
Amplifying and Fine-Tuning Rsm sRNAs Expression and Stability to Optimize the Survival of Pseudomonas brassicacerum in Nutrient-Poor Environments

 , , and
, , and
Abstract
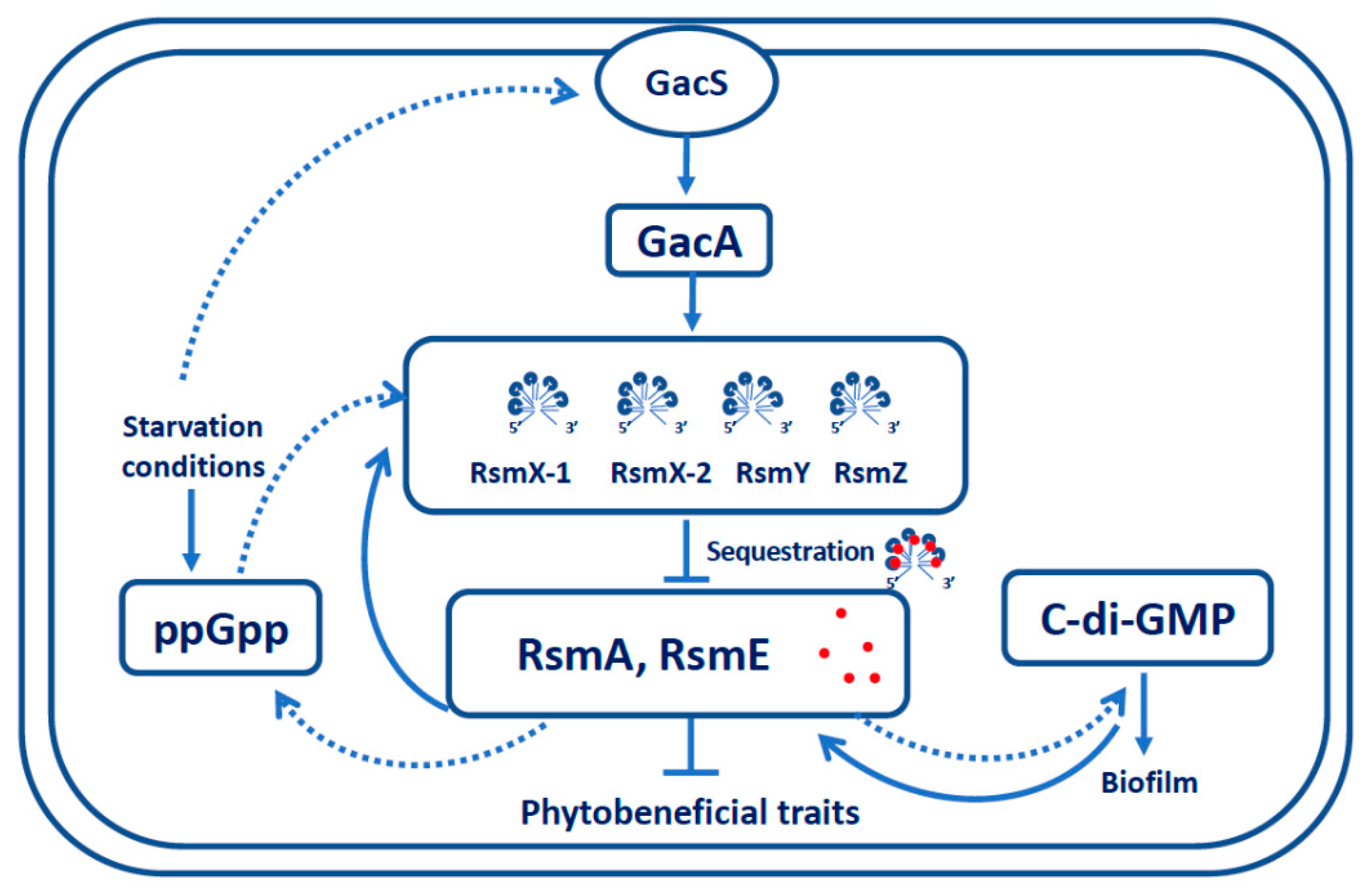
:1. Introduction
2. Experimental Procedures
2.1. Bacterial Strains, Plasmids and Growth Conditions
2.2. DNA Manipulation
2.3. RNA Manipulation
2.4. 5′ RACE
2.5. Overexpression of sRNAs
2.6. Construction of Transcriptional lacZ Fusions
2.7. Northern Blot Analysis
2.8. Protease Activity Assays
2.9. β-Galactosidase Assays
2.10. Biofilm Assays
2.11. Stability of Rsm sRNAs
2.12. High Resolution Mass Spectrometry Analysis
2.13. MassTRIX Metabolite Annotation
3. Results
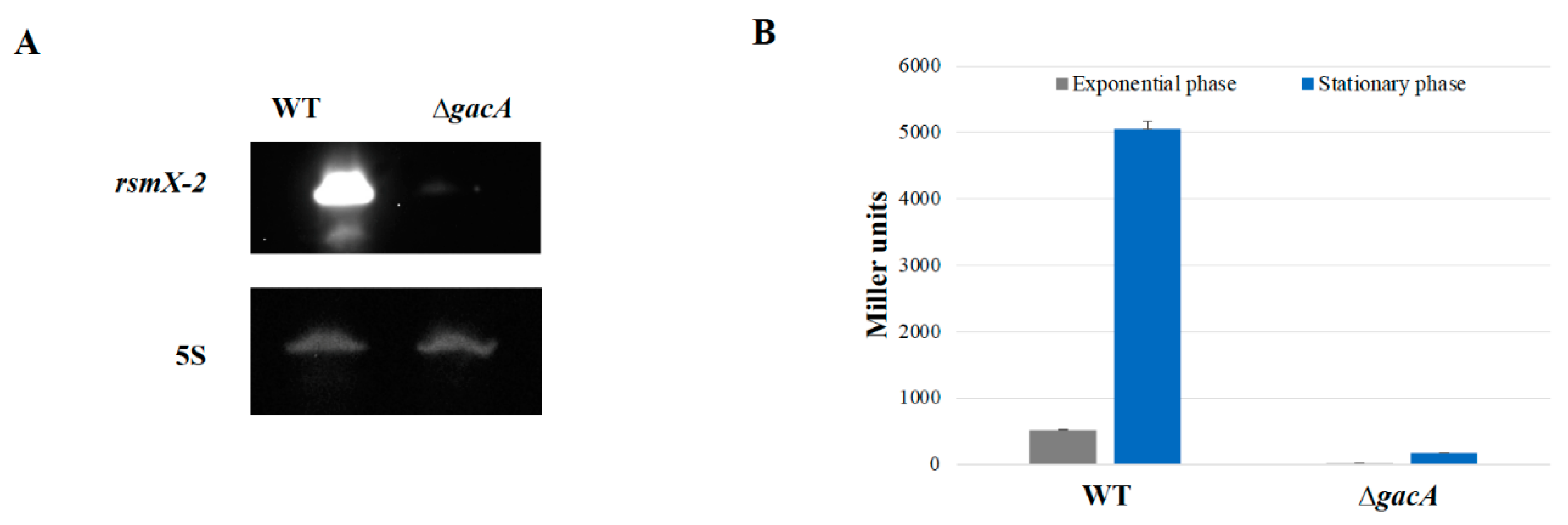
3.1. Identification of a Fourth Rsm sRNA in P. brassicacearum
3.2. rsmX-2 Expression Is Exclusively GacA-Dependent
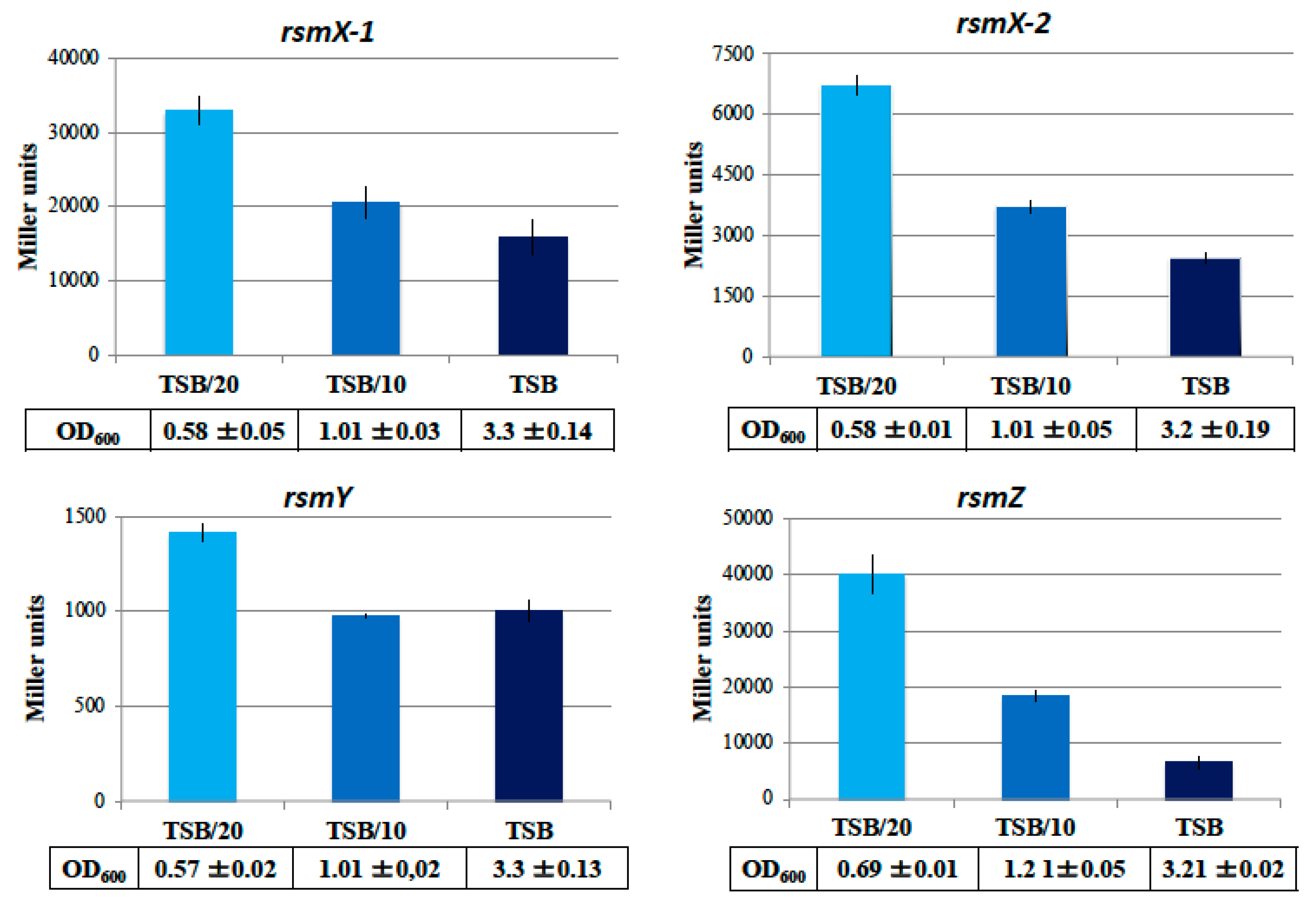
3.3. Stringent Conditions Activate rsm sRNAs Genes Expression
3.4. Stability of Rsm sRNA
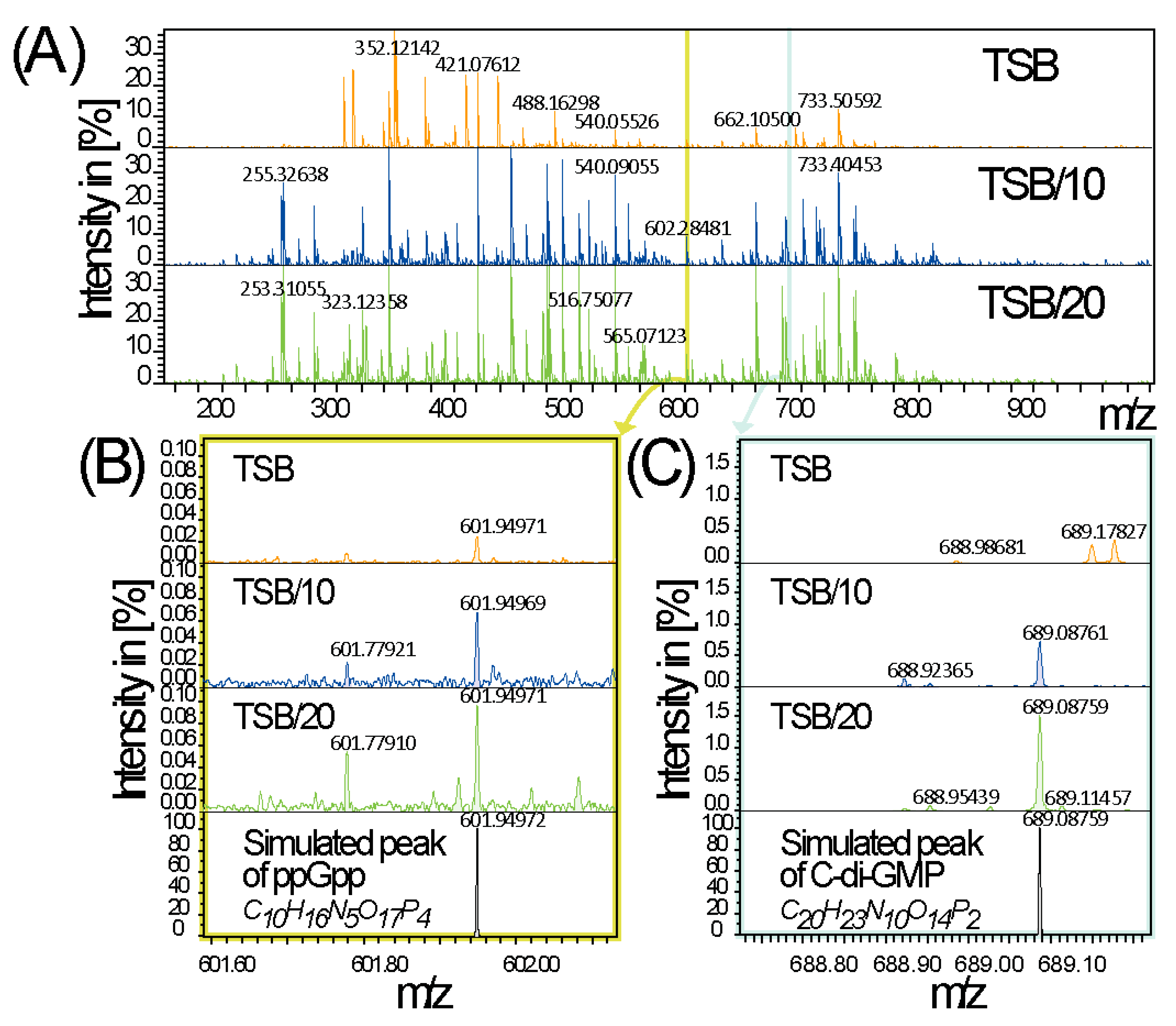
3.5. Stringent Conditions and Sedentary Lifestyle Signalling Molecules
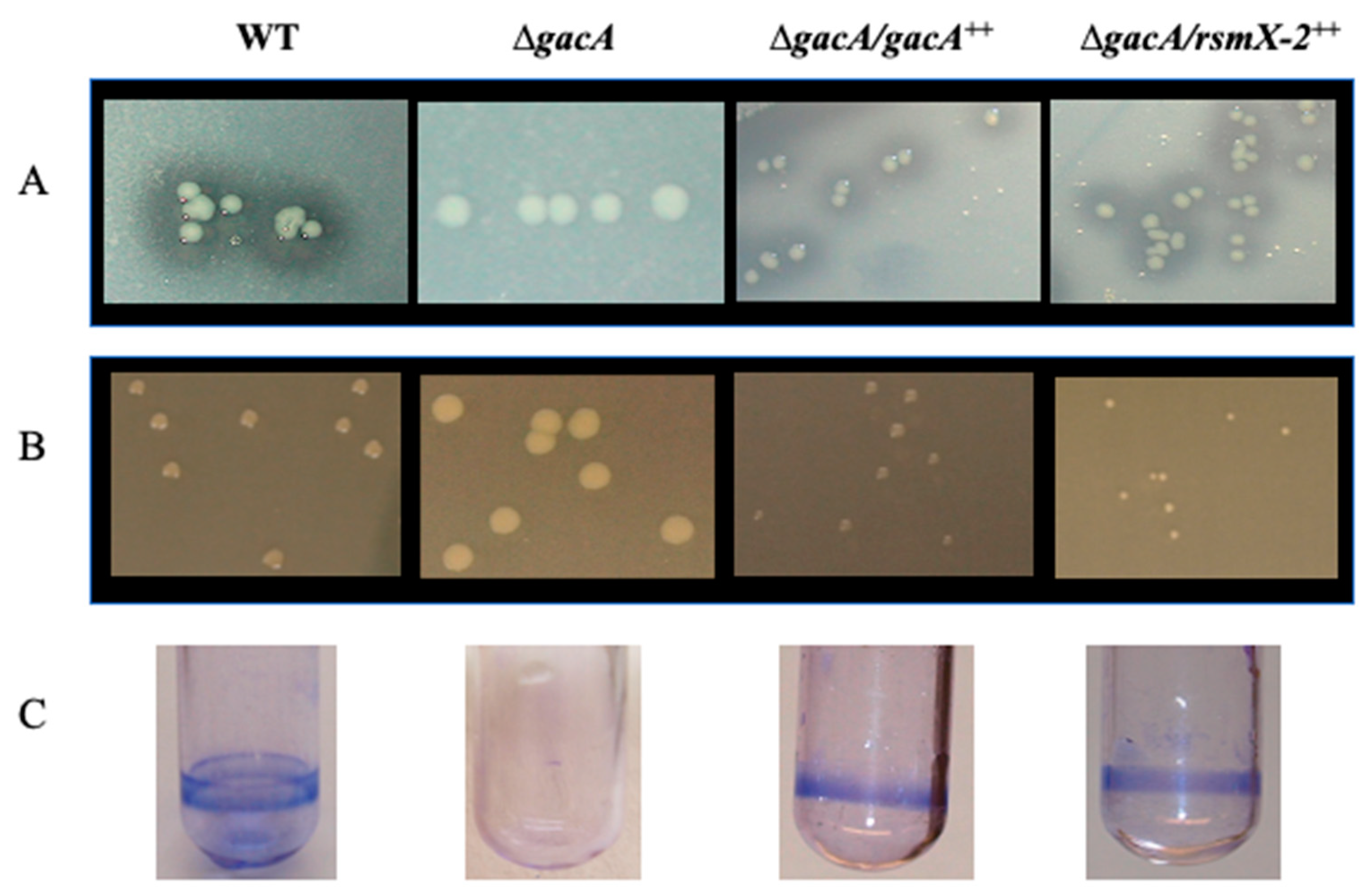
4. Discussion
4.1. P. brassicacearum Rsm sRNA Amplification by a Duplication of rsmX
4.2. RsmX-2 Is Part of Gac-Rsm System under Exclusive Control of GacA
4.3. Regulation of the Amount of RNA by Degradation
4.4. Modulation of Rsm sRNAs Expression in Response to Nutrient-Poor Conditions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Romeo, T. Global regulation by the small RNA-binding protein CsrA and the non-coding RNA molecule CsrB. Mol. Microbiol. 1998, 29, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Fortune, D.R.; Suyemoto, M.; Altier, C. Identification of CsrC and characterization of its role in epithelial cell invasion in Salmonella enterica serovar Typhimurium. Infect. Immun. 2006, 74, 331–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenz, D.H.; Miller, M.B.; Zhu, J.; Kulkarni, R.V.; Bassler, B.L. CsrA and three redundant small RNAs regulate quorum sensing in Vibrio cholerae. Mol. Microbiol. 2005, 58, 1186–1202. [Google Scholar] [CrossRef] [PubMed]
