
Streptococcal Infections in Marine Mammals


1
Institute for Microbiology, University of Veterinary Medicine Hannover, Foundation, Bischofsholer Damm 15, 30173 Hannover, Germany
2
Institute for Terrestrial and Aquatic Wildlife Research, University of Veterinary Medicine Hannover, Foundation, Werftstraße 6, 25761 Buesum, Germany
3
Institute of Microbiology and Epizootics, Department of Veterinary Medicine, Freie Universität Berlin, Robert-von-Ostertag-Strasse 7-13, 14163 Berlin, Germany
*
Author to whom correspondence should be addressed.
Microorganisms 2021, 9(2), 350; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020350
Submission received: 2 December 2020
/
Revised: 29 January 2021
/
Accepted: 7 February 2021
/
Published: 10 February 2021
(This article belongs to the Special Issue Epidemiology and Pathogenicity of Animal-Adapted Streptococci)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Marine mammals are sentinels for the marine ecosystem and threatened by numerous factors including infectious diseases. One of the most frequently isolated bacteria are beta-hemolytic streptococci. However, knowledge on ecology and epidemiology of streptococcal species in marine mammals is very limited. This review summarizes published reports on streptococcal species, which have been detected in marine mammals. Furthermore, we discuss streptococcal transmission between and adaptation to their marine mammalian hosts. We conclude that streptococci colonize and/or infect marine mammals very frequently, but in many cases, streptococci isolated from marine mammals have not been further identified. How these bacteria disseminate and adapt to their specific niches can only be speculated due to the lack of respective research. Considering the relevance of pathogenic streptococci for marine mammals as part of the marine ecosystem, it seems that they have been neglected and should receive scientific interest in the future.
1. Introduction
The marine ecosystem is challenged by numerous factors such as anthropogenic pollution including wastewater [1,2], plastic [3], chemical [4,5], and noise pollution [6], fisheries and prey depletion [7,8], offshore-construction [4], shipping [9,10], harmful algal blooms [11,12], climate change, and acidification [13,14,15,16,17]. Marine mammals are considered sentinels for the marine ecosystem [18,19,20] and, thus, the assessment of their health status should be of global interest and importance. Cumulative effects caused by different anthropogenic activities (e.g., pollution of the environment with PCBs, PBDEs, microplastic) can have suppressive effects on the immune system of marine mammals and this might result in a higher susceptibility for infectious diseases [21,22,23,24]. There are a number of studies indicating a higher prevalence of or risk for infectious disease in correlation with those (anthropogenic induced) threatens. For instance, in a recent study, microplastic was found in each of 50 tested animals belonging to 10 different marine mammal species. Furthermore, animals that died due to infectious diseases showed a slightly higher amount of microplastic in the intestines [3]. Sanderson and Alexander [17] found that climate related factors such as sea surface temperature have a significant effect on the occurrence of infectious disease-induced mass mortality events. In another study, the risk for infectious disease mortality increased by 2% for each 1 mg/kg increase of polychlorinated biphenyls (PCBs) in the blubber of harbor porpoises [25].
Infectious diseases are one of the most frequent causes of death in marine mammals [26,27,28,29,30,31,32,33] and beta-hemolytic streptococci are frequently isolated and associated with disease [26,30,31,34,35,36,37]. The genus Streptococcus belongs to the family Streptococcaceae, the order Lactobacillales, the class Bacilli, and the phylum Firmicutes. Streptococci are non-motile, Gram-positive, catalase-negative, non-spore forming, and chemo-organotrophic with a fermentative metabolism [38,39]. They appear as ovoid cocci in pairs or chains and can be classified by their ability to cause different forms of hemolysis and their serologically active antigens according to the Lancefield classification scheme. Most of the currently described 79 streptococcal species are summarized in groups based on their 16S rRNA sequence, their pathogenic potential and specific characteristics. The pyogenic group includes human and animal pathogens, while the anginosus, mitis and salivarius groups also include commensals of the oral cavity and pharynx of humans, which, however, can also cause disease, such as endocarditis. Members of the mutans group colonize tooth surfaces and the bovis group contains members of the colon microflora [38,39,40,41].
Pathogenic streptococci can express a large arsenal of virulence(-associated) factors, such as exotoxins and enzymes, antiphagocytic hyaluronic acid containing capsule, host matrix protein binding surface proteins complement evasion factors, and, last but not least, antiphagocytic M- and M-like proteins [42,43,44].
However, the etiology and pathogenicity of streptococci to marine mammals are largely unknown. There is some debate as to whether streptococci are primary or opportunistic pathogens [34,45,46,47,48,49]. This discussion is further hampered by the lack of species identification in many reports. On the other hand, new species are regularly found in marine mammals such as S. halichoeri [50], S. marimammalium [51], and S. phocae [34].
Besides, some streptococcal species are known as zoonotic agents [52]. For instance, human meningitis was acquired as zoonoses from S. equi subsp. zooepidemicus after contact with domestic animals [53]. S. suis, a commensal and opportunistic pathogen of pigs, is also known to cause infections in humans [54,55]. S. canis and S. iniae are also zoonotic pathogens [56,57,58]. Thus, the ubiquitous distribution and zoonotic potential of streptococcal species represents a global health risk for animals and humans.
This review attempts to summarize what is currently known about streptococcal species detected in marine mammals and discusses important issues that deserve more attention in future research.
2. Streptococcal Findings in Marine Mammals
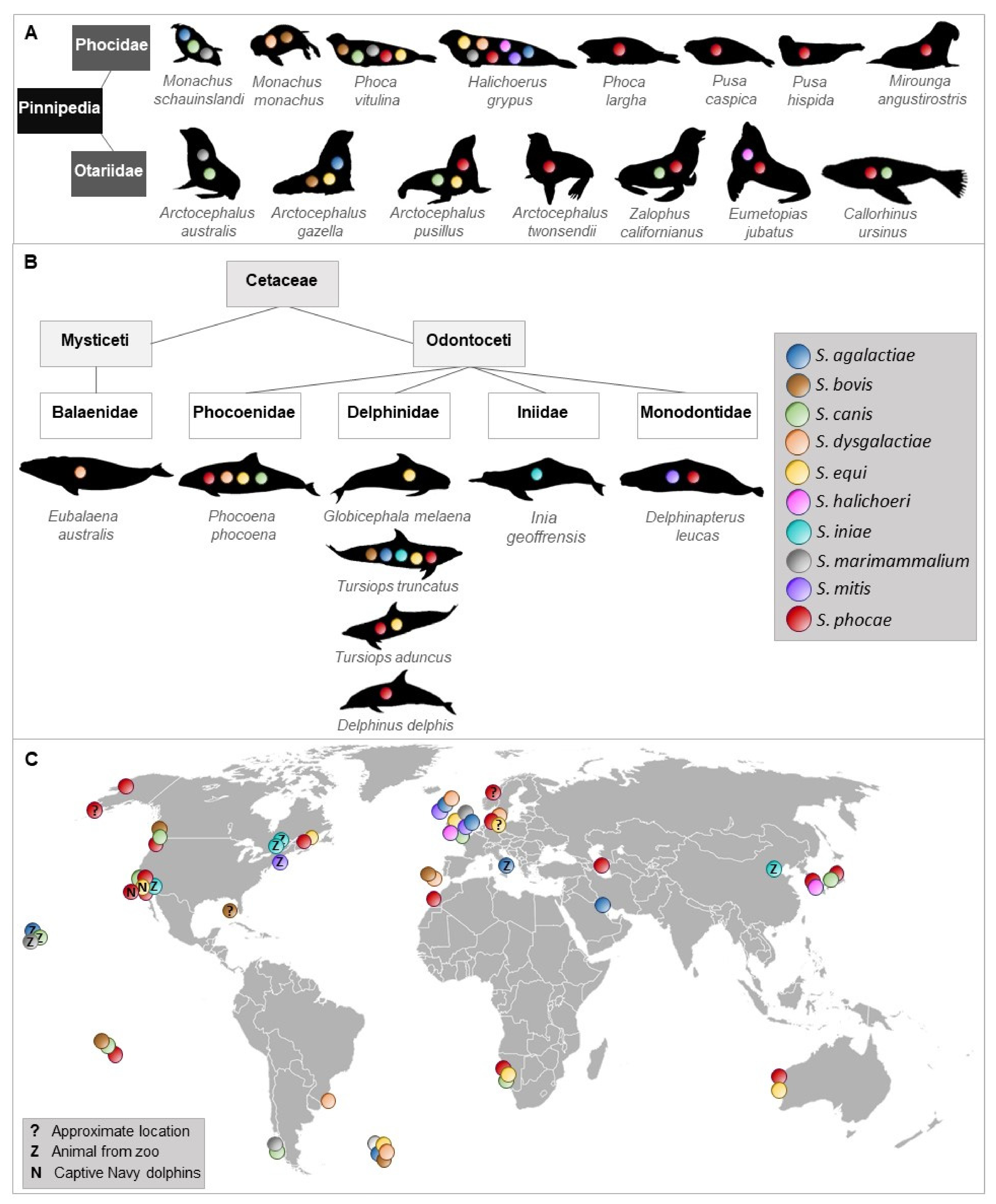
To the best of our knowledge, 10 streptococcal species were isolated and identified more than once from 23 species of Pinnipedia and Cetaceae worldwide (Figure 1, Supplementary Table S1).
2.1. Streptococcus agalactiae
S. agalactiae, also known as a bovine and human pathogen [59,60,61], was isolated from a wound and navel infection of grey seals (Halichoerus grypus) in North Rona in 1985 and 1986 [62] and from lesions of fight wounds, pneumonia, lymphadenitis as well as from lung and spleen samples of Antarctic fur seals (Arctocephalus gazella) from 1984–1987 on Bird Island, South Georgia [63]. Later, it was isolated from lesions and visceral organs (liver and lung) of a captive common bottlenose dolphin (Tursiops truncatus) that suffered from fatal necrotizing fasciitis and myositis [64]. One year later, the isolation of S. agalactiae from epaxial muscles of a wild stranded bottlenose dolphin was reported [65]. This strain caused 90% mortalities in tilapia in experimental infections and showed high similarity with strains associated with mullet kill in the concurrent Kuwait Bay. A mullet was found in the stomach of the dead dolphin, which might have served as a possible way of transmission. A study of human S. agalactiae strains from fish, seals, a dolphin, and a frog indicated zoonotic and anthroponotic hazard by causing severe disease in fish and compromising food security [66]. Between 2012 and 2014, S. agalactiae was isolated from a stranded grey seal on the British coast with ocular pathology [67]. In the Waikiki Aquarium, Honolulu, Haiwaii, S. agalactiae was isolated from two male healthy Hawaiian monk seals (M. schauinslandi) as part of their aerobic bacterial flora in the nasal cavity [68].
S. agalactiae is also known as serious fish pathogen [69,70,71]. In Brazil, high virulent strains were isolated from diseased Nile tilapia and transmission occurred by direct contact or through water [70]. Infection experiments confirmed the disease and revealed low LD50 for Nile tilapia. However, isolates from cattle did not cause any clinical signs in Nile tilapia and channel catfish indicating host specification and adaptation [72]. Human and bovine strains of S. agalactiae were able to cause disease in Nile tilapia, although there was no genetic relatedness of strains from fish, bovine, and human origin [73]. This suggests that the ability to cross host-specific barrier is not necessarily reflected by genetic linkage. Virulence gene profiling of S. agalctiae isolated from diseased tilapia in Thailand revealed a positive correlation of virulence genes content and pathogenicity [74]. Virulence genes for adhesion, invasion, and immune evasion were identified. Another study demonstrated that there were fish-specific genes or loci that were associated with disease in fish, while strains missing these regions were not able to cause morbidity in tilapia [75]. In addition, these fish-specific genes were mainly clustered in regions with signatures of mobile genetic elements and one fish-specific gene was found in the region, where the virulence genes rib or bca are in the human strain indicating genetic adaptation to the fish host.
2.2. Streptococcus bovis
S. bovis has been isolated from the gastrointestinal tract and feces of cattle, sheep, goats [76], and dogs [77]. It has also been identified as human pathogen associated with endocarditis [78], meningitis [79], septic arthritis [80], bacteremia, and gastrointestinal disease [81]. Virulence factors associated with S. bovis infection were, for instance, extracellular proteins [82] and antigens [83]. S. bovis was detected in fur seals with pneumonia that was characterized by extensive polymorphonuclear infiltrations and necrosis or very widespread abscess formation and, frequently, by additionally fibrinous exudative pleurisy [63]. Together with S. phocae and S. canis it was also isolated from dead herpesvirus-positive harbor seal pups at the Smith Island, Washington [32]. A monk seal pup (Monachus monachus) found on the island Deserta Grande, Portugal died due to septicemia and S. bovis was isolated and considered as a potential causative agent [84]. In 2006, S. bovis isolation (together with S. equisimilis/mitis) from free-ranging bottlenose dolphins that were captured, sampled, and released in coastal Gulf of Mexico and Atlantic ocean waters was reported [85].
2.3. Streptococcus canis
S. canis was first isolated from cows with mastitis and from dogs with different pathological findings [86], but can also cause infections in humans [56,57,87]. It has also been isolated from minks [88], feral cats [89] and cats [90]. Its virulence and pathogenicity were confirmed by the presence of virulence genes such as for fibronectin-binding protein, M proteins, protective antigen, and streptolysin [91,92,93,94]. The M protein of S. canis has also a high-affinity immunoglobulin G binding activity, which is not species-specific and facilitate S. canis to interact with different hosts [95].
A beta-hemolytic Streptococcus sp., biochemically similar to S. canis, was cultured from pyogranulomatus lesion of the laryngeal cartilages and epiglottis of an adult harbor seal (Phoca vitulina) [27]. S. canis was also isolated from peritoneal effusion of a captive California sea lion (Zalophus californianus) of the US Navy’s marine mammal program [96] and from a California sea lion with bilateral corneal ulceration of the London Zoo, UK [97]. During an increased mortality among South American fur seal (Arctocephalus australis) pups on Guafo Island, Chile, South America, S. canis together with S. marimammalium were isolated and associated with moderate to marked, multifocal, mucopurulent bronchopneumonia [98]. In August 1994, S. canis (and also S. phocae, S. equi subsp. zooepidemicus) was isolated from spleen, liver, and kidney of Cape fur seals (Arctocephalus pusillus pusillus) at Cape Cross, Namibia that suffered from respiratory infections and abortions associated with starvation [99]. Seven cases of stranded harbor porpoises (Phocoena phocoena) in England and Wales between 1990 and 1996 had a S. canis septicemia, which was isolated from lungs with pulmonary abscesses and enlarged pulmonary associated lymph nodes [100]. S. canis (together with S. phocae) was cultured from blood and lung samples of a dead, stranded Northern fur seal (Callorhinus ursinus) with necrotizing and fibrinous pneumonia infiltrated by band neutrophils with intraluminal abscess of bronchi at the coast of Niigata, Japan [46]. In 2005, the isolation of S. canis from two dead harbor seal pups on Smith Island, Washington was reported [32,101]. One died from omphalophlebitis and the other from omphalitis with subsequent peritonitis. S. canis was also isolated from the oral cavity of a male, healthy Hawaiian monk seal of the Waikiki Aquarium, Honolulu, Hawaii [68].
