
Biocontrol of Soft Rot Caused by Pectobacterium odoriferum with Bacteriophage phiPccP-1 in Kimchi Cabbage

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain Identification and Growth Conditions
2.2. Bacteriophage Purification and Propagation
2.3. Transmission Electron Microscopy
2.4. Determination of Host Range of phiPccP-1
2.5. Determination of Killing Curves of phiPccP-1
2.6. The Frequency Measurement of Bacteriophage-Insensitive Mutant (BIM) Development
2.7. Thermal, pH, and UV Radiation Stability of phiPccP-1
2.8. Genome Analysis of phiPccP-1 Genomic DNA
2.9. Control Efficacy of phiPccP-1 in the Detached Mature Leaves of Kimchi Cabbage
2.10. The Prophylactic and Therapeutic Effects of phiPccP-1 in Kimchi Cabbage Seedlings
2.11. Statistical Analysis
3. Results
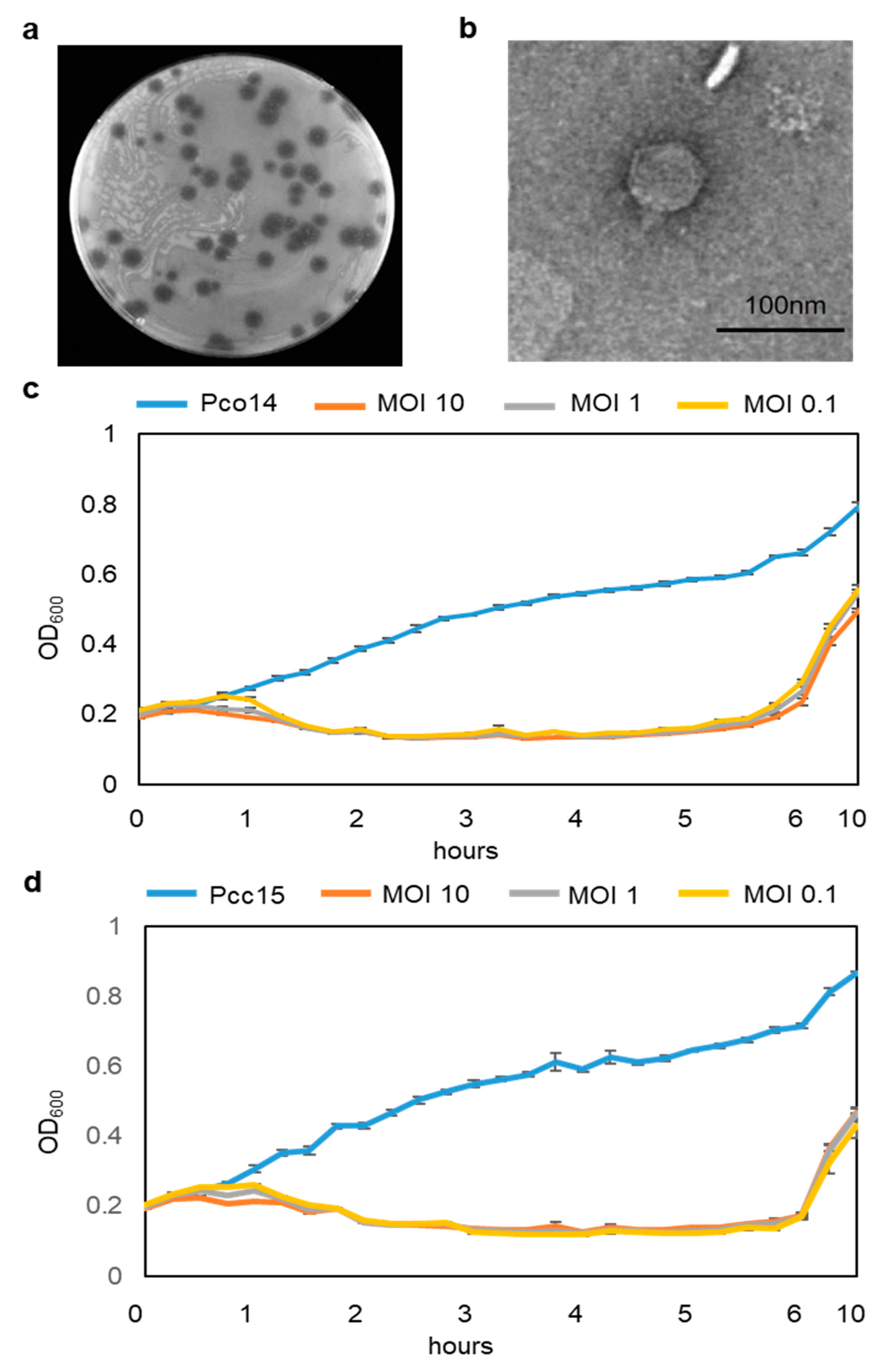
3.1. Isolation and Classification of phiPccP-1
3.2. Determination of phiPccP-1 Host Range and its Killing Curve
3.3. The Frequency of BIM Development
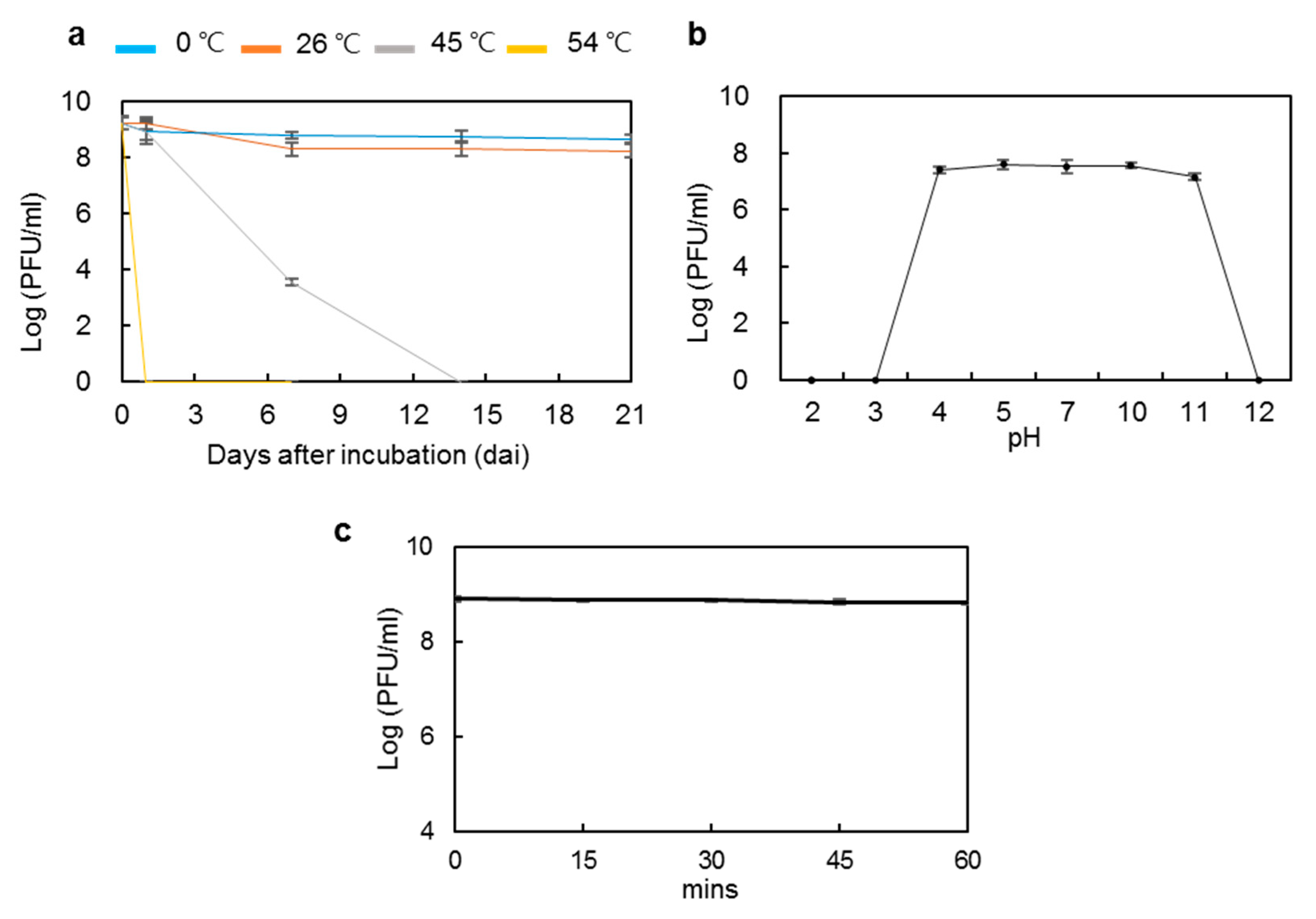
3.4. Stability of phiPccP-1 under Diverse Temperature, pH, and UV Irradiation
3.5. Analysis of phiPccP-1 Genome
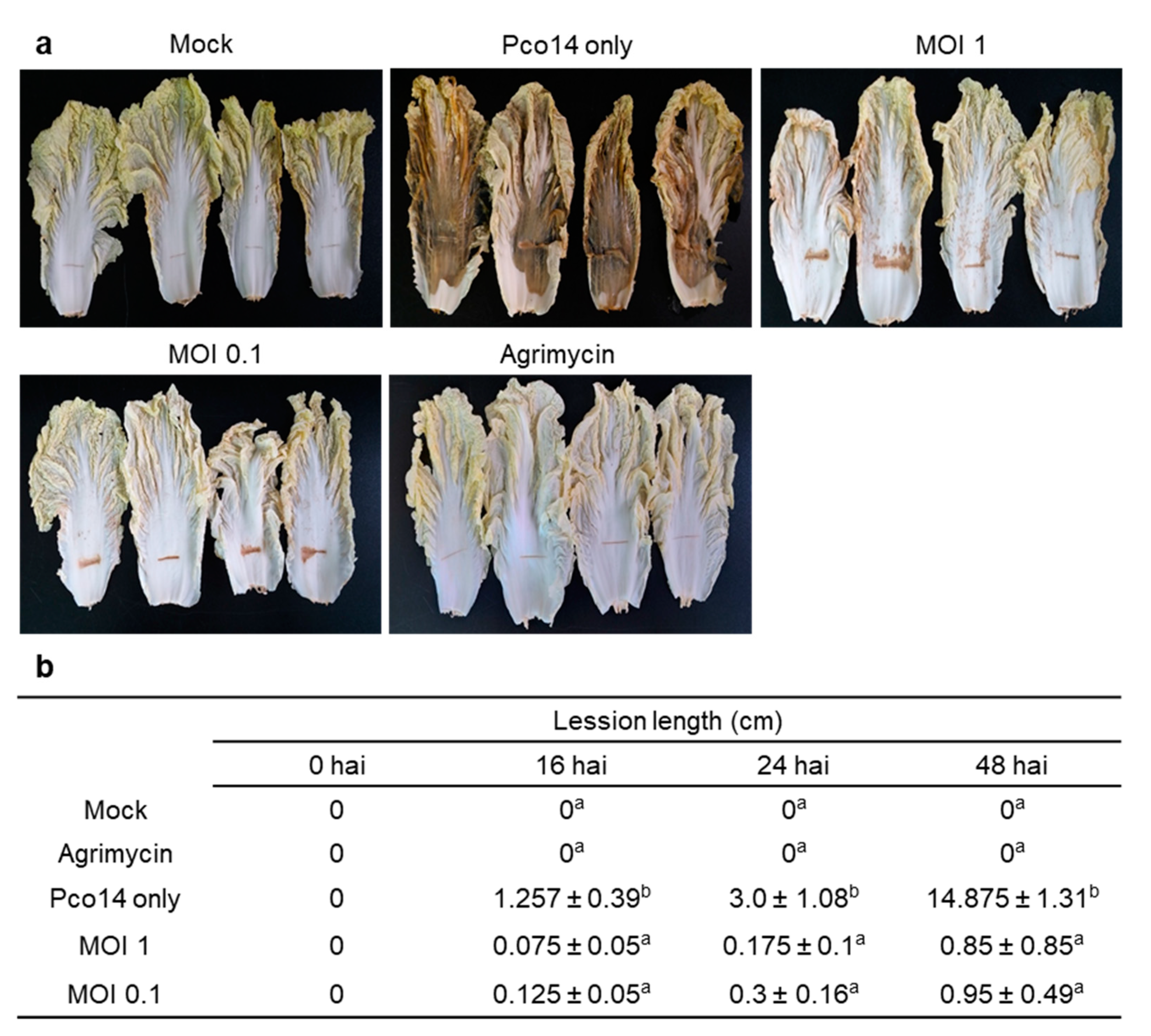
3.6. Control Efficacy of phiPccP-1 Against Soft-Rot Disease in the Detached Mature Leaves of Kimchi Cabbage
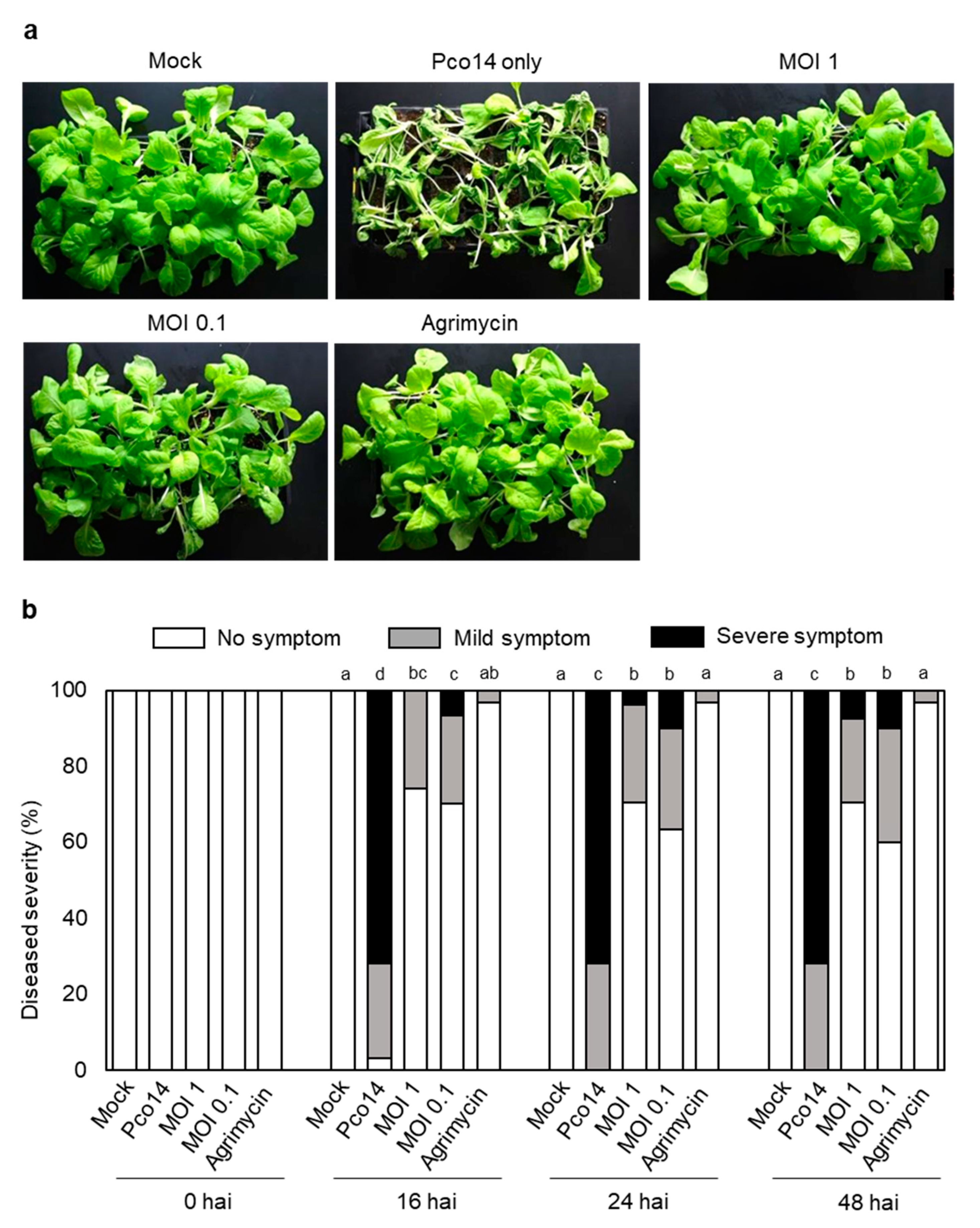
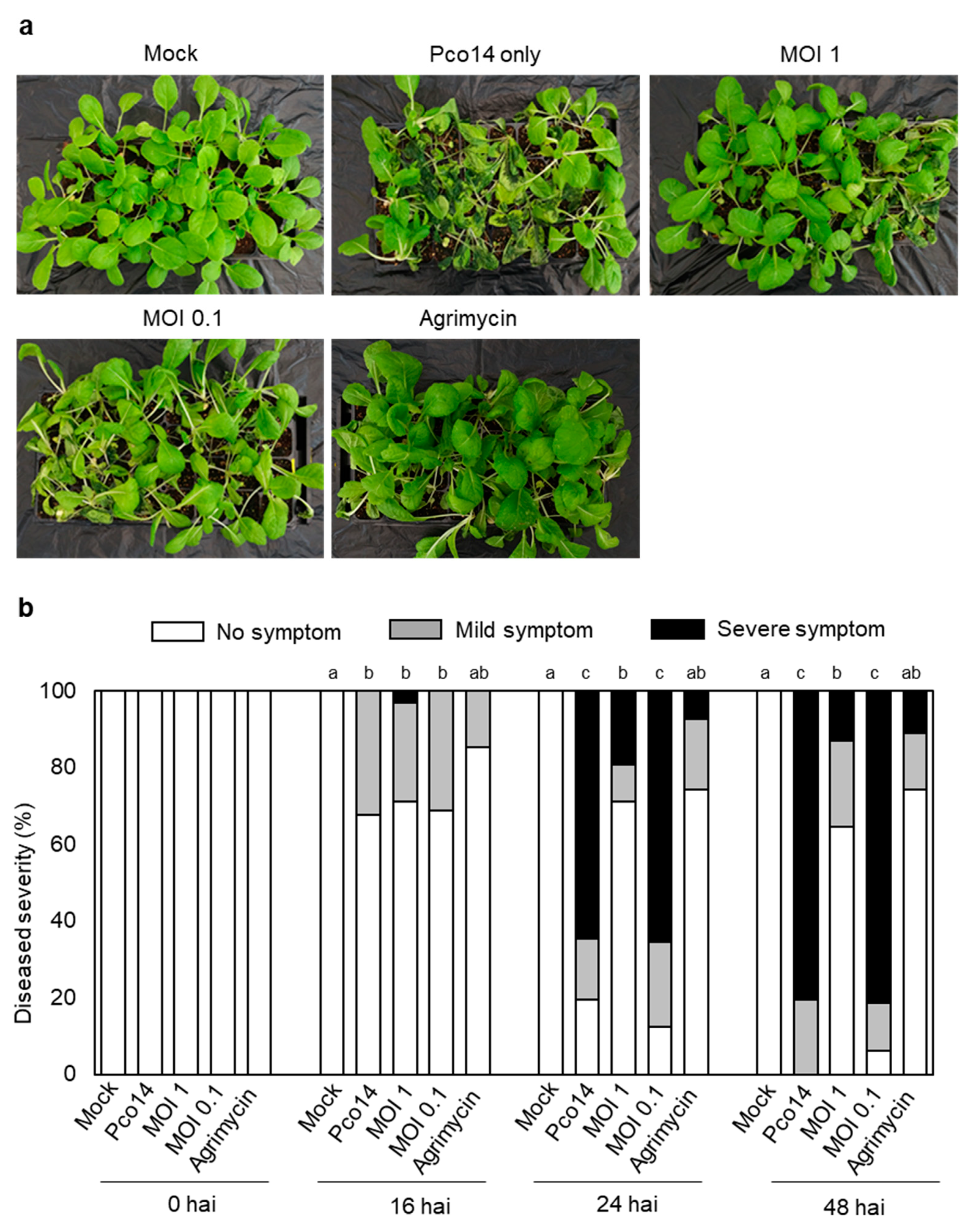
3.7. Control Efficacy of phiPccP-1 against Soft-Rot Disease in Kimchi Cabbage Seedlings
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Charkowski, A.O. The changing face of bacterial soft-rot diseases. Annu. Rev. Phytopathol. 2018, 56, 269–288. [Google Scholar] [CrossRef]
- Gardan, L.; Gouy, C.; Christen, R.; Samson, R. Elevation of three subspecies of Pectobacterium carotovorum to species level: Pectobacterium atrosepticum sp. nov., Pectobacterium betavasculorum sp. nov. and Pectobacterium wasabiae sp. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 381–391. [Google Scholar] [CrossRef]
