
Interactions between Rainbow Trout Eyed Eggs and Flavobacterium spp. Using a Bath Challenge Model: Preliminary Evaluation of Bacteriophages as Pathogen Control Agents

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria
2.2. Bacteriophages
2.3. Rainbow Trout Eyed Eggs
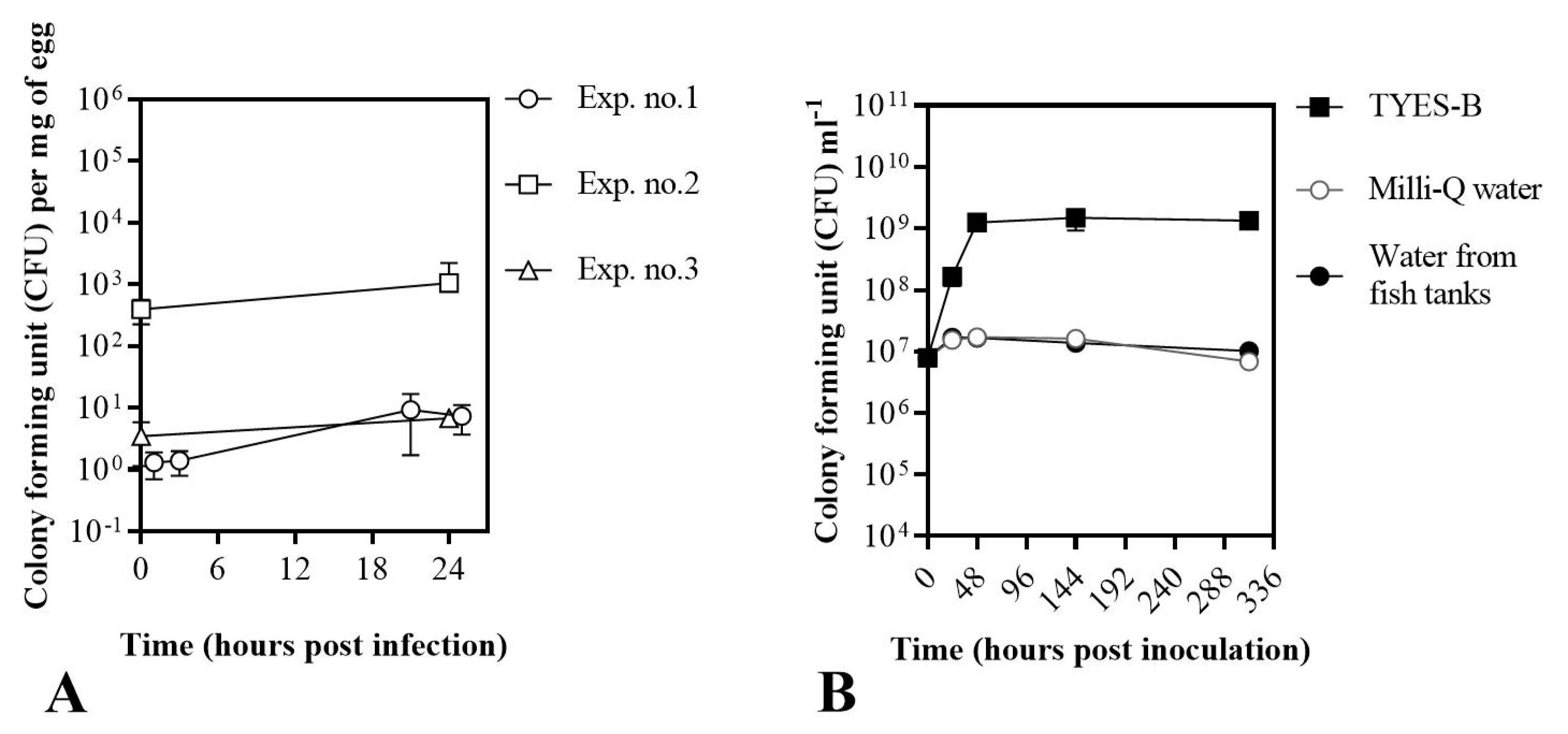
2.4. Establishment of a Bath Bacterial Challenge Method (Section A)
2.5. Interactions of Phages with Rainbow Trout Eyed Eggs (Section B)
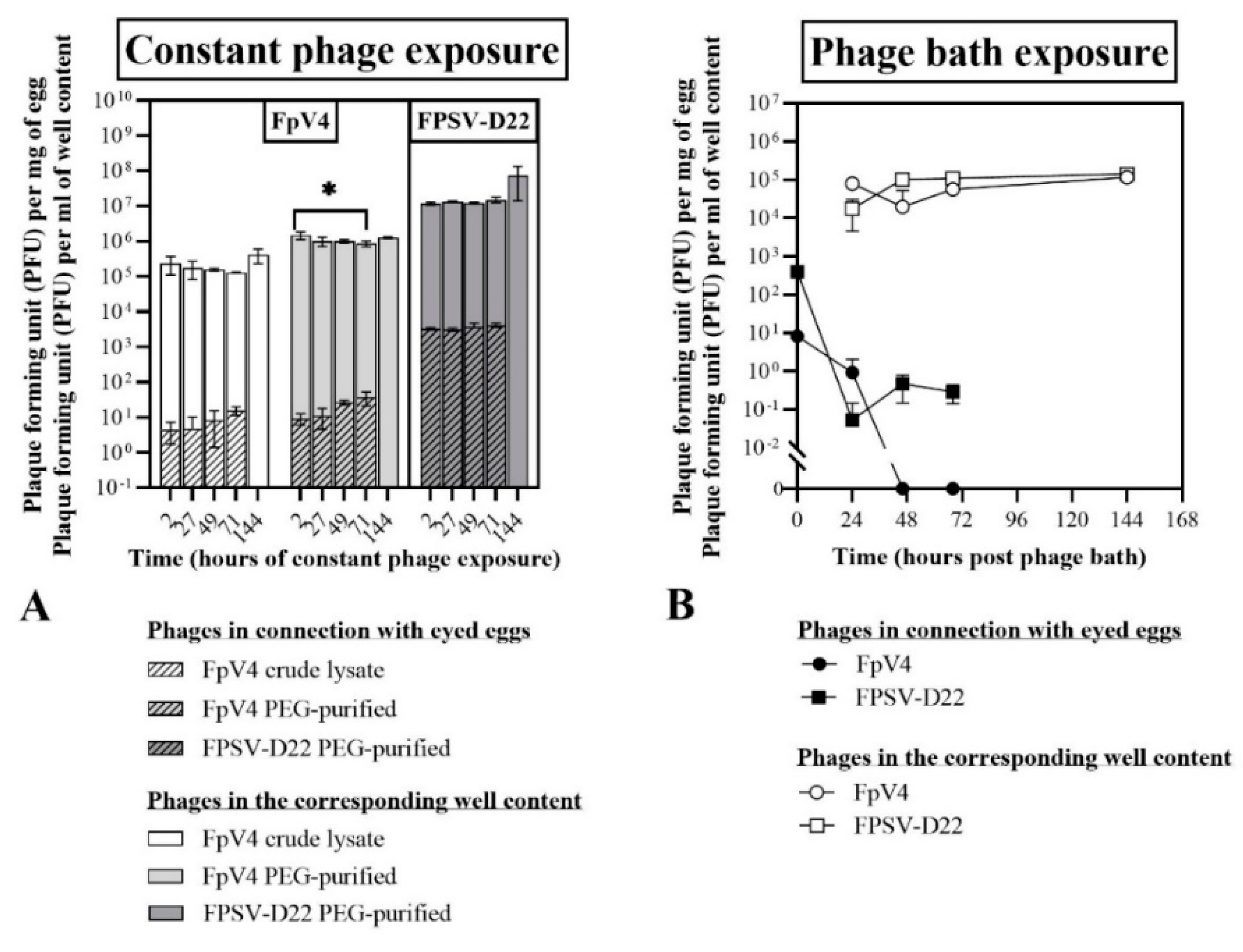
2.5.1. Constant Exposure of Eyed Eggs to F. psychrophilum Phages (Exp. I, Section B)
2.5.2. Bath Exposure of Eyed Eggs to F. psychrophilum Phages (Exp. II, Section B)
2.5.3. Bath and Constant Exposure of Eyed Eggs to F. columnare Phages (Exp. III, Section B)
2.6. Evaluation of Phages as Pathogen Control Agents (Section C)
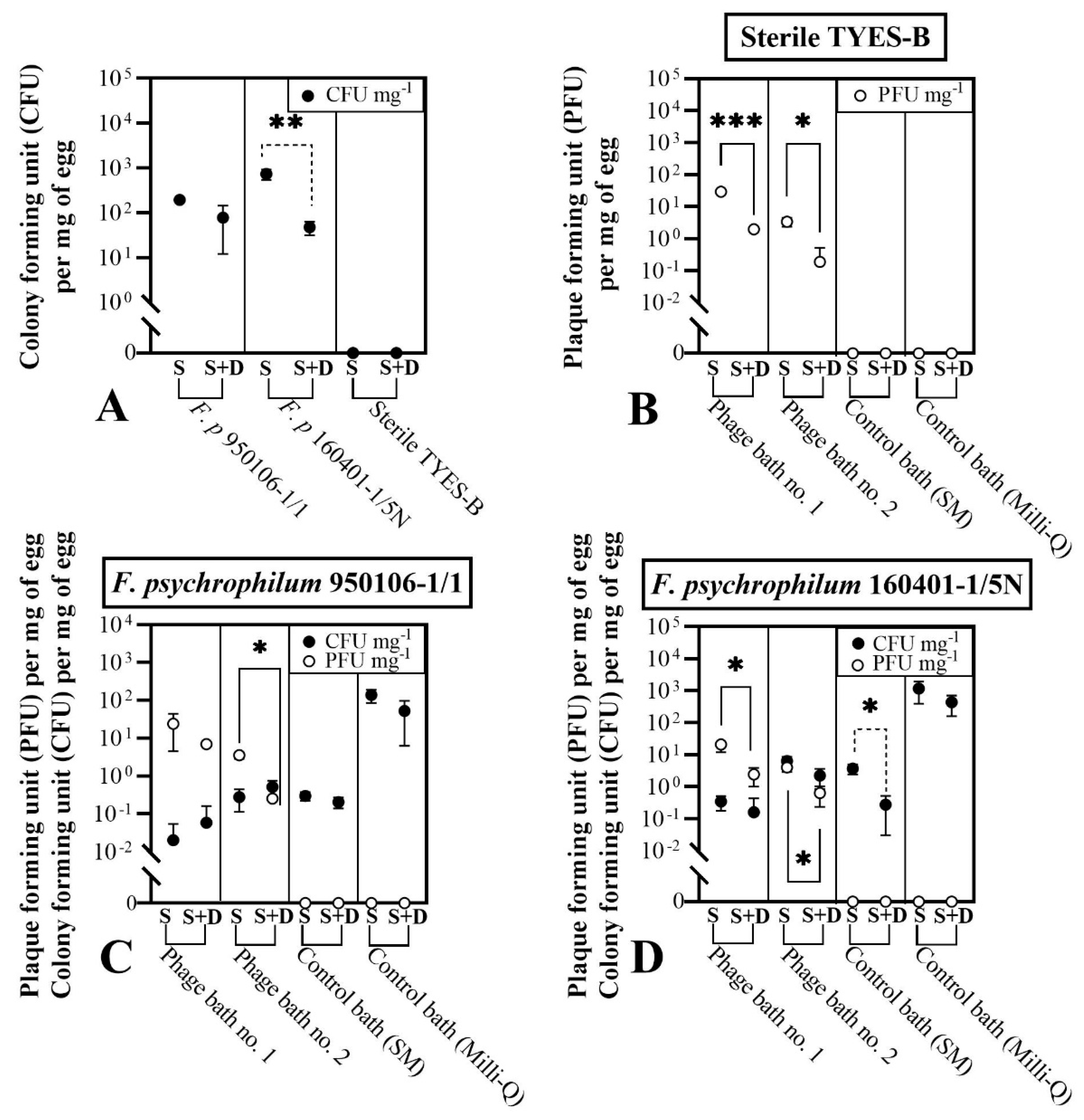
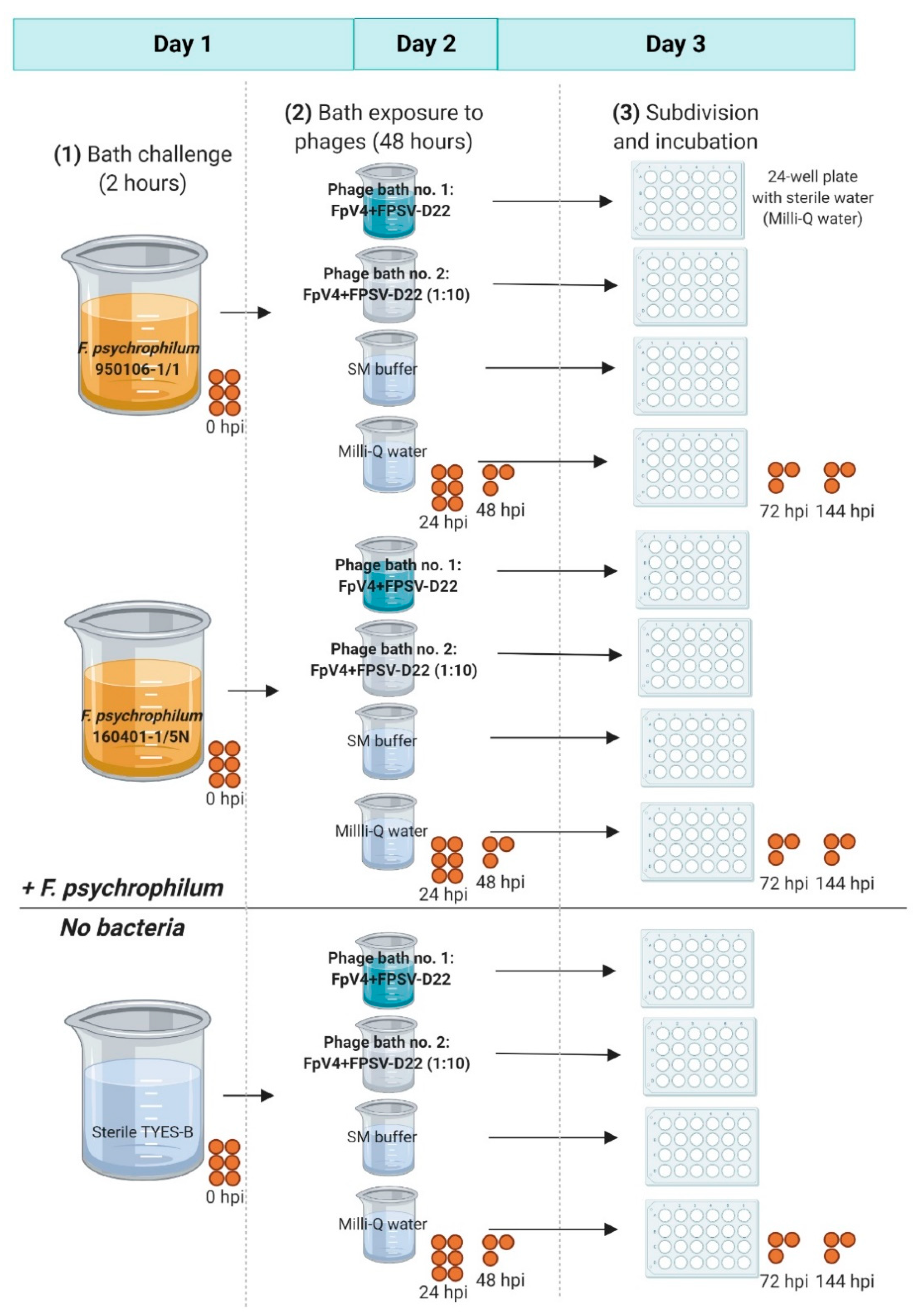
2.6.1. Phage Bath of F. psychrophilum Challenged Eggs (Exp. I, Section C)
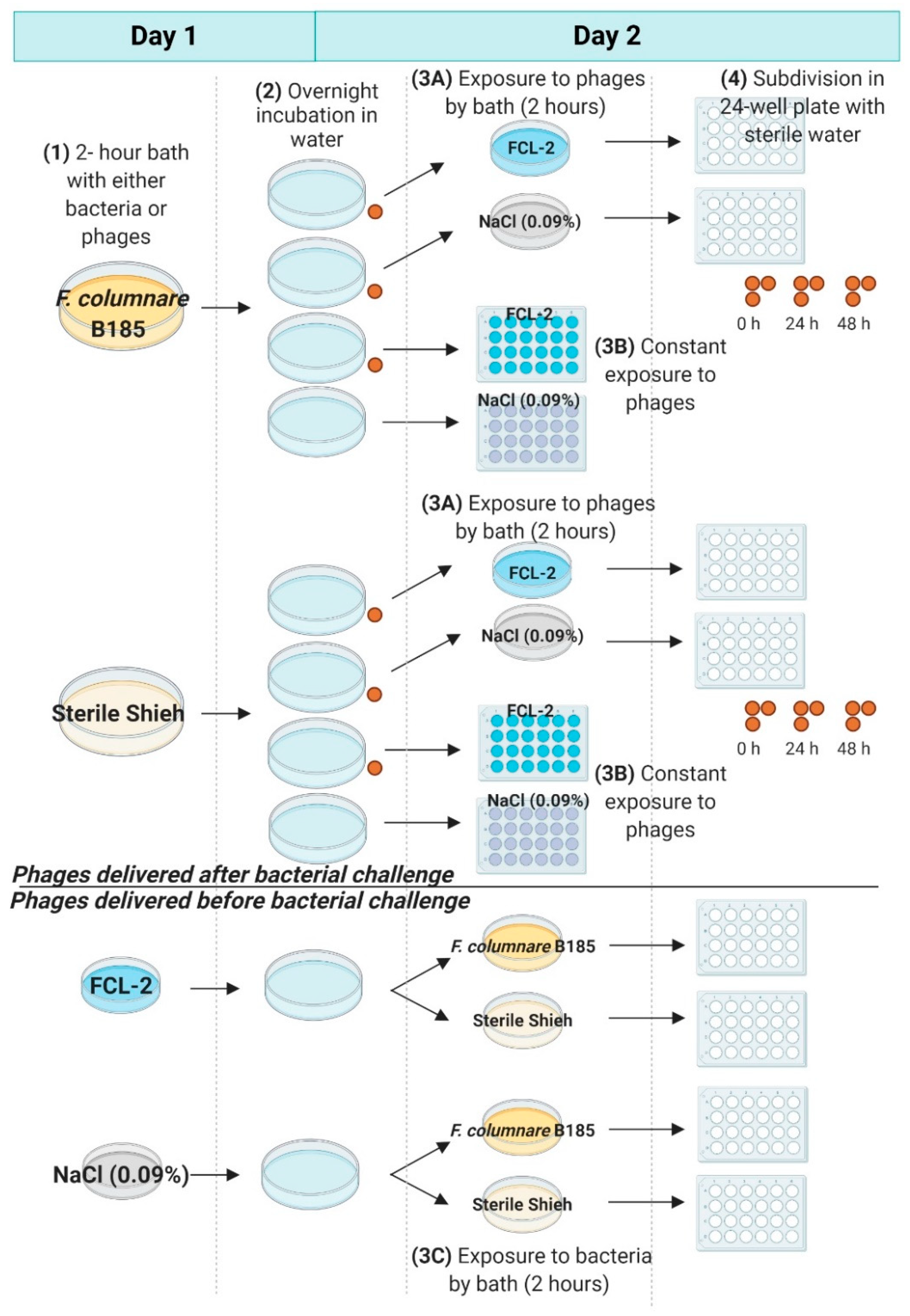
2.6.2. Phage Exposure of F. columnare Challenged Eggs (Exp. II, Section C)
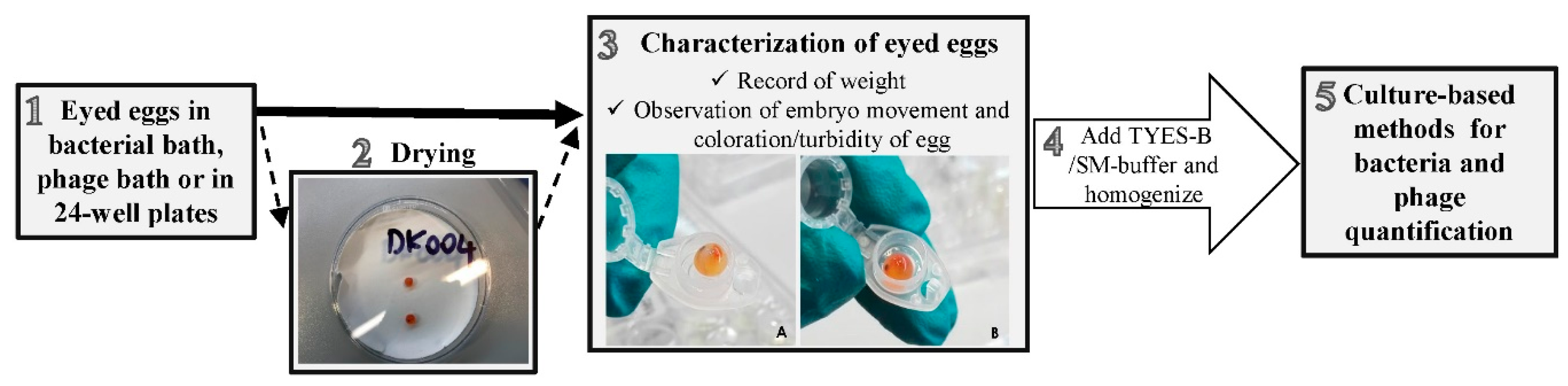
2.7. Eyed Eggs Sampling Procedure
2.8. Detection and Quantification of Bacteriophages
2.9. Statistics
3. Results
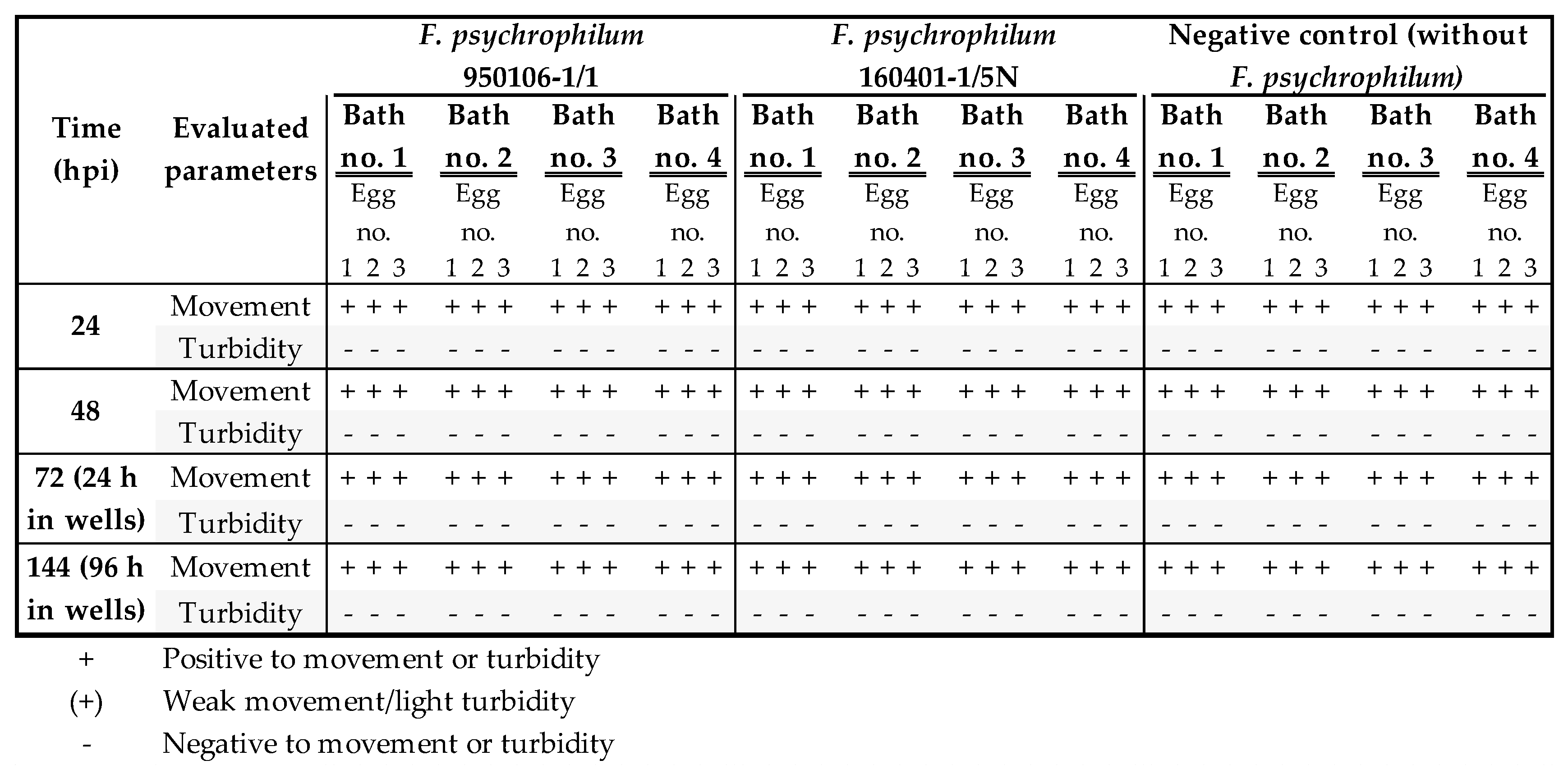
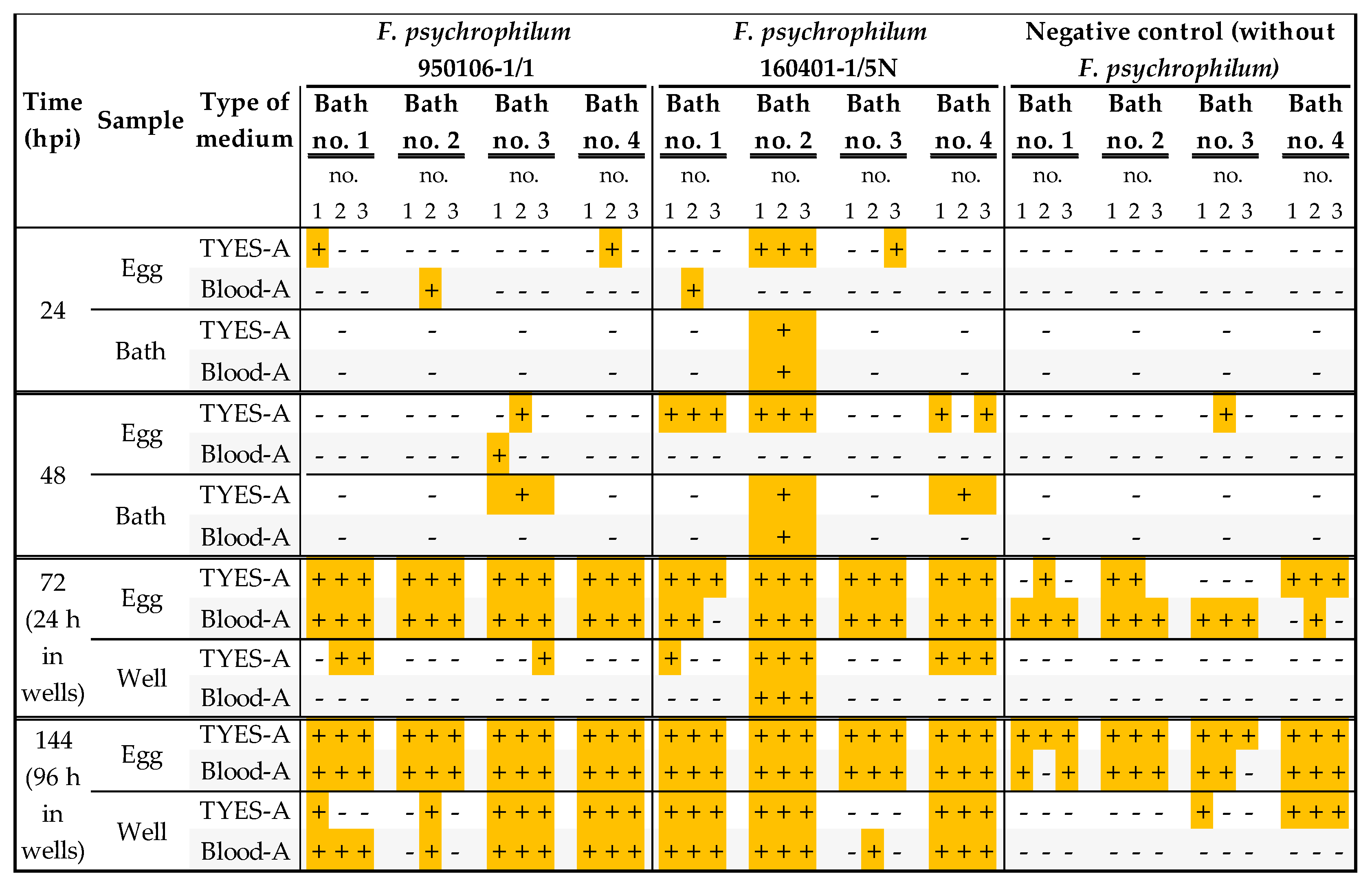
3.1. Establishment of a Bath Bacterial Challenge Method (Section A)
3.2. Interactions of Phages with Rainbow Trout Eyed Eggs (Section B)
3.2.1. Constant Exposure of Eyed Eggs to F. psychrophilum Phages (Exp. I, Section B)
3.2.2. Bath Exposure of Eyed Eggs to F. psychrophilum Phages (Exp. II, Section B)
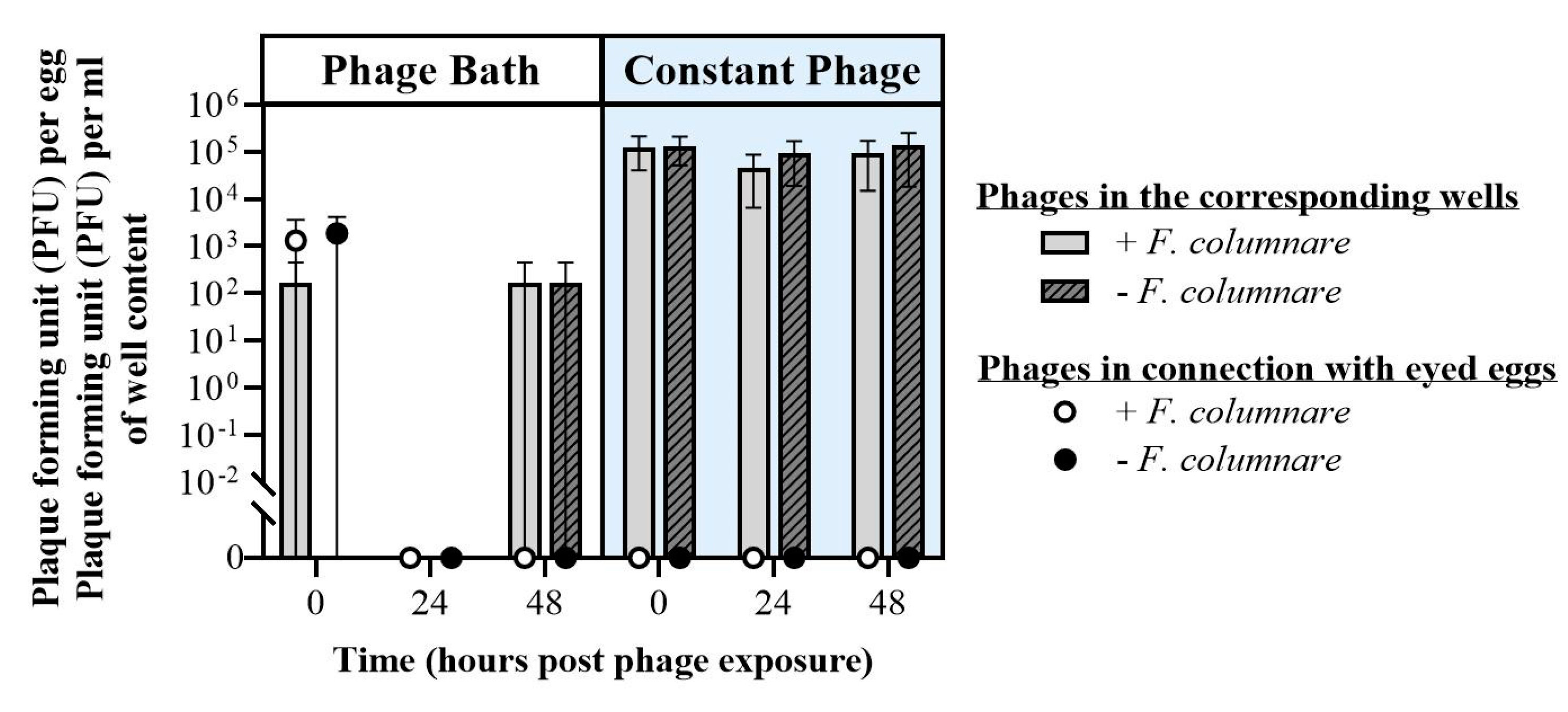
3.2.3. Bath and Constant Exposure of Eyed Eggs to F. columnare Phages (Exp. III, Section B)
3.3. Experiments to Evaluate the Use of Phages as Control Agents (Section C)
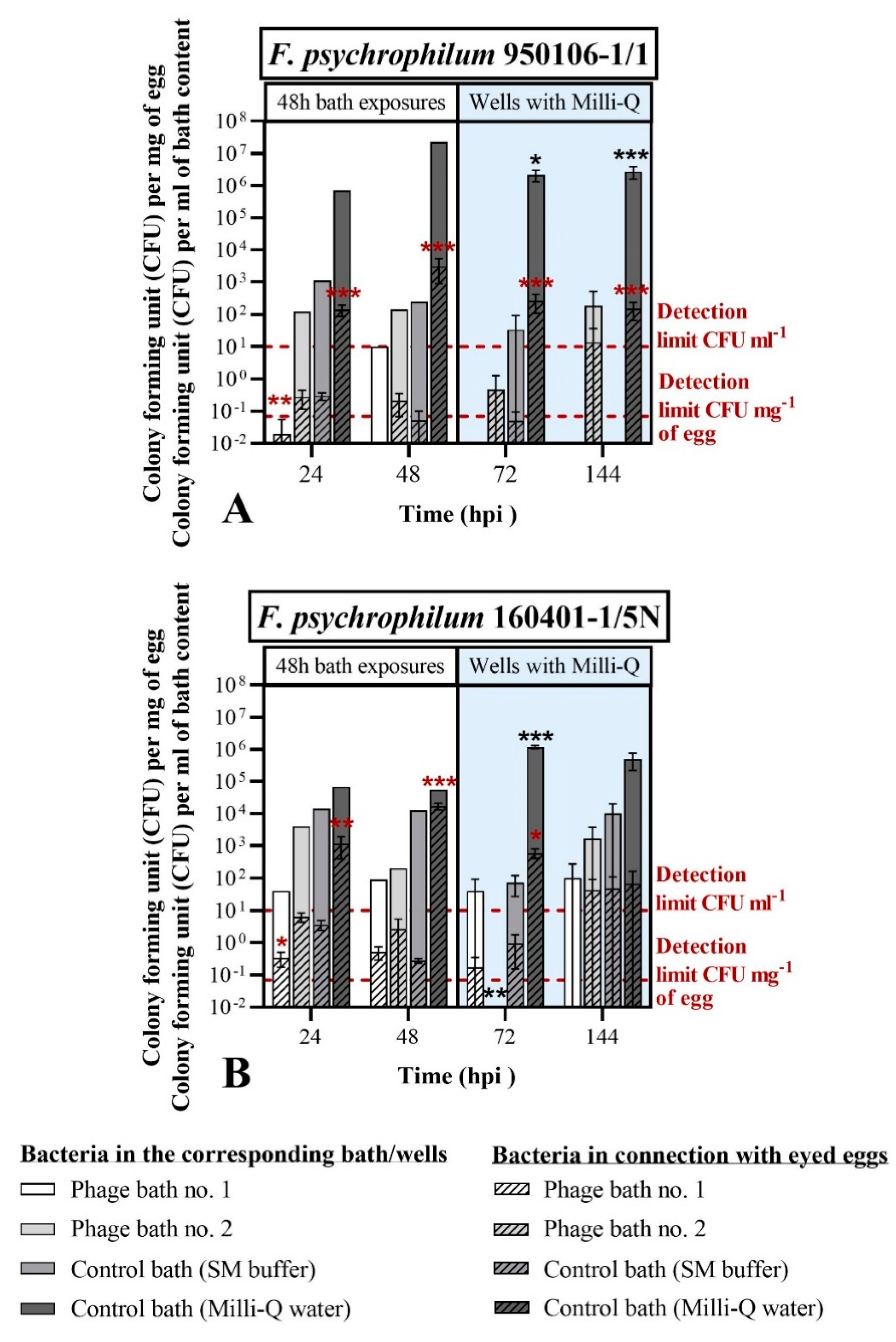
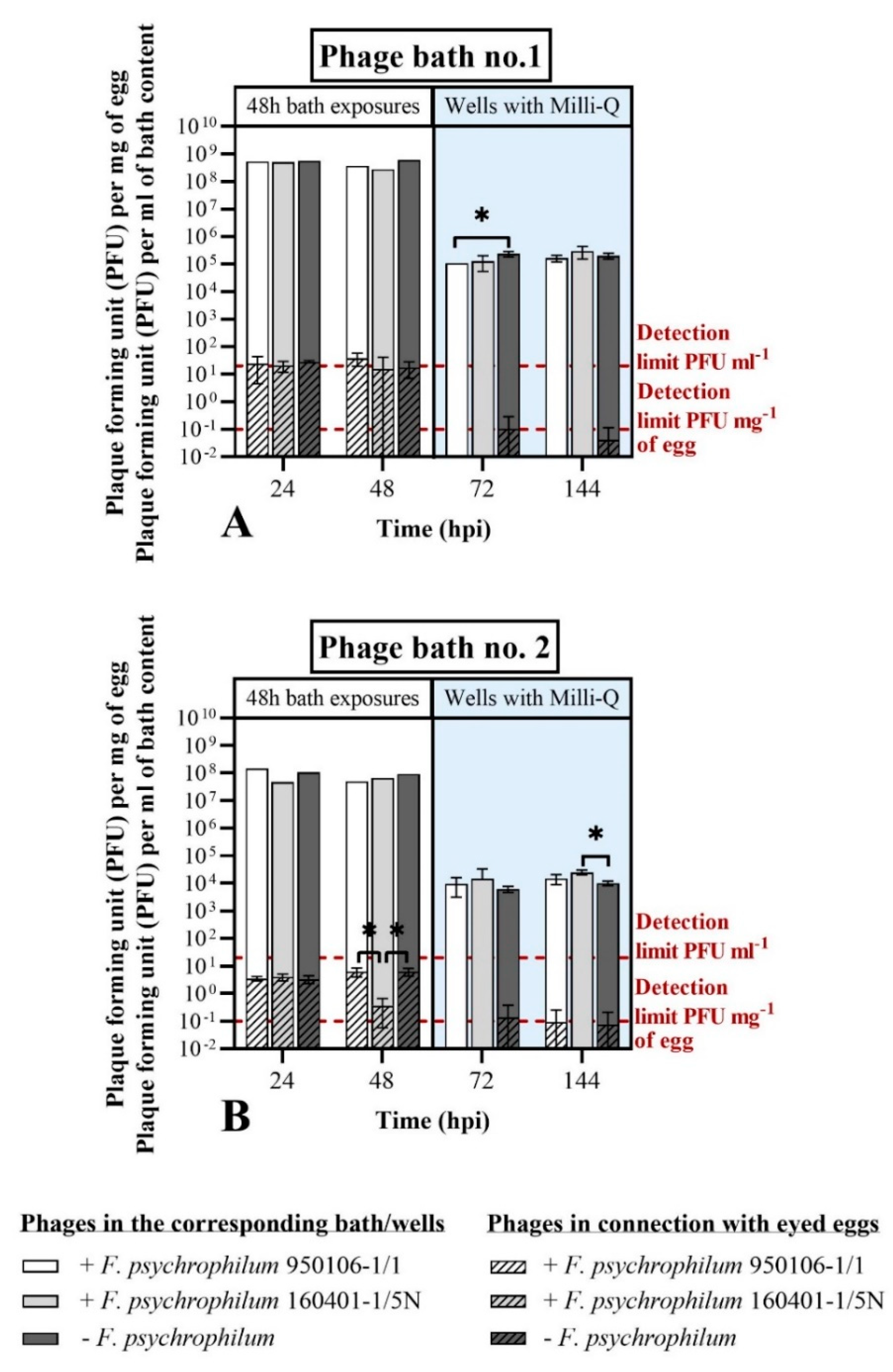
3.3.1. Phage Bath of F. psychrophilum Challenged Eggs (Exp. I, Section C)
3.3.2. Phage Exposure of F. columnare Challenged Eggs (Exp. II, Section C)
4. Discussion
4.1. Experimental Infection Method
4.2. Rainbow Trout Eyed Eggs–Bacteriophage Interactions
4.3. Phages as Control Agents for F. psychrophilum in Eyed Eggs
4.4. Phages as Control Agents for F. columnare in Eyed Eggs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (hpi) | Evaluated Parameters | + F. psychrophilum | − F. psychrophilum | |||||
|---|---|---|---|---|---|---|---|---|
| No. 1 | No. 2 | No. 3 | No. 1 | No. 2 | No. 3 | |||
| Exp. no. 1 | 21 | Movement | (+) | + | + | + | + | + |
| Turbidity | (+) | - | (+) | - | - | - | ||
| 25 | Movement | + | + | + | + | + | + | |
| Turbidity | (+) | (+) | (+) | - | - | - | ||
| Exp. no. 2 | 0 | Movement | + | + | + | + | + | + |
| Turbidity | - | - | - | - | - | - | ||
| 24 | Movement | + | - | + | + | + | + | |
| Turbidity | - | (+) | - | - | - | - | ||
| Exp. no. 3 | 0 | Movement | + | + | + | + | + | + |
| Turbidity | - | - | - | - | - | - | ||
| 24 | Movement | + | - | + | + | + | + | |
| Turbidity | - | - | - | - | - | - | ||
| Time (hpi) | Sample (Medium Type) | + F. psychrophilum | − F. psychrophilum | |||||
|---|---|---|---|---|---|---|---|---|
| No. 1 | No. 2 | No. 3 | No. 1 | No. 2 | No. 3 | |||
| Exp. no. 1 | 25 | Egg | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Well (Blood-A) | - | - | - | - | - | - | ||
| Exp. no. 2 | 0 | Egg (TYES-A) | + | - | + | + | + | - |
