
From Dysbiosis to Healthy Skin: Major Contributions of Cutibacterium acnes to Skin Homeostasis

 , ,
, ,
Abstract
:1. Cutibacterium and Cutibacterium acnes in the Skin Microbiome


2. Major Contributions of Cutibacterium acnes to Skin Homeostasis
2.1. Lipid Modulation
2.2. Follicular Niche Competition
2.3. Immune Modulation
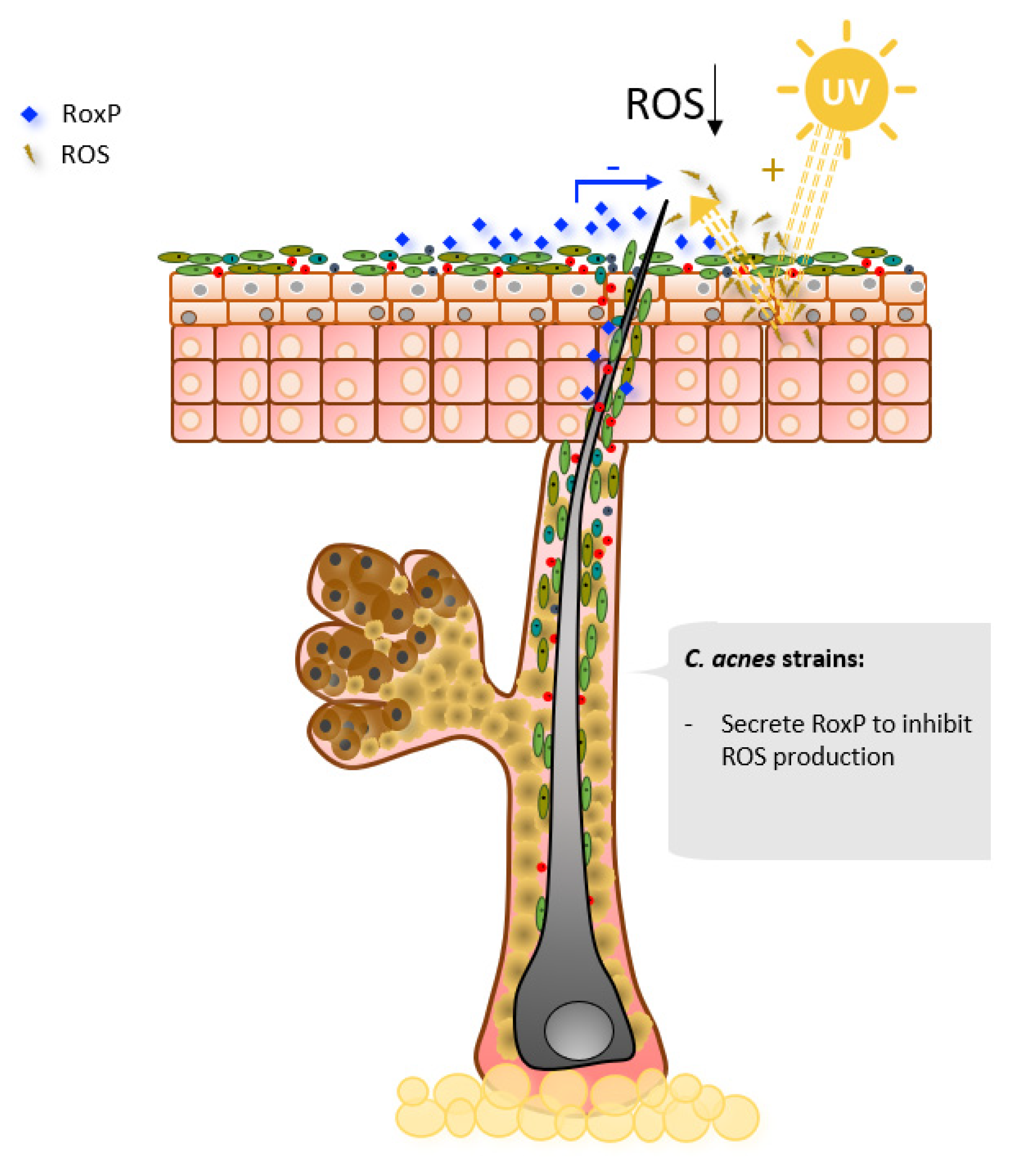
2.4. Oxidative Stress Mitigation
3. How to Prevent and/or Treat “On Site” Dysbiosis Disorders
3.1. Skin Microbiome Modulation Strategies
3.2. Bacteriotherapy in Dermatology
3.3. Probiotics Used in Dermatology
3.4. Safety Functionality and Technical Feasibility for Skin Probiotics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mosteller, R.D. Simplified Calculation of Body-Surface Area. N. Engl. J. Med. 1987, 317, 1098. [Google Scholar] [CrossRef] [PubMed]
- Kabashima, K.; Honda, T.; Ginhoux, F.; Egawa, G. The Immunological Anatomy of the Skin. Nat. Rev. Immunol. 2019, 19, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.; Geyer, S.; Weninger, W.; Guimberteau, J.-C.; Wong, J.K. The Dynamic Anatomy and Patterning of Skin. Exp. Dermatol. 2016, 25, 92–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, R.L. Human Skin Is the Largest Epithelial Surface for Interaction with Microbes. J. Investig. Dermmatol. 2017, 137, 1213–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahns, A.C.; Alexeyev, O.A. Three Dimensional Distribution of Propionibacterium Acnes Biofilms in Human Skin. Exp. Dermmatol. 2014, 23, 687–689. [Google Scholar] [CrossRef] [Green Version]
- Nakatsuji, T.; Chiang, H.-I.; Jiang, S.B.; Nagarajan, H.; Zengler, K.; Gallo, R.L. The Microbiome Extends to Subepidermal Compartments of Normal Skin. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.; Byrd, A.L.; Deming, C.; Conlan, S.; Kong, H.H.; Segre, J.A. Biogeography and Individuality Shape Function in the Human Skin Metagenome. Nature 2014, 514, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlotta De, L.R. Mycobiota: Micro-Eukaryotes Inhabiting Our Body as Commensals or Opportunistic Pathogens. Fungal. Genom. Biol. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Underhill, D.M.; Iliev, I.D. The Mycobiota: Interactions between Commensal Fungi and the Host Immune System. Nat. Rev. Immunol. 2014, 14, 405–416. [Google Scholar] [CrossRef]
- Hannigan, G.D.; Meisel, J.S.; Tyldsley, A.S.; Zheng, Q.; Hodkinson, B.P.; SanMiguel, A.J.; Minot, S.; Bushman, F.D.; Grice, E.A. The Human Skin Double-Stranded DNA Virome: Topographical and Temporal Diversity, Genetic Enrichment, and Dynamic Associations with the Host Microbiome. mBio 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Lecuit, M.; Eloit, M. The Human Virome: New Tools and Concepts. Trends Microbiol. 2013, 21, 510–515. [Google Scholar] [CrossRef]
- Grice, E.A.; Kong, H.H.; Renaud, G.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Wolfsberg, T.G.; Turner, M.L.; Segre, J.A. A Diversity Profile of the Human Skin Microbiota. Genome Res. 2008, 18, 1043–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; Green, E.D.; et al. Topographical and Temporal Diversity of the Human Skin Microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, E.K.; Lauber, C.L.; Hamady, M.; Fierer, N.; Gordon, J.I.; Knight, R. Bacterial Community Variation in Human Body Habitats Across Space and Time. Science 2009, 326, 1694–1697. [Google Scholar] [CrossRef] [Green Version]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The Human Skin Microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Grice, E.A.; Segre, J.A. The Skin Microbiome. Nat. Rev. Microbiol 2011, 9, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.; Schoenfeld, D.; Nomicos, E.; Park, M.; Becker, J.; et al. Topographic Diversity of Fungal and Bacterial Communities in Human Skin. Nature 2013, 498. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.; Zeng, D.-N.; Chi, L.; Tan, Y.; Galzote, C.; Cardona, C.; Lax, S.; Gilbert, J.; Quan, Z.-X. The Influence of Age and Gender on Skin-Associated Microbial Communities in Urban and Rural Human Populations. PLoS ONE 2015, 10, e0141842. [Google Scholar] [CrossRef]
- Oh, J.; Byrd, A.L.; Park, M.; Kong, H.H.; Segre, J.A. Temporal Stability of the Human Skin Microbiome. Cell 2016, 165, 854–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebeer, S.; Spacova, I. Exploring Human Host–Microbiome Interactions in Health and Disease—How to Not Get Lost in Translation. Genome Biol 2019, 20, 56. [Google Scholar] [CrossRef] [Green Version]
- Scholz, C.F.P.; Kilian, M. The Natural History of Cutaneous Propionibacteria, and Reclassification of Selected Species within the Genus Propionibacterium to the Proposed Novel Genera Acidipropionibacterium Gen. Nov., Cutibacterium Gen. Nov. and Pseudopropionibacterium Gen. Nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 4422–4432. [Google Scholar] [CrossRef] [PubMed]
- Fournière, M.; Latire, T.; Souak, D.; Feuilloley, M.G.J.; Bedoux, G. Staphylococcus Epidermidis and Cutibacterium acnes: Two Major Sentinels of Skin Microbiota and the Influence of Cosmetics. Microorganisms 2020, 8, 1752. [Google Scholar] [CrossRef] [PubMed]
- Schommer, N.N.; Gallo, R.L. Structure and Function of the Human Skin Microbiome. Trends Microbiol. 2013, 21, 660–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grice, E.A. The Skin Microbiome: Potential for Novel Diagnostic and Therapeutic Approaches to Cutaneous Disease. Semin. Cutan. Med. Surg. 2014, 33, 98–103. [Google Scholar] [CrossRef] [Green Version]
- Chien, A.L.; Tsai, J.; Leung, S.; Mongodin, E.F.; Nelson, A.M.; Kang, S.; Garza, L.A. Association of Systemic Antibiotic Treatment of Acne With Skin Microbiota Characteristics. JAMA Dermmatol. 2019, 155, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, S.; Novak, N. Atopic Dermatitis. Available online: https://pubmed.ncbi.nlm.nih.gov/26377142/ (accessed on 26 January 2021).