- Nabhan, S.; Boer, S.; Maiss, E.; Wydra, K. Taxonomic relatedness between Pectobacterium carotovorum subsp. carotovorum, Pectobacterium carotovorum subsp. odoriferum and Pectobacterium carotovorum subsp. brasiliense subsp. nov. J. Appl. Microbiol. 2012, 113, 904–913. [Google Scholar] [CrossRef] [PubMed]
- Nabhan, S.; De Boer, S.H.; Maiss, E.; Wydra, K. Pectobacterium aroidearum sp. nov., a soft rot pathogen with preference for monocotyledonous plants. Int. J. Syst. Evol. Microbiol. 2013, 63, 2520–2525. [Google Scholar] [CrossRef] [Green Version]
- Waleron, M.; Misztak, A.; Waleron, M.; Franczuk, M.; Wielgomas, B.; Waleron, K. Transfer of Pectobacterium carotovorum subsp. carotovorum strains isolated from potatoes grown at high altitudes to Pectobacterium peruviense sp. nov. Syst. Appl. Microbiol. 2018, 41, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Dees, M.W.; Lysøe, E.; Rossmann, S.; Perminow, J.; Brurberg, M.B. Pectobacterium polaris sp. nov., isolated from potato (Solanum tuberosum). Int. J. Syst. Evol. Microbiol. 2017, 67, 5222–5229. [Google Scholar] [CrossRef]
- Duarte, V.; De Boer, S.; Ward, L.; De Oliveira, A.M.R. Characterization of atypical Erwinia carotovora strains causing blackleg of potato in Brazil. J. Appl. Microbiol. 2004, 96, 535–545. [Google Scholar] [CrossRef]
- Khayi, S.; Cigna, J.; Chong, T.M.; Quêtu-Laurent, A.; Chan, K.-G.; Hélias, V.; Faure, D. Transfer of the potato plant isolates of Pectobacterium wasabiae to Pectobacterium parmentieri sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5379–5383. [Google Scholar] [CrossRef]
