
Development of a Fluorescent Tool for Studying Legionella bozemanae Intracellular Infection

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria, Plasmids, Primers, and Growth Conditions
2.2. Construction of GFP-Expressing Legionella Strains
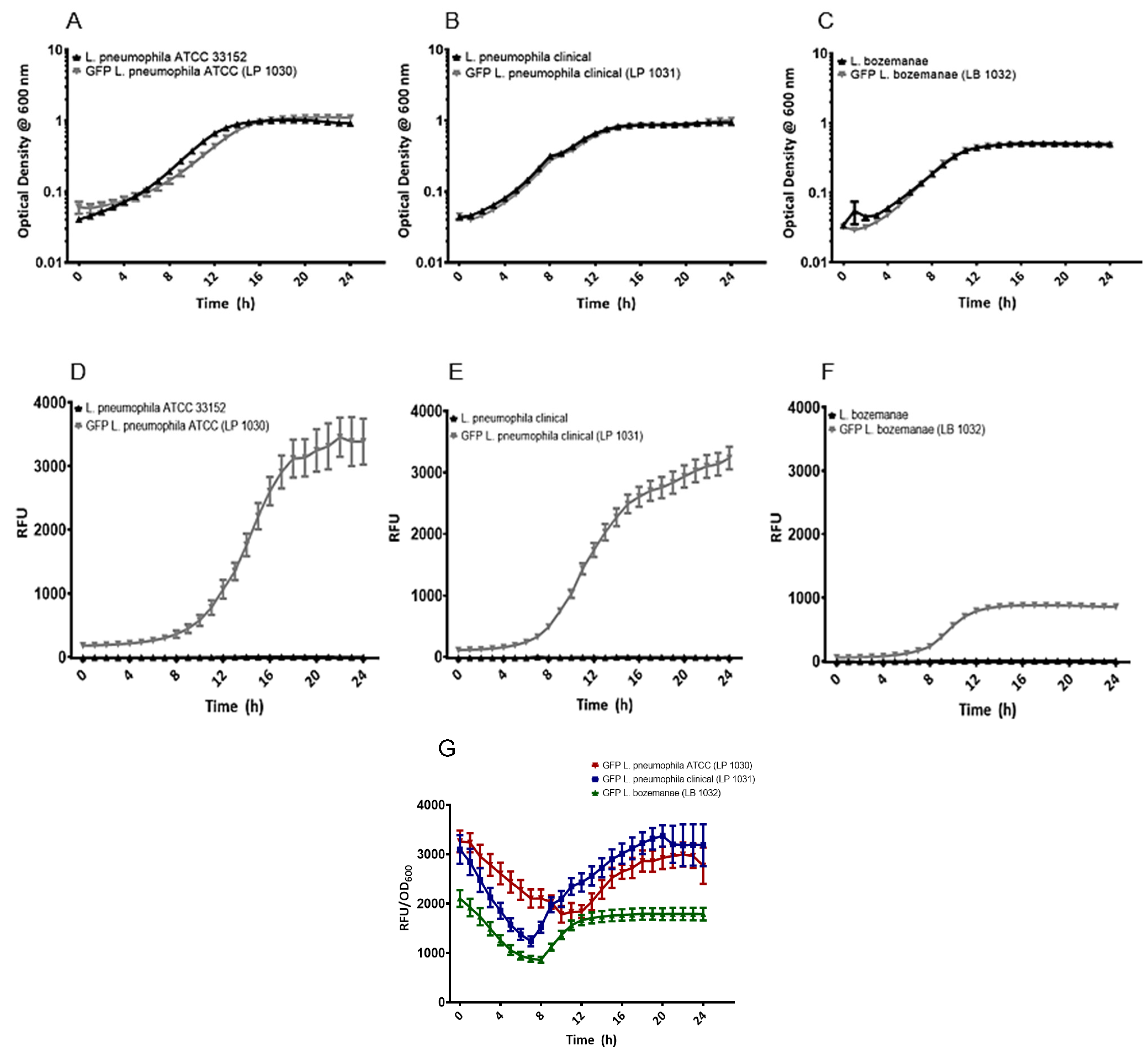
2.3. In Vitro Growth Kinetics and Relative Fluorescence Assays
2.4. Intracellular Growth Assays in Acanthamoeba castellanii
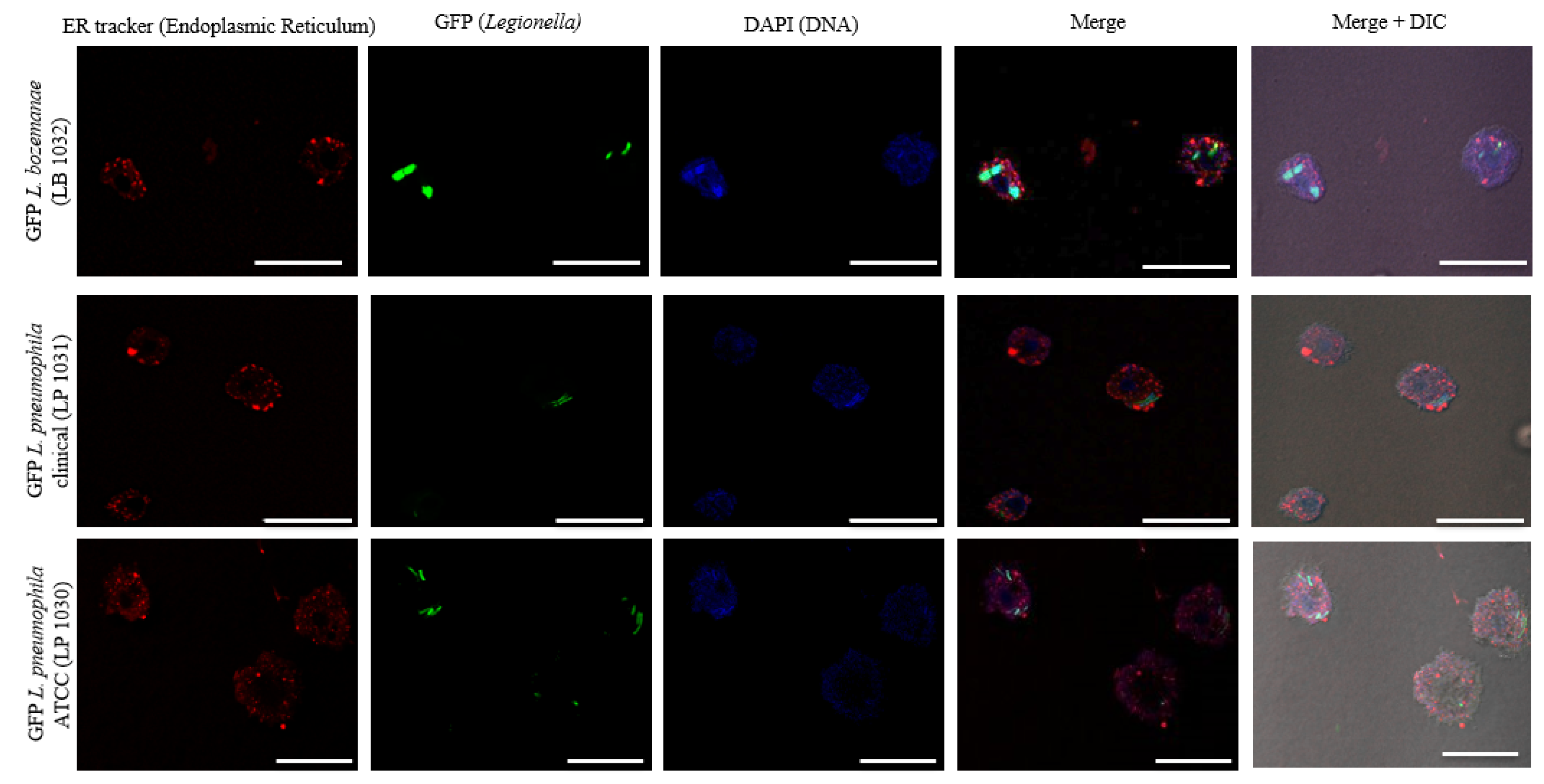
2.5. In Vitro and In Vivo Microscopic Imaging of the GFP-Expressing Legionella Strains
2.6. Statistical Analyses
3. Results
3.1. Generation of GFP-Expressing Legionella Strains
3.2. Plasmid-Borne GFP Expression Does Not Affect the Growth of Legionella In Vitro
3.3. Plasmid-Borne GFP Expression Does Not Affect Legionella Growth within A. castellanii protozoa
3.4. The Intracellular Life Cycle of L. bozemanae Is Similar to That of L. pneumophila
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benedict, K.M.; Reses, H.; Vigar, M.; Roth, D.M.; Roberts, V.A.; Mattioli, M.; Cooley, L.A.; Hilborn, E.D.; Wade, T.J.; Fullerton, K.E.; et al. Surveillance for waterborne disease outbreaks associated with drinking water- United States, 2013–2014. Morb. Mortal. Wkly. Rep. 2017, 66, 1216–1221. [Google Scholar] [CrossRef] [PubMed]