- Sahr, T.; Bruggemann, H.; Jules, M.; Lomma, M.; Albert-Weissenberger, C.; Cazalet, C.; Buchrieser, C. Two small ncRNAs jointly govern virulence and transmission in Legionella pneumophila. Mol. Microbiol. 2009, 72, 741–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heroven, A.K.; Bohme, K.; Rohde, M.; Dersch, P. A Csr-type regulatory system, including small non-coding RNAs, regulates the global virulence regulator RovA of Yersinia pseudotuberculosis through RovM. Mol. Microbiol. 2008, 68, 1179–1195. [Google Scholar] [CrossRef] [PubMed]
- Kay, E.; Dubuis, C.; Haas, D. Three small RNAs jointly ensure secondary metabolism and biocontrol in Pseudomonas fluorescens CHA0. Proc. Natl. Acad. Sci. USA 2005, 102, 17136–17141. [Google Scholar] [CrossRef] [Green Version]
- Kay, E.; Humair, B.; Denervaud, V.; Riedel, K.; Spahr, S.; Eberl, L.; Valverde, C.; Haas, D. Two GacA-dependent small RNAs modulate the quorum-sensing response in Pseudomonas aeruginosa. J. Bacteriol. 2006, 188, 6026–6033. [Google Scholar] [CrossRef] [Green Version]
- Moll, S.; Schneider, D.J.; Stodghill, P.; Myers, C.R.; Cartinhour, S.W.; Filiatrault, M.J. Construction of an rsmX co-variance model and identification of five rsmX non-coding RNAs in Pseudomonas syringae pv. tomato DC3000. RNA Biol. 2010, 7, 3–11. [Google Scholar] [CrossRef] [Green Version]
- White, D.; Hart, M.E.; Romeo, T. Phylogenetic distribution of the global regulatory gene csrA among eubacteria. Gene 1996, 182, 221–223. [Google Scholar] [CrossRef]
- Harfouche, L.; Haichar, F.Z.; Achouak, W. Small regulatory RNAs and the fine-tuning of plant-bacteria interactions. New Phytol. 2015, 206, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Sobrero, P.M.; Valverde, C. Comparative genomics and evolutionary analysis of Rna-binding proteins of the CsrA family in the genus pseudomonas. Front. Mol. Biosci. 2020, 7, 127. [Google Scholar] [CrossRef] [PubMed]
- Babitzke, P.; Romeo, T. CsrB sRNA family, sequestration of RNA-binding regulatory proteins. Curr. Opin. Microbiol. 2007, 10, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, M.J.; Kambara, T.K.; Ramsey, K.M.; Dove, S.L. Widespread targeting of nascent transcripts by RsmA in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2020, 117, 10520–10529. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Y.; Gui, G.; Wei, B.; Preston, J.F.; Oakford, L.; Yuksel, U.; Giedroc, D.P.; Romero, T. The RNA molecule CsrB binds to the global regulatory protein CsrA and antagonizes its activity in Escherichia coli. J. Biol. Chem. 1997, 272, 17502–17510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weilbacher, T.; Suzuki, K.; Dubey, A.K.; Wang, X.; Gudapaty, S.; Morozov, I.; Baker, C.S.; Georgellis, D.; Babitzke, P.; Romeo, T. A novel sRNA component of the carbon storage regulatory system of Escherichia coli. Mol. Microbiol. 2003, 48, 657–670. [Google Scholar] [CrossRef]