- Koh, Y.; Kim, G.; Lee, Y.; Sohn, S.; Koh, H.; Kwon, S.; Heu, S.; Jung, J. Pectobacterium carotovorum subsp. Actinidiae subsp. nov., a new bacterial pathogen causing canker-like symptoms in yellow kiwifruit, Actinidia chinensis. N. Z. J. Crop. Hortic. Sci. 2012, 40, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Oulghazi, S.; Cigna, J.; Lau, Y.Y.; Moumni, M.; Chan, K.G.; Faure, D. Transfer of the waterfall source isolate Pectobacterium carotovorum M022 to Pectobacterium fontis sp. nov., a deep-branching species within the genus Pectobacterium. Int. J. Syst. Evol. Microbiol. 2019, 69, 470–475. [Google Scholar] [CrossRef]
- Jee, S.; Choi, J.-G.; Lee, Y.-G.; Kwon, M.; Hwang, I.; Heu, S. Distribution of Pectobacterium Species Isolated in South Korea and Comparison of Temperature Effects on Pathogenicity. Plant Pathol. J. 2020, 36, 346–354. [Google Scholar] [CrossRef]
- Pédron, J.; Bertrand, C.; Taghouti, G.; Portier, P.; Barny, M.-A. Pectobacterium aquaticum sp. nov., isolated from waterways. Int. J. Syst. Evol. Microbiol. 2019, 69, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Portier, P.; Pédron, J.; Taghouti, G.; Saux, M.F.-L.; Caullireau, E.; Bertrand, C.; Laurent, A.; Chawki, K.; Oulgazi, S.; Moumni, M.; et al. Elevation of Pectobacterium carotovorum subsp. odoriferum to species level as Pectobacterium odoriferum sp. nov., proposal of Pectobacterium brasiliense sp. nov. and Pectobacterium actinidiae sp. nov., emended description of Pectobacterium carotovorum and description of Pectobacterium versatile sp. nov., isolated from streams and symptoms on diverse plants. Int. J. Syst. Evol. Microbiol. 2019, 69, 3207–3216. [Google Scholar] [CrossRef]
- Sarfraz, S.; Riaz, K.; Oulghazi, S.; Cigna, J.; Sahi, S.T.; Khan, S.H.; Faure, D. Pectobacterium punjabense sp. nov., isolated from blackleg symptoms of potato plants in Pakistan. Int. J. Syst. Evol. Microbiol. 2018, 68, 3551–3556. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toth, I.K.; Bell, K.S.; Holeva, M.C.; Birch, P.R.J. Soft rot erwiniae: From genes to genomes. Mol. Plant Pathol. 2002, 4, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Petzoldt, R.; Dickson, M.H. Screening and identification of resistance to bacterial soft rot in Brassica rapa. Euphytica 2001, 118, 271–280. [Google Scholar] [CrossRef]
