
Impacts of Low Temperature and Ensiling Period on the Bacterial Community of Oat Silage by SMRT

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Forage and Ensiling
2.2. Fermentation Quality and Chemical Composition Analysis
2.3. Microbial Population Analysis by Plate Culture
2.4. Microbial Community Analysis
2.4.1. DNA Extraction and Bacterial Composition Analysis by SMRT Method
2.4.2. SMRT Sequencing of Bacterial Diversity
2.5. Statistical Analyses
3. Results and Discussion
3.1. General Characteristics of Fresh Oat
3.2. Chemical Composition and Microbial Population by Plate Culture of Oat Silage
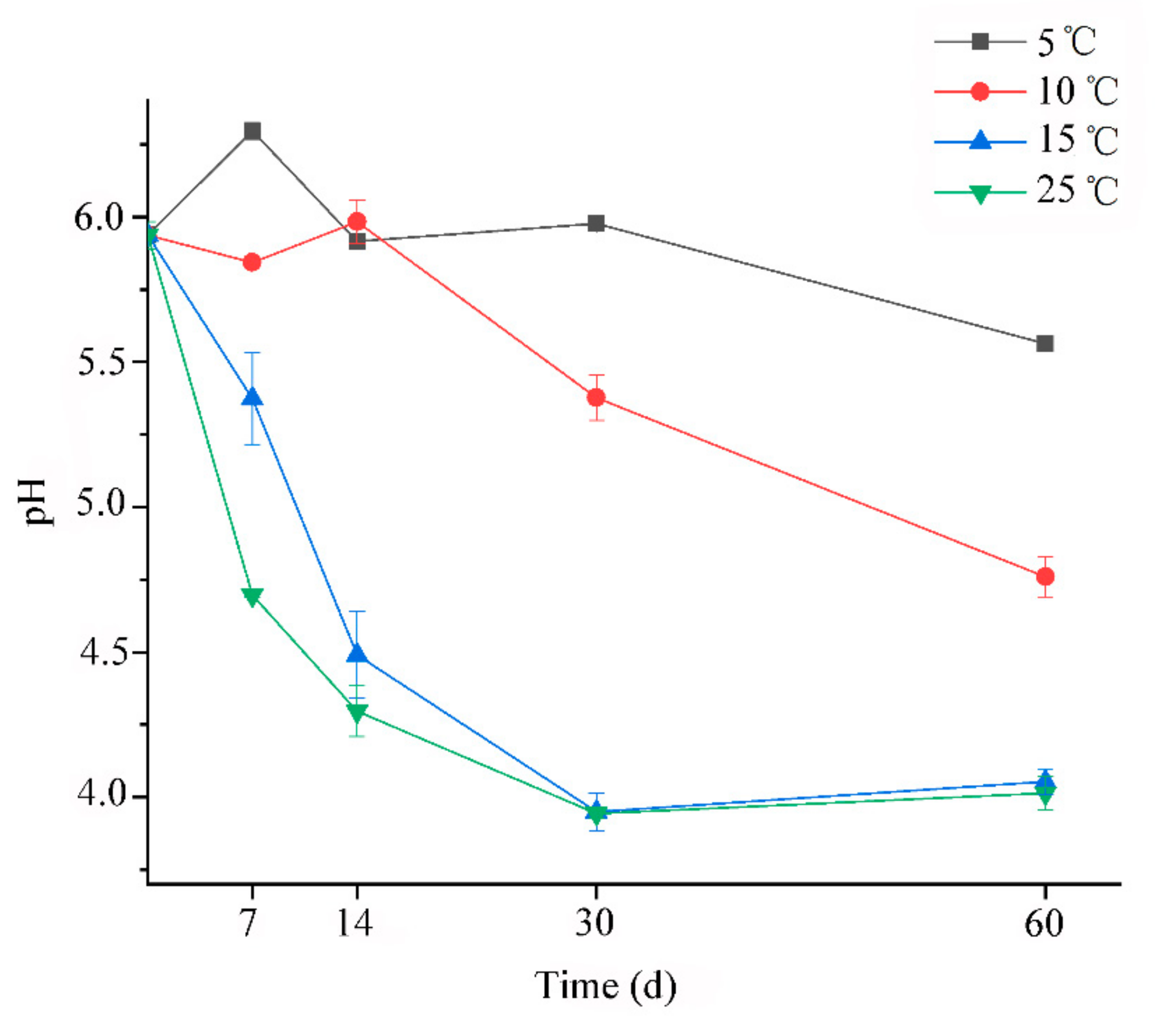
3.3. The Changes of pH and Organic Acid during Ensiling Process
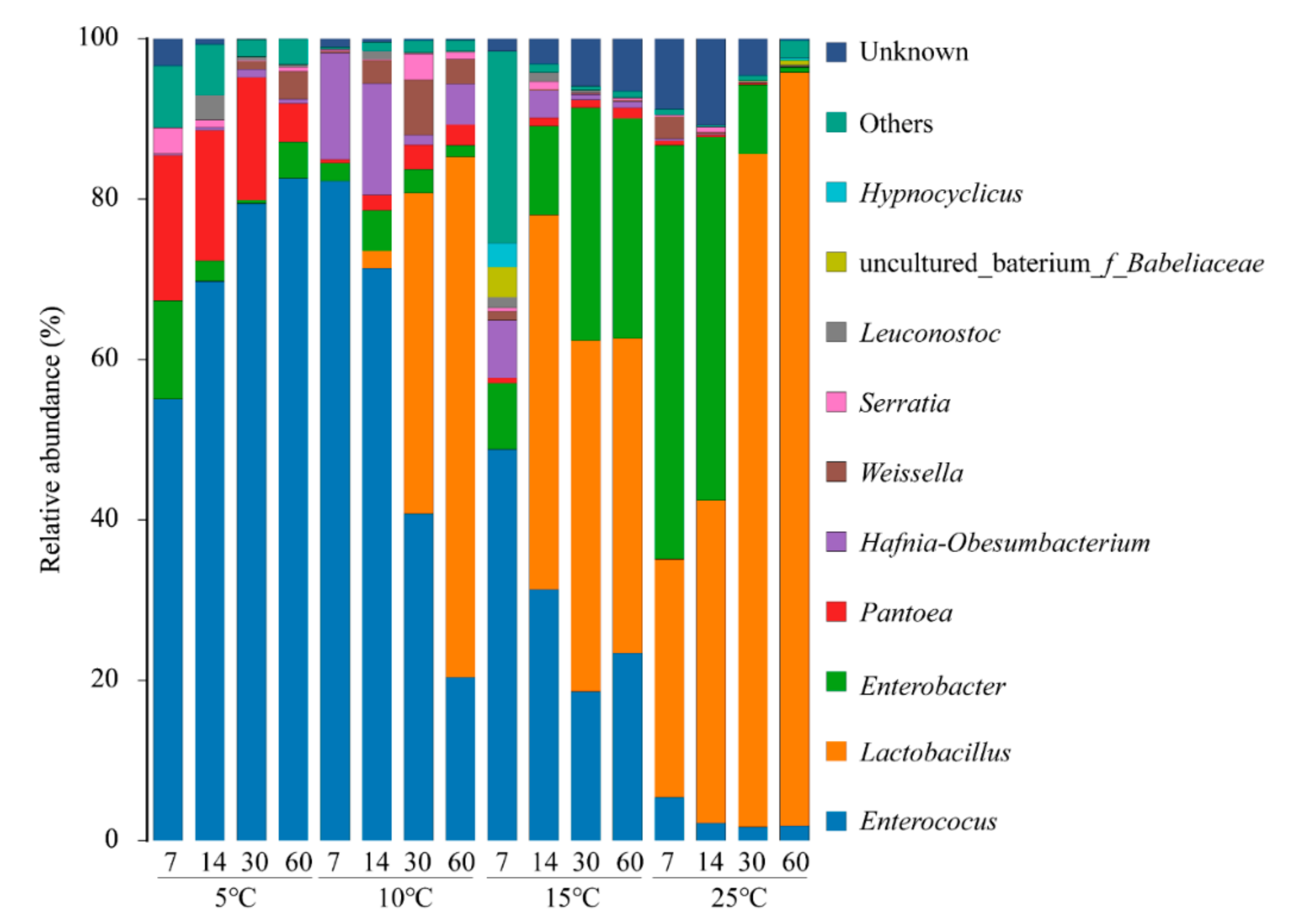
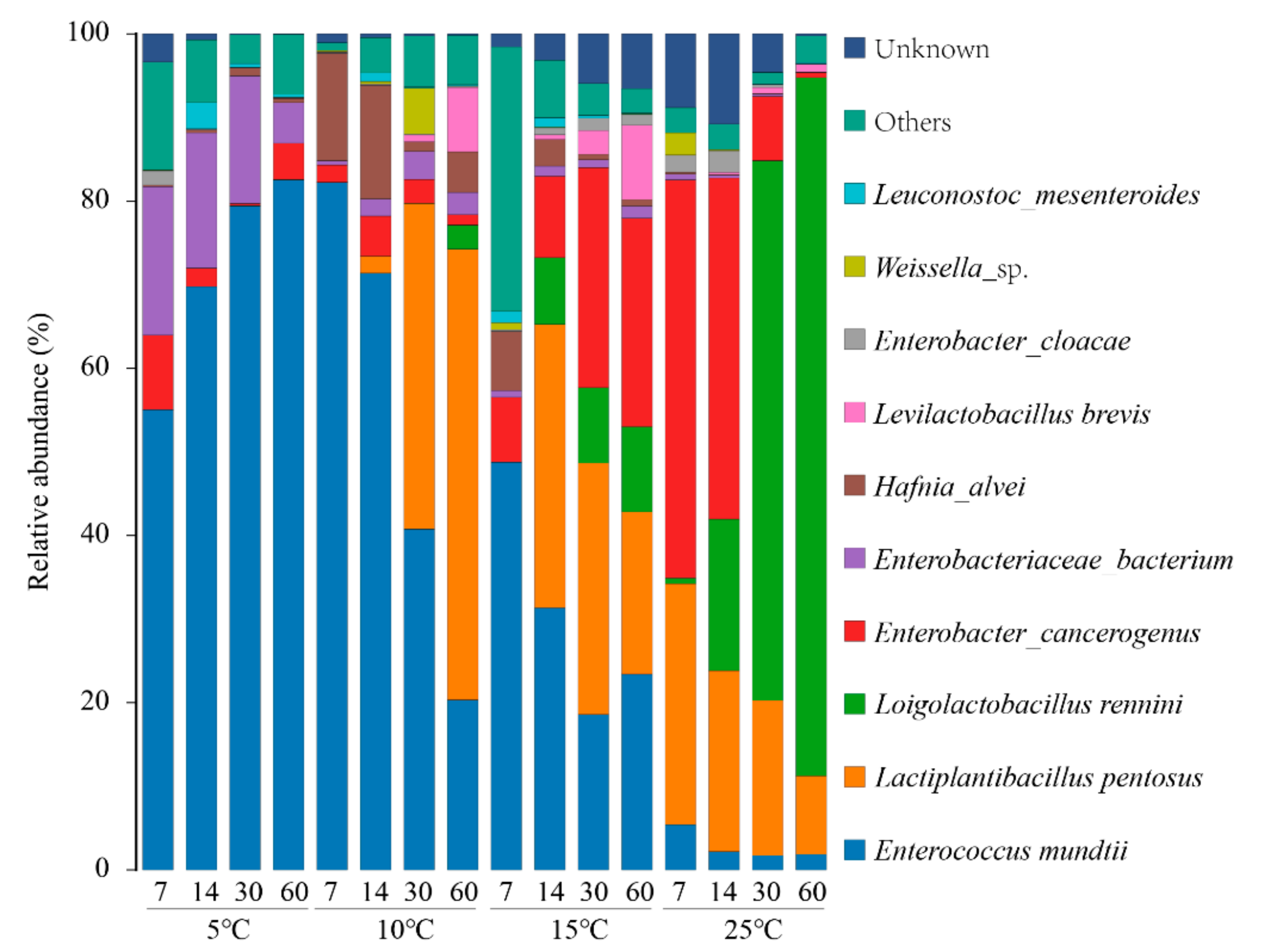
3.4. Bacterial Community during Ensiling Process
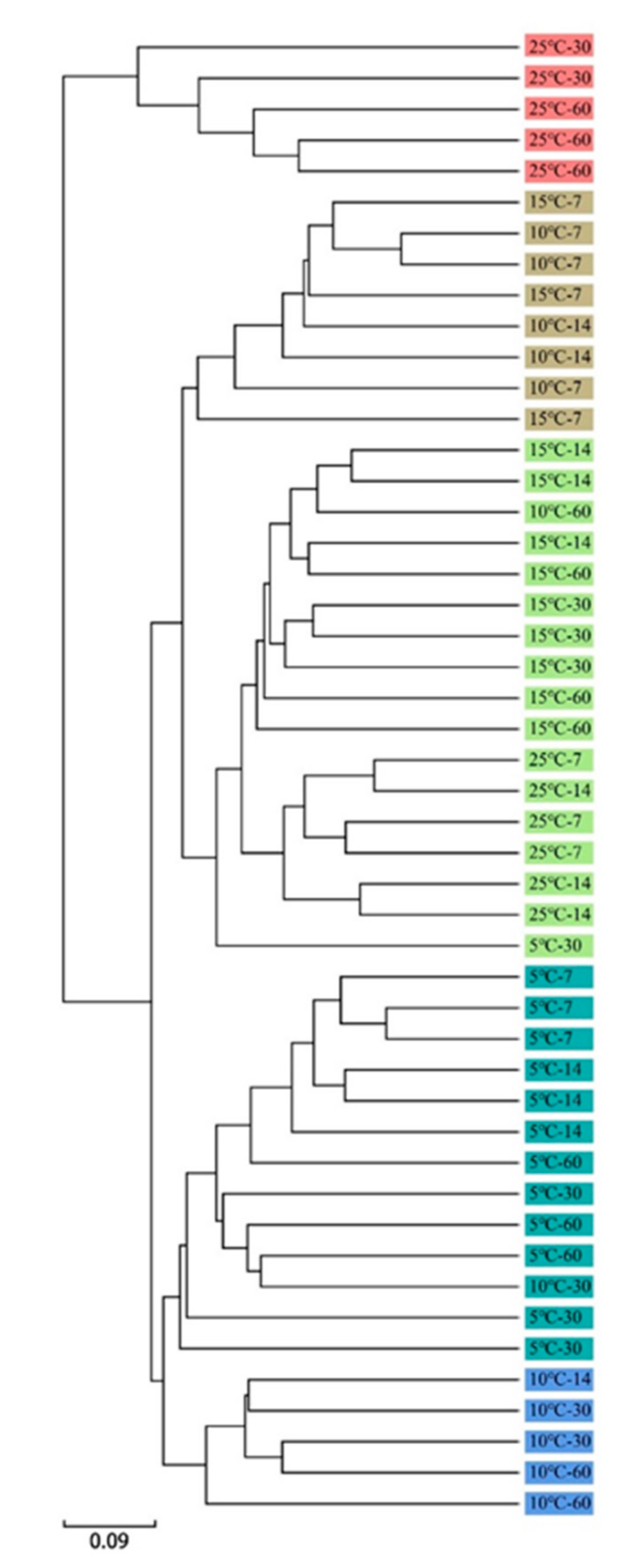
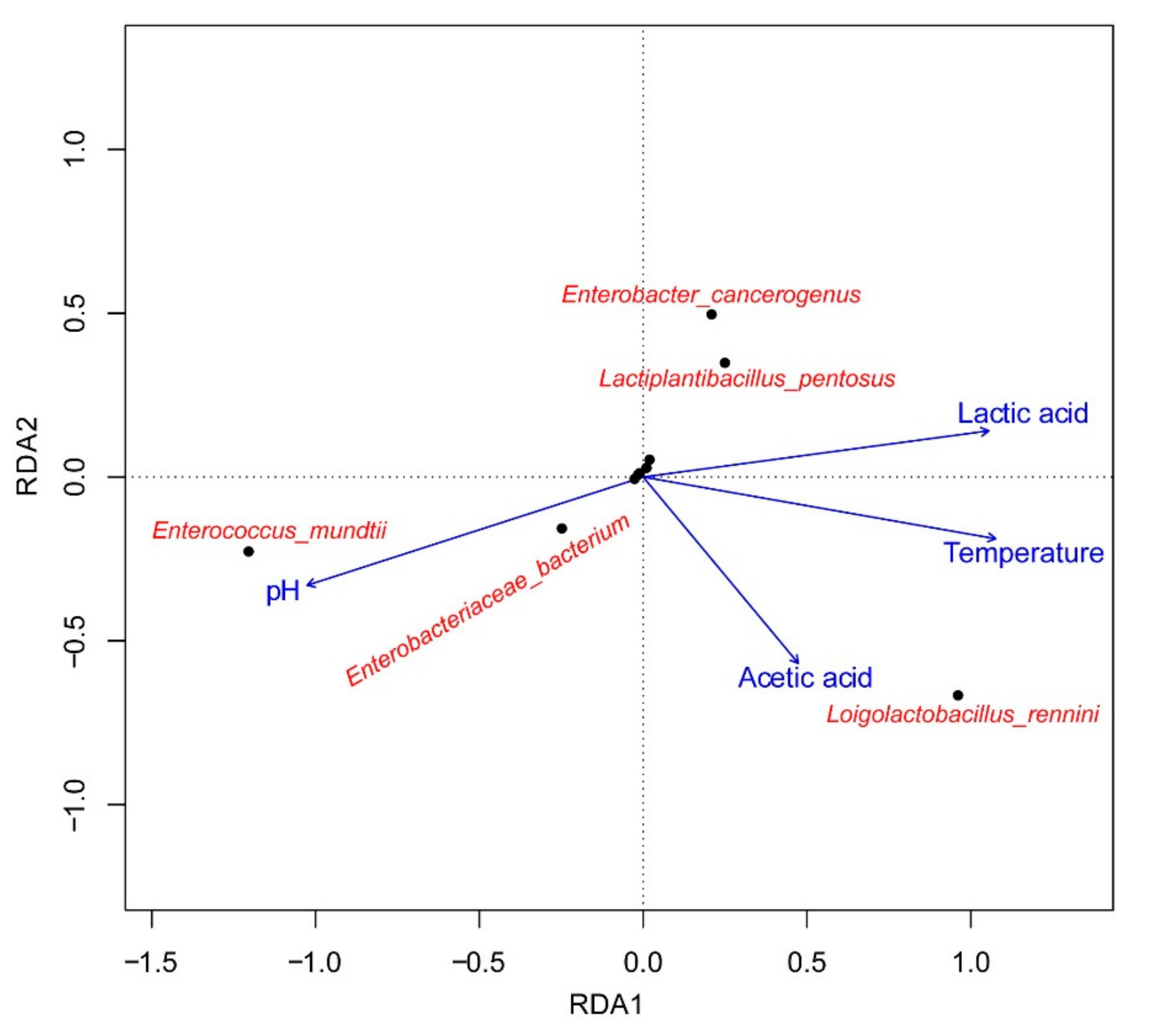
3.5. Cluster Analysis of the Bacterial Community and Its Correlation with Fermentation Products
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Storm, I.M.L.D.; Kristensen, N.B.; Raun, B.M.L.; Smedsgaard, J.; Thrane, U. Dynamics in the microbiologyof maize silage during whole-season storage. J. Appl. Microbiol. 2010, 109, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.M.; Benno, Y.; Ogawa, M.; Ohmomo, S.; Kumai, S.; Nakase, T. Influence of Lactobacillus spp. from an inoculant and of Weissella and Leuconostoc spp. from forage crops on silage fermentation. Appl. Environ. Microb. 