- Larsen, F.S.; Hanifin, J.M. Epidemiology of Atopic Dermatitis. Immunol. Allergy Clin. North. Am. 2002, 22, 1–24. [Google Scholar] [CrossRef]
- Williams, M.R.; Gallo, R.L. Evidence That Human Skin Microbiome Dysbiosis Promotes Atopic Dermatitis. J. Investig. Dermatol. 2017, 137, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Francuzik, W.; Franke, K.; Schumann, R.; Heine, G.; Worm, M. Propionibacterium Acnes Abundance Correlates Inversely with Staphylococcus Aureus: Data from Atopic Dermatitis Skin Microbiome. Acta Dermmatol. Venereol. 2018, 98. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.H.; Oh, J.; Deming, C.; Conlan, S.; Grice, E.A.; Beatson, M.A.; Nomicos, E.; Polley, E.C.; Komarow, H.D.; Murray, P.R.; et al. Temporal Shifts in the Skin Microbiome Associated with Disease Flares and Treatment in Children with Atopic Dermatitis. Genome Res. 2012, 22, 850–859. [Google Scholar] [CrossRef] [Green Version]
- Nakatsuji, T.; Chen, T.H.; Narala, S.; Chun, K.A.; Two, A.M.; Yun, T.; Shafiq, F.; Kotol, P.F.; Bouslimani, A.; Melnik, A.V.; et al. Antimicrobials from Human Skin Commensal Bacteria Protect against Staphylococcus Aureus and Are Deficient in Atopic Dermatitis. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Gether, L.; Overgaard, L.K.; Egeberg, A.; Thyssen, J.P. Incidence and Prevalence of Rosacea: A Systematic Review and Meta-Analysis. Br. J. Dermatol. 2018, 179, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Casas, C.; Paul, C.; Lahfa, M.; Livideanu, B.; Lejeune, O.; Alvarez-Georges, S.; Saint-Martory, C.; Degouy, A.; Mengeaud, V.; Ginisty, H.; et al. Quantification of Demodex Folliculorum by PCR in Rosacea and Its Relationship to Skin Innate Immune Activation. Exp. Dermmatol. 2012, 21, 906–910. [Google Scholar] [CrossRef] [PubMed]
- Moran, E.M.; Foley, R.; Powell, F.C. Demodex and Rosacea Revisited. Clin. Dermmatol. 2017, 35, 195–200. [Google Scholar] [CrossRef]
- Clanner-Engelshofen, B.M.; French, L.E.; Reinholz, M. Corynebacterium Kroppenstedtii Subsp. Demodicis Is the Endobacterium of Demodex Folliculorum. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 1043–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubiak, K.; Sielawa, H.; Chen, W.; Dzika, E. Endosymbiosis and Its Significance in Dermatology. J. Eur. Acad. Dermmatol. Venereol. 2018, 32, 347–354. [Google Scholar] [CrossRef]
- Chen, W.; Plewig, G. Are Demodex Mites Principal, Conspirator, Accomplice, Witness or Bystander in the Cause of Rosacea? Am. J. Clin. Dermmatol. 2015, 16, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Woo, Y.R.; Lee, S.H.; Cho, S.H.; Lee, J.D.; Kim, H.S. Characterization and Analysis of the Skin Microbiota in Rosacea: Impact of Systemic Antibiotics. J. Clin. Med. 2020, 9, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Farhat, M.; Na, J.; Li, R.; Wu, Y. Bacterial and Fungal Microbiome Characterization in Patients with Rosacea and Healthy Controls. Br. J. Dermmatol. 2020, 183, 1112–1114. [Google Scholar] [CrossRef]
- Nestle, F.O.; Kaplan, D.H.; Barker, J. Psoriasis. N. Engl. J. Med. 2009, 361, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, C.E.; Barker, J.N. Pathogenesis and Clinical Features of Psoriasis. Lancet 2007, 370, 263–271. [Google Scholar] [CrossRef]
- Alekseyenko, A.V.; Perez-Perez, G.I.; De Souza, A.; Strober, B.; Gao, Z.; Bihan, M.; Li, K.; Methé, B.A.; Blaser, M.J. Community Differentiation of the Cutaneous Microbiota in Psoriasis. Microbiome 2013, 1, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Tseng, C.; Strober, B.E.; Pei, Z.; Blaser, M.J. Substantial Alterations of the Cutaneous Bacterial Biota in Psoriatic Lesions. PLoS ONE 2008, 3, e2719. [Google Scholar] [CrossRef] [PubMed]
- Fahlén, A.; Engstrand, L.; Baker, B.S.; Powles, A.; Fry, L. Comparison of Bacterial Microbiota in Skin Biopsies from Normal and Psoriatic Skin. Arch. Dermmatol. Res. 2012, 304, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Tett, A.; Pasolli, E.; Farina, S.; Truong, D.T.; Asnicar, F.; Zolfo, M.; Beghini, F.; Armanini, F.; Jousson, O.; Sanctis, V.D.; et al. Unexplored Diversity and Strain-Level Structure of the Skin Microbiome Associated with Psoriasis. NPJ Biofilms Microbiomes 2017, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Stehlikova, Z.; Kostovcik, M.; Kostovcikova, K.; Kverka, M.; Juzlova, K.; Rob, F.; Hercogova, J.; Bohac, P.; Pinto, Y.; Uzan, A.; et al. Dysbiosis of Skin Microbiota in Psoriatic Patients: Co-Occurrence of Fungal and Bacterial Communities. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Chang, H.-W.; Yan, D.; Singh, R.; Liu, J.; Lu, X.; Ucmak, D.; Lee, K.; Afifi, L.; Fadrosh, D.; Leech, J.; et al. Alteration of the Cutaneous Microbiome in Psoriasis and Potential Role in Th17 Polarization. Microbiome 2018, 6, 154. [Google Scholar] [CrossRef]