- Bhat, K.; Masood, S.; Bhat, N.; Bhat, M.A.; Razvi, S.; Mir, M.; Akhtar, S.; Wani, N.; Habib, M. Current Status of Post Harvest Soft Rot in Vegetables: A Review. Asian J. Plant Sci. 2010, 9, 200–208. [Google Scholar] [CrossRef] [Green Version]
- Gallois, A.; Samson, R.; Ageron, E.; Grimont, P.A.D. Erwinia carotovora subsp. odorifera subsp. nov., Associated with Odorous Soft Rot of Chicory (Cichorium intybus L.). Int. J. Syst. Bacteriol. 1992, 42, 582–588. [Google Scholar] [CrossRef]
- Lan, W.W.; Nishiwaki, Y.; Akino, S.; Kondo, N. Soft rot of root chicory in Hokkaido and its causal bacteria. J. Gen. Plant Pathol. 2013, 79, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Waleron, M.; Lojkowska, E. Characterization of Pectobacterium carotovorum subsp. odoriferum causing soft rot of stored vegetables. Eur. J. Plant Pathol. 2014, 139, 457–469. [Google Scholar] [CrossRef] [Green Version]
- Oskiera, M.; Kałużna, M.; Kowalska, B.; Smolinska, U. Pectobacterium carotovorum subsp. odoriferum on cabbage and Chinese cabbage: Identification, characterization and taxonomic relatedness of bacterial soft rot causal agents. J. Plant Pathol. 2017, 99, 149–160. [Google Scholar]
- Li, X.; Fu, L.; Chen, C.; Sun, W.; Tian, Y.; Xie, H. Characteristics and Rapid Diagnosis of Pectobacterium carotovorum ssp. Associated with Bacterial Soft Rot of Vegetables in China. Plant Dis. 2020, 104, 1158–1166. [Google Scholar] [CrossRef]
- Kumar, K.; Gupta, S.C.; Chander, Y.; Singh, A.K. Antibiotic Use in Agriculture and Its Impact on the Terrestrial Environment. Adv. Agron. 2005, 87, 1–54. [Google Scholar] [CrossRef]