- Valverde, C. Artificial sRNAs activating the Gac/Rsm signal transduction pathway in Pseudomonas fluorescens. Arch. Microbiol. 2009, 191, 349–359. [Google Scholar] [CrossRef]
- Brencic, A.; McFarland, K.A.; McManus, H.R.; Castang, S.; Mogno, I.; Dove, S.L.; Lory, S. The GacS/GacA signal transduction system of Pseudomonas aeruginosa acts exclusively through its control over the transcription of the RsmY and RsmZ regulatory small RNAs. Mol. Microbiol. 2009, 73, 434–445. [Google Scholar] [CrossRef] [Green Version]
- Lalaouna, D.; Fochesato, S.; Sanchez, L.; Schmitt-Kopplin, P.; Haas, D.; Heulin, T.; Achouak, W. Phenotypic switching involves GacS- and GacA-dependent Rsm small RNAs in Pseudomonas brassicacearum. Appl. Environ. Microbiol. 2012, 78, 1658–1665. [Google Scholar] [CrossRef] [Green Version]
- Humair, B.; Wackwitz, B.; Haas, D. GacA-controlled activation of promoters for small RNA genes in Pseudomonas fluorescens. Appl. Environ. Microbiol. 2010, 76, 1497–1506. [Google Scholar] [CrossRef] [Green Version]
- Bordi, C.; Lamy, M.C.; Ventre, I.; Termine, E.; Hachani, A.; Fillet, S.; Roche, B.; Bleves, C.; Méjean, V.; Lazdunski, A.; et al. Regulatory RNAs and the HptB/RetS signalling pathways fine-tune Pseudomonas aeruginosa pathogenesis. Mol. Microbiol. 2010, 76, 1427–1443. [Google Scholar] [CrossRef] [Green Version]
- Achouak, W.; Sutra, L.; Heulin, T.; Meyer, J.M.; Fromin, N.; Degraeve, S.; Christen, R.; Gardan, L. Pseudomonas brassicacearum sp nov and Pseudomonas thivervalensis sp nov.; two root-associated bacteria isolated from Brassica napus and Arabidopsis thaliana. Int. J. Syst. Evol. Microbiol. 2000, 50, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achouak, W.; Conrod, S.; Cohen, V.; Heulin, T. Phenotypic variation of Pseudomonas brassicacearum as a plant root-colonization strategy. Mol. Plant-Microbe Interact. 2004, 17, 872–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achouak, W.; Conrod, S.; Robert, S.; Heulin, T. Phase variable effects of Pseudomonas brassicacearum on Arabidopsis thaliana root architecture. In Biology of Molecular Plant-Microbe Interactions; Tikhonovich, I., Lugtenberg, B., Provorov, N., Eds.; International Society for Molecular Plant-Microbe Interactions: St. Paul, MN, USA, 2004; Volume 4, pp. 359–363. [Google Scholar]