- NNDSS Annual Report Wroking Group. Australia’s Notifiable Disease Status, 2014: Annual Report of the National Notifiable Diseases Surveillance. System Commun. Dis. Intell. 2016, 40, 1. [Google Scholar]
- European Centre for Disease Prevention and Control (ECDC) Surveillance Atlas—Legionnaires Disease. Available online: https://www.ecdc.europa.eu/en/legionnaires-disease/surveillance/atlas (accessed on 25 January 2021).
- Public Health Agency of Canada Legionellosis Reported Cases from 1986 to 2018 in Canada. Available online: https://diseases.canada.ca/notifiable/charts?c=yl (accessed on 2 February 2021).
- European Centre for Disease Prevention and Contol. Annual Epidemiological Report for 2017 Legionnaires’ Disease; European Centre for Disease Prevention and Contol: Stockholm, Sweden, 2019. [Google Scholar]
- Centers for Disease Control and Prevention. Legionellosis—United States, 2000–2009; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2011; Volume 60. [Google Scholar]
- Collier, S.A.; Deng, L.; Adam, E.A.; Benedict, K.M.; Beshearse, E.M.; Blackstock, A.J.; Bruce, B.B.; Derado, G.; Edens, C.; Fullerton, K.E.; et al. Estimate of burden and direct healthcare cost of infectious waterborne disease in the United States. Emerg. Infect. Dis. 2021, 27, 140–149. [Google Scholar] [CrossRef]
- World Health Organization/Pan American Health Organization. Legionella and the Prevention of Legionellosis; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Cosson, P.; Soldati, T. Eat, kill or die: When amoeba meets bacteria. Curr. Opin. Microbiol. 2008, 11, 271–276. [Google Scholar] [CrossRef]
- Moffat, J.F.; Tompkins, L.S. A quantitative model of intracellular growth of Legionella pneumophila in Acanthamoeba castellanii. Infect. Immun. 1992, 60, 296–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gast, R.J.; Moran, D.M.; Dennett, M.R.; Wurtsbaugh, W.A.; Amaral-Zettler, L.A. Amoebae and Legionella pneumophila in saline environments. J. Water Health 2011, 9, 37–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- www.bacterio.net LPSN. Available online: http://www.bacterio.net/legionella.html (accessed on 5 July 2018).