- Durso, L.M.; Cook, K.L. Impacts of antibiotic use in agriculture: What are the benefits and risks? Curr. Opin. Microbiol. 2014, 19, 37–44. [Google Scholar] [CrossRef]
- Agrios, G. Plant Pathology, 5th ed.; Elsevier: Burlington, MA, USA, 1964; p. 952. [Google Scholar]
- Roh, E.; Park, T.-H.; Kim, M.-I.; Lee, S.; Ryu, S.; Oh, C.-S.; Rhee, S.; Kim, D.-H.; Park, B.-S.; Heu, S. Characterization of a New Bacteriocin, Carocin D, from Pectobacterium carotovorum subsp. carotovorum Pcc21. Appl. Environ. Microbiol. 2010, 76, 7541–7549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.-W.; Derilo, R.C.; Lagitnay, R.B.J.S.; Wu, H.-P.; Chen, K.-I.; Chuang, D.-Y. Identification and characterization of the bacteriocin Carocin S3 from the multiple bacteriocin producing strain of Pectobacterium carotovorum subsp. carotovorum. BMC Microbiol. 2020, 20, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hassan, E.A.; Bagy, H.M.K.; Bashandy, S.R. Efficacy of potent antagonistic yeast Wickerhamiella versatilis against soft rot disease of potato caused by Pectobacterium carotovorum subsp. carotovorum. Arch. Phytopathol. Plant Prot. 2019, 52, 1125–1148. [Google Scholar] [CrossRef]
- Krzyzanowska, D.M.; Maciag, T.; Siwinska, J.; Krychowiak, M.; Jafra, S.; Czajkowski, R. Compatible Mixture of Bacterial Antagonists Developed to Protect Potato Tubers from Soft Rot Caused by Pectobacterium spp. and Dickeya spp. Plant Dis. 2019, 103, 1374–1382. [Google Scholar] [CrossRef] [PubMed]
- Vu, N.T.; Oh, C.-S. Bacteriophage Usage for Bacterial Disease Management and Diagnosis in Plants. Plant Pathol. J. 2020, 36, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.H.; Lee, J.-H.; Shin, H.; Ji, S.; Roh, E.; Jung, K.; Ryu, S.; Choi, J.; Heu, S. Complete Genome Sequence of Pectobacterium carotovorum subsp. carotovorum Bacteriophage My1. J. Virol. 2012, 86, 11410–11411. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.