- Chabeaud, P.; de Groot, A.; Bitter, W.; Tommassen, J.; Heulin, T.; Achouak, W. Phase-variable expression of an operon encoding extracellular alkaline protease, a serine protease homolog, and lipase in Pseudomonas brassicacearum. J. Bacteriol. 2001, 183, 2117–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tillett, D.; Burns, B.P.; Neilan, B.A. Optimized rapid amplification of cDNA ends (RACE) for mapping bacterial mRNA transcripts. Biotechniques 2000, 28, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Heeb, S.; Blumer, C.; Haas, D. Regulatory RNA as mediator in GacA/RsmA-dependent global control of exoproduct formation in Pseudomonas fluorescens CHA0. J. Bacteriol. 2002, 184, 1046–1056. [Google Scholar] [CrossRef] [Green Version]
- Schnider-Keel, U.; Seematter, A.; Maurhofer, M.; Blumer, C.; Duffy, B.; Gigot-Bonnefoy, C.; Reimmann, C.; Notz, R.; Defago, G.; Haas, D.; et al. Autoinduction of 2,4-diacetylphloroglucinol biosynthesis in the biocontrol agent Pseudomonas fluorescens CHA0 and repression by the bacterial metabolites salicylate and pyoluteorin. J. Bacteriol. 2000, 182, 1215–1225. [Google Scholar] [CrossRef] [Green Version]
- Boisset, S.; Geissmann, T.; Huntzinger, E.; Fechter, P.; Bendridi, N.; Possedko, M.; Chevalier, C.; Helfer, A.C.; Benito, Y.; Jacquier, A.; et al. Staphylococcus aureus RNAIII coordinately represses the synthesis of virulence factors and the transcription regulator Rot by an antisense mechanism. Genes Dev. 2007, 21, 1353–1366. [Google Scholar] [CrossRef] [Green Version]
- Newell, P.D.; Monds, R.D.; O’Toole, G.A. LapD is a bis-(3′,5′)-cyclic dimeric GMP-binding protein that regulates surface attachment by Pseudomonas fluorescens Pf0-1. Proc. Natl. Acad. Sci. USA 2009, 106, 3461–3466. [Google Scholar] [CrossRef] [Green Version]
- Suhre, K.; Schmitt-Kopplin, P. MassTRIX: Mass translator into pathways. Nucl. Acids Res. 2008, 36, W481–W484. [Google Scholar] [CrossRef] [Green Version]
- Darty, K.; Denise, A.; Ponty, Y. VARNA, Interactive drawing and editing of the RNA secondary structure. Bioinformatics 2009, 25, 1974–1975. [Google Scholar] [CrossRef] [Green Version]