1998, 64, 2982–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage; Chalcombe Publications: Marlow, UK, 1991. [Google Scholar]
- Vila, C.L.S.; Carvalho, B.F. Silage fermentation-updates focusing on the performance of micro-organisms. J. Appl. Microbiol. 2020, 128, 966–984. [Google Scholar] [CrossRef] [Green Version]
- Borreani, G.; Tabacco, E.; Schmidt, R.J.; Holmes, B.J.; Muck, R.E. Silage review: Factors affecting dry matter and quality losses in silages. J. Dairy Sci. 2018, 101, 3952–3979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Bai, C.; Liu, L.; Cao, B. Effects of lactic acid bacteria inoculant on the fermentation quality of reed grass (Phragmites australis Cav. Trin. ex Sterd.) at low temperature. Acta Agrestia Sinica 2011, 19, 127–131. [Google Scholar]
- Zhou, Y.; Drouin, P.; Lafreniere, C. Effect of temperature (5–25 °C) on epiphytic lactic acid bacteria populations and fermentation of whole-plant corn silage. J. Appl. Microbiol. 2016, 121, 657–671. [Google Scholar] [CrossRef]
- Liu, Q.H.; Shao, T.; Bai, Y.F. The effect of fibrolytic enzyme, Lactobacillus plantarum and two food antioxidants on the fermentation quality, alpha-tocopherol and beta-carotene of high moisture napier grass silage ensiled at different temperatures. Anim. Feed Sci. Tech. 2016, 221, 1–11. [Google Scholar] [CrossRef]