- Quan, C.; Chen, X.-Y.; Li, X.; Xue, F.; Chen, L.-H.; Liu, N.; Wang, B.; Wang, L.-Q.; Wang, X.-P.; Yang, H.; et al. Psoriatic Lesions Are Characterized by Higher Bacterial Load and Imbalance between Cutibacterium and Corynebacterium. J. Am. Acad. Dermatol. 2020, 82, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Tuchayi, S.M.; Makrantonaki, E.; Ganceviciene, R.; Dessinioti, C.; Feldman, S.R.; Zouboulis, C.C. Acne Vulgaris. Nat. Rev. Dis. Primers 2015, 1, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Ju, Q.; Tao, T.; Hu, T.; Karadağ, A.S.; Al-Khuzaei, S.; Chen, W. Sex Hormones and Acne. Clin. Dermatol. 2017, 35, 130–137. [Google Scholar] [CrossRef]
- Heng, A.H.S.; Chew, F.T. Systematic Review of the Epidemiology of Acne Vulgaris. Sci. Rep. 2020, 10, 5754. [Google Scholar] [CrossRef] [Green Version]
- Williams, H.C.; Dellavalle, R.P.; Garner, S. Acne Vulgaris. Lancet 2012, 379, 361–372. [Google Scholar] [CrossRef]
- Collier, C.N.; Harper, J.C.; Cafardi, J.A.; Cantrell, W.C.; Wang, W.; Foster, K.W.; Elewski, B.E. The Prevalence of Acne in Adults 20 Years and Older. J. Am. Acad. Dermmatol. 2008, 58, 56–59. [Google Scholar] [CrossRef]
- Mahmood, N.F.; Shipman, A.R. The Age-Old Problem of Acne. Int J. Womens Dermmatol. 2016, 3, 71–76. [Google Scholar] [CrossRef]
- Dréno, B.; Dagnelie, M.A.; Khammari, A.; Corvec, S. The Skin Microbiome: A New Actor in Inflammatory Acne. Am. J. Clin. Dermmatol. 2020, 21, 18–24. [Google Scholar] [CrossRef]
- Lee, Y.B.; Byun, E.J.; Kim, H.S. Potential Role of the Microbiome in Acne: A Comprehensive Review. J. Clin. Med. 2019, 8, 987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitz-Gibbon, S.; Tomida, S.; Chiu, B.-H.; Nguyen, L.; Du, C.; Liu, M.; Elashoff, D.; Erfe, M.C.; Loncaric, A.; Kim, J.; et al. Propionibacterium Acnes Strain Populations in the Human Skin Microbiome Associated with Acne. J. Investig. Dermatol. 2013, 133, 2152–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dessinioti, C.; Katsambas, A. Propionibacterium Acnes and Antimicrobial Resistance in Acne. Clin. Dermmatol. 2017, 35, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Miura, Y.; Ishige, I.; Soejima, N.; Suzuki, Y.; Uchida, K.; Kawana, S.; Eishi, Y. Quantitative PCR of Propionibacterium Acnes DNA in Samples Aspirated from Sebaceous Follicles on the Normal Skin of Subjects with or without Acne. J. Med. Dent. Sci 2010, 57, 65–74. [Google Scholar] [PubMed]
- Omer, H.; McDowell, A.; Alexeyev, O.A. Understanding the Role of Propionibacterium Acnes in Acne Vulgaris: The Critical Importance of Skin Sampling Methodologies. Clin. Dermmatol. 2017, 35, 118–129. [Google Scholar] [CrossRef]
- Li, C.; You, Z.; Lin, Y.; Liu, H.; Su, J. Skin Microbiome Differences Relate to the Grade of Acne Vulgaris. J. Dermatol. 2019, 46, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Pécastaings, S.; Roques, C.; Nocera, T.; Peraud, C.; Mengeaud, V.; Khammari, A.; Dréno, B. Characterisation of Cutibacterium acnes Phylotypes in Acne and in vivo Exploratory Evaluation of Myrtacine®. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Corvec, S.; Dagnelie, M.-A.; Khammari, A.; Dréno, B. Taxonomy and Phylogeny of Cutibacterium (Formerly Propionibacterium) Acnes in Inflammatory Skin Diseases. Ann. De Dermatol. Et De Vénéréologie 2019, 146, 26–30. [Google Scholar] [CrossRef]
- McLaughlin, J.; Watterson, S.; Layton, A.M.; Bjourson, A.J.; Barnard, E.; McDowell, A. Propionibacterium Acnes and Acne Vulgaris: New Insights from the Integration of Population Genetic, Multi-Omic, Biochemical and Host-Microbe Studies. Microorganisms 2019, 7, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dagnelie, M.-A.; Corvec, S.; Saint-Jean, M.; Bourdès, V.; Nguyen, J.; Khammari, A.; Dréno, B. Decrease in Diversity of Propionibacterium Acnes Phylotypes in Patients with Severe Acne on the Back. Acta Dermmatol. Venereol. 2017, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v4: Recent Updates and New Developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDowell, A.; Barnard, E.; Nagy, I.; Gao, A.; Tomida, S.; Li, H.; Eady, A.; Cove, J.; Nord, C.E.; Patrick, S. An Expanded Multilocus Sequence Typing Scheme for Propionibacterium Acnes: Investigation of ‘Pathogenic’, ‘Commensal’ and Antibiotic Resistant Strains. PLoS ONE 2012, 7, e41480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-Access Bacterial Population Genomics: BIGSdb Software, the PubMLST.Org Website and Their Applications. Wellcome Open Res. 2018, 3. [Google Scholar] [CrossRef]