- Doleans, A.; Aurell, H.; Reyrolle, M.; Lina, G.; Freney, J.; Vandenesch, F.; Etienne, J.; Jarraud, S. Clinical and Environmental Distributions of Legionella Strains in France Are Different. J. Clin. Microbiol. 2004, 42, 458–460. [Google Scholar] [CrossRef] [Green Version]
- Mercante, J.W.; Winchell, J.M. Current and emerging Legionella diagnostics for laboratory and outbreak investigations. Clin. Microbiol. Rev. 2015, 28, 95–133. [Google Scholar] [CrossRef] [Green Version]
- Jarraud, S.; Descours, G.; Ginevra, C.; Lina, G.; Etienne, J. Identification of Legionella in Clinical Samples. In Legionella: Methods and Protocols; Buchrieser, C., Hilbi, H., Eds.; Springer: New York, NY, USA, 2013; Volume 945, pp. 27–56. ISBN 9781627032384. [Google Scholar]
- Ng, V.; Tang, P.; Jamieson, F.; Guyard, C.; Low, D.E.; Fisman, D.N. Laboratory-based evaluation of legionellosis epidemiology in Ontario, Canada, 1978 to 2006. BMC Infect. Dis. 2009, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Phin, N.; Parry-Ford, F.; Harrison, T.; Stagg, H.R.; Zhang, N.; Kumar, K.; Lortholary, O.; Zumla, A.; Abubakar, I. Epidemiology and clinical management of Legionnaires’ disease. Lancet Infect. Dis. 2014, 14, 1011–1021. [Google Scholar] [CrossRef]
- Head, B.M.; Trajtman, A.; Bernard, K.; Burdz, T.; Vélez, L.; Herrera, M.; Vanessa, Z.; Keynan, Y. Legionella co-infection in HIV-associated pneumonia. Diagn. Microbiol. Infect. Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Mancini, G.; Erario, L.; Gianfreda, R.; Oliva, A.; Massetti, A.P.; Mastroianni, C.M.; Vullo, V. Tuberculosis and Legionella pneumophila pneumonia in a patient receiving anti-tumour necrosis factor- a (anti-TNF- a) treatment. Eur. Soc. Clin. Microbiol. Infect. Dis. 2007, 13, 1036–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, K.L.; Miceli, M.H.; Tarrand, J.J.; Kontoyiannis, D.P. Legionella pneumonia in cancer patients. Medicine (Baltimore) 2008, 87, 152–159. [Google Scholar] [CrossRef]
- Just, S.A.; Knudsen, J.B.; Uldum, S.A.; Holt, H.M. Detection of Legionella bozemanae, a new cause of septic arthritis, by PCR followed by specific culture. J. Clin. Microbiol. 2012, 50, 4180–4182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neiderud, C.-J.; Lagerqvist Vidh, A.; Salaneck, E. Soft tissue infection caused by Legionella bozemanii in a patient with ongoing immunosuppressive treatment. Infect. Ecol. Epidemiol. 2013, 3, 20739. [Google Scholar] [CrossRef]
- Becton Dickinson and Company BCYE Agar. Quality Control Procedures; Becton Dickinson and Company: East Rutherford, NJ, USA, 2016; pp. 1–5. [Google Scholar]
- Gogoi, S.K.; Gopinath, P.; Paul, A.; Ramesh, A.; Ghosh, S.S.; Chattopadhyay, A. Green fluorescent protein-expressing Escherichia coli as a model system for investigating the antimicrobial activities of silver nanoparticles. Langmuir 2006, 22, 9322–9328. [Google Scholar] [CrossRef]
- Stretton, S.; Techkarnjanaruk, S.; Mclennan, A.M.; Goodman, A.E. Use of green fluorescent protein to tag and investigate gene expression in marine bacteria. Appl. Environ. Microbiol. 1998, 64, 2554–2559. [Google Scholar] [CrossRef] [Green Version]
- Morash, M.G.; Brassinga, A.K.C.; Warthan, M.; Gourabathini, P.; Garduño, R.A.; Goodman, S.D.; Hoffman, P.S. Reciprocal expression of integration host factor and HU in the developmental cycle and infectivity of Legionella pneumophila. Appl. Environ. Microbiol. 2009, 75, 1826–1837. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, C.; Harrison, C.F.; Hilbi, H. The natural alternative: Protozoa as cellular models for Legionella infection. Cell. Microbiol. 2014, 16, 15–26. [Google Scholar] [CrossRef]