- Ren, H.; Feng, Y.; Pei, J.; Li, J.; Wang, Z.; Fu, S.; Zheng, Y.; Li, Z.; Peng, Z. Effects of Lactobacillus plantarum additive and temperature on the ensiling quality and microbial community dynamics of cauliflower leaf silages. Bioresour. Technol. 2020, 307, 123. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.; Wang, B.; Yu, Z.; Wu, Z. The effects of stage of maturity and lactic acid bacteria inoculants on the ensiling characteristics, aerobic stability and in vitro digestibility of whole-crop oat silages. Grassl. Sci. 2020, 67, 55–62. [Google Scholar] [CrossRef]
- Ni, K.K.; Wang, F.; Zhu, B.; Yang, J.; Zhou, G.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Yan, Y.H.; Li, X.L.; Li, X.M.; Shuai, Y.; Feng, G.Y.; Ran, Q.F.; Cai, Y.M.; Li, Y.; Zhang, X.Q. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Amnon, A.; Amit, Z.; Or, Z.; Michael, E.; Shay, S.; Ohad, S. High-resolution microbial community reconstruction by integrating short reads from multiple 16S rRNA regions. Nucleic Acids Res. 2013, 22, 205. [Google Scholar]
- Yan, Y.; Li, X.; Guan, H.; Huang, L.; Ma, X.; Peng, Y.; Li, Z.; Nie, G.; Zhou, J.; Yang, W.; et al. Microbial community and fermentation characteristic of Italian ryegrass silage prepared with corn stover and lactic acid bacteria. Bioresour. Technol. 2019, 279, 166–173. [Google Scholar] [CrossRef]