| Bath (Blood-A) | - | - | ||||||
| 24 | Egg (TYES-A) | - | - | - | + | + | - | |
| Well (TYES-A; Blood-A) | -; - | -; - | -; - | +; - | -; - | -; - | ||
| Exp. no. 3 | 0 | Egg | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Bath (Blood-A) | - | - | ||||||
| 24 | Egg (TYES-A) | + | - | - | + | + | + | |
| Well (TYES-A) | - | - | - | + | + | + | ||
| Time (h) | Medium Type | + FpV4 | + FPSV-D22 | Control | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Crude Lysate | PEG-Purified | PEG-Purified | |||||||||||
| No. 1 | No. 2 | No. 3 | No. 1 | No. 2 | No. 3 | No. 1 | No. 2 | No. 3 | No. 1 | No. 2 | No. 3 | ||
| 2 | TYES-A | - | - | - | - | - | - | - | - | - | - | - | - |
| Blood-A | - | - | - | - | - | - | - | - | - | - | - | - | |
| 27 | TYES-A | + | - | - | + | - | - | - | - | - | - | - | - |
| Blood-A | + | - | - | + | - | - | - | - | - | - | - | - | |
| 49 | TYES-A | - | - | - | - | - | - | - | - | - | - | - | - |
| Blood-A | + | - | + | - | - | - | + | - | + | - | + | - | |
| 71 | TYES-A | n.d. | n.d. | n.d. | n.d. | ||||||||
| Blood-A | - | - | - | - | - | - | - | - | - | - | - | - | |
| 144 | TYES-A | n.d. | n.d. | n.d. | n.d. | ||||||||
| Blood-A | - | - | - | - | - | + | + | - | - | + | - | - | |
| Time (h) | Medium Type | + Crude Lysate | Control | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| + FpV4 | + FPSV-D22 | |||||||||
| No. 1 | No. 2 | No. 3 | No. 1 | No. 2 | No. 3 | No. 1 | No. 2 | No. 3 | ||
| 0 * | TYES-A | - | - | - | ||||||
| Blood-A | - | - | - | |||||||
| 24 | TYES-A | - | - | - | - | - | - | - | - | - |
| Blood-A | - | - | - | - | - | - | - | - | - | |
| 46 | TYES-A | - | - | - | - | - | - | - | - | - |
| Blood-A | - | - | - | - | - | - | - | - | - | |
| 68 | TYES-A | n.d. | n.d. | n.d. | ||||||
| Blood-A | - | - | - | - | - | - | - | + | - | |
| 144 | TYES-A | n.d. | n.d. | n.d. | ||||||
| Blood-A | - | - | + | + | - | - | + | + | - | |




References
- Groot, E.P.; Alderdice, D.F. Fine structure of the external egg membrane of five species of Pacific salmon and steelhead trout. Can. J. Zool. 1985, 63, 552–566. [Google Scholar] [CrossRef]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost microbiomes: The state of the art in their characterization, manipulation and importance in aquaculture and fisheries. Front. Microbiol. 2014, 5, 207. [Google Scholar] [CrossRef] [Green Version]
- Hansen, G.H.; Olafsen, J.A. Bacterial Interactions in Early Life Stages of Marine Cold Water Fish. Microb. Ecol. 1999, 38, 1–26. [Google Scholar] [CrossRef]
- Sauter, R.W.; Williams, C.; Meyer, E.A.; Celnik, B.; Banks, J.L.; Leith, D.A. A study of bacteria present within unfertilized salmon eggs at the time of spawning and their possible relation to early lifestage disease. J. Fish Dis. 1987, 10, 193–203. [Google Scholar] [CrossRef]
- Trust, T.J. The Bacterial Population in Vertical Flow Tray Hatcheries During Incubation of Salmonid Eggs. J. Fish. Res. Board Can. 1972, 29, 567–571. [Google Scholar] [CrossRef]
- Hansen, G.H.; Olafsen, J.A. Bacterial Colonization of Cod (Gadus morhua L.) and Halibut (Hippoglossus hippoglossus) Eggs in Marine Aquaculture. Appl. Environ. Microbiol. 1989, 55, 1435–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Swaef, E.; Broeck, W.V.D.; Dierckens, K.; Decostere, A. Disinfection of teleost eggs: A review. Rev. Aquac. 2016, 8, 321–341. [Google Scholar] [CrossRef]