- Lapouge, K.; Schubert, M.; Allain, F.H.T.; Haas, D. Gac/Rsm signal transduction pathway of gamma-proteobacteria, from RNA recognition to regulation of social behaviour. Mol. Microbiol. 2008, 67, 241–253. [Google Scholar] [CrossRef]
- Edwards, A.N.; Patterson-Fortin, L.M.; Vakulskas, C.A.; Mercante, J.W.; Potrykus, K.; Vinella, D.; Camacho, M.I.; Fields, J.A.; Thopson, S.A.; Georgellis, D.; et al. Circuitry linking the Csr and stringent response global regulatory systems. Mol. Microbiol. 2011, 80, 1561–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, K.; Yamada, K.; Haas, D. ppGpp controlled by the Gac/Rsm regulatory pathway sustains biocontrol activity in Pseudomonas fluorescens CHA0. Mol. Plant Microbe Interact. 2012, 25, 1440–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moscoso, J.A.; Mikkelsen, H.; Heeb, S.; Williams, P.; Filloux, A. The Pseudomonas aeruginosa sensor RetS switches type III and type VI secretion via c-di-GMP signalling. Environ. Microbiol. 2011, 13, 3128–3138. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.; Zhang, B.; Yang, Q.; Zhang, Z.; Zheng, D.; Zhang, L.Q.; Yan, Q.; Wu, X. Cyclic-di-GMP regulates the quorum-sensing system and the biocontrol activity of Pseudomonas fluorescens 2P24 through RsmA and RsmE proteins. Appl. Environ. Microbiol. 2020. [Google Scholar] [CrossRef]
- Frangipani, E.; Visaggio, D.; Heeb, S.; Kaever, V.; Camara, M.; Visca, P.; Imperi, F. The Gac/-Rsm and cyclic-di-GMP signalling networks coordinately regulate iron uptake in Pseudomonas aeruginosa. Environ. Microbiol. 2014, 16, 676–688. [Google Scholar] [CrossRef]
- Hammer, B.K.; Bassler, B.L. Regulatory small RNAs circumvent the conventional quorum sensing pathway in pandemic Vibrio cholerae. Proc. Natl. Acad. Sci. USA 2007, 104, 11145–11149. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, S.T.; Valastyan, J.S.; Taillefumier, T.; Wingreen, N.S.; Bassler, B.L. Comprehensive analysis reveals how single nucleotides contribute to noncoding RNA function in bacterial quorum sensing. Proc. Natl. Acad. Sci. USA 2015, 112, E6038–E6047. [Google Scholar] [CrossRef] [Green Version]
- Wilderman, P.J.; Sowa, N.A.; FitzGerald, D.J.; FitzGerald, P.C.; Gottesman, S.; Ochsner, U.A.; Vasil, M.L. Identification of tandem duplicate regulatory small RNAs in Pseudomonas aeruginosa involved in iron homeostasis. Proc. Natl. Acad. Sci. USA 2004, 101, 9792–9797. [Google Scholar] [CrossRef] [Green Version]