2.4. Streptococcus dysgalactiae
S. dysgalactiae subsp. equisimilis, previously known as S. equisimilis [102] and found in humans [103] and different animals such as dogs, cows, pigs, and horses [104], was isolated from Antarctic fur seal pups with septicemia and rhinitis in South Georgia, UK between 1979–1982 [105] and 1986 from a grey seal cow on North Rona, Scotland [62]. In the years 1988 and 1989, an increased number of harbor porpoise carcasses was observed in North and Baltic Seas [36]. Thirty-five isolates of beta-hemolytic streptococci were classified in Lancefield group L and identified as S. dysgalactiae subsp. dysgalactiae. In 1997, S. dysgalactiae and S. dysgalactiae subsp. equisimilis isolates were found in a dead, wild monk seal pup in association with a septicemia on the island Deserta Grande, Portugal [84]. Three isolates identified as S. dysgalactiae subsp. dysgalactiae were obtained from phocid seals (harbor and grey seals) stranded in the North and Baltic Seas between 1995 and 2000 [106]. Between 2005–2011, pathologic and microbiological findings of a southern right whale (Eubalaena australis) calf from Brazil indicated that beta-hemolytic S. dysgalactiae septicemia was responsible for the death of the animal [107].
2.5. Streptococcus equi
S. equi causes infections in horses [108] and was associated with canine infectious respiratory disease [109]. A systemic infection with S. equi in a horse handler has also been reported [110]. Further studies confirmed the zoonotic potential of S. equi [53,111]. In November 1978, a female North Atlantic pilot whale (Globicephala melaena) suffering from bronchopneumonia with lesions stranded on Metis Beach, Canada and S. equi (no further identification) was isolated from lung parenchyma, pharynx, and pericardial fluid [35]. A study from 1980 reported the isolation of S. equi subsp. zooepidemicus (previously S. zooepidemicus) from grey seals associated with mild, purulent pneumonia [49]. In 1994, it was isolated from the conjunctiva and trachea of two adult female Cape fur seals that had septicemic S. phocae in Namibia [99]. A total of 32 beta-hemolytic streptococcal isolates, collected during the phocine distemper outbreak in 2002 from 28 different harbor seals of the German North Sea, were identified as S. equi subsp. zooepidemicus [112]. Later, the same scientists isolated the same or at least very close related strains of S. equi subsp. zooepidemicus from grey seals and other harbor seals [113]. A retrospective study on 42 dead bottlenose dolphins from the US Navy Marine Mammal Program during 1980-2010 demonstrated an association of the isolation of S. equi subsp. zooepidemicus with pneumonia [114]. 16S rRNA sequences for S. equi (and S. phocae) were found in blow samples collected from four wild healthy Indo-Pacific bottlenose dolphins (T. aduncus) in Shark Bay (SB), Western Australia, in 2012 [115].
2.6. Streptococcus halichoeri
S. halichoeri, characterized as non-hemolytic and classified in Lancefield group B, was first isolated from dead grey seals in Iverness and Cornwall, UK [50] and few years later, in 2012, also from the kidney of a stranded, female Stellar sea lion (Eumetopias jubatus) in South Korea [116]. Also, in 2012, a severe case of a human infection with S. halichoeri was reported [117]. The patient had no contact to seals, but to fish, which could have been a possible transmission route. However, this was not tested. Another human infection was reported in 2018, where a man suffered from skin cellulitis due to S. halichoeri [118]. Shewmaker et al. [119] compared human and seal strains and concluded two subspecies S. halichoeri subsp. halichoeri for the seal isolates and S. halichoeri subsp. hominis for strains associated with human infections. The core genome of 20 S. halichoeri isolates from different hosts including dogs and minks contained 19 different streptococcal virulence factors, of which most were associated with adherence followed by proteases and toxins emphasizing its pathogenic potential [120,121].
2.7. Streptococcus iniae
S. iniae was described as new species in 1976, when it was first isolated from a captive Amazon freshwater dolphin (Inia geoffrensis) suffering from a dermatologic syndrome called “golf ball disease” in the Steinhart Aquarium, San Francisco, USA [122]. Further isolates were obtained from captive freshwater dolphins housed at the Niagara Falls Aquarium in New York, USA two years later [123], and in 1983 from a captive Amazon River dolphin at the Pittsburgh Zoo, USA that also developed the “golf ball disease” [124]. In 2015, a common dolphin died due to bacterial septicemia at Beijing aquarium, China, where S. iniae was isolated from hilar lymph nodes and pancreas of the dolphin [125].
S. iniae is also a serious fish pathogen [58,126]. Virulence mechanisms include a capsule with antiphagocytic function [127], the cytotoxin ß-hemolysin streptolysin S [128], an extracellular nuclease and s secreted nucleotidase that play an important role in immune evasion [129], a polysaccharide deacetylase involved in adherence, invasion, lysozyme resistance and survival in fish blood [130], and M-like protein [131]. Comparative genomics revealed genetic differences between strains from different hosts including I. geoffrensis and identified two plasticity zones that reflect adaptation to specific host environments [132]. Furthermore, the dolphin isolates differed from the fish and human isolates in lacking a capsule, forming denser and thicker biofilms, increased ability to withstand oxidative stress and were genetically highly divergent to the other isolates [133]. In addition, there were conserved mutation rates and mismatch/oxidized-guanine repair systems within phylogenetic clades, but significant differences between major phylogenetic lineages. Mutators might facilitate adaptation to novel hosts including immune escape. This indicates that S. iniae has the genetic repertoire to adapt very well to many different hosts.
2.8. Streptococcus marimammalium
S. marimammalium was first isolated from the lung of a dead harbor seal and a dead grey seal in Iverness, Scotland [51]. In 2007/2008, it was also isolated (together with S. agalactiae and S. canis) from nasal and oral swabs of two healthy Hawaiian monk seals from the Waikiki Aquarium, Honolulu, Hawaii [68]. In 2016, it was also isolated from South American Fur Seal Pups with moderate to marked, multifocal, mucopurulent bronchopneumonia on Guafo Island, Chile, South America [98]. To our knowledge, nothing is known about virulence factors and pathogenicity of S. marimammalium.
2.9. Streptococcus mitis
S. mitis is mainly known as member of the human oral cavity [134,135] and as opportunistic pathogen causing endocarditis and bloodstream infections in neutropenic and immunocompromised patients [136,137,138]. It is closely related to the human pathogen S. pneumoniae and its genome contain virulence genes involved in colonization and adherence, which might also be important for commensals to interact with host cells [139]. However, genes for hyaluronidase and capsular genes were absent.
S. mitis was isolated in 1985 from a blowhole swab of a captive, healthy white whale (Delphinapterus leucas) 139 days after captivity at the Mystic Marinelife Aquarium Connecticut, USA [140]. In 1985, it was isolated from lesions of a grey seal with peritonitis in North Rona, Scotland [62] and between 2012–2014 from clinically normal eyes of two grey seals stranded on the British coast [67]. These findings suggest that S. mitis might also be a commensal in some marine mammals. The commensalism and pathogenesis of S. mitis is reviewed by Mitchel, 2011 [141].
2.10. Streptococcus phocae
S. phocae was first isolated and identified from lung, liver, spleen, and kidney samples of harbor seals suffering from pneumonia with areas of consolidation, purulent exudate in the bronchi, interlobular edema, and emphysema during a phocine distemper virus outbreak in northwestern Europe [34]. Later, S. phocae was also isolated from diseased Atlantic salmon [142,143], stranded southern sea otters [144], and as gut commensal of Indian white shrimp [145]. Two subspecies are described, S. phocae subsp. salmonis for isolates from Atlantic salmon and S. phocae subsp. phocae for isolates from seals [146].
In August 1994, beta-hemolytic streptococci with high similarity to S. phocae were isolated from spleen, liver, and kidney of Cape fur seals at Cape Cross, Namibia that suffered from respiratory infections and abortions [99]. A total of 69 S. phocae isolates were obtained from harbor and grey seals from the North and the Baltic Sea investigated between 1995 and 2000 [106]. A study on phocid seals (harbor and grey seals) that were older than 19 months from the North Sea of Schleswig-Holstein, Germany reported two S. phocae isolates from intestines of phocid seals with intestinal displacements [147]. During diagnostic evaluation by the Animal Health Center, Abbotsford, British Columbia, Canada S. phocae was isolated from harbor seals with an increase of prevalence since 2000, ringed seal (P. hispida) pups from arctic Canada and two stranded harbor porpoises from Washington State [48]. In spring and summer 2000, more than 10,000 Caspian seals (Pusa caspica) were found dead with canine distemper virus infection as primary diagnosis [47]. The investigated animals suffered from broncho-interstitial pneumonia, lymphocytic necrosis and depletion in lymphoid organs, and the presence of typical intracytoplasmic inclusion bodies in multiple epithelia. S. phocae was isolated from three of eight animals. Between 2001 and 2003, vaginal and preputial swabs of California Sea Lions were collected for investigations of genital bacterial infections and urogenital carcinoma [37]. S. phocae was isolated from three specimen of cancer and three specimens of non-cancer animals stranded along the central and northern California coast. In November 2007, a short-beaked common dolphin (Delphinus delphis) stranded at La Graciosa, Canary Islands [45]. Diagnostic evaluation revealed bacterial septicemia, fibrino-necrotizing to pyogranulomatous dermatitis and panniculitis, embolic pneumonia, neutrophilic and lymphoplasmacytic meningo-choroiditis, random neutrophilic hepatitis, lymphoplasmacytic myocarditis and epicarditis, necrotizing adrenalitis, suppurative endometritis, and multicentric reactive lymphadenopathy cutaneous purulent nodules in the tail fluke, vegetative mitral valve endocarditis, and presumed postpartum pyometra. S. phocae could be cultured from lung, brain, and adrenal gland tissue. Morbillivirus was detected in the epithelium of the choroid plexus of the fourth ventricle. In November 2009, a female spotted seal (Phoca largha) stranded at Kotzebue Sound, Alaska and was diagnosed with pyometra [148]. S. phocae was isolated from the purulent discharge in uterine contents. Three Navy bottlenose dolphins (T. truncatus) developed in the time between 2009 and 2010 a strangles-like syndrome associated with S. phocae, which was isolated after the animals showed clinical signs such as inflammatory hemogram, neutrophilic leukocytosis, and unilateral cervical lymphadenopathy [149]. Between 2004 to 2010 S. phocae could be isolated from five harbor seal pups of the Smith Island in Washington, USA that were also tested positive for phocine herpes virus [32]. S. phocae was also isolated from five cases of bacterial septicemia of white whales stranded in St. Lawrence Estuary between 1983 to 2012 [33]. Necropsy of a total of 241 harbor porpoises stranded at the eastern Pacific and western Atlantic coasts of Canada between 1988 to 2011 revealed bacterial septicemia with S. phocae isolation [150]. In winter 2012, an adult female Stellar sea lion stranded in South Korea and S. phocae was cultured from the liver [116]. The cause of death was unknown. During 85 postmortem investigations of marine mammals of the northeastern Pacific and arctic Canada stranded between 2007–2012 resulted in S. phocae isolates from harbor seals (n = 61), ringed seals (n = 5), harbor porpoises (n = 5), California sea lion (n = 7), Stellar sea lion (n = 3), Guadalupe fur seal (Arctocephalus twonsendii, n = 1) and elephant seal (Mirounga angustirostris, n = 1) [151]. Sequencing of 16S rRNA V4 hyper variable regions of blow samples collected from four wild healthy Indo-Pacific bottlenose dolphins (T. aduncus) in Shark Bay (SB), Western Australia, in 2012 identified S. phocae (and S. equi) [115]. In February 2014, S. phocae was isolated from a carcass of a subadult male northern seal at the coast of Niigata, Japan that suffered from necrotizing and fibrinous pneumonia with diffuse abscesses of all lung lobes and massive necrosis of kidney and liver [46]. Between 2010 to 2017 the health of captive and stranded Alaskan ice seals were investigated and S. phocae isolates were obtained from blood, abscess, and lymph node samples from ringed seals [152]. Harbor seals stranded at the coast of San Juan County, Washington, USA between 2002 to 2018 were examined and from one adult female animal a fatal septicemia caused by S. phocae was reported [153].
While the presence of an antiphagocytic capsule and virulence of S. phocae subsp. salmonis to Atlantic salmon has been demonstrated in infectivity experiments [142,154,155], whole genome analyses of S. phocae subsp. phocae identified typical streptococcal virulence factors such as fibronectin-binding proteins, the toxin streptolysin S and genes encoding for a capsule [156]. Invasion of fish and mammalian cell lines by S. phocae subsp. phocae has also been shown and confirmed its pathogenic potential [154].
However, S. phocae subsp. phocae also seems to be a commensal of the oral cavity of grey seals as revealed by microbiome analyses and 16S rRNA sequencing. A transmission of S. phocae to harbor porpoises via bites is also indicated [157] and S. phocae might be an opportunistic pathogen, at least for seals.
2.11. Streptococcus viridans Group
In very few studies, streptococci isolated from marine mammals were identified as members of the S. viridans group (viridans streptococci), which includes streptococci that are usually alpha-hemolytic and inhabit the oral cavity, intestinal, and vaginal tract [158,159,160]. This group is very heterogeneous and includes species such as S. anginosus, S. mitis, S. sanguinis, S. mutans, and S. salivarius, which can also cause endocarditis [161], bacteremia [162], and respiratory infections [163].
Viridans streptococci were isolated from superficial abscesses, wounds, ocular and urethral discharges, and umbilici of live and from lung and liver samples of dead elephant seals, California sea lions and harbor seals stranded between January 1994 and December 1995 along the California Coast [164]. Viridans streptococci were isolated in mixed cultures with Arcanobacterium phocae from California sea lions, harbor seals, Northern elephant seals, sea otter and common dolphin stranded along the central California coast between 1994 and 2000 [165]. In Beluga whales that stranded at Cook Inlet (Alaska, USA) between 1998 and 2013 an isolate was identified as member of the S. viridans group [166]. Also, viridans streptococci were isolated from gastric fluid samples of free-ranging bottlenose dolphins from the southeastern United States during a catch and release health assessment between 2003 to 2005 [167].
2.12. One-Time Only Detections of Streptococcal Species from Marine Mammals
In studies described above, streptococcal species have been isolated and identified at least twice or more. In the following, reports on one-time only descriptions of streptococcal species are summarized.
S. uberis was found in dead free-ranging male Antarctic fur seals with pneumonia and extensive polymorphonuclear infiltrations and necrosis or very widespread abscess formation and frequently there was an associated fibrinous exudative pleurisy [63]. S. oralis was isolated and identified by API strips from three swabs taken from healthy eyes of free-ranging grey seals stranded on the British coast between November 2012 and February 2014 [67]. In a metagenome dataset of blood, muscle, and fecal samples of a living stranded sperm whale (Physeter catodon) S. anginosus, S. pneumoniae, and S. suis were detected in blood and fecal samples, but not in the muscles [168]. The animal died 79 h after rescue. S. intermedius was detected in blow samples of free-ranging and presumably healthy grey whales from Magdalena Bay and the Gulf of California by polymerase chain reaction [169].