- Borg, A.F. Studies on Myxobacteria Associated with Diseases in Salmonid Fishes. Ph.D. Thesis, University of Washington, Seattle, WA, USA, 1948. [Google Scholar]
- Bernardet, J.-F.; Segers, P.; Vancanneyt, M.; Berthe, F.; Kersters, K.; Vandamme, P. Cutting a Gordian Knot: Emended Classification and Description of the Genus Flavobacterium, Emended Description of the Family Flavobacteriaceae, and Proposal of Flavobacterium hydatis nom. nov. (Basonym, Cytophaga aquatilis Strohl and Tait 1978). Int. J. Syst. Bacteriol. 1996, 46, 128–148. [Google Scholar] [CrossRef]
- Madsen, L.; Dalsgaard, I. Comparative studies of Danish Flavobacterium psychrophilum isolates: Ribotypes, plasmid profiles, serotypes and virulence. J. Fish Dis. 2000, 23, 211–218. [Google Scholar] [CrossRef]
- Nematollahi, A.; Decostere, A.; Pasmans, F.; Haesebrouck, F. Flavobacterium psychrophilum infections in salmonid fish. J. Fish Dis. 2003, 26, 563–574. [Google Scholar] [CrossRef]
- Madsen, L.; Møller, J.D.; Dalsgaard, I. Flavobacterium psychrophilum in rainbow trout, Oncorhynchus mykiss (Walbaum), hatcheries: Studies on broodstock, eggs, fry and environment. J. Fish Dis. 2005, 28, 39–47. [Google Scholar] [CrossRef]
- Rangdale, R.E.; Richards, R.E.; Alderman, D.J. Isolation of Cytophaga psychrophila, causal agent of Rainbow Trout Fry Syndrome (RTFS) from reproductive fluids and egg surfaces of rainbow trout (Oncorhynchus mykiss). Bull. Eur. Ass. Fish Pathol. 1996, 16, 63–67. [Google Scholar]
- Brown, L.; Cox, W.; Levine, R. Evidence that the causal agent of bacterial cold-water disease Flavobacterium psychrophilum is transmitted within salmonid eggs. Dis. Aquat. Org. 1997, 29, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Madsen, L.; Dalsgaard, I. Water recirculation and good management: Potential methods to avoid disease outbreaks with Flavobacterium psychrophilum. J. Fish Dis. 2008, 31, 799–810. [Google Scholar] [CrossRef]
- Declercq, A.M.; Haesebrouck, F.; Broeck, W.V.D.; Bossier, P.; Decostere, A. Columnaris disease in fish: A review with emphasis on bacterium-host interactions. Veter. Res. 2013, 44, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzen, E. Studies on Flexibacter psychrophilus in Relation to Rainbow Trout Fry Syndrome (RTFS). Ph.D. Thesis, National Veterinary Laboratory, Århus & Royal Veterinary and Agricultural University, Copenhagen, Denmark, 1994. [Google Scholar]
- Nilsen, H.; Olsen, A.B.; Vaagnes, Ø.; Hellberg, H.; Bottolfsen, K.; Skjelstad, H.; Colquhoun, D.J. Systemic Flavobacterium psychrophilum infection in rainbow trout, Oncorhynchus mykiss (Walbaum), farmed in fresh and brackish water in Norway. J. Fish Dis. 2011, 34, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Suomalainen, L.-R.; Tiirola, M.A.; Valtonen, E.T. Effect of Pseudomonas sp. MT5 baths on Flavobacterium columnare infection of rainbow trout and on microbial diversity on fish skin and gills. Dis. Aquat. Org. 2005, 63, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Salmond, G.P.C.; Fineran, P.C. A century of the phage: Past, present and future. Nat. Rev. Genet. 2015, 13, 777–786. [Google Scholar] [CrossRef]
- Culot, A.; Grosset, N.; Gautier, M. Overcoming the challenges of phage therapy for industrial aquaculture: A review. Aquaculture 2019, 513, 734423. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, J.D.; Kazimierczak, J.; Sowińska, P.M.; Wójcik, E.A.; Siwicki, A.K.; Dastych, J. Growing Trend of Fighting Infections in Aquaculture Environment—Opportunities and Challenges of Phage Therapy. Antibiotics 2020, 9, 301. [Google Scholar] [CrossRef]