- Padalon-Brauch, G.; Hershberg, R.; Elgrably-Weiss, M.; Baruch, K.; Rosenshine, I.; Mar-galit, H.; Altuvia, S. Small RNAs encoded within genetic islands of Salmonella typhimurium show host-induced expression and role in virulence. Nucleic Acids Res. 2008, 36, 1913–1927. [Google Scholar] [CrossRef]
- Macintyre, R.J. Evolution and ecological value of duplicate genes. Annu. Rev. Ecol. Syst. 1976, 7, 421–468. [Google Scholar] [CrossRef]
- Reams, A.B.; Neidle, E.L. Selection for gene clustering by tandem duplication. Annu. Rev. Microbiol. 2004, 58, 119–142. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.P.; Roth, J.R. Tandem genetic duplications in phage and bacteria. Annu. Rev. Microbiol. 1977, 31, 473–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edlund, T.; Grundstrom, T.; Normark, S. Isolation and characterization of DNA repetitions carrying the chromosomal beta-lactamase gene of Escherichia coli K-12. Mol. Gen. Genet. 1979, 173, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Lalaouna, D.; Fochesato, S.; Barakat, M.; Ortet, P.; Achouak, W. Multiple transcription-activating sequences regulate the RsmZ regulatory small RNA of Pseudomonas brassicacearum. J. Bacteriol. 2012, 194, 4888–4893. [Google Scholar] [CrossRef] [Green Version]
- Janssen, K.H.; Diaz, M.R.; Gode, C.J.; Wolfgang, M.C.; Yahr, T.L. RsmV, a small noncoding regulatory RNA in Pseudomonas aeruginosa that sequesters RsmA and RsmF from target mRNAs. J. Bacteriol. 2018, 200, e00277-18. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.L.; Romero, M.; Karna, S.L.; Chen, T.; Heeb, S.; Leung, K.P. RsmW, Pseudomonas aeruginosa small non-coding RsmA-binding RNA upregulated in biofilm versus planktonic growth conditions. BMC Microbiol. 2016, 16, 155. [Google Scholar] [CrossRef]
- Storz, G.; Opdyke, J.A.; Zhang, A. Controlling mRNA stability and translation with small, noncoding RNAs. Curr. Opin. Microbio. 2004, 7, 140–144. [Google Scholar] [CrossRef]
- Suzuki, K.; Babitzke, P.; Kushner, S.R.; Romeo, T. Identification of a novel regulatory protein (CsrD) that targets the global regulatory RNAs CsrB and CsrC for degradation by RNase E. Genes Dev. 2006, 20, 2605–2617. [Google Scholar] [CrossRef] [Green Version]