3. Streptococcal Infections in Marine Mammals: Virulence and Mechanims of Pathogenicity
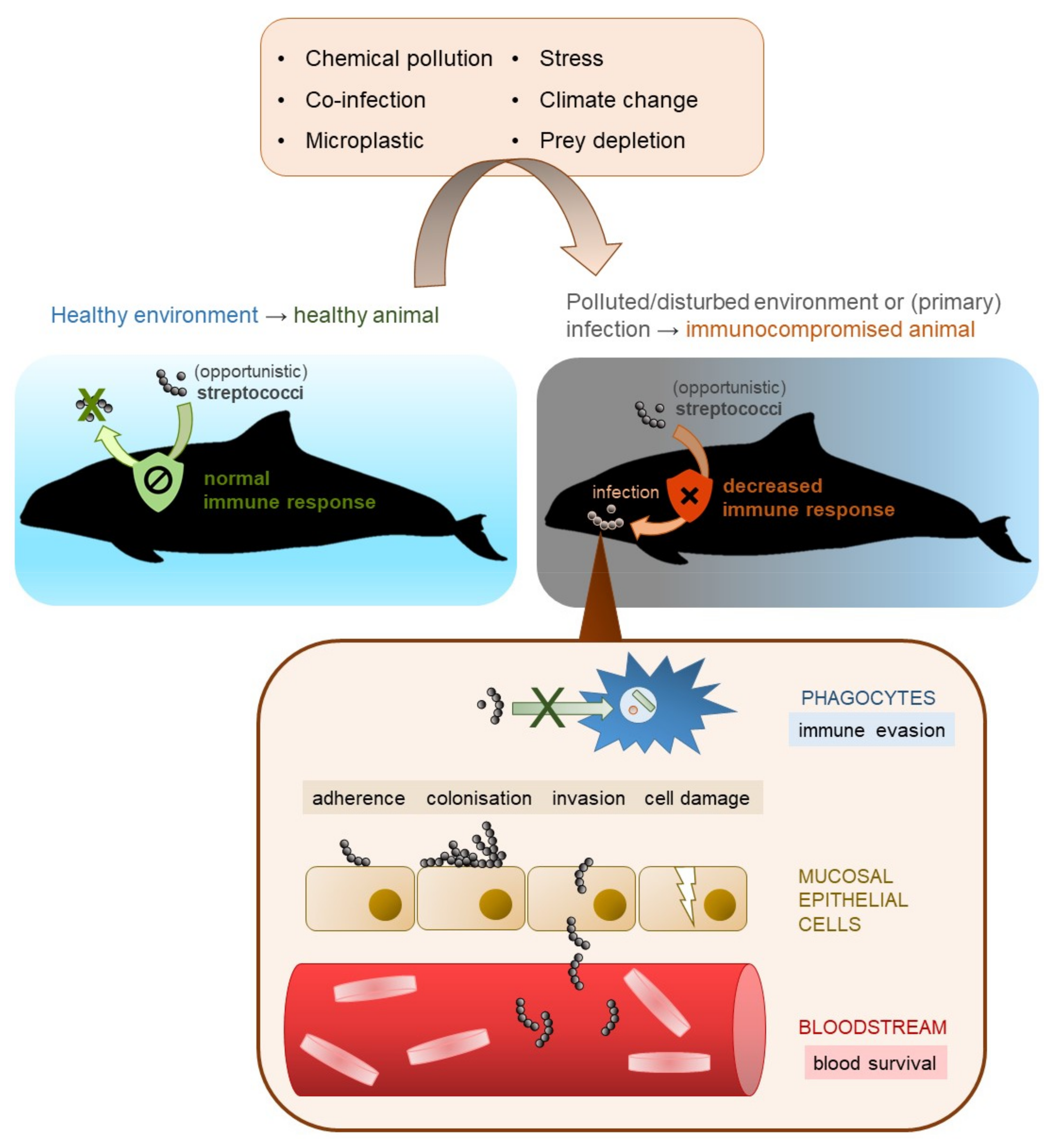
Streptococci are a phylogenetically diverse group and, hence, their virulence and infection mechanisms differ intensely. A general idea of streptococcal infection in (marine) mammals is displayed in Figure 2. One important requirement for a successful infection is the resistance of pathogenic streptococci to host phagocytosis. The major antiphagocytic factors are the polysaccharide capsule, which is also the basis for serotyping [127,170,171,172,173] and the streptococcal M protein of pyogenic streptococci [43,174]. The most critical phase in infection is the adhesion of streptococci to host cells. This is enabled by pili and/or adhesins such as fibronectin- and collagen-binding proteins and sortases [175,176]. It has also been demonstrated that streptococcal species are able to invade host cells [154,177,178] and produce toxins such as streptolysin S and streptolysin O [179,180]. There are numerous other virulence factors, for instance, streptokinases, secretory proteins that interacts with host plasminogen [181], or peptidoglycan deacetylases that protects the bacteria from host lysozymes [130,182]. To investigate the molecular mechanisms of streptococcal infection in marine mammals, primary cell cultures of marine mammals are required that can be infected and evaluated.
Many streptococci occur as opportunistic pathogens or as secondary infection [45,183,184,185,186,187]. This might also be the case for marine mammals. S. agalactiae, S. canis and S. marimammalium were found in the nasal cavity of two captured, healthy Hawaiian monk seals (M. schauinslandi) without any clinical signs [68]. A short-beaked common dolphin (Delphinus delphis) was coinfected by Streptococcus phocae and cetacean morbillivirus indicating S. phocae as secondary infection [45]. 16S rRNA sequencing of blow samples collected from four wild and healthy Indo-Pacific bottlenose dolphins (Tursiops aduncus) in Shark Bay, Western Australia identified S. phocae, S. equi and Streptococcus Group D as members of the blow microbiome without clinical signs [115].
Weakened or immunocompromised animals are more susceptible for infectious disease, even caused by opportunistic pathogens. The marine environment is threatened by climate change [16,17], ocean acidification [13,14,15], and pollutants (5,6,22,24), which can influence the immune system of marine mammals negatively [21,22,23,24]. This can result in an increase of infectious disease, which might also be true for streptococcal species [23].
The development of an infectious disease depends on the successful infection of a pathogen and on its virulence. This also includes the ability to evade host immune defense. Streptococci have evolved many mechanisms of immune invasion. For instance, the pore-forming toxin streptolysin O induces caspase-dependent macrophage apoptosis [188] and the gene sets for streptolysin S, which is responsible for the beta-hemolysis, was also found in S. phocae subsp. phocae [156]. The human pathogenic S. pyogenes can recruit and colonize collagen type IV via surface-bound fibronectin, and the collagen fibers protect the bacterial cells from opsonizing antibodies [189]. Proteins that interact with collagen were also found in other streptococcal species including S. phocae [190]. Group A streptococci (e.g., S. pyogenes) can bind red blood cells by S protein for immune evasion [191]. In S. phocae subsp. phocae, an immunoglobulin G degrading enzyme, called ideP, was identified, which cleaves IgG of seals and thus, contributes to immune evasion [192]. Hence, streptococci that infect marine mammals requires specific adaptation to their immune system, as only immune evasion guarantee virulence and infection. The immune system of marine mammals is similar to other (terrestrial) mammals regarding general mechanisms [193]. In grey seals, the main type of immunoglobulin was IgG with two subclasses [194]. Furthermore, the authors discussed whether the susceptibility for bacterial infections including streptococcal infections in grey seal pups is related to the observed low values of IgG in pup serum in comparison to the relatively high values in the colostrum. Immunoglobulin classes homologous to human IgG, IgM, and IgA were identified also in dolphins and sea lions [195] indicating that streptococci face similar conditions and molecules when jumping from terrestrial to marine mammals.
One difference in streptococcal infection between marine and terrestrial mammals could be the fact that plasminogen from marine mammals could be activated by human plasma including urokinase, but not by streptokinase vaccine [196,197]. Moreover, it is known that the genes for streptokinase of group A streptococci showed a high heterogeneity even in strains with the same serotype indicating immunological and chemical differences [198]. We did not find any information on streptokinases in streptococci of marine mammals and, hence, it can only be speculated if they just lost the gene or if there a genetic variations reflecting host adaptability.
4. Adaptation of Streptococci to Marine Mammals
Many streptococcal species found in marine mammals are also present in terrestrial mammals, although there are some physiological differences. That raises the question, to what extend and how these streptococci adapt to their specific marine host environments. Since this has not been studied yet in any detail here, we can only speculate and discuss it.
The body temperature of marine mammals is comparable to that of terrestrial mammals [199,200,201,202,203], thus, it does not require special adaptations. However, gas physiology and rapid change in hydrostatic pressure during diving might be a challenge for the bacteria. The hydrostatic pressure of seawater increases about 0.1 atmosphere per meter of depth. It has been shown that hydrostatic pressure can influence the growth and viability of marine and terrestrial bacteria negatively [204,205]. At least the closely related Lactococcus lactis (previously S. lactis) was able to survive 500 atmosphere representing around 5000 m depth, while many other bacteria including species of Bacillus, Clostridium, and Staphylococcus died at pressure of 400 or less atmospheres [205]. In conclusion, streptococcal species and other bacteria might not have a problem with the hydrostatic pressure that increases during diving of marine mammals. However, bacteria of marine mammals, depending on which body part they inhabit (e.g., skin, intestines, respiratory tract) and if they were acquired from the marine environment, have to challenge the salinity of seawater. Seawater has an average salinity of 3.5%, but salt tolerance of streptococcal species is usually tested at 6.5% NaCl and 40% bile salts, respectively, [34,142,206,207]. The salt tolerance of S. iniae was tested at 2.0%, 4.0%, and 6.5% NaCl and growth was observed for the first two conditions [122]. In addition, few S. thermophilus strains and S. uberis grow at 4.0% NaCl, but not at 6.5% [208,209]. The fish pathogen S. parauberis is known to persist in seawater, probably by switching to dormancy as a survival strategy [210]. Therefore, it is also possible for other streptococcal species to persist and maybe even grow in seawater.
Streptococci infecting marine mammals often use the respiratory tract as port of entry for colonization infection. Thus, it is likely that they have adapted to this special host niche. Notably, the respiratory tract differs between terrestrial and marine mammals, e.g., in anatomy, immune response, gas physiology (O2/CO2 exchange), humidity, and chemical composition of the mucus. For instance, it has been shown that lung surfactant of pinnipeds has higher amounts of anti-adhesive components compared to terrestrial mammals, which probably supports alveolar opening after collapse during diving [211].
To our knowledge, there are no specific studies on genetic adaptation of streptococcal species to their marine mammalian host. However, whole genome analyses of human and fish/frog S. agalactiae strains revealed genetic adaptation to fish host by gene reduction and different gene expression such as of virulence associated genes [212]. A comparative genomic study of S. dysgalactiae species suggest that changes in gene content, selection of orthologous protein-coding loci and operon promoters involving mobile elements enables streptococci to adapt to changing environments and new hosts [213]. S. phocae subsp. phocae showed host specificity by the immunoglobulin G degrading enzyme ideP that solely cleaves IgG of grey and harbor seals, but not from harbor porpoises or non-marine mammals indicating functional adaptation [192]. S. halichoeri is assumed to have marine origin, although it has also been isolated from dog, bluefox, finnraccoon, and mink, as it has a great number of adhesins and salt tolerance proteins [120]. Lateral gene transfer between different streptococcal species is discussed as potential way of host adaptation [214]. For instance, lateral gene transfer was observed between S. canis and human S. urinalis and bovine S. agalctiae and S, dysgalactiae subsp. dysgalactiae mediated by variety of mobile genetic elements. This might also be true for marine mammals, where streptococci adapted to the marine mammalian host such as S. phocae might exchange genetic elements with acquired streptococcal species from fish or other sources. Further studies are required to understand the adaptation and interaction of streptococcal species and their marine mammal host as there is a huge scientific gap.
5. Epidemiology and Possible Transmission Routes of Streptococci Species in Marine Mammals
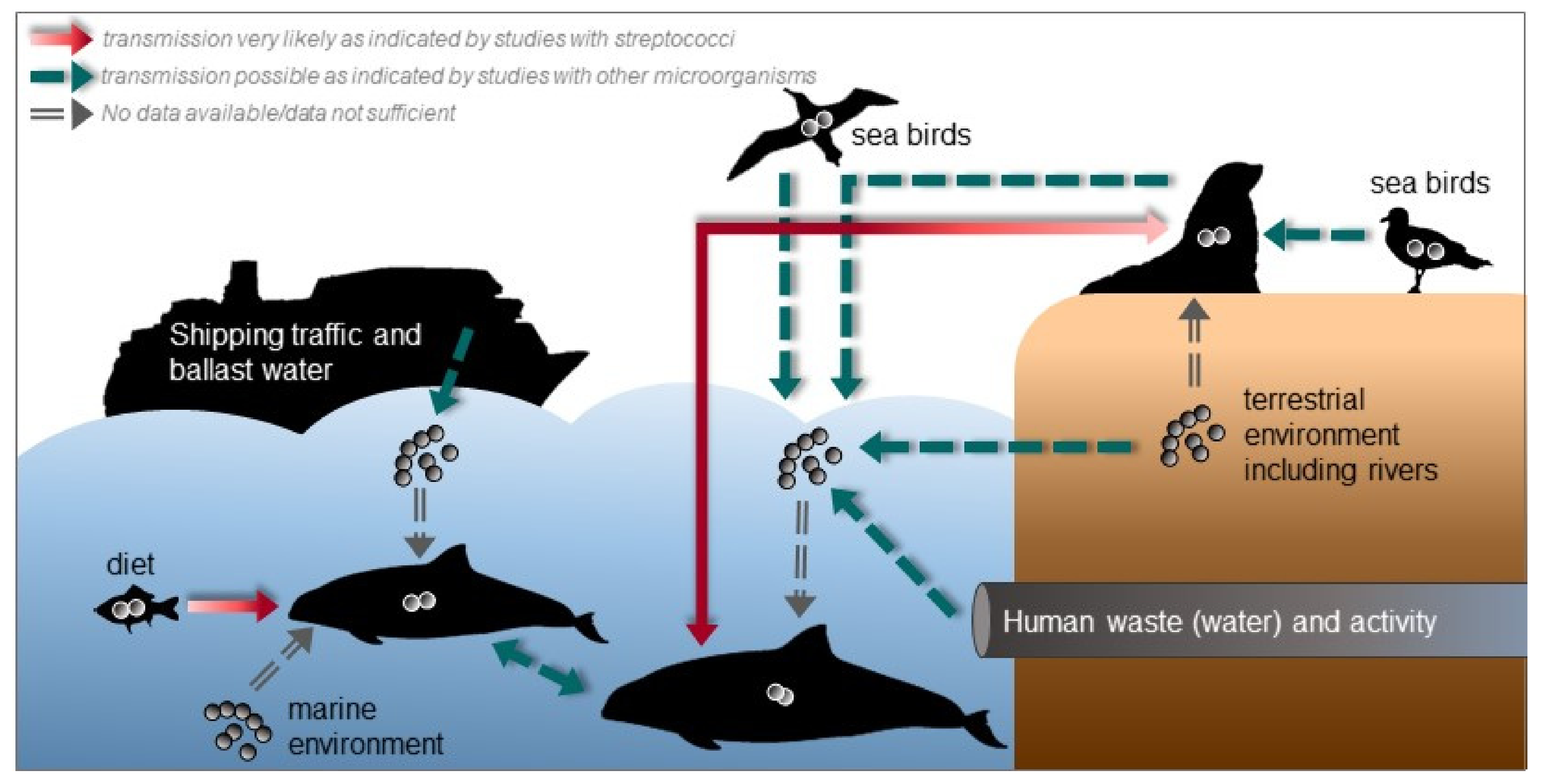
It is not known how marine mammals acquire or get infected with streptococci. Studies on the ecology and the environment could provide some insights about possible transmission routes and the epidemiology of streptococcal infections in marine mammals. In Figure 3, we give an overview about possible transmission routes and sources for streptococcal species. With many social species among marine mammals, transmission of pathogens are density- or frequency-dependent according to their social behavior [215,216].