- Barbier, M.; Damron, F.H. Rainbow vectors for broad-range bacterial fluorescence labeling. PLoS ONE 2016, 11, 1–16. [Google Scholar] [CrossRef]
- Tombolini, R.; Unge, A.; Davey, M.E.; De Bruijn, F.J.; Jansson, J.K. Flow cytometric and microscopic analysis of GFP-tagged Pseudomonas fluorescens bacteria. FEMS Microbiol. Ecol. 1997, 22, 17–28. [Google Scholar] [CrossRef]
- Aye, T.; Wachsmuth, K.; Feeley, J.C.; Gibson, R.J.; Johnson, S.R. Plasmid profiles of Legionella species. Curr. Microbiol. 1981, 6, 389–394. [Google Scholar] [CrossRef]
- Kain, S.R. Methods and protocols. Methods Biochem. Anal. 2005, 47, 407–421. [Google Scholar] [CrossRef]
- Feeley, J.C.; Gibson, R.J.; Gorman, G.W.; Langford, N.C.; Rasheed, J.K.; Mackel, D.C.; Baine, W.B. Charcoal-yeast extract agar: Primary isolation medium for Legionella pneumophila. J. Clin. Microbiol. 1979, 10, 437–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, J.J.; Zylstra, G.J. Plasposons: Modular self-cloning minitransposon derivatives for rapid genetic analysis of gram-negative bacterial genomes. Appl. Environ. Microbiol. 1998, 64, 2710–2715. [Google Scholar] [CrossRef] [Green Version]
- Bachman, M.A.; Swanson, M.S. Genetic evidence that Legionella pneumophila RpoS modulates expression of the transmission phenotype in both the exponential phase and the stationary phase. Infect. Immun. 2004, 72, 2468–2476. [Google Scholar] [CrossRef] [Green Version]
- Tanner, J.R.; Li, L.; Faucher, S.P.; Brassinga, A.K.C. The CpxRA two-component system contributes to Legionella pneumophila virulence. Mol. Microbiol. 2016, 100, 1017–1038. [Google Scholar] [CrossRef] [Green Version]
- Berger, K.H.; Isberg, R.R. Two distinct defects in intracellular growth complemented by a single genetic locus in Legionella pneumophila. Mol. Microbiol. 1993, 7, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Pitre, C.A.J.; Tanner, J.R.; Patel, P.; Brassinga, A.K.C. Regulatory control of temporally expressed integration host factor (IHF) in Legionella pneumophila. Microbiol. (U. K.) 2013, 159, 475–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietersdorfer, E.; Cervero-Aragó, S.; Sommer, R.; Kirschner, A.K.; Walochnik, J. Optimized methods for Legionella pneumophila release from its Acanthamoeba hosts. BMC Microbiol. 2016, 16, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M. Intracellular survival and replication of legionella pneumophila within host cells. Yakugaku zasshi J. Pharm. Soc. Japan 2008, 128, 1763–1770. [Google Scholar] [CrossRef]
- Joshi, A.D.; Swanson, M.S. Comparative analysis of Legionella pneumophila and Legionella micdadei virulence traits. Infect. Immun. 1999, 67, 4134–4142. [Google Scholar] [CrossRef] [Green Version]
- Terry Alli, O.A.; Zink, S.; von Lackum, N.K.; Abu-kwaik, Y. Comparative assessment of virulence traits in Legionella spp. Microbiology 2003, 149, 631–641. [Google Scholar] [CrossRef] [Green Version]
- Qin, T.; Zhou, H.; Ren, H.; Liu, W. Distribution of secretion systems in the genus Legionella and its correlation with pathogenicity. Front. Microbiol. 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Saito, M.; Tanaka, T.; Amako, K.; Yoshida, S. ichi Comparative analysis of virulence traits between a Legionella feeleii strain implicated in Pontiac fever and a strain that caused Legionnaires’ disease. Microb. Pathog. 