-A.; Jee, S.; Lee, D.H.; Roh, E.; Jung, K.; Oh, C.; Heu, S. Biocontrol of Pectobacterium carotovorum subsp. carotovorum using bacteriophage PP1. J. Microbiol. Biotechnol. 2013, 23, 1147–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.-A.; Lee, N.H.; Heu, S. Isolation and Genomic Characterization of the T4-Like Bacteriophage PM2 Infecting Pectobacterium carotovorum subsp. carotovorum. Plant Pathol. J. 2015, 31, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Hirata, H.; Kashihara, M.; Horiike, T.; Suzuki, T.; Dohra, H.; Netsu, O.; Tsuyumu, S. Genome Sequence of Pectobacterium carotovorum Phage PPWS1, Isolated from Japanese Horseradish [Eutrema japonicum (Miq.) Koidz] Showing Soft-Rot Symptoms. Genome Announc. 2016, 4, e01625-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.-A.; Heu, S.; Park, J.; Roh, E. Genomic characterization of bacteriophage vB_PcaP_PP2 infecting Pectobacterium carotovorum subsp. carotovorum, a new member of a proposed genus in the subfamily Autographivirinae. Arch. Virol. 2017, 162, 2441–2444. [Google Scholar] [CrossRef] [Green Version]
- Voronina, M.V.; Bugaeva, E.N.; Vasiliev, D.M.; Kabanova, A.P.; Barannik, A.P.; Shneider, M.M.; Kulikov, E.E.; Korzhenkov, A.A.; Toschakov, S.V.; Ignatov, A.N.; et al. Characterization of Pectobacterium carotovorum subsp. carotovorum Bacteriophage PP16 Prospective for Biocontrol of Potato Soft Rot. Microbiology 2019, 88, 451–460. [Google Scholar] [CrossRef]
- Kim, H.; Kim, M.; Bai, J.; Lim, J.-A.; Heu, S.; Ryu, S. Colanic Acid Is a Novel Phage Receptor of Pectobacterium carotovorum subsp. carotovorum Phage POP72. Front. Microbiol. 2019, 10, 143. [Google Scholar] [CrossRef] [PubMed]
- Heuer, H.; Krsek, M.; Baker, P.; Smalla, K.; Wellington, E.M. Analysis of actinomycete communities by specific amplification of genes encoding 16S rRNA and gel-electrophoretic separation in denaturing gradients. Appl. Environ. Microbiol. 1997, 63, 3233–3241. [Google Scholar] [CrossRef] [Green Version]