The diet, e.g., fish could serve as reservoir for (pathogenic) streptococcal species and consequently as source of infection for marine mammals, which is also indicated by the study of Evans et al. [65], where the same S. agalactiae strain was found in both a dead dolphin and its diet, a mullet which was found in its stomach. S. iniae and S. agalactiae were also isolated from wild fish indicating fish as potential source for streptococci [217,218]. In addition, S. halichoeri is supposed to be transmitted to mink and Finnish dogs via fed fish [120]. S. phocae is also a well-known pathogen of Atlantic salmon [143]. However, the fish isolates differ from isolates of marine mammals, which was also the reason to suggest two subspecies of S. phocae [146]. The genetic differences could be a result of adaptation from fish to the mammalian host.
Pathogenic streptococci and other bacteria could be introduced into seawater by shipping traffic including ballast water [219,220], human activities including recreational activity [221,222,223,224,225] and waste (water) [66,226,227,228,229] and from the terrestrial environment via rivers and storm water [230,231], wind transport [232,233] or animals, e.g., semi-aquatic mammals, such as pinnipeds, from which streptococci are frequently isolated. For instance, human pathogenic S. agalactiae strains were identified in grey seals indicating that sea mammals were exposed to human pathogens via human effluents that contaminate coastal surface waters [66]. S. phocae has been found in the oral cavity of grey seals and in bite wounds of harbor porpoises probably caused by grey seals suggesting an interspecies transmission [157]. Seabirds can also shed pathogenic organisms into seawater [228,234,235] and streptococci were also detected in the gastrointestinal tract of seabirds [236]. If these transmission routes are real, streptococcal contamination and infections might increase with increasing human activities including growing cities in coastal regions and higher rates of shipping traffic, e.g., an increase of pollution and a higher likelihood of ship strikes or fisheries interactions. In addition, with higher temperatures due to climate change the persistence of pathogens in seawater is probably enhanced and, thus, there is a higher risk of infection [17,215]. The mortality of tilapias due to S. agalactiae infection was increased in higher water temperature [237]. This could also be the case for marine mammals, but there are no studies yet. In addition, habitat loss can enhance transmission by leading to higher population densities with higher contact rates.
To our knowledge streptococci are no or not abundant members of the natural marine microbiome [238]. This raises the question, where they come from. Nevertheless, streptococci that are introduced by the routes mentioned above might cause dermal disease or enter marine mammals through open wounds and other traumata. Díaz-Delgado et al. [45] proposed that the S. phocae infection in a short-beaked common dolphin occurred through cutaneous penetration after a skin traumata, as the dolphin showed cutaneous disease with firm, raised, and occasionally ulcerated purulent subcutaneous nodules along the ventral, dorsal, and cranial edge of the caudal fluke, bilaterally, and more prominently at its insertion with the peduncle.
However, studies on the distribution and presence of streptococci in healthy animals including the screening of the environment are necessary to investigate possible transmission routes or confirm these streptococcal species as commensals or members of the normal microbiome.
6. Conclusions and Outlook
Taken together, this is an overview about streptococcal species that were identified in marine mammals. Streptococcal species play an important role in the health of marine mammals all over the world. However, while beta-hemolytic streptococci are frequently isolated from marine mammals, only relatively few isolates were further identified (to the species level). There are further marine mammals from which (beta-hemolytic) streptococci have been isolated, but not identified to the species level, such as the blue whale (Balaenoptera musculus) [239], the gray whale (Eschrichtius robustus) [239], the sperm whale (Physeter macrocephalus) [240], the killer whale (Orcinus orca) [241], and the pacific walrus (Odobenus rosmarus divergens) [242]. This scientific gap makes it difficult to evaluate the diversity, distribution, and epidemiology of streptococcal species among marine mammals and needs to be filled. In addition, while there are lineages of streptococcal species that are quite host-specific, there are others that seem to have a more groad host spectrum and are easily transmissible between different hosts. For instance, in addition to marine mammals, S. canis can be found in dogs and cows [86], S. halichoeri in humans [119], and S. phocae in Atlantic salmon [143] and shrimps [145]. For some of these species, subspecies were defined based on their host-related differences such as S. phocae subsp. phocae and S. phocae subsp. salmonis [146] or S. halichoeri subsp. halichoeri and S. halichoeri subsp. hominis [119]. Furthermore, some of the streptococcal species found in marine mammals are major fish pathogens such as S. agalactiae [243], S. iniae [244], and S. phocae [143] and even zoonotic infections are possible, but the lack of data does not allow clear risk assessments. S. iniae caused infections in humans that handled live or freshly killed fish [58,245]. S. canis infections in humans are summarized by Galpérine et al. [56] and an endocarditis due to S. canis has been reported by Ansallem et al. [246]. S. equi subsp. zooepidemicus is also known to cause zoonotic disease in humans [53,247]. Hence, people working with marine mammals should also be aware of the zoonotic potential of streptococcal species.
It is also not fully understood how the different streptococcal species are involved in diseases of marine mammals, although they are frequently isolated from sick or dead animals. With increasing chemical pollution and other anthropogenic activities in the marine ecosystem, the health of marine mammals is threatened and thus, they are more susceptible to infectious diseases. Hence, more research is needed on the epidemiology and pathogenic potential of streptococcal species in marine mammals.
In conclusion, streptococcal species are isolated from many different marine mammal species world-wide. Further investigations on the role of the different streptococci species on the health status of marine mammals is urgently needed as streptococci are found with high prevalence in diseased marine mammals. This also underlines the need of additional information on the zoonotic potential of streptococci species found in marine mammals.
Supplementary Materials
The following is available online at https://0-www-mdpi-com.brum.beds.ac.uk/2076-2607/9/2/350/s1, Table S1: Prevalence of isolated and identified streptococcal species in marine mammals.
Author Contributions
Conceptualization, D.N., P.V.-W., M.F., and U.S.; writing—original draft preparation, D.N.; writing—review and editing, D.N., P.V.-W., M.F., and U.S.; visualization, D.N.; supervision, P.V.-W., M.F., and U.S.; project administration, D.N., P.V.-W., M.F., and U.S.; funding acquisition, D.N. and P.V.-W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the Deutsche Forschungsgemeinschaft (DFG), Bonn, Germany (NU 422/1).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This publication was supported by Deutsche Forschungsgemeinschaft, Bonn, Germany, and the University of Veterinary Medicine Hannover, Foundation within the funding programme Open Access Publishing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shuval, H. Estimating the global burden of thalassogenic diseases: Human infectious diseases caused by wastewater pollution of the marine environment. J. Water Health 2003, 1, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Dorsch, D.E.; Bay, S.M.; Maruya, K.; Snyder, S.A.; Trenholm, R.A.; Vanderford, B.J. Contaminants of emerging concern in municipal wastewater effluents and marine receiving water. Environ. Toxicol. Chem. 2012, 31, 2674–2682. [Google Scholar] [CrossRef] [PubMed]
- Nelms, S.E.; Barnett, J.; Brownlow, A.; Davison, N.J.; Deaville, R.; Galloway, T.S.; Lindeque, P.K.; Santillo, D.; Godley, B.J. Microplastics in marine mammals stranded around the British coast: Ubiquitous but transitory? Sci. Rep. 2019, 9, 1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tornero, V.; Hanke, G. Chemical contaminants entering the marine environment from sea-based sources: A review with a focus on European seas. Mar. Pollut. Bull. 2016, 112, 17–38. [Google Scholar] [CrossRef] [PubMed]
- Desforges, J.-P.; Hall, A.; McConnell, B.; Rosing-Asvid, A.; Barber, J.L.; Brownlow, A.; Guise, S.D.; Eulaers, I.; Jepson, P.D.; Letcher, R.J.; et al. Predicting global killer whale population collapse from PCB pollution. Science 2018, 361, 1373–1376. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.J.; Soto, N.A.; Baldwin, A.L.; Bateson, M.; Beale, C.M.; Clark, C.; Deak, T.; Edwards, E.F.; Fernández, A.; Godinho, A.; et al. Do marine mammals experience stress related to anthropogenic noise? Int. J. Comp. Psychol. 2007, 20. Available online: https://escholarship.org/uc/item/6t16b8gw (accessed on 12 August 2020).
- La Cruz, D.B.-D.; DeRango, E.; Johnson, S.P.; Simeone, C.A. Evidence of anthropogenic trauma in marine mammals stranded along the central California coast, 2003–2015. Mar. Mammal Sci. 2018, 34, 330–346. [Google Scholar] [CrossRef]
- Bearzi, G.; Politi, E.; Agazzi, S.; Azzellino, A. Prey depletion caused by overfishing and the decline of marine megafauna in eastern Ionian Sea coastal waters (central Mediterranean). Biol. Conserv. 2006, 127, 373–382. [Google Scholar] [CrossRef]
- Rolland, R.M.; McLellan, W.A.; Moore, M.J.; Harms, C.A.; Burgess, E.A.; Hunt, K.E. Fecal glucocorticoids and anthropogenic injury and mortality in North Atlantic right whales Eubalaena glacialis. Endanger. Species Res. 2017, 34, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Bexton, S.; Thompson, D.; Brownlow, A.; Barley, J.; Milne, R.; Bidewell, C. Unusual mortality of pinnipeds in the United Kingdom associated with helical (corkscrew) injuries of anthropogenic origin. Aquat. Mamm. 2012, 38, 229–240. [Google Scholar] [CrossRef]
- Fire, S.E.; Wang, Z.; Byrd, M.; Whitehead, H.R.; Paternoster, J.; Morton, S.L. Co-occurrence of multiple classes of harmful algal toxins in bottlenose dolphins (Tursiops truncatus) stranding during an unusual mortality event in Texas, USA. Harmful Algae 2011, 10, 330–336. [Google Scholar] [CrossRef]
- Fire, S.E.; Dolah, F.V. Marine Biotoxins: Emergence of Harmful Algal Blooms as Health Threats to Marine Wildlife Publications, Agencies and Staff of the U.S. Department of Commerce. 2012. Available online: https://digitalcommons.unl.edu/usdeptcommercepub/553 (accessed on 12 August 2020).
- Hofmann, G.E.; Barry, J.P.; Edmunds, P.J.; Gates, R.D.; Hutchins, D.A.; Klinger, T.; Sewell, M.A. The effect of ocean acidification on calcifying organisms in marine ecosystems: An organism-to-ecosystem perspective. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 127–147. [Google Scholar] [CrossRef]
- Brewer, P.G.; Hester, K. Ocean acidification and the increasing transparency of the ocean to low-frequency sound. Oceanography 2009, 22, 86–93. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, S.; Ishida, A.; King, R.; Raymond, B.; Waller, N.; Constable, A.; Nicol, S.; Wakita, M.; Ishimatsu, A. Risk maps for Antarctic krill under projected Southern Ocean acidification. Nat. Clim. Chang. 2013, 3, 843–847. [Google Scholar] [CrossRef]
- Moore, S.E.; Huntington, H.P. Arctic marine mammals and climate change: Impacts and resilience. Ecol. Appl. 2008, 18, S157–S165. [Google Scholar] [CrossRef] [Green Version]
- Sanderson, C.E.; Alexander, K.A. Unchartered waters: Climate change likely to intensify infectious disease outbreaks causing mass mortality events in marine mammals. Glob. Chang. Biol. 2020, 26, 4284–4301. [Google Scholar] [CrossRef] [PubMed]
- Bossart, G.D. Marine mammals as sentinel species for oceans and human health. Vet. Pathol. 2011, 48, 676–690. [Google Scholar] [CrossRef] [Green Version]
- Moore, S.E. Marine mammals as ecosystem sentinels. J. Mammal 2008, 89, 534–540. [Google Scholar] [CrossRef]
- Ross, P.S. Marine mammals as sentinels in ecological risk assessment. Hum. Ecol. Risk Assess. Int. J. 2000, 6, 29–46. [Google Scholar] [CrossRef]
- Desforges, J.-P.W.; Sonne, C.; Levin, M.; Siebert, U.; De Guise, S.; Dietz, R. Immunotoxic effects of environmental pollutants in marine mammals. Environ. Int. 2016, 86, 126–139. [Google Scholar] [CrossRef]
- Kakuschke, A.; Prange, A. The influence of metal pollution on the immune system a potential stressor for marine mammals in the North Sea. Int. J. Comp. Psychol. 2007, 20. Available online: https://escholarship.org/uc/item/55p4w9tj (accessed on 6 July 2020).
- Ross, P.S. The role of immunotoxic environmental contaminants in facilitating the emergence of infectious diseases in marine mammals. Hum. Ecol. Risk Assess. Int. J. 2002, 8, 277–292. [Google Scholar] [CrossRef]
- Mos, L.; Morsey, B.; Jeffries, S.J.; Yunker, M.B.; Raverty, S.; Guise, S.D.; Ross, P.S. Chemical and biological pollution contribute to the immunological profiles of free-ranging harbor seals. Environ. Toxicol. Chem. 2006, 25, 3110–3117. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.J.; Hugunin, K.; Deaville, R.; Law, R.J.; Allchin, C.R.; Jepson, P.D. The Risk of infection from polychlorinated biphenyl exposure in the harbor porpoise (Phocoena phocoena): A case–control approach. Environ. Health Perspect. 2006, 114, 704–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, J.C.; Ridgway, S.H. Common diseases of small cetaceans. J. Am. Vet. Med. Assoc. 1975, 167, 533–540. [Google Scholar]
- Stroud, R.K.; Roffe, T.J. Causes of death in marine mammals stranded along the oregon coast. J. Wildl. Dis. 1979, 15, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Harwood, J.; Hall, A. Mass mortality in marine mammals: Its implications for population dynamics and genetics. Trends Ecol. Evol. 1990, 5, 254–257. [Google Scholar] [CrossRef]
- Bogomolni, A.L.; Pugliares, K.R.; Sharp, S.M.; Patchett, K.; Harry, C.T.; LaRocque, J.M.; Touhey, K.M.; Moore, M. Mortality trends of stranded marine mammals on Cape Cod and southeastern Massachusetts, USA, 2000 to 2006. Dis. Aquat. Organ. 2010, 88, 143–155. [Google Scholar] [CrossRef] [Green Version]
- Siebert, U.; Wohlsein, P.; Lehnert, K.; Baumgärtner, W. Pathological findings in harbour seals (Phoca vitulina): 1996–2005. J. Comp. Pathol. 2007, 137, 47–58. [Google Scholar] [CrossRef]
- Siebert, U.; Wünschmann, A.; Weiss, R.; Frank, H.; Benke, H.; Frese, K. Post-mortem findings in harbour porpoises (Phocoena phocoena) from the German North and Baltic Seas. J. Comp. Pathol. 2001, 124, 102–114. [Google Scholar] [CrossRef]
- Huggins, J.L.; Leahy, C.L.; Calambokidis, J. Causes and patterns of harbor seal (Phoca vitulina) pup mortality at Smith Island, Washington, 2004–2010. Northwest Nat. 2013, 94, 198–208. [Google Scholar] [CrossRef]
- Lair, S.; Martineau, D.; Measures, L.N. Causes of Mortality in St. Lawrence Estuary Beluga (Delphinapterus leucas), from 1983 to 2012; Canadian Science Advisory Secretariat: Ottawa, ON, Canada, 2014. [Google Scholar]
- Skaar, I.; Gaustad, P.; Tønjum, T.; Holm, B.; Stenwig, H. Streptococcus phocae sp. nov., a new species isolated from clinical specimens from seals. Int. J. Syst. Evol. Microbiol. 1994, 44, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Higgins, R.; Claveau, R.; Roy, R. Bronchopneumonia caused by Streptococcus equi in a North Atlantic pilot whale (Globicephala melaena). J. Wildl. Dis. 1980, 16, 319–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swenshon, M.; Lämmler, C.; Siebert, U. Identification and molecular characterization of beta-hemolytic streptococci isolated from harbor porpoises (Phocoena phocoena) of the North and Baltic Seas. J. Clin. Microbiol. 1998, 36, 1902–1906. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.; Lowenstine, L.; Gulland, F.; Jang, S.; Imai, D.; Almy, F.; DeLong, R.; Gardner, I. Aerobic bacterial flora of the vagina and prepuce of California sea lions (Zalophus californianus) and investigation of associations with urogenital carcinoma. Vet. Microbiol. 2006, 114, 94–103. [Google Scholar] [CrossRef]
- Hardie, J.M.; Whiley, R.A. The genus Streptococcus. In The Genera of Lactic Acid Bacteria; Wood, B.J.B., Holzapfel, W.H., Eds.; Springer: Boston, MA, USA, 1995; pp. 55–124. [Google Scholar] [CrossRef]
- du Toit, M.; Huch, M.; Cho, G.-S.; Franz, C.M.A.P. The genus Streptococcus. In The Lactic Acid Bacteria; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1995; pp. 457–505. [Google Scholar] [CrossRef]
- Tagg, J.R.; Wescombe, P.A.; Burton, J.P.; Wescombe, P.A. Streptococcus: A brief update on the current taxonomic status of the genus. In Lactic Acid Bacteria; Lahtinen, S., Ouwehand, A.C., Salminen, S., von Wright, A., Eds.; CRC Press: Boca Raton, FI, USA, 2011. [Google Scholar] [CrossRef]
- Facklam, R. What happened to the streptococci: Overview of taxonomic and nomenclature changes. Clin. Microbiol. Rev. 2002, 15, 613–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellen, R.P.; Gibbons, R.J. M protein-associated adherence of Streptococcus pyogenes to epithelial surfaces: Prerequisite for Virulence. Infect. Immun. 1972, 5, 826–830. [Google Scholar] [CrossRef] [Green Version]
- Horstmann, R.D.; Sievertsen, H.J.; Knobloch, J.; Fischetti, V.A. Antiphagocytic activity of streptococcal M protein: Selective binding of complement control protein factor H. Proc. Natl. Acad. Sci. USA 1988, 85, 1657–1661. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.Y.H.; Nizet, V. Extracellular virulence factors of group B streptococci. Front. Biosci. 2004, 9, 1794–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Delgado, J.; Sierra, E.; Vela, A.I.; Arbelo, M.; Zucca, D.; Groch, K.R.; Fernández, A. Coinfection by Streptococcus phocae and Cetacean morbillivirus in a short-beaked common dolphin Delphinus delphis. Dis. Aquat. Organ. 2017, 124, 247–252. [Google Scholar] [CrossRef]
- Iwao, H.; Yanagisawa, M.; Kino, S.; Takamori, J.; Okamoto, M. Two beta-hemolytic streptococci Streptococcus canis and S. phocae isolated from a northern fur seal with septicemia from Niigata, Japan. In Proceedings of the 46th Annual Conference of the International Association for Aquatic Animal Medicine, Chicago, IL, USA, 6–10 April 2015; Available online: https://www.vin.com/doc/?id=7009572 (accessed on 6 July 2020).