- Du, Z.M.; Sun, L.; Chen, C.; Lin, J.; Yang, F.Y.; Cai, Y. Exploring microbial community structure and metabolic gene clusters during silage fermentation of paper mulberry, a high-protein woody plant. Anim. Feed. Sci. Tech 2020. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Vansoest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Guo, L.; Yao, D.; Li, D.; Lin, Y.; Bureenok, S.; Ni, K.; Yang, F. Effects of Lactic Acid Bacteria Isolated from Rumen Fluid and Feces of Dairy Cows on Fermentation Quality, Microbial Community, and in vitro Digestibility of Alfalfa Silage. Front. Microbiol. 2020, 10, 2998. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Gloeckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Saldinger, S.S. Bacterial Dynamics of Wheat Silage. Front. Microbiol. 2019, 10, 1532. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.P.; Li, Z.Z.; Yu, Z.; Zhang, Q.; Li, X.J. Interactive effect of inoculant and dried jujube powder on the fermentation quality and nitrogen fraction of alfalfa silage. Anim. Sci. J. 2017, 88, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.L.M.; Jacovaci, F.A.; Bolson, D.C.; Nussio, L.G.; Jobim, C.C. Effects of light wilting and heterolactic inoculant on the formation of volatile organic compounds, fermentative losses and aerobic stability of oat silage. Anim. Feed. Sci. Tech. 2019, 247, 94–198. [Google Scholar] [CrossRef]