- Viegas, S.C.; Silva, I.J.; Saramago, M.; Domingues, S.; Arraiano, C.M. Regulation of the small regulatory RNA MicA by ribonuclease III, a target-dependent pathway. Nucleic Acids Res. 2011, 39, 2918–2930. [Google Scholar] [CrossRef]
- Reimmann, C.; Valverde, C.; Kay, E.; Haas, D. Posttranscriptional repression of GacS/GacA-controlled genes by the RNA-binding protein RsmE acting together with RsmA in Biocontrol strain Pseudomonas fluorescens CHA0. J. Bacteriol. 2005, 187, 276–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, A.; Cui, Y.; Chatterjee, A.K. RsmA and the quorum-sensing signal, N-[3-oxohexanoyl]-L-homoserine lactone, control the levels of rsmB RNA in Erwinia carotovora subsp. carotovora by affecting its stability. J. Bacteriol. 2002, 184, 4089–4095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masse, E.; Escorcia, F.E.; Gottesman, S. Coupled degradation of a small regulatory RNA and its mRNA targets in Escherichia coli. Genes Dev. 2003, 17, 2374–2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorger-Domenigg, T.; Sonnleitner, E.; Kaberdin, V.R.; Blasi, U. Distinct and overlapping binding sites of Pseudomonas aeruginosa Hfq and RsmA proteins on the non-coding RNA RsmY. Biochem. Biophys. Res. Commun. 2007, 352, 769–773. [Google Scholar] [CrossRef]
- Janssen, K.H.; Corley, J.M.; Djapgne, L.; Cribbs, J.T.; Voelker, D.; Slusher, Z.; Nordell, R.; Regulski, E.E.; Kazmierczak, B.I.; McMackin, E.W.; et al. Hfq and sRNA 179 inhibit expression of the Pseudomonas aeruginosa cAMP-Vfr and type III secretion regulons. mBio 2020, 11, e00363-20. [Google Scholar] [CrossRef] [PubMed]
- Haas, D.; Defago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [PubMed]
- Takeuchi, K.; Kiefer, P.; Reimmann, C.; Keel, C.; Dubuis, C.; Rolli, J.; Vorgolt, J.A.; Haas, D. Small RNA-dependent Expression of Secondary Metabolism Is Controlled by Krebs Cycle Function in Pseudomonas fluorescens. J. Biol. Chem. 2009, 284, 34976–34985. [Google Scholar] [CrossRef] [Green Version]
- Romling, U.; Galperin, M.Y.; Gomelsky, M. Cyclic di-GMP: The first 25 years of a universal bacterial second messenger. Microbiol. Mol. Biol. Rev. 2013, 77, 1–52. [Google Scholar] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Gene | Begin | End | Size (nt) | Sequence Identity (%) | Number of rsmX-Like Genes |
|---|---|---|---|---|---|---|
| Pseudomonas brassicacearum NFM421 | rsmX-1 | 4839892 | 4840003 | 111 | 75 | 2 |
| rsmX-2 | 4839681 | 4839791 | 110 | |||