- Rao, B.M.; Lalitha, K. Bacteriophages for aquaculture: Are they beneficial or inimical. Aquaculture 2015, 437, 146–154. [Google Scholar] [CrossRef]
- Christiansen, R.H.; Dalsgaard, I.; Middelboe, M.; Lauritsen, A.H.; Madsen, L. Detection and Quantification of Flavobacterium psychrophilum-Specific Bacteriophages In Vivo in Rainbow Trout upon Oral Administration: Implications for Disease Control in Aquaculture. Appl. Environ. Microbiol. 2014, 80, 7683–7693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, L.; Bertelsen, S.K.; Dalsgaard, I.; Middelboe, M. Dispersal and Survival of Flavobacterium psychrophilum Phages In Vivo in Rainbow Trout and In Vitro under Laboratory Conditions: Implications for Their Use in Phage Therapy. Appl. Environ. Microbiol. 2013, 79, 4853–4861. [Google Scholar] [CrossRef] [Green Version]
- Castillo, D.; Higuera, G.; Villa, M.; Middelboe, M.; Dalsgaard, I.; Madsen, L.; Espejo, R.T. Diversity of Flavobacterium psychrophilum and the potential use of its phages for protection against bacterial cold water disease in salmonids. J. Fish Dis. 2012, 35, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Donati, V.L.; Dalsgaard, I.; Sundell, K.; Castillo, D.; Er-Rafik, M.; Clark, J.; Wiklund, T.; Middelboe, M.; Madsen, L. Phage-Mediated Control of Flavobacterium psychrophilum in Aquaculture: In vivo Experiments to Compare Delivery Methods. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Elaanto, E.; Bamford, J.K.H.; Ravantti, J.J.; Esundberg, L.-R. The use of phage FCL-2 as an alternative to chemotherapy against columnaris disease in aquaculture. Front. Microbiol. 2015, 6, 829. [Google Scholar] [CrossRef] [Green Version]
- Dalsgaard, I.; Madsen, L. Bacterial pathogens in rainbow trout, Oncorhynchus mykiss (Walbaum), reared at Danish freshwater farms. J. Fish Dis. 2000, 23, 199–209. [Google Scholar] [CrossRef]
- Madsen, L.; Dalsgaard, I. Reproducible methods for experimental infection with Flavobacterium psychrophilum in rainbow trout Oncorhynchus mykiss. Dis. Aquat. Org. 1999, 36, 169–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundell, K.; Landor, L.; Nicolas, P.; Jørgensen, J.; Castillo, D.; Middelboe, M.; Dalsgaard, I.; Donati, V.L.; Madsen, L.; Wiklund, T. Phenotypic and Genetic Predictors of Pathogenicity and Virulence in Flavobacterium psychrophilum. Front. Microbiol. 2019, 10, 1711. [Google Scholar] [CrossRef] [PubMed]
- Holt, R.A.; Rohovec, J.S.; Fryer, J.L. Bacterial cold-water disease. Bact. Dis. Fish 1993, 3–22. [Google Scholar]
- Runtuvuori-Salmela, A.; Kunttu, H.M.T.; Laanto, E.; Almeida, G.M.F.; Mäkelä, K.; Middelboe, M.; Sundberg, L.-R. Prevalence of genetically similar Flavobacterium columnare phages across aquaculture environments reveals a strong potential for pathogen control. bioRxiv 2020. [Google Scholar] [CrossRef]
- Laanto, E.; Sundberg, L.-R.; Bamford, J.K.H. Phage Specificity of the Freshwater Fish Pathogen Flavobacterium columnare. Appl. Environ. Microbiol. 2011, 77, 7868–7872. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.L.; Fryer, J.L.; Rohovec, J.S. Comparison of six media for the cultivation of Flexibacter columnaris. Fish Pathol. 1988, 23, 91–94. [Google Scholar] [CrossRef] [Green Version]