- Neville, B.A.; Forster, S.C.; Lawley, T.D. Commensal Koch’s Postulates: Establishing Causation in Human Microbiota Research. Curr. Opin. Microbiol. 2018, 42, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Heintz-Buschart, A.; Wilmes, P. Human Gut Microbiome: Function Matters. Trends Microbiol. 2018, 26, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Visconti, A.; Le Roy, C.I.; Rosa, F.; Rossi, N.; Martin, T.C.; Mohney, R.P.; Li, W.; de Rinaldis, E.; Bell, J.T.; Venter, J.C.; et al. Interplay between the Human Gut Microbiome and Host Metabolism. Nat. Commun. 2019, 10, 4505. [Google Scholar] [CrossRef] [Green Version]
- Platsidaki, E.; Dessinioti, C. Recent Advances in Understanding Propionibacterium Acnes (Cutibacterium acnes) in Acne. F1000Res 2018, 7, 1953. [Google Scholar] [CrossRef] [Green Version]
- Bruggemann, H. The Complete Genome Sequence of Propionibacterium Acnes, a Commensal of Human Skin. Science 2004, 305, 671–673. [Google Scholar] [CrossRef]
- Holland, C.; Mak, T.N.; Zimny-Arndt, U.; Schmid, M.; Meyer, T.F.; Jungblut, P.R.; Brüggemann, H. Proteomic Identification of Secreted Proteins of Propionibacterium Acnes. BMC Microbiol. 2010, 10, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and Functional Importance in the Gut. Proc. Nutr. Soc. 2020, 1–13. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of Short Chain Fatty Acids by the Gut Microbiota and Their Impact on Human Metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, G.J.M.; Brüggemann, H. Bacterial Skin Commensals and Their Role as Host Guardians. Benef. Microbes 2014, 5, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Proksch, E. PH in Nature, Humans and Skin. J. Dermmatol. 2018, 45, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Proksch, E.; Brandner, J.M.; Jensen, J.-M. The Skin: An Indispensable Barrier. Exp. Dermatol. 2008, 17, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Boer, D.E.C.; van Smeden, J.; Al-Khakany, H.; Melnik, E.; van Dijk, R.; Absalah, S.; Vreeken, R.J.; Haenen, C.C.P.; Lavrijsen, A.P.M.; Overkleeft, H.S.; et al. Skin of Atopic Dermatitis Patients Shows Disturbed β-Glucocerebrosidase and Acid Sphingomyelinase Activity That Relates to Changes in Stratum Corneum Lipid Composition. Biochim. Et Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2020, 1865, 158673. [Google Scholar] [CrossRef] [PubMed]
- Surber, C.; Abels, C.; Maibach, H. pH in Atopic Dermatitis. Curr. Probl. Dermatol. 2018, 54, 95–107. [Google Scholar]
- Claesen, J.; Spagnolo, J.B.; Ramos, S.F.; Kurita, K.L.; Byrd, A.L.; Aksenov, A.A.; Melnik, A.V.; Wong, W.R.; Wang, S.; Hernandez, R.D.; et al. A Cutibacterium acnes Antibiotic Modulates Human Skin Microbiota Composition in Hair Follicles. Sci. Transl. Med. 2020, 12, eaay5445. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; O’Neill, A.M.; Williams, M.R.; Cau, L.; Nakatsuji, T.; Horswill, A.R.; Gallo, R.L. Short Chain Fatty Acids Produced by Cutibacterium acnes Inhibit Biofilm Formation by Staphylococcus Epidermidis. Sci. Rep. 2020, 10, 21237. [Google Scholar] [CrossRef]
- Parsek, M.R.; Singh, P.K. Bacterial Biofilms: An Emerging Link to Disease Pathogenesis. Annu. Rev. Microbiol. 2003, 57, 677–701. [Google Scholar] [CrossRef]
- Fujimura, S.; Nakamura, T. Purification and Properties of a Bacteriocin-Like Substance (Acnecin) of Oral Propionibacterium Acnes. Antimicrob. Agents Chemother. 1978, 14, 893–898. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.V.; Soulika, A.M. The Dynamics of the Skin’s Immune System. IJMS 2019, 20, 1811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naik, S.; Bouladoux, N.; Wilhelm, C.; Molloy, M.J.; Salcedo, R.; Kastenmuller, W.; Deming, C.; Quinones, M.; Koo, L.; Conlan, S.; et al. Compartmentalized Control of Skin Immunity by Resident Commensals. Science 2012, 337, 1115–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanke, I.; Steffen, H.; Christ, C.; Krismer, B.; Götz, F.; Peschel, A.; Schaller, M.; Schittek, B. Skin Commensals Amplify the Innate Immune Response to Pathogens by Activation of Distinct Signaling Pathways. J. Investig. Dermatol. 2011, 131, 382–390. [Google Scholar] [CrossRef] [Green Version]
- Sanford, J.A.; O’Neill, A.M.; Zouboulis, C.C.; Gallo, R.L. Short-Chain Fatty Acids from Cutibacterium acnes Activate Both a Canonical and Epigenetic Inflammatory Response in Human Sebocytes. J. Immunol. 2019, 202, 1767–1776. [Google Scholar] [CrossRef] [Green Version]
- Sanford, J.A.; Zhang, L.-J.; Williams, M.R.; Gangoiti, J.A.; Huang, C.-M.; Gallo, R.L. Inhibition of HDAC8 and HDAC9 by Microbial Short-Chain Fatty Acids Breaks Immune Tolerance of the Epidermis to TLR Ligands. Sci. Immunol. 2016, 1, eaah4609. [Google Scholar] [CrossRef]