2015, 89, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Neumeister, B.; Schöniger, S.; Faigle, M.; Eichner, M.; Dietz, K. Multiplication of different Legionella species in Mono Mac 6 cells and in Acanthamoeba castellanii. Appl. Environ. Microbiol. 1997, 63, 1219–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, H.J.; Ang, D.K.Y.; Van Driel, I.R.; Hartland, E.L. Molecular pathogenesis of infections caused by Legionella pneumophila. Clin. Microbiol. Rev. 2010, 23, 274–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kjos, M.; Aprianto, R.; Fernandes, V.E.; Andrew, P.W.; Van Strijp, J.A.G.; Nijland, R.; Veening, J.W. Bright fluorescent Streptococcus pneumoniae for live-cell imaging of host-pathogen interactions. J. Bacteriol. 2015, 197, 807–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jørgensen, M.G.; van Raaphorst, R.; Veening, J.W. Noise and Stochasticity in Gene Expression. A Pathogenic Fate Determinant. Methods Microbiol. 2013, 40, 157–175. [Google Scholar] [CrossRef]
- Hiltz, M.F.; Sisson, G.R.; Brassinga, A.K.C.; Garduno, E.; Garduno, R.A.; Hoffman, P.S. Expression of magA in Legionella pneumophila Philadelphia-1 Is Developmentally Regulated and a Marker of Formation of Mature Intracellular Forms. J. Bacteriol. 2004, 186, 3038–3045. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organisms | Genotype | Reference/Source |
|---|---|---|
| Legionella pneumophila ATCC 33,152 Philadelphia-1 | Parental | ATCC |
| Legionella pneumophila 950 clinical isolate | Parental | Health Sciences Centre (Winnipeg, Canada) |
| Legionella bozemanae 948 clinical isolate | Parental | Health Sciences Centre (Winnipeg, Canada) |
| LP 1030 | L. pneumophila ATCC 33,152 pGFP-Kan; Kanr | This study |
| LP 1031 | L. pneumophila 950 pGFP-Kan; Kanr | This study |
| LB 1032 | L. bozemanae 948 pGFP-Kan; Kanr | This study |
| E. coli DH5α | F’ endA1 hsdR17(rk- mk-) supE44 thi-1 recA1 gyrA (Nalr) relA1 Δ(lacZYA-argF)U169 deoR(Φ80lacΔ(lacZ)M15) | Laboratory stock (New England Biolabs) |
| Acanthamoeba castellanii (ATCC 30234) | Douglas | ATCC |
| Plasmids | ||
| p34S-Kan | Cloning vector with Kanr cassette | Laboratory stock Dennis et al., 1998 [33] |
| pKB127 | 245 bp PmagA region cloned into BamHI and XbaI sites of pBH6119; Ampr, Thy+ | Morash et al., 2009 [26] |
| pGFP-Kan | pBH6119 PmagA; Kanr | This study |
| Primer | Sequence (5′ to 3′) | Annealing Temperature (°C) | Amplicon Size (bp) |
|---|---|---|---|
| pF KanR EcoRI | CGCATAgaattcCCACGTTGTGTCTCAAAATCTCTG | 60 | 1120 |
| pR KanR SacII | CGCATAccgcggGGTTGATGAGAGCTTTGTTGTAG | 60 | |
| pF KanR Conf Int | GTTGCATTCGATTCCTGTTTG | 60 | 179 |
| pR KanR Conf Int | GTGAGAATGGCAAAAGCTTATG | 60 | |
| pF KanR Conf Ext | GTGCCCATTAACATCACCATC | 60 | 1200 a |
| pR KanR Conf Rev | GTTATTTCTCCGGATTTAATTCG | 60 | 726 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Head, B.M.; Graham, C.I.; MacMartin, T.; Keynan, Y.; Brassinga, A.K.C. Development of a Fluorescent Tool for Studying Legionella bozemanae Intracellular Infection. Microorganisms 2021, 9, 379. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020379
Head BM, Graham CI, MacMartin T, Keynan Y, Brassinga AKC. Development of a Fluorescent Tool for Studying Legionella bozemanae Intracellular Infection. Microorganisms. 2021; 9(2):379. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020379
Chicago/Turabian StyleHead, Breanne M., Christopher I. Graham, Teassa MacMartin, Yoav Keynan, and Ann Karen C. Brassinga. 2021. "Development of a Fluorescent Tool for Studying Legionella bozemanae Intracellular Infection" Microorganisms 9, no. 2: 379. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020379