- Stenholm, A.R.; Dalsgaard, I.; Middelboe, M. Isolation and Characterization of Bacteriophages Infecting the Fish Pathogen Flavobacterium psychrophilum. Appl. Environ. Microbiol. 2008, 74, 4070–4078. [Google Scholar] [CrossRef] [Green Version]
- Castillo, D.; Middelboe, M. Genomic diversity of bacteriophages infecting the fish pathogen Flavobacterium psychrophilum. FEMS Microbiol. Lett. 2016, 363, fnw272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, D.; Andersen, N.; Kalatzis, P.G.; Middelboe, M. Large Phenotypic and Genetic Diversity of Prophages Induced from the Fish Pathogen Vibrio anguillarum. Viruses 2019, 11, 983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cipriano, R.C. Intraovum Infection Caused by Flavobacterium psychrophilum among Eggs from Captive Atlantic Salmon Broodfish. J. Aquat. Anim. Health 2005, 17, 275–283. [Google Scholar] [CrossRef]
- Cipriano, R.C. Bacterial Analysis of Fertilized Eggs of Atlantic Salmon from the Penobscot, Naraguagus, and Machias Rivers, Maine. J. Aquat. Anim. Health. 2015, 27, 172–177. [Google Scholar] [CrossRef]
- Cipriano, R.C.; Ford, L.A.; Teska, J.D. Association of Cytophaga psychrophila with mortality among eyed eggs of Atlantic Salmon (Salmo salar). J. Wildl. Dis. 1995, 31, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Jansson, E.; Haenen, O.L.M.; Nonnemann, B.; Madsen, L.; Gelderen, v.E.; Aspán, A.; Säker, E.; Boonstra, M.; Gulla, S.; Colquhoun, D.J. MALDI-TOF MS: A diagnostic tool for identification of bacterial fish pathogens. Bull. Eur. Ass. Fish Pathol. 2020, 40, 240–248. [Google Scholar]
- Clokie, M.R.J.; Kropinski, A.M. (Eds.) Bacteriophages: Methods and Protocols, Volume 1: Isolation, Characterization, and Interactions, vol. 501; Humana Press, a part of Springer Science+Business Media: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Ashrafi, R.; Bruneaux, M.; Sundberg, L.-R.; Pulkkinen, K.; Valkonen, J.; Ketola, T. Broad thermal tolerance is negatively correlated with virulence in an opportunistic bacterial pathogen. Evol. Appl. 2018, 11, 1700–1714. [Google Scholar] [CrossRef] [PubMed]
- Woynarovich, G.; Hoitsy, A.; Moth-Poulsen, T. Small-Scale Rainbow Trout Farming; FAO Fisheries and Aquaculture Technical Paper No. 561; FAO: Rome, Italy, 2011; 81p. [Google Scholar]
- Rangdale, R.E.; Richards, R.H.; Alderman, D.J. Colonization of eyed rainbow trout ova with Flavobacterium psychrophilum leads to rainbow trout fry syndrome in fry. Bull. Eur. Ass. Fish Pathol. 1997, 17, 108–111. [Google Scholar]
- Ekman, E.; Åkerman, G.; Balk, L.; Norrgren, L. Nanoinjection as a tool to mimic vertical transmission of Flavobacterium psychrophilum in rainbow trout Oncorhynchus mykiss. Dis. Aquat. Org. 2003, 55, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Barnes, M.E. A Review of Flavobacterium psychrophilum Biology, Clinical Signs, and Bacterial Cold Water Disease Prevention and Treat. Open Fish Sci. J. 2011, 4, 40–48. [Google Scholar] [CrossRef]
- Vatsos, I.; Thompson, K.; Adams, A. Starvation of Flavobacterium psychrophilum in broth, stream water and distilled water. Dis. Aquat. Org. 2003, 56, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Madetoja, J.; Nystedt, S.; Wiklund, T. Survival and virulence of Flavobacterium psychrophilum in water microcosms. FEMS Microbiol. Ecol. 2003, 43, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Sundberg, L.-R.; Kunttu, H.M.T.; Valtonen, E.T. Starvation can diversify the population structure and virulence strategies of an environmentally transmitting fish pathogen. BMC Microbiol. 2014, 14, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FINEFISH Collective Research Project Consortium. Control of Malformations in Fish Aquaculture. Science and Practice. 2009. Available online: http://feap.info/index.php/about/projects/ (accessed on 27 February 2021).