- Yu, Y.; Champer, J.; Agak, G.W.; Kao, S.; Modlin, R.L.; Kim, J. Different Propionibacterium Acnes Phylotypes Induce Distinct Immune Responses and Express Unique Surface and Secreted Proteomes. J. Investig. Dermatol. 2016, 136, 2221–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Megyeri, K.; Orosz, L.; Bolla, S.; Erdei, L.; Rázga, Z.; Seprényi, G.; Urbán, E.; Szabó, K.; Kemény, L. Propionibacterium Acnes Induces Autophagy in Keratinocytes: Involvement of Multiple Mechanisms. J. Investig. Dermatol. 2018, 138, 750–759. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, J.L.; Sumpter, R.; Levine, B.; Hooper, L.V. Intestinal Epithelial Autophagy Is Essential for Host Defense against Invasive Bacteria. Cell Host Microbe 2013, 13, 723–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kistowska, M.; Meier, B.; Proust, T.; Feldmeyer, L.; Cozzio, A.; Kuendig, T.; Contassot, E.; French, L.E. Propionibacterium Acnes Promotes Th17 and Th17/Th1 Responses in Acne Patients. J. Investig. Dermatol. 2015, 135, 110–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, S.J.; Sullivan, B.M.; Peng, S.L.; Glimcher, L.H. Molecular Mechanisms RegulatinG Th1 Immune Responses. Annu. Rev. Immunol. 2003, 21, 713–758. [Google Scholar] [CrossRef]
- Brandt, E.B. Th2 Cytokines and Atopic Dermatitis. J. Clin. Cell Immunol 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, H.; Yamanaka, K.; Kakeda, M.; Inada, H.; Imai, Y.; Gabazza, E.C.; Kurokawa, I.; Mizutani, H. Propionibacterium Acnes Vaccination Induces Regulatory T Cells and Th1 Immune Responses and Improves Mouse Atopic Dermatitis: Letter to the Editor. Exp. Dermatol. 2011, 20, 157–158. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, K.; Yamanaka, K.; Linan, W.; Miyahara, Y.; Akeda, T.; Nakanishi, T.; Kitagawa, H.; Kakeda, M.; Kurokawa, I.; Shiku, H.; et al. Intratumoral Injection of Propionibacterium Acnes Suppresses Malignant Melanoma by Enhancing Th1 Immune Responses. PLoS ONE 2011, 6, e29020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu-Smith, F.; Jia, J.; Zheng, Y. UV-Induced Molecular Signaling Differences in Melanoma and Non-melanoma Skin Cancer. In Ultraviolet Light in Human Health, Diseases and Environment; Ahmad, S.I., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2017; Volume 996, pp. 27–40. ISBN 978-3-319-56016-8. [Google Scholar]
- Oberley, T.D. Oxidative Damage and Cancer. Am. J. Pathol. 2002, 160, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Xian, D.; Lai, R.; Song, J.; Xiong, X.; Zhong, J. Emerging Perspective: Role of Increased ROS and Redox Imbalance in Skin Carcinogenesis. Oxidative Med. Cell. Longev. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzaga, E.R. Role of UV Light in Photodamage, Skin Aging, and Skin Cancer: Importance of Photoprotection. Am. J. Clin. Dermatol. 2009, 10, 19–24. [Google Scholar] [CrossRef]
- Allhorn, M.; Arve, S.; Brüggemann, H.; Lood, R. A Novel Enzyme with Antioxidant Capacity Produced by the Ubiquitous Skin Colonizer Propionibacterium Acnes. Sci. Rep. 2016, 6, 36412. [Google Scholar] [CrossRef] [Green Version]
- Andersson, T.; Ertürk Bergdahl, G.; Saleh, K.; Magnúsdóttir, H.; Stødkilde, K.; Andersen, C.B.F.; Lundqvist, K.; Jensen, A.; Brüggemann, H.; Lood, R. Common Skin Bacteria Protect Their Host from Oxidative Stress through Secreted Antioxidant RoxP. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Sander, C.S.; Hamm, F.; Elsner, P.; Thiele, J.J. Oxidative Stress in Malignant Melanoma and Non-Melanoma Skin Cancer. Br. J. Dermatol. 2003, 148, 913–922. [Google Scholar] [CrossRef]
- Wood, D.L.A.; Lachner, N.; Tan, J.-M.; Tang, S.; Angel, N.; Laino, A.; Linedale, R.; Lê Cao, K.-A.; Morrison, M.; Frazer, I.H.; et al. A Natural History of Actinic Keratosis and Cutaneous Squamous Cell Carcinoma Microbiomes. mBio 2018, 9, e01432-18. [Google Scholar] [CrossRef] [Green Version]
- Ertürk, G.; Hedström, M.; Mattiasson, B.; Ruzgas, T.; Lood, R. Highly Sensitive Detection and Quantification of the Secreted Bacterial Benevolence Factor RoxP Using a Capacitive Biosensor: A Possible Early Detection System for Oxidative Skin Diseases. Plos ONE 2018, 13, e0193754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, J.; Maldonado-Gómez, M.X.; Martínez, I. To Engraft or Not to Engraft: An Ecological Framework for Gut Microbiome Modulation with Live Microbes. Curr. Opin. Biotechnol. 2018, 49, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Schrezenmeir, J.; de Vrese, M. Probiotics, Prebiotics, and Synbiotics—Approaching a Definition. Am. J. Clin. Nutr. 2001, 73, 361s–364s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakken, J.S.; Borody, T.; Brandt, L.J.; Brill, J.V.; Demarco, D.C.; Franzos, M.A.; Kelly, C.; Khoruts, A.; Louie, T.; Martinelli, L.P.; et al. Treating Clostridium Difficile Infection With Fecal Microbiota Transplantation. Clin. Gastroenterol. Hepatol. 