- Wang, T.; Teng, K.; Cao, Y.; Shi, W.; Xuan, Z.; Zhou, J.; Zhang, J.; Zhong, J. Effects of Lactobacillus hilgardii 60TS-2, with or without homofermentative Lactobacillus plantarum B90, on the aerobic stability, fermentation quality and microbial community dynamics in sugarcane top silage. Bioresour. Technol. 2020, 312, 123. [Google Scholar] [CrossRef] [PubMed]
- Vasco-Correa, J.; Li, Y. Solid-state anaerobic digestion of fungal pretreated Miscanthus sinensis harvested in two different seasons. Bioresour. Technol. 2015, 185, 211–217. [Google Scholar] [CrossRef]
- Ni, K.; Zhao, J.; Zhu, B.; Su, R.; Pan, Y.; Ma, J.; Zhou, G.; Tao, Y.; Liu, X.; Zhong, J. Assessing the fermentation quality and microbial community of the mixed silage of forage soybean with crop corn or sorghum. Bioresour. Technol. 2018, 265, 563–567. [Google Scholar] [CrossRef]
- Rooke, J.A.; Hatfield, R.D. Biochemistry of ensiling. Silage Sci. Technol. 2003, 42, 95–139. [Google Scholar]
- Bigwood, T.; Hudson, J.A.; Cooney, J.; McIntyre, L.; Billington, C.; Heinemann, J.A.; Wall, F. Inhibition of Listeria monocytogenes by Enterococcus mundtii isolated from soil. Food Microbiol. 2012, 32, 354–360. [Google Scholar] [CrossRef]
- Klein, G. Taxonomy, ecology and antibiotic resistance of enterococci from food and the gastro-intestinal tract. Int. J. Food Microbiol. 2003, 88, 123–131. [Google Scholar] [CrossRef]