- Evenhuis, J.P.; Lipscomb, R.; Birkett, C. Virulence variations of Flavobacterium columnare in rainbow trout (Oncorhynchus mykiss) eyed eggs and alevin. J. Fish Dis. 2021, 1–7. [Google Scholar] [CrossRef]
- Dong, H.T.; LaFrentz, B.; Pirarat, N.; Rodkhum, C. Phenotypic characterization and genetic diversity of Flavobacterium columnare isolated from red tilapia, Oreochromis sp., in Thailand. J. Fish Dis. 2015, 38, 901–913. [Google Scholar] [CrossRef]
- Barony, G.M.; Tavares, G.C.; Assis, G.B.N.; Luz, R.K.; Figueiredo, H.C.P.; Leal, C.A.G. New hosts and genetic diversity of Flavobacterium columnare isolated from Brazilian native species and Nile tilapia. Dis. Aquat. Org. 2015, 117, 1–11. [Google Scholar] [CrossRef]
- Barnes, M.E.; Bergmann, D.; Jacobs, J.; Gabel, M. Effect of Flavobacterium columnare inoculation, antibiotic treatments and resident bacteria on rainbow trout Oncorhynchus mykiss eyed egg survival and external membrane structure. J. Fish Biol. 2009, 74, 576–590. [Google Scholar] [CrossRef] [PubMed]
- Al-Harbi, A.H.; Uddin, M.N. Seasonal variation in the intestinal bacterial flora of hybrid tilapia (Oreochromis niloticus × Oreochromis aureus) cultured in earthen ponds in Saudi Arabia. Aquaculture 2004, 229, 37–44. [Google Scholar] [CrossRef]
- Rørbo, N.; Rønneseth, A.; Kalatzis, P.G.; Rasmussen, B.B.; Engell-Sørensen, K.; Kleppen, H.P.; Wergeland, H.I.; Gram, L.; Middelboe, M. Exploring the Effect of Phage Therapy in Preventing Vibrio anguillarum Infections in Cod and Turbot Larvae. Antibiotics 2018, 7, 42. [Google Scholar] [CrossRef] [Green Version]
- Zawada, A.; Polechoński, R.; Bronowska, A. Iodine Disinfection of Sea Trout, Salmo Trutta (L.), Eggs and the Affect on Egg Surfaces. Arch. Pol. Fish. 2014, 22, 121–126. [Google Scholar] [CrossRef]
- Silva, Y.J.; Costa, L.; Pereira, C.; Mateus, C.; Cunha, Â.; Calado, R.; Gomes, N.C.M.; Pardo, M.A.; Hernandez, I.; Almeida, A. Phage Therapy as an Approach to Prevent Vibrio anguillarum Infections in Fish Larvae Production. PLoS ONE 2014, 9, e114197. [Google Scholar] [CrossRef] [PubMed]
- Yamagami, K. 7 Mechanisms of Hatching in Fish. Fish Physiol. 1988, 11, 447–499. [Google Scholar] [CrossRef]
- Castillo, D.; Jørgensen, J.; Sundell, K.; Madsen, L.; Dalsgaard, I.; Wiklund, T.; Middelboe, M. Genome-informed approach to identify genetic determinants of Flavobacterium psychrophilum phage susceptibility. Environ. Microbiol. 2021. in review. [Google Scholar]
- Almeida, G.M.F.; Laanto, E.; Ashrafi, R.; Sundberg, L.-R. Bacteriophage Adherence to Mucus Mediates Preventive Protection against Pathogenic Bacteria. mBio 2019, 10, e01984-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardet, J.F.; Kerouault, B. Phenotypic and genomic studies of “Cytophaga psychrophila” isolated from diseased rainbow trout (Oncorhynchus mykiss) in France. Appl. Environ. Microbiol. 1989, 55, 1796–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatsos, I.; Thompson, K.; Adams, A. Adhesion of the fish pathogen Flavobacterium psychrophilum to unfertilized eggs of rainbow trout (Oncorhynchus mykiss) and n-hexadecane. Lett. Appl. Microbiol. 2001, 33, 178–182. [Google Scholar] [CrossRef]