2011, 9, 1044–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Agriculture Organization of the United Nations. Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation; World Health Organization, Ed.; FAO Food and Nutrition Paper; Food and Agriculture Organization of the United Nations; World Health Organization: Rome, Italy, 2006; ISBN 978-92-5-105513-7. [Google Scholar]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, Gut Microbiota, and Their Influence on Host Health and Disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernet, M.F.; Brassart, D.; Neeser, J.R.; Servin, A.L. Adhesion of Human Bifidobacterial Strains to Cultured Human Intestinal Epithelial Cells and Inhibition of Enteropathogen-Cell Interactions. Appl. Env. Microbiol. 1993, 59, 4121–4128. [Google Scholar] [CrossRef] [Green Version]
- Ouwehand, A.C.; Kirjavainen, P.V.; Grönlund, M.-M.; Isolauri, E.; Salminen, S.J. Adhesion of Probiotic Micro-Organisms to Intestinal Mucus. Int. Dairy J. 1999, 9, 623–630. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic Effects: Metabolic and Health Benefits. Br. J. Nutr. 2010, 104, S1–S63. [Google Scholar] [CrossRef] [Green Version]
- Piqué, N.; Berlanga, M.; Miñana-Galbis, D. Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar] [CrossRef] [Green Version]
- Collado, M.C.; Vinderola, G.; Salminen, S. Postbiotics: Facts and Open Questions. A Position Paper on the Need for a Consensus Definition. Benef. Microbes 2019, 10, 711–719. [Google Scholar] [CrossRef]
- Weill, F.S.; Cela, E.M.; Paz, M.L.; Ferrari, A.; Leoni, J.; González Maglio, D.H. Lipoteichoic Acid from Lactobacillus Rhamnosus GG as an Oral Photoprotective Agent against UV-Induced Carcinogenesis. Br. J. Nutr. 2013, 109, 457–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.-C.; Tomita, S.; Kleerebezem, M.; Bron, P.A. The Quest for Probiotic Effector Molecules--Unraveling Strain Specificity at the Molecular Level. Pharm. Res. 2013, 69, 61–74. [Google Scholar] [CrossRef]
- Jain, V.V.; Kitagaki, K.; Kline, J.N. CpG DNA and Immunotherapy of Allergic Airway Diseases. Clin. Exp. Allergy 2003, 33, 1330–1335. [Google Scholar] [CrossRef] [PubMed]
- Hynönen, U.; Palva, A. Lactobacillus Surface Layer Proteins: Structure, Function and Applications. Appl. Microbiol. Biotechnol. 2013, 97, 5225–5243. [Google Scholar] [CrossRef] [Green Version]
- Epstein, S. The Phenomenon of Microbial Uncultivability. Curr. Opin. Microbiol. 2013, 16, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Forssten, S.D.; Sindelar, C.W.; Ouwehand, A.C. Probiotics from an Industrial Perspective. Anaerobe 2011, 17, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Rong, J.; Shan, C.; Liu, S.; Zheng, H.; Liu, C.; Liu, M.; Jin, F.; Wang, L. Skin Resistance to UVB-Induced Oxidative Stress and Hyperpigmentation by the Topical Use of Lactobacillus Helveticus NS8-Fermented Milk Supernatant. J. Appl. Microbiol. 2017, 123, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Shu, M.; Wang, Y.; Yu, J.; Kuo, S.; Coda, A.; Jiang, Y.; Gallo, R.L.; Huang, C.-M. Fermentation of Propionibacterium Acnes, a Commensal Bacterium in the Human Skin Microbiome, as Skin Probiotics against Methicillin-Resistant Staphylococcus Aureus. PLoS ONE 2013, 8, e55380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callewaert, C.; Knödlseder, N.; Karoglan, A.; Güell, M.; Paetzold, B. Skin Microbiome Transplantation and Manipulation: Current State of the Art. Comput. Struct. Biotechnol. J. 2021, 19, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Gbassi, G.K.; Vandamme, T. Probiotic Encapsulation Technology: From Microencapsulation to Release into the Gut. Pharmaceutics 2012, 4, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Lisko, D.J.; Johnston, G.P.; Johnston, C.G. Effects of Dietary Yogurt on the Healthy Human Gastrointestinal (GI) Microbiome. Microorganisms 2017, 5, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Dunaway, S.; Champer, J.; Kim, J.; Alikhan, A. Changing Our Microbiome: Probiotics in Dermatology. Br. J. Dermatol. 2020, 182, 39–46. [Google Scholar] [CrossRef]
- Dimarzio, L.; Cinque, B.; Cupelli, F.; De Simone, C.; Cifone, M.G.; Giuliani, M. Increase of Skin-Ceramide Levels in Aged Subjects Following a Short-Term Topical Application of Bacterial Sphingomyelinase from Streptococcus Thermophilus. Int J. Immunopathol Pharm. 2008, 21, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Nodake, Y.; Matsumoto, S.; Miura, R.; Honda, H.; Ishibashi, G.; Matsumoto, S.; Dekio, I.; Sakakibara, R. Pilot Study on Novel Skin Care Method by Augmentation with Staphylococcus Epidermidis, an Autologous Skin Microbe—A Blinded Randomized Clinical Trial. J. Dermatol. Sci. 2015, 79, 119–126. [Google Scholar] [CrossRef]