- Kropinski, A.M.; Mazzocco, A.; Waddell, T.E.; Lingohr, E.; Johnson, R.P. Enumeration of Bacteriophages by Double Agar Overlay Plaque Assay. In Bacteriophages; Springer: Berlin/Heidelberg, Germany, 2009; pp. 69–76. [Google Scholar]
- Yamamoto, K.R.; Alberts, B.M.; Benzinger, R.; Lawhorne, L.; Treiber, G. Rapid bacteriophage sedimentation in the presence of polyethylene glycol and its application to large-scale virus purification. Virology 1970, 40, 734–744. [Google Scholar] [CrossRef]
- King, A.M.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: San Diego, CA, USA, 2011; Volume 9. [Google Scholar]
- Frampton, R.A.; Taylor, C.; Moreno, A.V.H.; Visnovsky, S.B.; Petty, N.K.; Pitman, A.R.; Fineran, P.C. Identification of Bacteriophages for Biocontrol of the Kiwifruit Canker Phytopathogen Pseudomonas syringae pv. actinidiae. Appl. Environ. Microbiol. 2014, 80, 2216–2228. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, K.; Cazarez-Montoya, C.; Lopez-Moreno, H.S.; Campo, N.C.-D. Bacteriophage cocktail for biocontrol of Escherichia coli O157:H7: Stability and potential allergenicity study. PLoS ONE 2018, 13, e0195023. [Google Scholar] [CrossRef] [Green Version]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Jończyk, E.; Kłak, M.; Międzybrodzki, R.; Górski, A. The influence of external factors on bacteriophages—Review. Folia Microbiol. 2011, 56, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Ignoffo, C.; Garcia, C. Antioxidant and oxidative enzyme effects on the inactivation of inclusion bodies of the Heliothis baculovirus by simulated sunlight-UV. Environ. Entomol. 1994, 23, 1025–1029. [Google Scholar] [CrossRef]