| Pseudomonas fluorescens F113 | rsmX-1 * | 2113839 | 2113950 | 111 | 75 | 2 |
| rsmX-2 * | 2114052 | 2114160 | 108 | |||
| Pseudomonas stutzeri A150 | rsmX-1 * | 356904 | 357013 | 109 | 81.5 | 2 |
| rsmX-2 * | 357143 | 357250 | 107 | |||
| Pseudomonas syringae pv. phaseolicola 1448A | rsmX-4 ** | 160448 | 160336 | 112 | 85 | 5 |
| rsmX-3 ** | 160740 | 160627 | 113 | |||
| Pseudomonas syringae pv. syringae B728a | rsmX-3 ** | 5867169 | 5867282 | 113 | 84 | 5 |
| rsmX-4 ** | 5867461 | 5867572 | 111 | |||
| Pseudomonas syringae pv. tomato str. DC3000 | rsmX-3 | 6144830 | 6144943 | 113 | 87 | 5 |
| rsmX-4 | 6145122 | 6145235 | 113 |
| Medium | Phase | Half-Lives in WT (in Min) | ||||
|---|---|---|---|---|---|---|
| OD600nm | RsmX-1 | RsmX-2 | RsmY | RsmZ | ||
| TSB | Expo | 1.5 | 12 ± 1.1 | 5 ± 0.6 | 8 ± 0.2 | 37 ± 2.6 |
| Stat | 4.5 | 23 ± 5.4 | 16 ± 1.5 | 45 ± 5.2 | 53 ± 0.9 | |
| TSB/10 | Expo | 0.5 | 7 ± 1.5 | 5 ± 1.6 | 8 ± 1.7 | 26 ± 3.6 |
| Stat | 0.9 | 37 ± 4.4 | 40 ± 8.6 | ≥60 | ≥90 | |
| Half-Lives (in Min) | ||||
|---|---|---|---|---|
| Genes | GGA Motifs | GGA Motifs Exposed * | TSB/10 24 h | TSB 24 h |
| RsmX-1 | 6 | 2–4 | 37 ± 4.4 | 23 ± 5.4 |
| RsmX-2 | 6 | 3 | 40 ± 8.6 | 16 ± 1.5 |
| RsmY | 7 | 5–4 | ≥60 | 45 ± 5.2 |
| RsmZ | 10 | 8–7 | ≥90 | 53 ± 0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lalaouna, D.; Fochesato, S.; Harir, M.; Ortet, P.; Schmitt-Kopplin, P.; Heulin, T.; Achouak, W. Amplifying and Fine-Tuning Rsm sRNAs Expression and Stability to Optimize the Survival of Pseudomonas brassicacerum in Nutrient-Poor Environments. Microorganisms 2021, 9, 250. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020250
Lalaouna D, Fochesato S, Harir M, Ortet P, Schmitt-Kopplin P, Heulin T, Achouak W. Amplifying and Fine-Tuning Rsm sRNAs Expression and Stability to Optimize the Survival of Pseudomonas brassicacerum in Nutrient-Poor Environments. Microorganisms. 2021; 9(2):250. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020250
Chicago/Turabian StyleLalaouna, David, Sylvain Fochesato, Mourad Harir, Philippe Ortet, Philippe Schmitt-Kopplin, Thierry Heulin, and Wafa Achouak. 2021. "Amplifying and Fine-Tuning Rsm sRNAs Expression and Stability to Optimize the Survival of Pseudomonas brassicacerum in Nutrient-Poor Environments" Microorganisms 9, no. 2: 250. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020250