- Guéniche, A.; Bastien, P.; Ovigne, J.M.; Kermici, M.; Courchay, G.; Chevalier, V.; Breton, L.; Castiel-Higounenc, I. Bifidobacterium Longum Lysate, a New Ingredient for Reactive Skin. Exp. Dermatol. 2010, 19, e1–e8. [Google Scholar] [CrossRef]
- Karoglan, A.; Paetzold, B.; De Lima, J.P.; Brüggemann, H.; Tüting, T.; Schanze, D.; Güell, M.; Gollnick, H. Safety and Efficacy of Topically Applied Selected Cutibacterium acnes Strains over Five Weeks in Patients with Acne Vulgaris: An Open-Label, Pilot Study. Acta Dermmatol. Venereol. 2019, 99, 1253–1257. [Google Scholar] [CrossRef] [Green Version]
- Butler, É.; Lundqvist, C.; Axelsson, J. Lactobacillus Reuteri DSM 17938 as a Novel Topical Cosmetic Ingredient: A Proof of Concept Clinical Study in Adults with Atopic Dermatitis. Microorganisms 2020, 8, 1026. [Google Scholar] [CrossRef]
- Kottner, J.; Lichterfeld, A.; Blume-Peytavi, U. Maintaining Skin Integrity in the Aged: A Systematic Review. Br. J. Dermatol. 2013, 169, 528–542. [Google Scholar] [CrossRef] [PubMed]
- Castillo, D.E.; Nanda, S.; Keri, J.E. Propionibacterium (Cutibacterium) Acnes Bacteriophage Therapy in Acne: Current Evidence and Future Perspectives. Dermatol. Ther. 2019, 9, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Wright, A. von Regulatory Aspects of Probiotics and Other Microbial Products Intended for Skin Care. In Skin Microbiome Handbook; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 321–341. ISBN 978-1-119-59305-8. [Google Scholar]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Paetzold, B.; Willis, J.R.; Pereira de Lima, J.; Knödlseder, N.; Brüggemann, H.; Quist, S.R.; Gabaldón, T.; Güell, M. Skin Microbiome Modulation Induced by Probiotic Solutions. Microbiome 2019, 7, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, C.F.P.; Jensen, A.; Lomholt, H.B.; Brüggemann, H.; Kilian, M. A Novel High-Resolution Single Locus Sequence Typing Scheme for Mixed Populations of Propionibacterium Acnes In Vivo. PLoS ONE 2014, 9, e4199. [Google Scholar] [CrossRef] [Green Version]
- Mayslich, C.; Grange, P.A.; Dupin, N. Cutibacterium acnes as an Opportunistic Pathogen: An Update of Its Virulence-Associated Factors. Microorganisms 2021, 9, 303. [Google Scholar] [CrossRef]
- REHBERGERt, T.G.; Glatz, B.A. Characterization of Propionibacterium Plasmidst. Appl. Env. Microbiol. 1990, 56, 8. [Google Scholar]
- Christensen, G.J.M.; Scholz, C.F.P.; Enghild, J.; Rohde, H.; Kilian, M.; Thürmer, A.; Brzuszkiewicz, E.; Lomholt, H.B.; Brüggemann, H. Antagonism between Staphylococcus Epidermidis and Propionibacterium Acnes and Its Genomic Basis. Bmc Genom. 2016, 17, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dréno, B.; Araviiskaia, E.; Berardesca, E.; Gontijo, G.; Sanchez Viera, M.; Xiang, L.F.; Martin, R.; Bieber, T. Microbiome in Healthy Skin, Update for Dermatologists. J. Eur. Acad. Dermatol. Venereol 2016, 30, 2038–2047. [Google Scholar] [CrossRef] [PubMed]
- Brzuszkiewicz, E.; Weiner, J.; Wollherr, A.; Thürmer, A.; Hüpeden, J.; Lomholt, H.B.; Kilian, M.; Gottschalk, G.; Daniel, R.; Mollenkopf, H.-J.; et al. Comparative Genomics and Transcriptomics of Propionibacterium Acnes. PLoS ONE 2011, 6, e21581. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, S.; Iwamoto, T.; Takada, K.; Okuda, K.-I.; Tajima, A.; Iwase, T.; Mizunoe, Y. Staphylococcus Epidermidis Esp Degrades Specific Proteins Associated with Staphylococcus Aureus Biofilm Formation and Host-Pathogen Interaction. J. Bacteriol. 2013, 195, 1645–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zipperer, A.; Konnerth, M.C.; Laux, C.; Berscheid, A.; Janek, D.; Weidenmaier, C.; Burian, M.; Schilling, N.A.; Slavetinsky, C.; Marschal, M.; et al. Human Commensals Producing a Novel Antibiotic Impair Pathogen Colonization. Nature 2016, 535, 511–516. [Google Scholar] [CrossRef]
- Gribbon, E.M.; Shoesmith, J.G.; Cunliffe, W.J.; Holland, K.T. The Microaerophily and Photosensitivity of Propionibacterium Acnes. J. Appl. Bacteriol. 1994, 77, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yan, R.; Zhong, Q.; Ngo, S.; Bangayan, N.J.; Nguyen, L.; Lui, T.; Liu, M.; Erfe, M.C.; Craft, N.; et al. The Diversity and Host Interactions of Propionibacterium Acnes Bacteriophages on Human Skin. ISME J. 2015, 9, 2078–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]




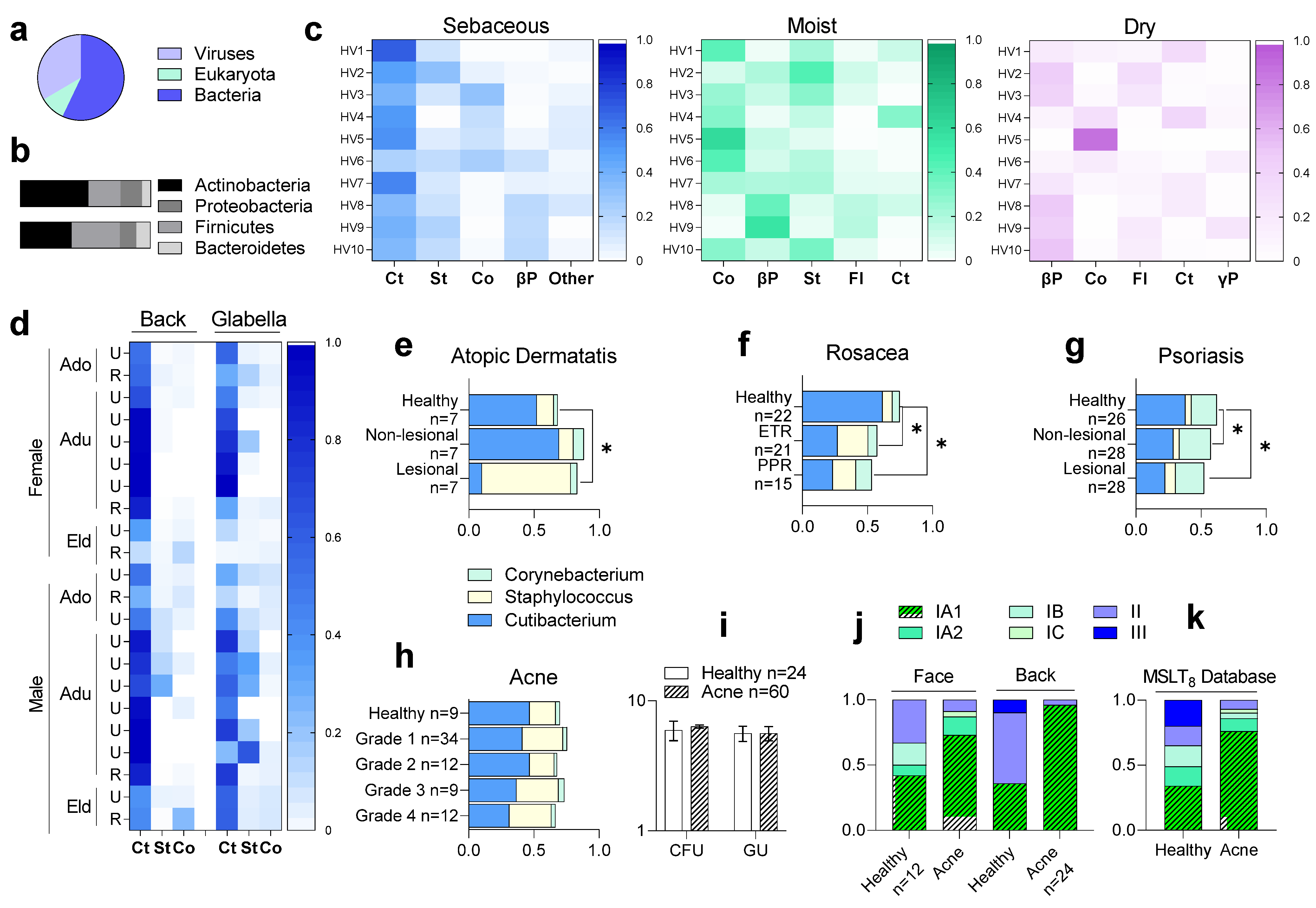{kind=link}
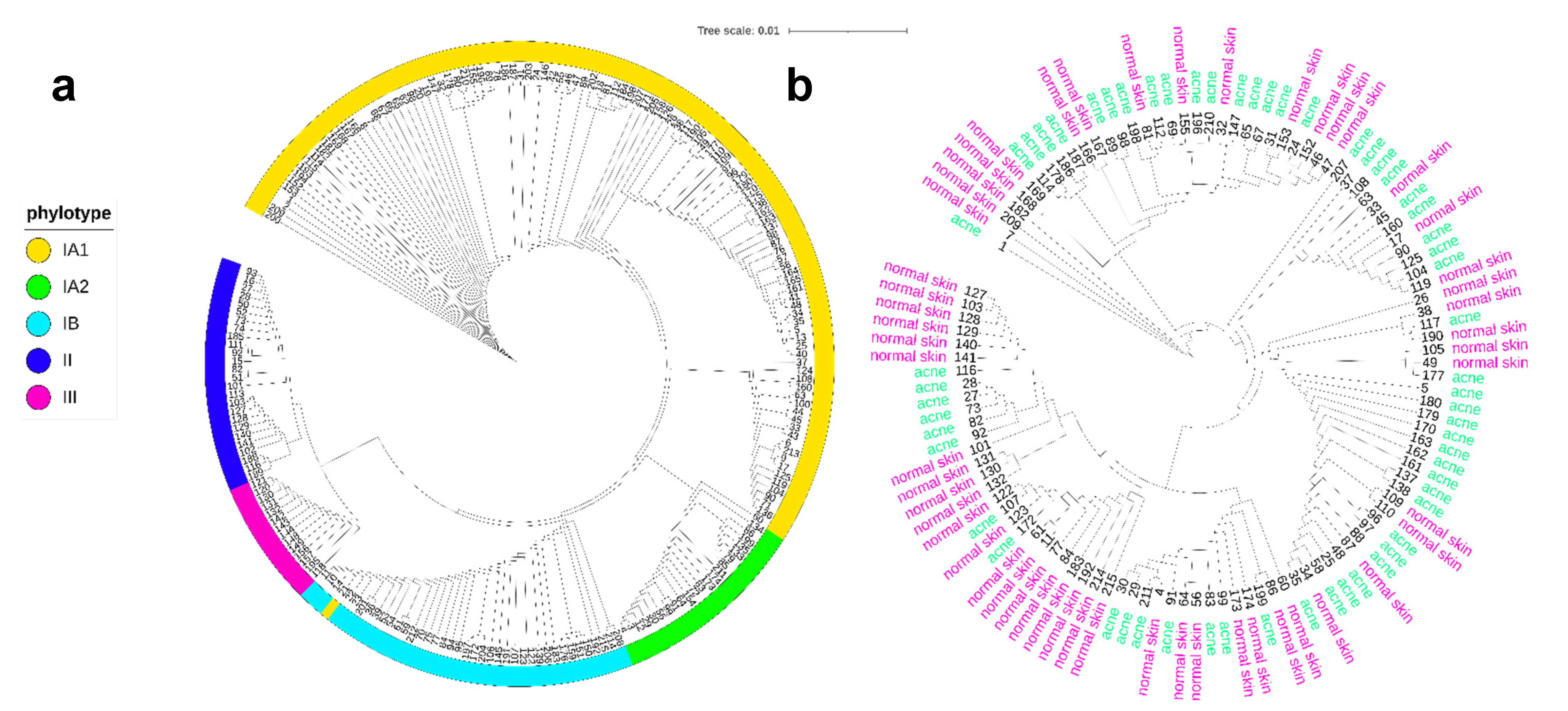{kind=link}
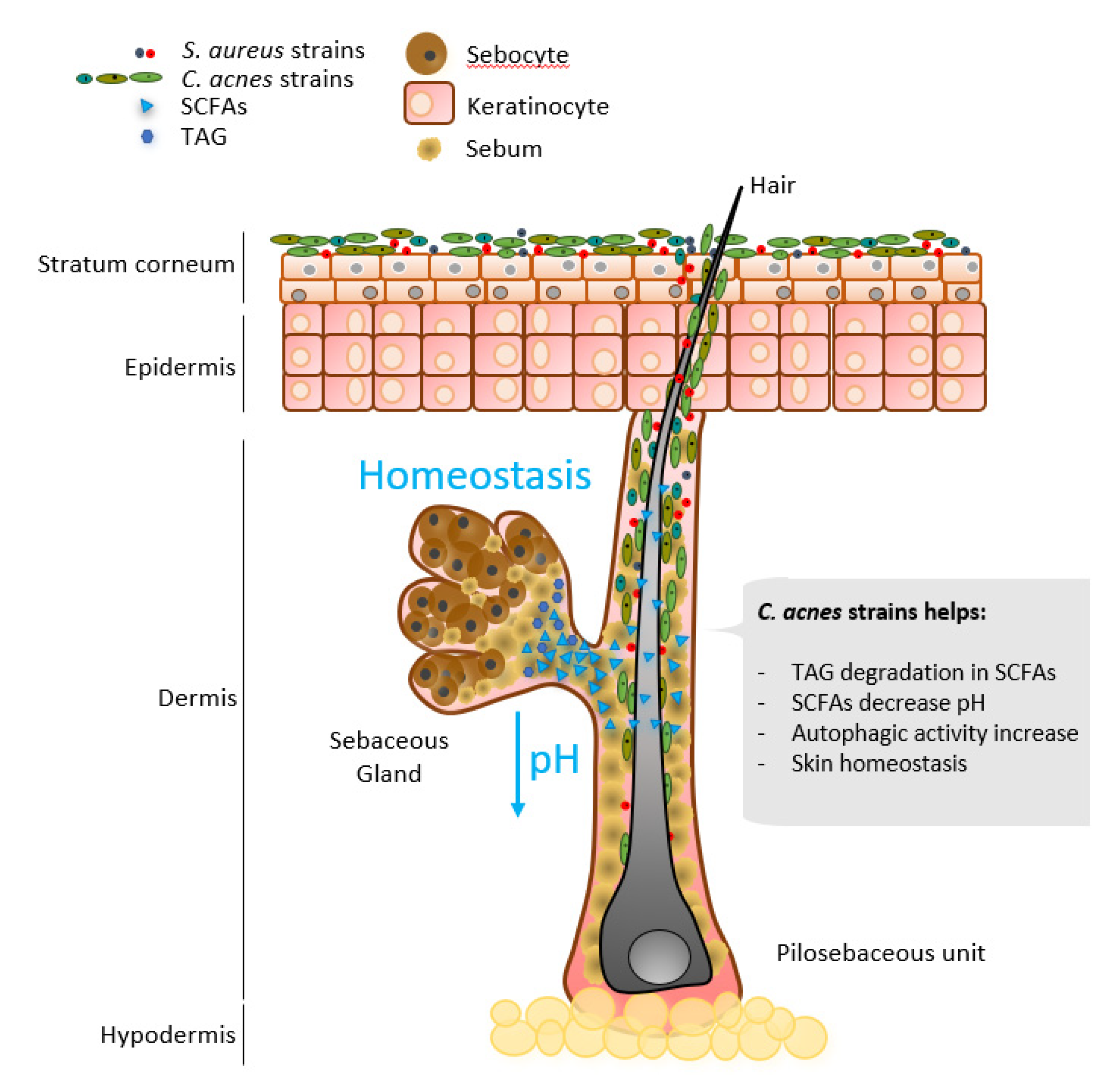{kind=link}
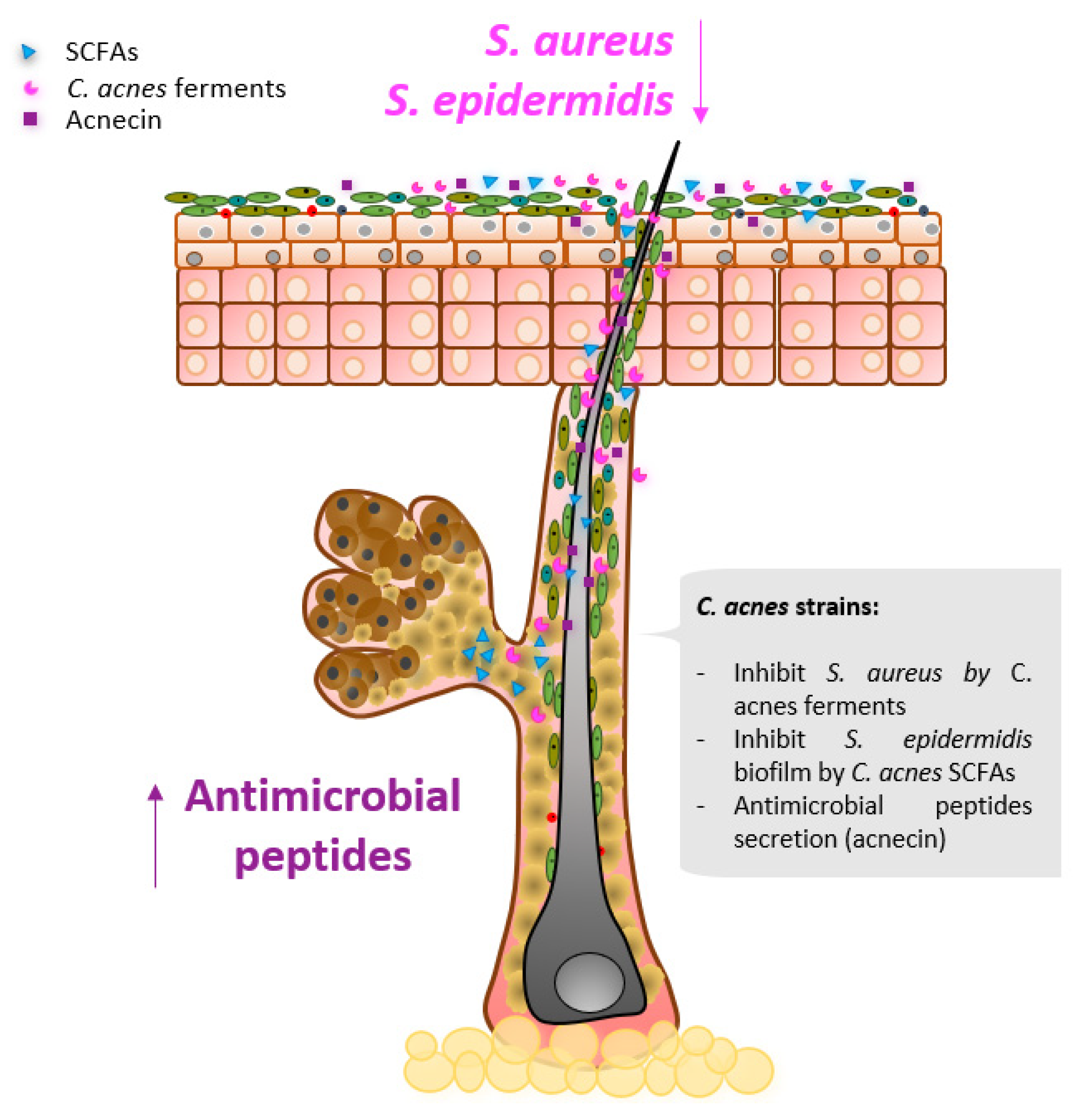{kind=link}
{kind=link}
| Safety | Statement |
| Human or animal origin | The novel genus Cutibacterium contains the human cutaneous species formerly known as Propionibacterium acnes, Propionibacterium avidum, Propionibacterium granulosum [21]. |