- Kuiken, T.; Kennedy, S.; Barrett, T.; Van de Bildt, M.W.G.; Borgsteede, F.H.; Brew, S.D.; Codd, G.A.; Duck, C.; Deaville, R.; Eybatov, T.; et al. The 2000 canine distemper epidemic in caspian seals (Phoca caspica): Pathology and analysis of contributory factors. Vet. Pathol. 2006, 43, 321–338. [Google Scholar] [CrossRef] [Green Version]
- Raverty, S.A.; Gaydos, J.K.; Nielsen, O.; Ross, P.S. Pathologic and clinical implications of Streptococcus phocae isolated from pinnipeds along coastal Washington State, British Columbia and Arctic Canada. In Proceedings of the 35th Annual Conference of the International Association of Aquatic Animal Medicine, Galveston, TX, USA, 4–9 April 2004; Available online: https://www.vin.com/doc/?id=6696240 (accessed on 6 July 2020).
- Baker, J.R. The pathology of the grey seal (Haliochoerus grypus). II. Juveniles and adults. Br. Vet. J. 1980, 136, 443–447. [Google Scholar] [CrossRef]
- Lawson, P.A.; Foster, G.; Falsen, E.; Davison, N.; Collins, M.D. Streptococcus halichoeri sp. nov., isolated from grey seals (Halichoerus grypus). Int. J. Syst. Evol. Microbiol. 2004, 54, 1753–1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, P.A.; Foster, G.; Falsen, E.; Collins, M.D. Streptococcus marimammalium sp. nov., isolated from seals. Int. J. Syst. Evol. Microbiol. 2005, 55, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Fulde, M.; Valentin-Weigand, P. Epidemiology and pathogenicity of zoonotic streptococci. In Host-Pathogen Interactions in Streptococcal Diseases; Current Topics in Microbiology and Immunology; Chhatwal, G.S., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 49–81. [Google Scholar] [CrossRef]
- Minces, L.R.; Brown, P.J.; Veldkamp, P.J. Human meningitis from Streptococcus equi subsp. zooepidemicus acquired as zoonoses. Epidemiol. Infect. 2011, 139, 406–410. [Google Scholar] [CrossRef] [Green Version]
- Strangmann, E.; Fröleke, H.; Kohse, K.P. Septic shock caused by Streptococcus suis: Case report and investigation of a risk group. Int. J. Hyg. Environ. Health 2002, 205, 385–392. [Google Scholar] [CrossRef]
- Yu, H.; Jing, H.; Chen, Z.; Zheng, H.; Zhu, X.; Wang, H.; Wang, S.; Liu, L.; Zu, R.; Luo, L.; et al. Human Streptococcus suis outbreak, Sichuan, China. Emerg. Infect. Dis. 2006, 12, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Galpérine, T.; Cazorla, C.; Blanchard, E.; Boineau, F.; Ragnaud, J.-M.; Neau, D. Streptococcus canis infections in humans: Retrospective study of 54 patients. J. Infect. 2007, 55, 23–26. [Google Scholar] [CrossRef]
- Bert, F.; Lambert-Zechovsky, N. Septicemia caused by streptococcus canis in a human. J. Clin. Microbiol. 1997, 35, 777–779. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, M.R.; Litt, M.; Kertesz, D.A.; Wyper, P.; Rose, D.; Coulter, M.; McGeer, A.; Facklam, R.; Ostach, C.; Willey, B.M.; et al. Invasive infections due to a fish pathogen, Streptococcus iniae. N. Engl. J. Med. 1997, 337, 589–594. [Google Scholar] [CrossRef] [Green Version]
- Anthony, B.F.; Okada, D.M. The emergence of group B streptococci in infections of the newborn infant. Annu. Rev. Med. 1977, 28, 355–369. [Google Scholar] [CrossRef] [PubMed]
- Musser, J.M.; Mattingly, S.J.; Quentin, R.; Goudeau, A.; Selander, R.K. Identification of a high-virulence clone of type III Streptococcus agalactiae (group B Streptococcus) causing invasive neonatal disease. Proc. Natl. Acad. Sci. USA 1989, 86, 4731–4735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keefe, G.P. Streptococcus agalactiae mastitis: A review. Can. Vet. J. 1997, 38, 429–437. [Google Scholar]
- Baker, J.R. Further studies on grey seal (Halichoerus grypus) pup mortality on North Rona. Br. Vet. J. 1988, 144, 497–506. [Google Scholar] [CrossRef]
- Baker, J.R.; McCann, T.S. Pathology and bacteriology of adult male antarctic fur seals, Arctocephalus gazella, dying at Bird Island, South Georgia. Br. Vet. J. 1989, 145, 263–275. [Google Scholar] [CrossRef]
- Zappulli, V.; Mazzariol, S.; Cavicchioli, L.; Petterino, C.; Bargelloni, L.; Castagnaro, M. Fatal necrotizing fasciitis and myositis in a captive common bottlenose dolphin (Tursiops truncatus) associated with Streptococcus agalactiae. J. Vet. Diagn. Investig. 2005, 17, 617–622. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.J.; Pasnik, D.J.; Klesius, P.H.; Al-Ablani, S. First report of Streptococcus agalactiae and Lactococcus garvieae from a wild bottlenose dolphin (Tursiops truncatus). J. Wildl. Dis. 2006, 42, 561–569. [Google Scholar] [CrossRef]
- Delannoy, C.M.; Crumlish, M.; Fontaine, M.C.; Pollock, J.; Foster, G.; Dagleish, M.P.; Turnbull, J.F.; Zadoks, R.N. Human Streptococcus agalactiae strains in aquatic mammals and fish. BMC Microbiol. 2013, 13, 41. [Google Scholar] [CrossRef] [Green Version]
- Fleming, M.; Bexton, S. Conjunctival flora of healthy and diseased eyes of grey seals (Halichoerus grypus): Implications for treatment. Vet. Rec. 2016, 179, 99. [Google Scholar] [CrossRef]
- Kissel, L.N.; Bankowski, M.J.; Koyamatsu, T.L.; Nagai, R.Y.; Seifried, S.E.; Crow, G.L. Aerobic microorganisms identified over a fourteen-month period from the upper respiratory tract of captive Hawaiian monk seals (Monachus schauinslandi). Aquat. Mamm. 2011, 37, 377. [Google Scholar] [CrossRef]
- Evans, J.J.; Klesius, P.H.; Gilbert, P.M.; Shoemaker, C.A.; Sarawi, M.A.A.; Landsberg, J.; Duremdez, R.; Marzouk, A.A.; Zenki, S.A. Characterization of β-haemolytic Group B Streptococcus agalactiae in cultured seabream, Sparus auratus L., and wild mullet, Liza klunzingeri (Day), in Kuwait. J. Fish Dis. 2002, 25, 505–513. [Google Scholar] [CrossRef]
- Mian, G.F.; Godoy, D.T.; Leal, C.A.G.; Yuhara, T.Y.; Costa, G.M.; Figueiredo, H.C.P. Aspects of the natural history and virulence of S. agalactiae infection in Nile tilapia. Vet. Microbiol. 2009, 136, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Duremdez, R.; Al-Marzouk, A.; Qasem, J.A.; Al-Harbi, A.; Gharabally, H. Isolation of Streptococcus agalactiae from cultured silver pomfret, Pampus argenteus (Euphrasen), in Kuwait. J. Fish Dis. 2004, 27, 307–310. [Google Scholar] [CrossRef]
- Garcia, J.C.; Klesius, P.H.; Evans, J.J.; Shoemaker, C.A. Non-infectivity of cattle Streptococcus agalactiae in Nile tilapia, Oreochromis niloticus and channel catfish, Ictalurus punctatus. Aquaculture 2008, 281, 151–154. [Google Scholar] [CrossRef]
- Pereira, U.P.; Mian, G.F.; Oliveira, I.C.M.; Benchetrit, L.C.; Costa, G.M.; Figueiredo, H.C.P. Genotyping of Streptococcus agalactiae strains isolated from fish, human and cattle and their virulence potential in Nile tilapia. Vet. Microbiol. 2010, 140, 186–192. [Google Scholar] [CrossRef]
- Kannika, K.; Pisuttharachai, D.; Srisapoome, P.; Wongtavatchai, J.; Kondo, H.; Hirono, I.; Unajak, S.; Areechon, N. Molecular serotyping, virulence gene profiling and pathogenicity of Streptococcus agalactiae isolated from tilapia farms in Thailand by multiplex PCR. J. Appl. Microbiol. 2017, 122, 1497–1507. [Google Scholar] [CrossRef] [PubMed]
- Delannoy, C.M.J.; Zadoks, R.N.; Crumlish, M.; Rodgers, D.; Lainson, F.A.; Ferguson, H.W.; Turnbull, J.; Fontaine, M.C. Genomic comparison of virulent and non-virulent Streptococcus agalactiae in fish. J. Fish Dis. 2016, 39, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Higginbottom, C.; Wheater, D.W.F. The incidence of Streptococcus bovis in cattle. J. Agric. Sci. 1954, 44, 434–442. [Google Scholar] [CrossRef]
- Greetham, H.L.; Giffard, C.; Hutson, R.A.; Collins, M.D.; Gibson, G.R. Bacteriology of the Labrador dog gut: A cultural and genotypic approach. J. Appl. Microbiol. 2002, 93, 640–646. [Google Scholar] [CrossRef] [Green Version]
- Moellering, R.C.; Watson, B.K.; Kunz, L.J. Endocarditis due to group D streptococci: Comparison of disease caused by Streptococcus bovis with that produced by the enterococci. Am. J. Med. 1974, 57, 239–250. [Google Scholar] [CrossRef]
- Fikar, C.R.; Levy, J. Streptococcus bovis meningitis in a neonate. Am. J. Dis. Child. 1979, 133, 1149–1150. [Google Scholar] [CrossRef] [PubMed]
- García-Porrúa, C.; González-Gay, M.A.; Monterroso, J.R.; Sánchez-Andrade, A.; González-Ramirez, A. Septic arthritis due to Streptococcus bovis as presenting sign of ‘silent’ colon carcinoma. Rheumatology 2000, 39, 338–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, H.W.; Roberts, R.B. Streptococcus bovis bacteremia and underlying gastrointestinal disease. Arch. Intern Med. 1978, 138, 1097–1099. [Google Scholar] [CrossRef] [PubMed]
- Vanrobaeys, M.; De Herdt, P.; Ducatelle, R.; Creten, W.; Haesebrouck, F. Extracellular proteins and virulence in Streptococcus bovis isolates from pigeons. Vet. Microbiol. 1997, 59, 59–66. [Google Scholar] [CrossRef]
- Vanrobaeys, M.; De Herdt, P.; Haesebrouck, F.; Ducatelle, R.; Devriese, L.A. Secreted antigens as virulence associated markers in Streptococcus bovis strains from pigeons. Vet. Microbiol. 1996, 53, 339–348. [Google Scholar] [CrossRef]
- Neves, H.C.; Pires, R. Recuperation of a Mediterranean monk seal pup, Monachus monachus, in Desertas Islands, Madeira archipelago: Conditions for its success. In Arquipelago: Life and Marine Sciences; The Monachus Guardian: Ponta Delgada, Portugal, 2001; Supplement 2 Part B; pp. 111–116. ISBN 0873-4704. [Google Scholar]
- Buck, J.D.; Wells, R.S.; Rhinehart, H.L.; Hansen, L.J. Aerobic microorganisms associated with free-ranging bottlenose dolphins in coastal Gulf of Mexico and Atlantic Ocean waters. J. Wildl. Dis. 2006, 42, 536–544. [Google Scholar] [CrossRef]
- Devriese, L.A.; Hommez, J.; Kilpper-Bälz, R.; Schleifer, K.-H. Streptococcus canis sp. nov.: A species of Group G streptococci from animals. Int. J. Syst. Evol. Microbiol. 1986, 36, 422–425. [Google Scholar] [CrossRef] [Green Version]
- Lam, M.M.; Clarridge, J.E.; Young, E.J.; Mizuki, S. The other Group G Streptococcus: Increased detection of Streptococcus canis ulcer infections in dog owners. J. Clin. Microbiol. 2007, 45, 2327–2329. [Google Scholar] [CrossRef] [Green Version]
- Chalmers, G.; McLean, J.; Hunter, D.B.; Brash, M.; Slavic, D.; Pearl, D.L.; Boerlin, P. Staphylococcus spp., Streptococcus canis, and Arcanobacterium phocae of healthy Canadian farmed mink and mink with pododermatitis. Can. J. Vet. Res. 2015, 79, 129–135. [Google Scholar]
- Hariharan, H.; Matthew, V.; Fountain, J.; Snell, A.; Doherty, D.; King, B.; Shemer, E.; Oliveira, S.; Sharma, R.N. Aerobic bacteria from mucous membranes, ear canals, and skin wounds of feral cats in Grenada, and the antimicrobial drug susceptibility of major isolates. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 129–134. [Google Scholar] [CrossRef]
- Timoney, J.F.; Velineni, S.; Ulrich, B.; Blanchard, P. Biotypes and ScM types of isolates of Streptococcus canis from diseased and healthy cats. Vet. Rec. 2017, 180, 358. [Google Scholar] [CrossRef] [PubMed]
- DeWinter, L.M.; Low, D.E.; Prescott, J.F. Virulence of Streptococcus canis from canine streptococcal toxic shock syndrome and necrotizing fasciitis. Vet. Microbiol. 1999, 70, 95–110. [Google Scholar] [CrossRef]
- Fulde, M.; Rohde, M.; Polok, A.; Preissner, K.T.; Chhatwal, G.S.; Bergmann, S. Cooperative plasminogen recruitment to the surface of Streptococcus canis via M protein and enolase enhances bacterial survival. mBio 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichhorn, I.; Linden, M.; van der Jarek, M.; Fulde, M. Draft genome sequence of zoonotic Streptococcus canis isolate G361. Genome Announc. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Liu, Y.; Xu, J.; Li, B. Characterization of a new protective antigen of Streptococcus canis. Vet. Res. Commun. 2010, 34, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, S.; Eichhorn, I.; Kohler, T.P.; Hammerschmidt, S.; Goldmann, O.; Rohde, M.; Fulde, M. SCM, the M protein of Streptococcus canis binds immunoglobulin G. Front. Cell Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Van Bonn, W.G.; Ridgway, S.H.; Williams, B.H. Chronic refractory emesis associated with a colonic lesion in a california sea lion (Zalophus californianus). J. Zoo Wildl. Med. 1995, 26, 286–292. [Google Scholar]
- Williams, D.L.; MacGregor, S.; Sainsbury, A.W. Evaluation of bacteria isolated from infected eyes of captive, non-domestic animals. Vet. Rec. 2000, 146, 515–518. [Google Scholar] [CrossRef]
- Seguel, M.; Gutiérrez, J.; Hernández, C.; Montalva, F.; Verdugo, C. Respiratory mites (Orthohalarachne diminuata) and β-hemolytic streptococci-associated bronchopneumonia outbreak in South American fur seal pups (Arctocephalus australis). J. Wildl. Dis. 2018, 54, 380–385. [Google Scholar] [CrossRef]
- Henton, M.M.; Zapke, O.; Basson, P.A. Streptococcus phocae infections associated with starvation in Cape fur seals: Case report. J. S. Afr. Vet. Assoc. 1999, 70, 98–99. [Google Scholar] [CrossRef] [Green Version]
- Jepson, P.D.; Baker, J.R.; Kuiken, T.; Simpson, V.R.; Kennedy, S.; Bennett, P.M. Pulmonary pathology of harbour porpoises (Phocoena phocoena) stranded in England. Vet. Rec. 2000, 146, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Leahy, C.L. Causes and Patterns of Harbor Seal (Phoca vitulina) Pup Mortality at Smith Island, Washington, 2004–2009. Ph.D. Thesis, Evergreen State College, Olympia, WA, USA, 2010. [Google Scholar]