- Adriaenssens, E.M.; Sullivan, M.B.; Knezevic, P.; van Zyl, L.J.; Sarkar, B.; Dutilh, B.E.; Alfenas-Zerbini, P.; Łobocka, M.; Tong, Y.; Brister, J.R. Taxonomy of prokaryotic viruses: 2018–2019 update from the ICTV Bacterial and Archaeal Viruses Subcommittee. Arch. Virol. 2020, 165, 1253–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Kim, S.; Park, T. Diversity of bacteriophages infecting Pectobacterium from potato fields. J. Plant Pathol. 2017, 99, 453–460. [Google Scholar]
- Gill, J.; Abedon, S. Bacteriophage Ecology and Plants. APSnet Featur. Artic. 2003, 1–17. [Google Scholar] [CrossRef]
- Perombelon, M.C.M.; Kelman, A. Ecology of the Soft Rot Erwinias. Annu. Rev. Phytopathol. 1980, 18, 361–387. [Google Scholar] [CrossRef]
- Elphinstone, J. Soft Rot and Blackleg of Potato: Erwinia spp; Technical Information Bulletin 21; CIP: Lima, Peru, 1987. [Google Scholar]
- Perombelon, M.C.M.; Lumb, V.M.; Zutra, D. Pathogenicity of soft rot erwinias to potato plants in Scotland and Israel. J. Appl. Bacteriol. 1987, 63, 73–84. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Vu, N.-T.; Oh, E.-J.; Rahimi-Midani, A.; Thi, T.-N.; Song, Y.-R.; Hwang, I.-S.; Choi, T.-J.; Oh, C.-S. Biocontrol of Soft Rot Caused by Pectobacterium odoriferum with Bacteriophage phiPccP-1 in Kimchi Cabbage. Microorganisms 2021, 9, 779. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040779
Lee S, Vu N-T, Oh E-J, Rahimi-Midani A, Thi T-N, Song Y-R, Hwang I-S, Choi T-J, Oh C-S. Biocontrol of Soft Rot Caused by Pectobacterium odoriferum with Bacteriophage phiPccP-1 in Kimchi Cabbage. Microorganisms. 2021; 9(4):779. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040779
Chicago/Turabian StyleLee, Soohong, Nguyen-Trung Vu, Eom-Ji Oh, Aryan Rahimi-Midani, Thuong-Nguyen Thi, Yu-Rim Song, In-Sun Hwang, Tae-Jin Choi, and Chang-Sik Oh. 2021. "Biocontrol of Soft Rot Caused by Pectobacterium odoriferum with Bacteriophage phiPccP-1 in Kimchi Cabbage" Microorganisms 9, no. 4: 779. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040779