- Agarussi, M.C.N.; Pereira, O.G.; de Paula, R.A.; da Silva, V.P.; Roseira, J.P.S.; Silva, F.F.E. Novel lactic acid bacteria strains as inoculants on alfalfa silage fermentation. Sci. Rep. 2019, 9, 8007. [Google Scholar] [CrossRef] [Green Version]
- Chenoll, E.; Carmen, M.M.; Aznar, R. Lactobacillus rennini sp. nov., isolated from rennin and associated with cheese spoilage. Int. J. Syst. Evol. Micr 2006, 56, 449–452. [Google Scholar] [CrossRef]
- Pérez-Díaz, I.M.; Hayes, J.; Medina, E.; Webber, A.M.; Butz, N.; Dickey, A.N.; Lu, Z.J.; Azcarate-Peril, M.A. Assessment of the Non-lactic acid bacteria microbiota in fresh cucumbers and commercially fermented cucumber pickles brined with 6% NaCl. Food Microbiol. 2019, 77, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Nishino, N. Effects of ensiling fermentation and aerobic deterioration on the bacterial community in Italian ryegrass, Guinea grass, and whole-crop maize silages stored at high moisture content. Asian Austral. J. Anim. 2013, 26, 1304–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Fresh Oat |
|---|---|
| Dry matter (%) | 23.70 ± 1.31 |
| Crude protein (%DM) | 13.25 ± 1.19 |
| Acid detergent fiber (%DM) | 32.80 ± 1.07 |
| Neutral detergent fiber (%DM) | 49.64 ± 2.04 |
| Organic matter (%DM) | 95.23 ± 3.20 |
| Ether extract (%DM) | 5.12 ± 0.13 |
| Lactic acid bacteria (Log10 cfu/g FM−1) | 3.61 ± 0.12 |
| Yeasts (Log10 cfu/g FM−1) | 4.37 ± 0.23 |