- Vandamme, P.; Pot, B.; Falsen, E.; Kersters, K.; Devriese, L.A. Taxonomic study of Lancefield streptococcal Groups C, G, and L (Streptococcus dysgalactiae) and proposal of S. dysgalactiae subsp. equisimilis subsp. nov. Int. J. Syst. Evol. Microbiol. 1996, 46, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.M.; Wilson, M.E.; Brandt, C.M.; Spellerberg, B. Human infections due to Streptococcus dysgalactiae subspecies equisimilis. Clin. Infect. Dis. 2009, 49, 766–772. [Google Scholar] [CrossRef] [Green Version]
- Jensen, A.; Kilian, M. Delineation of Streptococcus dysgalactiae, its subspecies, and its clinical and phylogenetic relationship to Streptococcus pyogenes. J. Clin. Microbiol. 2012, 50, 113–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, J.R.; Doidge, D.W. Pathology of the antarctic fur seal (Arctocephalus gazella) in South Georgia. Br. Vet. J. 1984, 140, 210–219. [Google Scholar] [CrossRef]
- Vossen, A.; Abdulmawjood, A.; Lämmler, C.; Weiß, R.; Siebert, U. Identification and molecular characterization of beta-hemolytic streptococci isolated from harbor seals (Phoca vitulina) and grey seals (Halichoerus grypus) of the German North and Baltic Seas. J. Clin. Microbiol. 2004, 42, 469–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, M.V.; Ehlers, L.P.; Vargas, T.P.; Lopes, B.C.; Taunde, P.A.; de Cecco, B.S.; Henker, L.C.; Vielmo, A.; Lorenzett, M.P.; Riboldi, C.I.; et al. Omphalitis, urachocystitis and septicemia by Streptococcus dysgalactiae in a southern right whale calf Eubalaena australis, Brazil. Dis. Aquat. Organ. 2018, 131, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Boyle, A.G.; Timoney, J.F.; Newton, J.R.; Hines, M.T.; Waller, A.S.; Buchanan, B.R. Streptococcus equi infections in horses: Guidelines for treatment, control, and prevention of strangles—Revised consensus statement. J. Vet. Intern Med. 2018, 32, 633–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalker, V.J.; Brooks, H.W.; Brownlie, J. The association of Streptococcus equi subsp. zooepidemicus with canine infectious respiratory disease. Vet. Microbiol. 2003, 95, 149–156. [Google Scholar] [CrossRef]
- Breiman, R.F.; Silverblatt, F.J. Systemic Streptococcus equi infection in a horse handler—A case of human strangles. West. J. Med. 1986, 145, 385–386. [Google Scholar]
- Eyre, D.W.S.; Kenkre, J.; Bowler, I.C.J.W.; McBride, S.J. Streptococcus equi subspecies zooepidemicus meningitis—A case report and review of the literature. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 1459–1463. [Google Scholar] [CrossRef] [PubMed]
- Akineden, Ö.; Hassan, A.A.; Alber, J.; El-Sayed, A.; Estoepangestie, A.T.S.; Lämmler, C.; Weiss, R.; Siebert, U. Phenotypic and genotypic properties of Streptococcus equi subsp. zooepidemicus isolated from harbor seals (Phoca vitulina) from the German North Sea during the phocine distemper outbreak in 2002. Vet. Microbiol. 2005, 110, 147–152. [Google Scholar] [CrossRef]
- Akineden, Ö; Alber, J.; Lämmler, C.; Weiss, R.; Siebert, U.; Foster, G.; Tougaard, S.; Brasseur, S.M.J.M.; Reijnders, P.J.H. Relatedness of Streptococcus equi subsp. zooepidemicus strains isolated from harbour seals (Phoca vitulina) and grey seals (Halichoerus grypus) of various origins of the North Sea during 1988–2005. Vet. Microbiol. 2007, 121, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Venn-Watson, S.; Daniels, R.; Smith, C. Thirty year retrospective evaluation of pneumonia in a bottlenose dolphin Tursiops truncatus population. Dis. Aquat. Organ. 2012, 99, 237–242. [Google Scholar] [CrossRef] [Green Version]
- Nelson, T.M.; Wallen, M.M.; Bunce, M.; Oskam, C.L.; Lima, N.; Clayton, L.; Mann, J. Detecting respiratory bacterial communities of wild dolphins: Implications for animal health. Mar. Ecol. Prog. Ser. 2019, 622, 203–217. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Kim, J.-Y.; Jung, S.C.; Lee, H.-S.; Her, M.; Chae, C. First isolation of Streptococcus halichoeri and Streptococcus phocae from a steller sea lion (Eumetopias jubatus) in South Korea. J. Wildl. Dis. 2015, 52, 183–185. [Google Scholar] [CrossRef] [PubMed]
- Foo, R.M.; Chan, D. A Fishy Tale: A man with empyema caused by Streptococcus halichoeri. J. Clin. Microbiol. 2014, 52, 681–682. [Google Scholar] [CrossRef] [Green Version]
- Giudice, P.D.; Plainvert, C.; Hubiche, T.; Tazi, A.; Fribourg, A.; Poyart, C. Infectious cellulitis caused by Streptococcus halichoeri. Acta Derm. Venereol. 2018, 98, 378–379. [Google Scholar] [CrossRef] [Green Version]
- Shewmaker, P.L.; Whitney, A.M.; Humrighouse, B.W. Phenotypic, genotypic, and antimicrobial characteristics of Streptococcus halichoeri isolates from humans, proposal to rename Streptococcus halichoeri as Streptococcus halichoeri subsp. halichoeri, and description of Streptococcus halichoeri subsp. hominis subsp. nov., a bacterium associated with human clinical infections. J. Clin. Microbiol. 2016, 54, 739–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aaltonen, K.; Kant, R.; Eklund, M.; Raunio-Saarnisto, M.; Paulin, L.; Vapalahti, O.; Grönthal, T.; Rantala, M.; Sironen, T. Streptococcus halichoeri: Comparative genomics of an emerging pathogen. Int. J. Genomics 2020. [Google Scholar] [CrossRef] [Green Version]
- Eklund, M.; Aaltonen, K.; Sironen, T.; Raunio-Saarnisto, M.; Grönthal, T.; Nordgren, H.; Pitkälä, A.; Vapalahti, O.; Rantala, M. Comparison of Streptococcus halichoeri isolates from canine and fur animal infections: Biochemical patterns, molecular characteristics and genetic relatedness. Acta Vet. Scand. 2020, 62, 26. [Google Scholar] [CrossRef] [PubMed]
- Pier, G.B.; Madin, S.H. Streptococcus iniae sp. nov., a beta-hemolytic Streptococcus isolated from an Amazon freshwater dolphin, Inia geoffrensis. Int. J. Syst. Evol. Microbiol. 1976, 26, 545–553. [Google Scholar] [CrossRef]
- Pier, G.B.; Madin, S.H.; Al-Nakeeb, S. Isolation and characterization of a second isolate of Streptococcus iniae. Int. J. Syst. Evol. Microbiol. 1978, 28, 311–314. [Google Scholar] [CrossRef] [Green Version]
- Bonar, C.J.; Wagner, R.A. A Third report of “golf ball disease” in an amazon river dolphin (Inia geoffrensis) associated with Streptococcus iniae. J. Zoo Wildl. Med. 2003, 34, 296–301. [Google Scholar] [CrossRef]
- Song, Z.; Yue, R.; Sun, Y.; Liu, C.; Khan, S.H.; Li, C.; Zhao, Y.; Zhou, X.; Yang, L.; Zhao, D. Fatal bacterial septicemia in a bottlenose dolphin Tursiops truncatus caused by Streptococcus iniae. Dis. Aquat. Organ. 2017, 122, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Rahmatullah, M.; Ariff, M.; Kahieshesfandiari, M.; Daud, H.M.; Zamri-Saad, M.; Sabri, M.Y.; Amal, M.N.A.; Ina-Salwany, M.Y. Isolation and pathogenicity of Streptococcus iniae in cultured red hybrid tilapia in Malaysia. J. Aquat. Anim. Health 2017, 29, 208–213. [Google Scholar] [CrossRef]
- Locke, J.B.; Colvin, K.M.; Datta, A.K.; Patel, S.K.; Naidu, N.N.; Neely, M.N.; Nizet, V.; Buchanan, J.T. Streptococcus iniae capsule impairs phagocytic clearance and contributes to virulence in fish. J. Bacteriol. 2007, 189, 1279–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locke, J.B.; Colvin, K.M.; Varki, N.; Vicknair, M.R.; Nizet, V.; Buchanan, J.T. Streptococcus iniae β-hemolysin streptolysin S is a virulence factor in fish infection. Dis. Aquat. Organ. 2007, 76, 17–26. [Google Scholar] [CrossRef]
- Soh, K.Y.; Loh, J.M.S.; Hall, C.; Proft, T. Functional analysis of two novel Streptococcus iniae virulence factors using a zebrafish infection model. Microorganisms 2020, 8, 1361. [Google Scholar] [CrossRef]
- Milani, C.J.E.; Aziz, R.K.; Locke, J.B.; Dahesh, S.; Nizet, V.; Buchanan, J.T. The novel polysaccharide deacetylase homologue Pdi contributes to virulence of the aquatic pathogen Streptococcus iniae. Microbiology 2010, 156, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Locke, J.B.; Aziz, R.K.; Vicknair, M.R.; Nizet, V.; Buchanan, J.T. Streptococcus iniae M-like protein contributes to virulence in fish and is a target for live attenuated vaccine development. PLoS ONE 2008, 3, e2824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Wang, Y.; Wang, K. Comparative genome analysis of Streptococcus iniae DX09 reveals new insights into niche adaptation and competitive host colonisation ability. Oncotarget 2017, 5. [Google Scholar] [CrossRef]
- Silayeva, O.; Engelstädter, J.; Barnes, A.C. Evolutionary epidemiology of Streptococcus iniae: Linking mutation rate dynamics with adaptation to novel immunological landscapes. bioRxiv 2020, 355412. [Google Scholar] [CrossRef] [PubMed]
- Aas, J.A.; Paster, B.J.; Stokes, L.N.; Olsen, I.; Dewhirst, F.E. Defining the normal bacterial flora of the oral cavity. J. Clin. Microbiol. 2005, 43, 5721–5732. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.J.; Anderson, J.M.; King, W.F.; Houte, J. van, Taubman, M.A. Oral streptococcal colonization of infants. Oral Microbiol. Immunol. 1993, 8, 1–4. [Google Scholar] [CrossRef]
- Matsui, N.; Ito, M.; Kuramae, H.; Inukai, T.; Sakai, A.; Okugawa, M. Infective endocarditis caused by multidrug-resistant Streptococcus mitis in a combined immunocompromised patient: An autopsy case report. J. Infect. Chemother. 2013, 19, 321–325. [Google Scholar] [CrossRef]
- Husain, E.; Whitehead, S.; Castell, A.; Thomas, E.E.; Speert, D.P. Viridans streptococci bacteremia in children with malignancy: Relevance of species identification and penicillin susceptibility. Pediatr. Infect. Dis. J. 2005, 24, 563–566. [Google Scholar] [CrossRef]
- Marron, A.; Carratalà, J.; González-Barca, E.; Fernández-Sevilla, A.; Alcaide, F.; Gudiol, F. Serious complications of bacteremia caused by viridans streptococci in neutropenic patients with cancer. Clin. Infect. Dis. 2000, 31, 1126–1130. [Google Scholar] [CrossRef]
- Denapaite, D.; Brückner, R.; Nuhn, M.; Reichmann, P.; Henrich, B.; Maurer, P.; Schähle, Y.; Selbmann, P.; Zimmermann, W.; Wambutt, R.; et al. The genome of Streptococcus mitis B6—What is a commensal? PLoS ONE 2010, 5, e9426. [Google Scholar] [CrossRef] [Green Version]
- Buck, J.D.; Shepard, L.L.; Bubucis, P.M.; Spotte, S.; McClave, K.; Cook, R.A. Microbiological characteristics of white whale (Delphinapterus leucas) from capture through extended captivity. Can. J. Fish Aquat. Sci. 1989, 46, 1914–1921. [Google Scholar] [CrossRef]
- Mitchell, J. Streptococcus mitis: Walking the line between commensalism and pathogenesis. Mol. Oral Microbiol. 2011, 26, 89–98. [Google Scholar] [CrossRef]
- Romalde, J.L.; Ravelo, C.; Valdés, I.; Magariños, B.; de la Fuente, E.; Martín, C.S.; Avendaño-Herrera, R.; Toranzo, A.E. Streptococcus phocae, an emerging pathogen for salmonid culture. Vet. Microbiol. 2008, 130, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Gibello, A.; Mata, A.I.; Blanco, M.M.; Casamayor, A.; Domínguez, L.; Fernández-Garayzabal, J.F. First identification of Streptococcus phocae isolated from Atlantic salmon (Salmo salar). J. Clin. Microbiol. 2005, 43, 526–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, D.; Jang, S.; Miller, M.; Conrad, P.A. Characterization of beta-hemolytic streptococci isolated from southern sea otters (Enhydra lutris nereis) stranded along the California coast. Vet. Microbiol. 2009, 136, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Satish, R.K.; Arul, V. Purification and characterization of phocaecin PI80: An anti-listerial bacteriocin produced by Streptococcus phocae PI80 Isolated from the gut of Peneaus indicus (Indian white shrimp). J. Microbiol. Biotechnol. 2009, 19, 1393–1400. [Google Scholar]
- Avendaño-Herrera, R.; Balboa, S.; Castro, N.; González-Contreras, A.; Magariños, B.; Fernández, J.; Toranzo, A.E.; Romalde, J.L. Comparative polyphasic characterization of Streptococcus phocae strains with different host origin and description of the subspecies Streptococcus phocae subsp. salmonis subsp. nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 1775–1781. [Google Scholar] [CrossRef]
- Ludes-Wehrmeister, E.; Wohlsein, P.; Prenger-Berninghoff, E.; Ewers, C.; Woelfing, B.; Lehnert, K.; Siebert, U. Intestinal displacements in older harbour and grey seals. Dis. Aquat. Organ. 2020, 138, 215–225. [Google Scholar] [CrossRef]
- Hueffer, K.; Lieske, C.L.; McGilvary, L.M.; Hare, R.F.; Miller, D.L.; O’Hara, T.M. Streptococcus phocae isolated from a spotted seal (Phoca largha) with pyometra in Alaska. J. Zoo Wildl. Med. 2011, 42, 108–112. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, C.L.; Jensen, E.D.; Johnson, S.P.; Lutmerding, B.A.; Smith, C.R.; Meegan, J.M.; Emory-Gomez, F.M.; Venn-Watson, S. Strangles-like clinical presentation of Streptococcus phocae in three bottlenose dolphins (Tursiops truncatus). In Proceedings of the 42nd Annual Conference of the International Association for Aquatic Animal Medicine, Las Vegas, NV, USA, 7–11 May 2011; Available online: https://www.vin.com/doc/?id=6698522 (accessed on 11 January 2021).