| Isolated from the skin of healthy individuals | C. acnes is a prevalent bacterial commensals on healthy skin [19]. Studies investigating probiotic C. acnes strains used isolates from the skin of healthy individuals [135,141]. |
| History of safe use | C. acnes has no long history of use yet, see “no adverse effects” for safety indication [135]. |
| Precise diagnostic identification (phenotype and genotype traits) | SLST analysis allows the strain-level identification of C. acnes species [142]. |
| Absence of data regarding an association with infective disease | Strains of C. acnes have been reported as opportunistic pathogens. Virulence is related to certain phylotypes or strains (e.g., phylotype IA has been associated with acne [143]). |
| No adverse effects | No adverse events were reported in two studies applying C. acnes probiotics [135,141]. |
| Absence of genes responsible for antibiotic resistance localized in non-stable elements | No detectable differences in the MIC of 21 antibiotics were observed between parent strains and their plasmid-cured derivatives for C. acnes [144]. |
| Functionality | Statement |
| Competitiveness with respect to the microbiota inhabiting the ecosystem of the skin | Antagonistic activity between S. epidermis and C. acnes strains [145]. |
| Ability to survive and maintain the metabolic activity and to grow in the target site | Applied probiotic strains of C. acnes could be detected well beyond application days, indicating their ability to survive in the target site [141]. |
| Resistance to skin salts and enzymes | C. acnes is adapted to lipid-rich anaerobic environment of sebaceous follicles [146]. |
| Resistance to low pH of the skin | C. acnes can compensate for the low pH by utilizing arginine deiminase [147]. |
| Competitiveness with respect to microbial species inhabiting the ecosystem of the skin (including closely related species) | C. acnes inhibits biofilm formation of S. epidermidis [83]. C. acnes produces an antibiotic that targets specific C. acnes strains [85]. |
| Antagonistic activity towards pathogens (e.g., S. aureus) | C. acnes produces an antibiotic against S. aureus and C. acnes ferments have been shown to inhibit methicillin-resistant S. aureus [82,127]. S. epidermis degrades proteins associated with S. aureus biofilm formation [148]. S. lugdunensis inhibits growth of S. aureus through production of AMP lugdunin [149]. |
| Resistance to bacteriocins and acids produced by the endogenic skin microbiota | C. acnes and S. epidermidis coexist in the skin as stable heterogeneous communities of strains [19]. |
| Adherence and ability to colonize some particular sites within the host organism, and an appropriate survival rate on the skin | Different body sites are shown to be colonized by different multi-phyletic communities of C. acnes [15]. |
| Technological feasibility | Statement |
| Easy production of large amounts of biomass and high productivity of cultures | Steady-state continuous culture of Propionibacterium acnes was achieved [150]. |
| Viability and stability of the desired properties of probiotic bacteria during the fixing process (freezing, freeze-drying), preparation, and distribution of probiotic products | Not yet established. |
| High storage survival rate in finished products (in aerobic and micro-aerophilic conditions) | C. acnes probiotic solution was stable for at least 1.5 months at room temperature [141] |
| Guarantee of desired sensory properties of finished products (in the case of the cosmetics industry) | Not yet established |
| Genetic stability | The sequenced strain exhibited 100% identity on the 16S ribosomal RNA to several isolated C. acnes [73]. |
| Resistance to bacteriophages | C. acnes phylotypes show selective resistance or sensitivity to bacteriophages [151]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rozas, M.; Hart de Ruijter, A.; Fabrega, M.J.; Zorgani, A.; Guell, M.; Paetzold, B.; Brillet, F. From Dysbiosis to Healthy Skin: Major Contributions of Cutibacterium acnes to Skin Homeostasis. Microorganisms 2021, 9, 628. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030628
Rozas M, Hart de Ruijter A, Fabrega MJ, Zorgani A, Guell M, Paetzold B, Brillet F. From Dysbiosis to Healthy Skin: Major Contributions of Cutibacterium acnes to Skin Homeostasis. Microorganisms. 2021; 9(3):628. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030628
Chicago/Turabian StyleRozas, Miquel, Astrid Hart de Ruijter, Maria Jose Fabrega, Amine Zorgani, Marc Guell, Bernhard Paetzold, and Francois Brillet. 2021. "From Dysbiosis to Healthy Skin: Major Contributions of Cutibacterium acnes to Skin Homeostasis" Microorganisms 9, no. 3: 628. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030628