| Aerobic bacteria (Log10 cfu/g FM−1) | 5.12 ± 0.19 |
| Mold (Log10 cfu/g FM−1) | ND |
| Coliform | 4.1 ± 0.03 |
| Temperature | DM(%) | Chemical Composition (%DM) | Microbial Population (Log10 cfu/g FM−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CP | ADF | NDF | OM | EE | LAB | Yeast | Aerobic Bacteria | Mold | Coliform | ||
| 5 °C | 27.87 | 11.82 b | 29.29 b | 45.40 b | 94.34 | 5.34 | 5.51 a | ND | 6.02 | ND | 3.16 b |
| 10 °C | 26.96 | 11.70 b | 31.73 a | 47.05 a | 94.22 | 5.33 | 5.62 a | ND | 7.21 | ND | 3.21 b |
| 15 °C | 25.85 | 12.68 a | 32.00 a | 47.49 a | 93.56 | 5.04 | 5.84 a | ND | 6.55 | ND | 4.45 a |
| 25 °C | 25.71 | 12.41 a | 31.27 a | 47.19 a | 94.45 | 5.23 | 4.25 b | ND | 6.43 | ND | 3.08 b |
| SEM | 1.58 | 0.26 | 0.86 | 1.56 | 0.88 | 0.23 | 0.53 | − | 0.56 | − | 0.88 |
| Significant analysis | NS | * | * | NS | NS | NS | * | − | NS | − | * |
| Temperature | Day | OTU | Shannon | Chao | Coverage |
|---|---|---|---|---|---|
| 5 °C | |||||
| 7 | 51a | 1.51 b | 61 c | 0.99 | |
| 14 | 48 a | 1.08 bc | 67 bc | 0.99 | |
| 30 | 31c | 0.55 c | 57 c | 0.99 | |
| 60 | 32 c | 0.82 bc | 78 b | 0.99 | |
| 10 °C | |||||
| 7 | 40 b | 0.63 c | 57 c | 0.99 | |
| 14 | 41 b | 1.04 bc | 65 c | 0.99 | |
| 30 | 49 a | 1.34 b | 75 a | 0.99 | |
| 60 | 51 a | 1.45 b | 88 a | 0.99 | |
| 15 °C | |||||
| 7 | 60 a | 2.10 a | 71 b | 0.99 | |
| 14 | 45 b | 1.79 b | 55 c | 0.99 | |