- Fenton, H.; Daoust, P.-Y.; Forzán, M.J.; Vanderstichel, R.V.; Ford, J.K.B.; Spaven, L.; Lair, S.; Raverty, S. Causes of mortality of harbor porpoises Phocoena phocoena along the Atlantic and Pacific coasts of Canada. Dis. Aquat. Organ. 2017, 122, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Taurisano, N.D.; Butler, B.P.; Stone, D.; Hariharan, H.; Fields, P.J.; Ferguson, H.W.; Haulena, M.; Cotrell, P.; Nielsen, O.; Raverty, S. Streptococcus phocae in marine mammals of Northeastern Pacific and Arctic Canada: A retrospective analysis of 85 postmortem investigations. J. Wildl. Dis. 2017, 54, 101–111. [Google Scholar] [CrossRef]
- Goertz, C.E.C.; Reichmuth, C.; Thometz, N.M.; Ziel, H.; Boveng, P. Comparative health assessments of Alaskan ice seals. Front. Vet. Sci. 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Ashley, E.A.; Olson, J.K.; Adler, T.E.; Raverty, S.; Anderson, E.M.; Jeffries, S.; Gaydos, J.K. Causes of mortality in a harbor seal (Phoca vitulina) population at equilibrium. Front. Mar. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Martin, M.C.-S.; González-Contreras, A.; Avendaño-Herrera, R. Infectivity study of Streptococcus phocae to seven fish and mammalian cell lines by confocal microscopy. J. Fish Dis. 2018, 35, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Salazar, S.; Oliver, C.; Yáñez, A.J.; Avendaño-Herrera, R. Comparative analysis of innate immune responses to Streptococcus phocae strains in Atlantic salmon (Salmo salar) and rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2016, 51, 97–103. [Google Scholar] [CrossRef]
- Avendaño-Herrera, R.; Poblete-Morales, M. Genome sequence of Streptococcus phocae subsp. phocae Strain ATCC 51973T isolated from a harbor seal (Phoca vitulina). Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, M.J.; IJsseldijk, L.L.; Rubio-García, A.; Gröne, A.; Duim, B.; Rossen, J.; Zomer, A.L.; Wagenaar, J.A. After the bite: Bacterial transmission from grey seals (Halichoerus grypus) to harbour porpoises (Phocoena phocoena). R. Soc. Open Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Coykendall, A.L. Classification and identification of the viridans streptococci. Clin. Microbiol. Rev. 1989, 2, 315–328. [Google Scholar] [CrossRef]
- Facklam, R.R. Physiological differentiation of viridans streptococci. J. Clin. Microbiol. 1977, 5, 184–201. [Google Scholar] [PubMed]
- Beighton, D.; Hardie, J.M.; Whiley, R.A. A scheme for the identification of viridans streptococci. J. Med. Microbiol. 1991, 35, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Douglas, C.W.I.; Heath, J.; Hampton, K.K.; Preston, F.E. Identity of viridans streptococci isolated from cases of infective endocarditis. J. Med. Microbiol. 1993, 39, 179–182. [Google Scholar] [CrossRef] [Green Version]
- Salavert, M.; Gomez, L.; Rodriguez-Carballeira, M.; Xercavins, M.; Freixas, N.; Garau, J. Seven-year review of bacteremia caused by Streptococcus milleri and other viridans streptococci. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, S.; Yatera, K.; Kawanami, T.; Yamasaki, K.; Naito, K.; Akata, K.; Shimabukuro, I.; Ishimoto, H.; Yoshii, C.; Mukae, H. The clinical features of respiratory infections caused by the Streptococcus anginosus group. BMC Pulm. Med. 2015, 15, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornton, S.M.; Nolan, S.; Gulland, F.M.D. Bacterial isolates from california sea lions (Zalophus californianus), harbor seals (Phoca vitulina), and northern elephant seals (Mirounga angustirostris) admitted to a rehabilitation center along the Central California Coast, 1994–1995. J. Zoo Wildl. Med. 1998, 29, 171–176. [Google Scholar] [PubMed]
- Johnson, S.P.; Jang, S.; Gulland, F.M.D.; Miller, M.A.; Casper, D.R.; Lawrence, J.; Herrera, J. Characterization and clinical manifestations of Arcanobacterium phocae infections in marine mammals stranded along the Central California Coast. J. Wildl. Dis. 2003, 39, 136–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burek-Huntington, K.A.; Dushane, J.L.; Goertz, C.E.C.; Measures, L.N.; Romero, C.H.; Raverty, S.A. Morbidity and mortality in stranded Cook Inlet beluga whales Delphinapterus leucas. Dis. Aquat. Organ. 2015, 114, 45–60. [Google Scholar] [CrossRef]
- Morris, P.J.; Johnson, W.R.; Pisani, J.; Bossart, G.D.; Adams, J.; Reif, J.S.; Fair, P.A. Isolation of culturable microorganisms from free-ranging bottlenose dolphins (Tursiops truncatus) from the southeastern United States. Vet. Microbiol. 2011, 148, 440–447. [Google Scholar] [CrossRef]
- Li, C.; Tan, X.; Bai, J.; Xu, Q.; Liu, S.; Guo, W.; Yu, C.; Fan, G.; Lu, Y.; Zhang, H.; et al. A survey of the sperm whale (Physeter catodon) commensal microbiome. PeerJ 2019, 7, e7257. [Google Scholar] [CrossRef] [Green Version]
- Rocha Goselin, A. Caracterización de la Carga Microbiana y Parasitaria de tres Especies de Misticetos en las ostas de la Península de Baja California, México. 2009. Available online: http://www.repositoriodigital.ipn.mx//handle/123456789/13932 (accessed on 9 July 2020).
- Benga, L.; Fulde, M.; Neis, C.; Goethe, R.; Valentin-Weigand, P. Polysaccharide capsule and suilysin contribute to extracellular survival of Streptococcus suis co-cultivated with primary porcine phagocytes. Vet. Microbiol. 2008, 132, 211–219. [Google Scholar] [CrossRef]
- Edwards, M.S.; Nicholson-Weller, A.; Baker, C.J.; Kasper, D.L. The role of specific antibody in alternative complement pathway-mediated opsonophagocytosis of type III, group B Streptococcus. J. Exp. Med. 1980, 151, 1275–1287. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.O.; Romero-Steiner, S.; Sørensen, U.B.; Blom, J.; Carvalho, M.; Barnard, S.; Carlone, G.; Weiser, J.N. Relationship between cell surface carbohydrates and intrastrain variation on opsonophagocytosis of Streptococcus pneumoniae. Infect. Immun. 1999, 67, 2327–2333. [Google Scholar] [CrossRef] [Green Version]
- Wessels, M.R.; Moses, A.E.; Goldberg, J.B.; DiCesare, T.J. Hyaluronic acid capsule is a virulence factor for mucoid group A streptococci. Proc. Natl. Acad. Sci. USA 1991, 88, 8317–8321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischetti, V.A. Streptococcal M protein: Molecular design and biological behavior. Clin. Microbiol. Rev. 1989, 2, 285–314. [Google Scholar] [CrossRef]
- Brouwer, S.; Barnett, T.C.; Rivera-Hernandez, T.; Rohde, M.; Walker, M.J. Streptococcus pyogenes adhesion and colonization. FEBS Lett. 2016, 590, 3739–3757. [Google Scholar] [CrossRef] [PubMed]
- Nobbs, A.H.; Lamont, R.J.; Jenkinson, H.F. Streptococcus adherence and colonization. Microbiol. Mol. Biol. Rev. 2009, 73, 407–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanier, G.; Segura, M.; Friedl, P.; Lacouture, S.; Gottschalk, M. Invasion of porcine brain microvascular endothelial cells by Streptococcus suis Serotype 2. Infect. Immun. 2004, 72, 1441–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greco, R.; De Martino, L.; Donnarumma, G.; Conte, M.P.; Seganti, L.; Valenti, P. Invasion of cultured human cells by Streptococcus pyogenes. Res. Microbiol. 1995, 146, 551–560. [Google Scholar] [CrossRef]
- Bhakdi, S.; Tranum-Jensen, J.; Sziegoleit, A. Mechanism of membrane damage by streptolysin-O. Infect. Immun. 1985, 47, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Alouf, J.E. Streptococcal toxins (streptolysin O, streptolysin S, erythrogenic toxin). Pharmacol. Ther. 1980, 11, 661–717. [Google Scholar] [CrossRef]
- Khil, J.; Im, M.; Heath, A.; Ringdahl, U.; Mundada, L.; Cary Engleberg, N.; Fay, W.P. Plasminogen enhances virulence of Group A streptococci by streptokinase-dependent and streptokinase-independent mechanisms. J. Infect. Dis. 2003, 188, 497–505. [Google Scholar] [CrossRef] [Green Version]
- Blair, D.E.; Schüttelkopf, A.W.; MacRae, J.I.; van Aalten, D.M.F. Structure and metal-dependent mechanism of peptidoglycan deacetylase, a streptococcal virulence factor. Proc. Natl. Acad. Sci. USA 2005, 102, 15429–15434. [Google Scholar] [CrossRef] [Green Version]
- Von Graevenitz, A. The role of opportunistic bacteria in human disease. Annu. Rev. Microbiol. 1977, 31, 447–471. [Google Scholar] [CrossRef] [PubMed]
- Lewthwaite, P.; Parsons, H.K.; Bates, C.J.; McKendrick, M.W.; Dockrell, D.H. Group G streptococcal bacteraemia: An opportunistic infection associated with immune senescence. Scand. J. Infect. Dis. 2002, 34, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Wilde, S.; Olivares, K.L.; Nizet, V.; Hoffman, H.M.; Radhakrishna, S.; LaRock, C.N. Opportunistic invasive infection by Group A Streptococcus during anti–interleukin-6 immunotherapy. J. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Louw, A.; Tikly, M. Purulent pericarditis due to co-infection with Streptococcus pneumoniae and Mycobacterium tuberculosis in a patient with features of advanced HIV infection. BMC Infect. Dis. 2007, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Blyth, C.C.; Webb, S.A.R.; Kok, J.; Dwyer, D.E.; van Hal, S.J.; Foo, H.; Ginn, A.N.; Kesson, A.M.; Seppelt, I.; Iredell, J.R. The impact of bacterial and viral co-infection in severe influenza. Influenza Other Respir. Viruses 2013, 7, 168–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmer, A.M.; Timmer, J.C.; Pence, M.A.; Hsu, L.-C.; Ghochani, M.; Frey, T.G.; Karin, M.; Salvesen, G.S.; Nizet, V. Streptolysin O promotes Group A Streptococcus immune evasion by accelerated macrophage apoptosis. J. Biol. Chem. 2009, 284, 862–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinkla, K.; Rohde, M.; Jansen, W.T.M.; Carapetis, J.R.; Chhatwal, G.S.; Talay, S.R. Streptococcus pyogenes recruits collagen via surface-bound fibronectin: A novel colonization and immune evasion mechanism. Mol. Microbiol. 2003, 47, 861–869. [Google Scholar] [CrossRef]
- Avendaño-Herrera, R.; Suarez, R.; Lazo, E.; Bravo, D.; Llegues, K.O.; Romalde, J.L.; Godoy, M.G. Genome sequence of Streptococcus phocae subsp. salmonis Strain C-4T, isolated from Atlantic salmon (Salmo salar). Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Wierzbicki, I.H.; Campeau, A.; Dehaini, D.; Holay, M.; Wei, X.; Greene, T.; Ying, M.; Sands, J.S.; Lamsa, A.; Zuniga, E.; et al. Group A streptococcal S protein utilizes red blood cells as immune camouflage and is a critical determinant for immune evasion. Cell Rep. 2019, 29, 2979–2989.e15. [Google Scholar] [CrossRef] [Green Version]
- Rungelrath, V.; Wohlsein, J.C.; Siebert, U.; Stott, J.; Prenger-Berninghoff, E.; von Pawel-Rammingen, U.; Valentin-Weigand, P.; Baums, C.G.; Seele, J. Identification of a novel host-specific IgG protease in Streptococcus phocae subsp. phocae. Vet. Microbiol. 2017, 201, 42–48. [Google Scholar] [CrossRef]
- Cavagnolo, R.Z.; Vedros, N.A. Identification and characterization of three immunoglobulin classes in the northern fur seal Callorhinus ursinus. Dev. Comp. Immunol. 1978, 2, 689–697. [Google Scholar] [CrossRef]
- Carter, S.D.; Hughes, D.E.; Baker, J.R. Characterization and measurement of immunoglobulins in the grey seal (Halichoerus grypus). J. Comp. Pathol. 1990, 102, 13–23. [Google Scholar] [CrossRef]
- Nash, D.R.; Mach, J.P. Immunoglobulin classes in aquatic mammals: Characterization by serologic cross-reactivity, molecular size and binding of human free secretory component. J. Immunol. 1971, 107, 1424–1430. [Google Scholar] [PubMed]
- Saito, H.; Poon, M.-C.; Goldsmith, G.H.; Ratnoff, O.D.; Árnason, Ú. Studies on the blood clotting and fibrinolytic system in the plasma from a sei (baleen) whale. Proc. Soc. Exp. Biol. Med. 1976, 152, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Robinson, A.J.; Kropatkin, M.; Aggeler, P.M. Hageman Factor (Factor XII) deficiency in marine mammals. Science 1969, 166, 1420–1422. [Google Scholar] [CrossRef]
- Huang, T.T.; Malke, H.; Ferretti, J.J. Heterogeneity of the streptokinase gene in group A streptococci. Infect. Immun. 1989, 57, 502–506. [Google Scholar] [CrossRef] [Green Version]
- Feltz, E.T.; Fay, F.H. Thermal requirements in vitro of epidermal cells from seals. Cryobiology 1966, 3, 261–264. [Google Scholar] [CrossRef]
- Katsumata, E.; Furuta, C.; Katsumata, H.; Watanabe, G.; Taya, K. Basal body temperature method for detecting ovarian cycle in the captive beluga (Delphinapterus leucas). J. Reprod. Dev. 2005. [Google Scholar] [CrossRef] [Green Version]
- Katsumata, E.; Jaroenporn, S.; Katsumata, H.; Konno, S.; Maeda, Y.; Watanabe, G.; Taya, K. Body temperature and circulating progesterone levels before and after parturition in killer whales (Orcinus orca). J. Reprod. Dev. 2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terasawa, F.; Yokoyama, Y.; Kitamura, M. Rectal temperatures before and after parturition in bottlenose dolphins. Zoo Biol. 1999, 18, 153–156. [Google Scholar] [CrossRef]
- Bartholomew, G.A.; Wilke, F. Body temperature in the northern fur seal, Callorhinus ursinus. J. Mammal 1956, 37, 327–337. [Google Scholar] [CrossRef]