- Barnes, M.; Bergmann, D.; Stephenson, H.; Gabel, M.; Cordes, R. Bacterial Numbers from Landlocked Fall Chinook Salmon Eyed Eggs Subjected to Various Formalin Treatments as Determined by Scanning Electron Microscopy and Bacteriological Culture Methods. N. Am. J. Aquac. 2005, 67, 23–33. [Google Scholar] [CrossRef]
- Loch, T.P.; Faisal, M. Gamete-associated flavobacteria of the oviparous Chinook salmon (Oncorhynchus tshawytscha) in lakes Michigan and Huron, North America. J. Microbiol. 2016, 54, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Suomalainen, L.-R.; Tiirola, A.M.; Valtonen, E.T. Influence of rearing conditions on Flavobacterium columnare infection of rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis. 2005, 28, 271–277. [Google Scholar] [CrossRef]
- Cai, W.; De La Fuente, L.; Arias, C.R. Biofilm Formation by the Fish Pathogen Flavobacterium columnare: Development and Parameters Affecting Surface Attachment. Appl. Environ. Microbiol. 2013, 79, 5633–5642. [Google Scholar] [CrossRef] [PubMed] [Green Version]








| Study Name | Infection with Flavobacterium spp. | Type of Exposure to Phages | Phages | Type of Preparation |
|---|---|---|---|---|
| Exp. I Section B | No | Constant | FpV4 and FPSV-D22 (singularly) | Crude lysates and PEG-purified in sterile TYES-B |
| Exp. II Section B | No | 4 h bath | FpV4 and FPSV-D22 (singularly) | Crude lysates |
| Exp. III Section B | No | 30 min bath; constant | FCL-2 and FCOV-F27 (singularly) | Crude lysates |
| Exp. I Section C | Yes | 48 h bath | FpV4 and FPSV-D22 (mixed 1:1) | PEG-purified in SM buffer |
| Exp. II Section C | Yes | 2 h bath; constant | FCL-2 | Diafiltration |
| (A) Constant Phage Exposure (Exp. I Section B) | |||||||||||||
| Time (h) | Evaluated Parameters | FpV4 | FPSV-D22 | Control | |||||||||
| Crude Lysate | PEG-Purified | PEG-Purified | |||||||||||
| no. 1 | no. 2 | no. 3 | no. 1 | no. 2 | no. 3 | no. 1 | no. 2 | no. 3 | no. 1 | no. 2 | no. 3 | ||
| 2 | Movement | + | - | + | (+) | + | + | + | + | (+) | + | + | + |
| Turbidity | - | - | - | - | - | - | - | - | - | - | - | - | |
| 27 | Movement | + | + | - | + | + | + | + | + | + | + | - | + |
| Turbidity | - | - | (+) | - | - | - | - | - | - | - | - | - | |
| 49 | Movement | + | + | - | + | + | + | + | + | + | + | + | + |
| Turbidity | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | - | (+) | (+) | |
| 71 | Movement | - | - | + | + | + | + | (+) | + | + | + | + | (+) |
| Turbidity | + | (+) | - | (+) | (+) | (+) | - | (+) | (+) | - | - | (+) | |
| 144 | Alive/Dead | Dead | Dead | Dead | Alive | Alive | Dead | Dead | Dead | Dead | Dead | Dead | Alive |
| Hatched or not | Yes | No | No | Yes | Yes | Yes | Yes | No | Yes | No | Yes | No | |
| (B) Phage bath exposure (Exp. II section B) | |||||||||||||
| Time (h) | Evaluated Parameters | Crude lysate | Control | ||||||||||
| FpV4 | FPSV-D22 | ||||||||||||
| no. 1 | no. 2 | no. 3 | no. 1 | no. 2 | no. 3 | no. 1 | no. 2 | no. 3 | |||||
| 0 | Movement | + | + | + | (+) | (+) | (+) | + | + | + | |||
| Turbidity | - | - | - | - | - | - | - | - | - | ||||
| 24 | Movement | (+) | (+) | (+) | + | (+) | (+) | + | + | + | |||
| Turbidity | - | - | - | - | - | - | - | - | - | ||||
| 46 | Movement | + | + | + | + | + | + | + | + | + | |||
| Turbidity | - | - | - | - | - | - | - | - | (+) | ||||
| 68 | Movement | + | (+) | + | (+) | + | + | + | + | + | |||
