190–196. [Google Scholar] [CrossRef] [Green Version]
- Chi, D.L.; Zegarra, G.; Huerta, E.C.V.; Castillo, J.L.; Milgrom, P.; Roberts, M.C.; Matta, A.C.; Merino, A.P.; Mancl, L. Milk Sweetened with Xylitol: A Proof-of-Principle Caries Prevention Randomized Clinical Trial. J. Dent. Child. 2016, 83, 152–160. [Google Scholar]
- Takahashi-Abbe, S.; Abbe, K.; Takahashi, N.; Tamazawa, Y.; Yamada, T. Inhibitory effect of sorbitol on sugar metabolism of Streptococcus mutans in vitro and on acid production in dental plaque in vivo. Oral Microbiol. Immunol. 2001, 16, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Lemos, J.A.; Palmer, S.R.; Zeng, L.; Wen, Z.T.; Kajfasz, J.K.; Freires, I.A.; Abranches, J.; Brady, L.J. The Biology of Streptococcus mutans. Microbiol. Spectr. 2019, 7, 7. [Google Scholar] [CrossRef]
- Cornejo, O.E.; Lefébure, T.; Bitar, P.D.P.; Lang, P.; Richards, V.; Eilertson, K.; Do, T.; Beighton, D.; Zeng, L.; Ahn, S.-J.; et al. Evolutionary and Population Genomics of the Cavity Causing Bacteria Streptococcus mutans. Mol. Biol. Evol. 2013, 30, 881–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.; Mao, Y.; Guo, H.; Zhao, Y.; Deng, Y. Analysis and identification of main antibacterial metabolites secreted by Lactobacillus plantarum DY6. Microbiol. China 2019, 46, 2258–2271. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Q.; Guo, M.; Zhao, J. Identification of metabolites secreted by Lactobacillus plantarum CCFM8724 on inhibiting dual-species biofilm. Microbiol. China 2021. [Google Scholar] [CrossRef]
- James, K.M.; Macdonald, K.W.; Chanyi, R.M.; Cadieux, P.A.; Burton, J. Inhibition of Candida albicans biofilm formation and modulation of gene expression by probiotic cells and supernatant. J. Med. Microbiol. 2016, 65, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Dehullu, J.; Valotteau, C.; Herman-Bausier, P.; Garcia-Sherman, M.; Mittelviefhaus, M.; Vorholt, J.A.; Lipke, P.N.; Dufrêne, Y.F. Fluidic Force Microscopy Demonstrates That Homophilic Adhesion by Candida albicans Als Proteins Is Mediated by Amyloid Bonds between Cells. Nano Lett. 2019, 19, 3846–3853. [Google Scholar] [CrossRef] [PubMed]
- Montanaro, L.; Poggi, A.; Visai, L.; Ravaioli, S.; Campoccia, D.; Speziale, P.; Arciola, C.R. Extracellular DNA in Biofilms. Int. J. Artif. Organs 2011, 34, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; Bastiaanssen, T.F.S.; Song, L.; Jiang, B.; Zhang, X.; Zhao, J.; Zhang, H.; Chen, W.; Cryan, J.F.; Wang, G. Unraveling the Microbial Mechanisms Underlying the Psychobiotic Potential of a Bifidobacterium breve Strain. Mol. Nutr. Food Res. 2021, 65, 2000704. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | C1 | C2 | T1 | T2 |
|---|---|---|---|---|
| Total reads (C. albicans) | 5,5819,264 | 54,481,928 | 56,579,976 | 53,032,306 |
| Total mapped (C. albicans) | 36,144,061 (64.75%) | 35,817,034 (65.74%) | 48,178,839 (85.15%) | 44,747,300 (84.38%) |
| Total reads (S. mutans) | 55,506,496 | 54,041,988 | 56,174,376 | 52,584,766 |
| Total mapped (S. mutans) | 13,877,006 (25%) | 12,460,931 (23.06%) | 18,215 (0.03%) | 45,311 (0.08%) |
| Organisms | Cq | Cells | ||
|---|---|---|---|---|
| C | T | C | T | |
| S. mutans | 15.18 a | 24.36 b | 109 a | 106 b |
| C. albicans | 16.15 a | 16.22 a | 1010 a | 1010 a |
| Compounds | Correlated DEGs |
|---|---|
| Mannose-6-phosphate | 131 |
| Kestose | 158 |
| Lysine | 127 |
| Adenosine-5-monophosphate | 140 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Li, J.; Lu, W.; Zhao, J.; Zhang, H.; Chen, W. Multi-Omics Reveals the Inhibition of Lactiplantibacillus plantarum CCFM8724 in Streptococcus mutans-Candida albicans Mixed-Species Biofilms. Microorganisms 2021, 9, 2368. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112368
Zhang Q, Li J, Lu W, Zhao J, Zhang H, Chen W. Multi-Omics Reveals the Inhibition of Lactiplantibacillus plantarum CCFM8724 in Streptococcus mutans-Candida albicans Mixed-Species Biofilms. Microorganisms. 2021; 9(11):2368. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112368
Chicago/Turabian StyleZhang, Qiuxiang, Jiaxun Li, Wenwei Lu, Jianxin Zhao, Hao Zhang, and Wei Chen. 2021. "Multi-Omics Reveals the Inhibition of Lactiplantibacillus plantarum CCFM8724 in Streptococcus mutans-Candida albicans Mixed-Species Biofilms" Microorganisms 9, no. 11: 2368. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112368