| 30 | 51 a | 1.81 b | 70 b | 0.99 | |
| 60 | 50 a | 1.89 b | 87 a | 0.99 | |
| 25 °C | |||||
| 7 | 40 b | 1.43 b | 58 c | 0.99 | |
| 14 | 36 bc | 1.43 b | 44 d | 0.99 | |
| 30 | 30 c | 0.94 c | 61 c | 0.99 | |
| 60 | 32 c | 0.71 c | 77 b | 0.99 | |
| SEM | 5.67 | 0.09 | 3.17 | 0.00 | |
| Significant analysis: | |||||
| Temperature (T) | NS | p < 0.05 | NS | NS | |
| Day (D) | p < 0.05 | p < 0.05 | p < 0.05 | NS | |
| T×D | p < 0.05 | p < 0.05 | NS | NS | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Chen, F.; Wang, X.; Sun, L.; Guo, L.; Xiong, Y.; Wang, Y.; Zhou, H.; Jia, S.; Yang, F.; et al. Impacts of Low Temperature and Ensiling Period on the Bacterial Community of Oat Silage by SMRT. Microorganisms 2021, 9, 274. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020274
Li X, Chen F, Wang X, Sun L, Guo L, Xiong Y, Wang Y, Zhou H, Jia S, Yang F, et al. Impacts of Low Temperature and Ensiling Period on the Bacterial Community of Oat Silage by SMRT. Microorganisms. 2021; 9(2):274. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020274
Chicago/Turabian StyleLi, Xiaomei, Fei Chen, Xuekai Wang, Lin Sun, Linna Guo, Yi Xiong, Yuan Wang, Hongzhang Zhou, Shangang Jia, Fuyu Yang, and et al. 2021. "Impacts of Low Temperature and Ensiling Period on the Bacterial Community of Oat Silage by SMRT" Microorganisms 9, no. 2: 274. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020274