| Turbidity | - | - | - | - | - | - | (+) | (+) | (+) | ||||
| 144 | Alive/Dead | Alive | Alive | Alive | Alive | Alive | Dead | Alive | Alive | Alive | |||
| Hatched or not | No | Yes | Yes | No | Yes | No | Yes | No | Yes | ||||
| (A) Exposure to Phage FCL-2 | ||||||||
|---|---|---|---|---|---|---|---|---|
| Medium | Sample | Phage exposure | Time (h) | |||||
| 0 | 24 | 48 | ||||||
| No. 1 | No. 2 | No. 1 | No. 2 | No. 1 | No. 2 | |||
| WATER | Well (PFU ml−1) | Bath | 3.0 × 106 | 9.0 × 105 | 3.0 × 106 | 5.0 × 105 | 4.0 × 106 | 3.0 × 106 |
| Constant | 9.0 × 107 | 1.0 × 109 | 9.0 × 106 | 1.0 × 109 | 3.0 × 107 | 2.5 × 107 | ||
| Control (no egg) | 2.3 × 109 | 1.0 × 109 | 1.0 × 107 | 1.0 × 107 | 1.5 × 107 | 5.0 × 106 | ||
| Egg (PFU egg−1) | Bath | 0 | 0 | 0 | 0 | 0 | 0 | |
| Constant | 0 | 2.0 × 101 | 2.0 × 102 | 0 | 9.3 × 101 | 5.0 × 101 | ||
| SHIEH | Well (PFU ml−1) | Bath | 8.0 × 106 | 7.0 × 107 | 2.0 × 108 | 1.0 × 107 | 9.0 × 106 | 7.0 × 107 |
| Constant | 3.5 × 109 | 2.0 × 109 | 8.0 × 109 | 2.0 × 109 | 4.5 × 109 | 5.5 × 109 | ||
| Control (no egg) | 3.5 × 109 | 2.0 × 109 | 3.5 × 109 | 4.0 × 109 | 2.0 × 109 | 8.0 × 108 | ||
| Egg (PFU egg−1) | Bath | 0 | 0 | 0 | 0 | 0 | 0 | |
| Constant | 0 | 0 | 0 | 0 | 0 | 0 | ||
| (B) Exposure to Phage FCOV-F27 | ||||||||
| Medium | Sample | Phage exposure | Time (h) | |||||
| 0 | 24 | 48 | ||||||
| No. 1 | No. 2 | No. 1 | No. 2 | No. 1 | No. 2 | |||
| WATER | Well (PFU ml−1) | Bath | 1.5 × 105 | 1.0 × 105 | + | + | + | 4.0 × 105 |
| Constant | 1.3 × 107 | 5.0 × 106 | 3.0 × 106 | 2.0 × 106 | + | 2.0 × 106 | ||
| Control (no egg) | 1.5 × 107 | 2.1 × 107 | + | 3.0 × 106 | 1.8 × 107 | + | ||
| Egg (PFU egg−1) | Bath | 0 | 0 | 0 | 0 | 0 | 0 | |
| Constant | 2.3 × 102 | 0 | 5.0 × 100 | 0 | 0 | 0 | ||
| SHIEH | Well (PFU ml−1) | Bath | 2.0 × 106 | 1.0 × 106 | 6.0 × 106 | 7.0 × 106 | 2.0 × 107 | 1.0 × 107 |
| Constant | 4.5 × 109 | 1.0 × 109 | 2.0 × 109 | 8.0 × 108 | 1.5 × 109 | 9.0 × 108 | ||
| Control (no egg) | 6.0 × 108 | 5.0 × 108 | 2.0 × 109 | 8.0 × 108 | 3.0 × 109 | 2.0 × 109 | ||
| Egg (PFU egg−1) | Bath | 0 | 0 | 0 | 0 | 0 | 0 | |
| Constant | 0 | 0 | 0 | 0 | 0 | 0 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donati, V.L.; Dalsgaard, I.; Runtuvuori-Salmela, A.; Kunttu, H.; Jørgensen, J.; Castillo, D.; Sundberg, L.-R.; Middelboe, M.; Madsen, L. Interactions between Rainbow Trout Eyed Eggs and Flavobacterium spp. Using a Bath Challenge Model: Preliminary Evaluation of Bacteriophages as Pathogen Control Agents. Microorganisms 2021, 9, 971. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9050971
Donati VL, Dalsgaard I, Runtuvuori-Salmela A, Kunttu H, Jørgensen J, Castillo D, Sundberg L-R, Middelboe M, Madsen L. Interactions between Rainbow Trout Eyed Eggs and Flavobacterium spp. Using a Bath Challenge Model: Preliminary Evaluation of Bacteriophages as Pathogen Control Agents. Microorganisms. 2021; 9(5):971. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9050971
Chicago/Turabian StyleDonati, Valentina L., Inger Dalsgaard, Anniina Runtuvuori-Salmela, Heidi Kunttu, Johanna Jørgensen, Daniel Castillo, Lotta-Riina Sundberg, Mathias Middelboe, and Lone Madsen. 2021. "Interactions between Rainbow Trout Eyed Eggs and Flavobacterium spp. Using a Bath Challenge Model: Preliminary Evaluation of Bacteriophages as Pathogen Control Agents" Microorganisms 9, no. 5: 971. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9050971