- Oppenheimer, C.H.; ZoBell, C.E. The growth and viability of sixty-three species of marine bacteria as influenced by hydrostatic pressure. J. Mar. Res. 1952, 11, 10–18. [Google Scholar]
- ZoBell, C.E.; Johnson, F.H. The influence of hydrostatic pressure on the growth and viability of terrestrial and marine bacteria. J. Bacteriol. 1949, 57, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Hoppes, W.L.; Lerner, P.I. Nonenterococcal Group-D streptococcal endocarditis caused by Streptococcus bovis. Ann. Intern Med. 1974, 81, 588–593. [Google Scholar] [CrossRef]
- Facklam, R.R.; Padula, J.F.; Thacker, L.G.; Wortham, E.C.; Sconyers, B.J. Presumptive identification of Group, A, B, and D streptococci. Appl. Microbiol. 1974, 27, 107–113. [Google Scholar] [CrossRef]
- Ayhan, K.; Durlu-Özkaya, F.; Tunail, N. Commercially important characteristics of Turkish origin domestic strains of Streptococcus thermophilus and Lactobacillus delbrueckii ssp. bulgaricus. Int. J. Dairy Technol. 2005, 58, 150–157. [Google Scholar] [CrossRef]
- Seeley, H.W. The physiology and nutrition of Streptococcus uberis. J. Bacteriol. 1951, 62, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Currás, M.; Magariños, B.; Toranzo, A.E.; Romalde, J.L. Dormancy as a survival strategy of the fish pathogen Streptococcus parauberis in the marine environment. Dis. Aquat. Organ. 2002, 52, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, D.B.; Fahlman, A.; Gardner, M.; Kleinhenz, D.; Piscitelli, M.; Raverty, S.; Haulena, M.; Zimba, P.V. Phosphatidylcholine composition of pulmonary surfactant from terrestrial and marine diving mammals. Respir. Physiol. Neurobiol. 2015, 211, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosinski-Chupin, I.; Sauvage, E.; Mairey, B.; Mangenot, S.; Ma, L.; Da Cunha, V.; Rusniok, C.; Bouchier, C.; Barbe, V.; Glaser, P. Reductive evolution in Streptococcus agalactiae and the emergence of a host adapted lineage. BMC Genomics 2013, 14, 252. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Lefébure, T.; Hubisz, M.J.; Pavinski Bitar, P.; Lang, P.; Siepel, A.; Stanhope, M.J. Comparative genomic analysis of the Streptococcus dysgalactiae species group: Gene content, molecular adaptation, and promoter evolution. Genome Biol. Evol. 2011, 3, 168–185. [Google Scholar] [CrossRef] [Green Version]
- Richards, V.P.; Zadoks, R.N.; Pavinski Bitar, P.D.; Lefébure, T.; Lang, P.; Werner, B.; Tikofsky, L.; Moroni, P.; Stanhope, M.J. Genome characterization and population genetic structure of the zoonotic pathogen, Streptococcus canis. BMC Microbiol. 2012, 12, 293. [Google Scholar] [CrossRef] [Green Version]
- Burek, K.A.; Gulland, F.M.D.; O’Hara, T.M. Effects of climate change on arctic marine mammal health. Ecol. Appl. 2008, 18, S126–S134. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S. Morbillivirus infections in aquatic mammals. J. Comp. Pathol. 1998, 119, 201–225. [Google Scholar] [CrossRef]
- Zlotkin, A.; Hershko, H.; Eldar, A. Possible transmission of Streptococcus iniae from wild fish to cultured marine fish. Appl. Environ. Microbiol. 1998, 64, 4065–4067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, M.; Delamare-Deboutteville, J.; Bowater, R.O.; Walker, M.J.; Beatson, S.; Zakour, N.L.B.; Barnes, A.C. Microevolution of Streptococcus agalactiae ST-261 from Australia indicates dissemination via imported tilapia and ongoing adaptation to marine hosts or environment. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmeyer, R. Diversity of bacteria in ships ballast water as revealed by next generation DNA sequencing. Mar. Pollut. Bull. 2016, 107, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Altug, G.; Gurun, S.; Cardak, M.; Ciftci, P.S.; Kalkan, S. The occurrence of pathogenic bacteria in some ships’ ballast water incoming from various marine regions to the Sea of Marmara, Turkey. Mar. Environ. Res. 2012, 81, 35–42. [Google Scholar] [CrossRef]
- Gerba, C.P. Assessment of enteric pathogen shedding by bathers during recreational activity and its impact on water quality. Quant Microbiol. 2000, 2, 55–68. [Google Scholar] [CrossRef]
- Elmir, S.M.; Wright, M.E.; Abdelzaher, A.; Solo-Gabriele, H.M.; Fleming, L.E.; Miller, G.; Rybolowik, M.; Peter Shih, M.-T.; Pillai, S.P.; Cooper, J.A.; et al. Quantitative evaluation of bacteria released by bathers in a marine water. Water Res. 2007, 41, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Plano, L.R.; Garza, A.C.; Shibata, T.; Elmir, S.M.; Kish, J.; Sinigalliano, C.D.; Gidley, M.L.; Miller, G.; Withum, K.; Fleming, L.E.; et al. Shedding of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus from adult and pediatric bathers in marine waters. BMC Microbiol. 2011, 11, 5. [Google Scholar] [CrossRef] [Green Version]
- Kalkan, S.; Altuğ, G. The composition of cultivable bacteria, bacterial pollution, and environmental variables of the coastal areas: An example from the Southeastern Black Sea, Turkey. Environ. Monit. Assess. 2020, 192, 1–23. [Google Scholar] [CrossRef]
- Nielsen, M.C.; Jiang, S.C. Alterations of the human skin microbiome after ocean water exposure. Mar. Pollut. Bull. 2019, 145, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Holeton, C.; Chambers, P.A.; Grace, L. Wastewater release and its impacts on Canadian waters. Can. J. Fish Aquat. Sci. 2011, 68, 1836–1859. [Google Scholar] [CrossRef]
- O’Malley, M.L.; Lear, D.W.; Adams, W.N.; Gaines, J.; Sawyer, T.K.; Lewis, E.J. Microbial contamination of continental shelf sediments by wastewater. J. Water Pollut. Control Fed. 1982, 54, 1311–1317. [Google Scholar]
- Nelson, M.; Jones, S.H.; Edwards, C.; Ellis, J.C. Characterization of Escherichia coli populations from gulls, landfill trash, and wastewater using ribotyping. Dis. Aquat. Organ. 2008, 81, 53–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleisher, J.M.; Kay, D.; Salmon, R.L.; Jones, F.; Wyer, M.D.; Godfree, A.F. Marine waters contaminated with domestic sewage: Nonenteric illnesses associated with bather exposure in the United Kingdom. Am. J. Public Health 1996, 86, 1228–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fries, J.S.; Characklis, G.W.; Noble, R.T. Sediment–water exchange of Vibrio sp. and fecal indicator bacteria: Implications for persistence and transport in the Neuse River Estuary, North Carolina, USA. Water Res. 2008, 42, 941–950. [Google Scholar] [CrossRef]
- Fries, J.S.; Characklis, G.W.; Noble, R.T. Attachment of fecal indicator bacteria to particles in the Neuse River Estuary, N.C. J. Environ. Eng. 2006, 132, 1338–1345. [Google Scholar] [CrossRef]
- Griffin, D.W.; Kellogg, C.A.; Garrison, V.H.; Shinn, E.A. The global transport of dust: An Intercontinental river of dust, microorganisms and toxic chemicals flows through the Earth’s atmosphere. Am. Sci. 2002, 90, 228–235. [Google Scholar] [CrossRef]
- Prospero, J.M.; Blades, E.; Mathison, G.; Naidu, R. Interhemispheric transport of viable fungi and bacteria from Africa to the Caribbean with soil dust. Aerobiologia 2005, 21, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Okazaki, K.; Kawaoka, Y.; Takada, A.; Webster, R.G.; Kida, H. Perpetuation of influenza A viruses in Alaskan waterfowl reservoirs. Arch. Virol. 1995, 140, 1163–1172. [Google Scholar] [CrossRef] [PubMed]
- Anthony, S.J.; Leger, J.A.S.; Pugliares, K.; Ip, H.S.; Chan, J.M.; Carpenter, Z.W.; Navarrete-Macias, I.; Sanchez-Leon, M.; Saliki, J.T.; Pedersen, J.; et al. Emergence of fatal avian influenza in New England harbor seals. mBio 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Dewar, M.L. Gastrointestinal Microbiota of Seabirds. Ph.D. Thesis, School of Exercise and Nutrition Sciences, Deakin University, Geelong, Australia, 2012. [Google Scholar]
- Zhao, X.; Han, Y.; Ren, S.; Ma, Y.; Li, H.; Peng, X. L-proline increases survival of tilapias infected by Streptococcus agalactiae in higher water temperature. Fish Shellfish Immunol. 2015, 44, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Troussellier, M.; Escalas, A.; Bouvier, T.; Mouillot, D. Sustaining rare marine microorganisms: Macroorganisms as repositories and dispersal agents of microbial diversity. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Acevedo-Whitehouse, K.; Rocha-Gosselin, A.; Gendron, D. A novel non-invasive tool for disease surveillance of free-ranging whales and its relevance to conservation programs. Anim. Conserv. 2010, 13, 217–225. [Google Scholar] [CrossRef]
- Hermosilla, C.; Hirzmann, J.; Silva, L.M.R.; Brotons, J.M.; Cerdà, M.; Prenger-Berninghoff, E.; Ewers, C.; Taubert, A. Occurrence of anthropozoonotic parasitic infections and faecal microbes in free-ranging sperm whales (Physeter macrocephalus) from the Mediterranean Sea. Parasitol. Res. 2018, 117, 2531–2541. [Google Scholar] [CrossRef]
- Greenwood, A.G.; Taylor, D.C. Captive killer whales in Europe. Aquat. Mamm. 1985, 1, 10–12. [Google Scholar]
- Goertz, C.E.C.; Polasek, L.; Burek, K.; Suydam, R.; Sformo, T. Demography and pathology of a pacific walrus (Odobenus rosmarus divergens) mass-mortality event at Icy Cape, Alaska, September 2009. Polar Biol. 2017, 40, 989–996. [Google Scholar] [CrossRef]
- Zamri-Saad, M.; Amal, M.N.A.; Siti-Zahrah, A. Pathological changes in red tilapias (Oreochromis spp.) naturally infected by Streptococcus agalactiae. J. Comp. Pathol. 2010, 143, 227–229. [Google Scholar] [CrossRef]
- Shoemaker, C.A.; Evans, J.J.; Klesius, P.H. Density and dose: Factors affecting mortality of Streptococcus iniae infected tilapia (Oreochromis niloticus). Aquaculture 2000, 188, 229–235. [Google Scholar] [CrossRef]
- Agnew, W.; Barnes, A.C. Streptococcus iniae: An aquatic pathogen of global veterinary significance and a challenging candidate for reliable vaccination. Vet. Microbiol. 2007, 122, 1–15. [Google Scholar] [CrossRef]
- Amsallem, M.; Iung, B.; Bouleti, C.; Armand-Lefevre, L.; Eme, A.-L.; Touati, A.; Kirsch, M.; Duval, X.; Vahanian, A. First reported human case of native mitral infective endocarditis caused by Streptococcus canis. Can. J. Cardiol 2014, 30, 1462.e1–1462.e2. [Google Scholar] [CrossRef]
- Abbott, Y.; Acke, E.; Khan, S.; Muldoon, E.G.; Markey, B.K.; Pinilla, M.; Leonard, F.C.; Steward, K.; Waller, A. Zoonotic transmission of Streptococcus equi subsp. zooepidemicus from a dog to a handler. J. Med. Microbiol. 2010, 59, 120–123. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Occurrence of streptococcal species described in different marine mammals. Streptococcal species that have been isolated and identified at least twice in pinnipeds (A) and cetaceans (B). (C) shows a world map indicating location of streptococcal species detected in marine mammals.
Figure 1.
Occurrence of streptococcal species described in different marine mammals. Streptococcal species that have been isolated and identified at least twice in pinnipeds (A) and cetaceans (B). (C) shows a world map indicating location of streptococcal species detected in marine mammals.

Figure 2.
Schematic representation of the infection of marine mammals with (opportunistic) streptococci.
Figure 2.
Schematic representation of the infection of marine mammals with (opportunistic) streptococci.

Figure 3.
Possible ways how streptococci are introduced into marine environments and how they may be transmitted to and between marine mammals. Most indicated transmission routes are hypothetical based on general microbiological observations. In the other cases, there are studies showing that the transmission route is very likely or even confirmed. Hence, this figure displays scientific gaps that has still to be filled and where data are insufficient.
Figure 3.
Possible ways how streptococci are introduced into marine environments and how they may be transmitted to and between marine mammals. Most indicated transmission routes are hypothetical based on general microbiological observations. In the other cases, there are studies showing that the transmission route is very likely or even confirmed. Hence, this figure displays scientific gaps that has still to be filled and where data are insufficient.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Numberger, D.; Siebert, U.; Fulde, M.; Valentin-Weigand, P. Streptococcal Infections in Marine Mammals. Microorganisms 2021, 9, 350. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020350
AMA Style
Numberger D, Siebert U, Fulde M, Valentin-Weigand P. Streptococcal Infections in Marine Mammals. Microorganisms. 2021; 9(2):350. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020350
Chicago/Turabian StyleNumberger, Daniela, Ursula Siebert, Marcus Fulde, and Peter Valentin-Weigand. 2021. "Streptococcal Infections in Marine Mammals" Microorganisms 9, no. 2: 350. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020350
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.