1. Introduction
Cryptosporidium spp. are intracellular protozoan parasites that cause moderate-to-severe diarrhea in humans and various animals. Two
Cryptosporidium species,
C. parvum and
C. hominis, are the major causative agents of human cryptosporidiosis [
1]. Children, neonatal animals, and immunocompromised individuals are especially susceptible to infection [
2]. In low-income countries, cryptosporidiosis is a common cause of diarrhea-associated mortality in young children [
3]. It was estimated that in 2016 alone, acute
Cryptosporidium infections caused more than 48,000 deaths and over 4.2 million disability-adjusted life-years in low- and middle-income countries [
4]. No effective vaccines are in clinical use, and nitazoxanide, the sole drug approved by the U.S. Food and Drug Administration for treating cryptosporidiosis, has poor efficacy in immunocompromised individuals [
5].
Various proteases and protein kinases secreted by several unique organelles are thought to be involved in host cell adhesion and invasion by apicomplexans parasites [
6]. Among them, insulinlike proteases (INS), members of the M16 family of metalloproteases, are characterized by the presence of a Zn
2+-binding motif His-Xaa-Xaa-Glu-His (HXXEH) [
7,
8]. The M16 family of proteases includes three subfamilies, M16A, M16B and M16C. M16A and M16C proteases have large molecular masses (>110 kDa) that can be subdivided into N- and C-terminal domains [
9], while M16B proteases are usually encoded by separate genes and form heterodimers after translation [
10].
The functions of INS in apicomplexans have attracted the attention of researchers. Falcilysin in
Plasmodium falciparum, a M16C protease, has been identified as a key component of the catabolic process, playing an important role in hemoglobin degradation [
11]. Toxolysin 4 (TLN4), a micronemal M16A in
Toxoplasma gondii, plays a potential role in host cell invasion in response to elevated calcium levels, as TLN4-knockout tachyzoites grow poorly [
12]. Another INS in
T. gondii, Toxolysin-1 (TLN1), a M16A protease secreted by the rhoptry, has been shown to participate in the initial interaction between the parasite and the host by means of protein C-terminal and prodomain cleavage, trafficking the C-terminal portion of the protein to the rhoptry for the formation of a detergent resistant complex [
13].
The results of comparative genomics analysis indicate that
C. parvum has 22 potential INS genes, 13 of which have the highest expression in early host cell infection [
14,
15]. In previous studies, INS-5, INS-15 and INS-20-19 were shown to be potentially involved in the early infection of
C. parvum [
16,
17,
18].
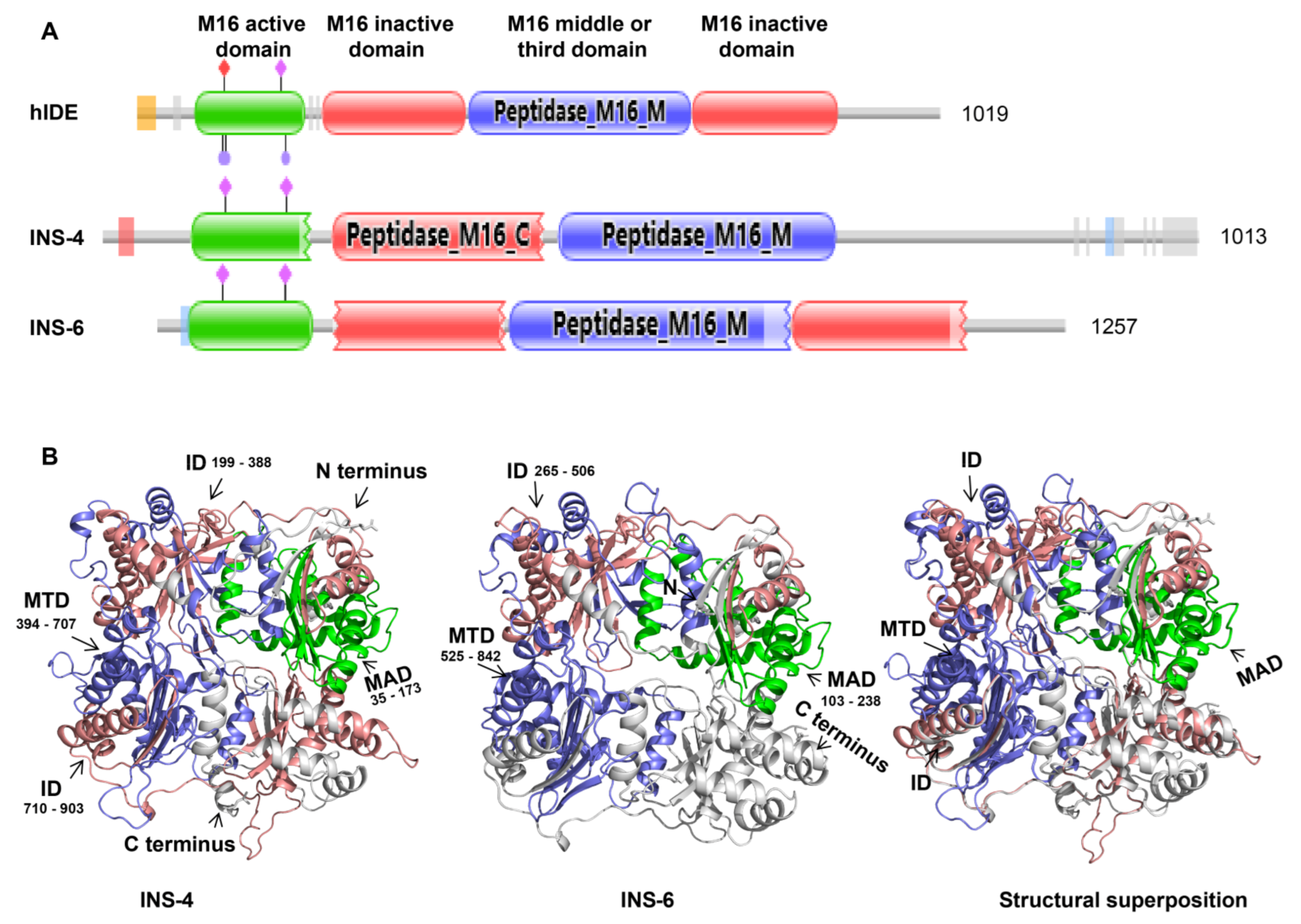
In this study, CpINS-4 and CpINS-6, two M16A proteases encoded by the cgd2_930 and cgd2_4270 genes, respectively, in chromosome 2 of C. parvum were studied. They were chosen because CpINS-4 is one of the few INS in C. parvum that have all four domains in functional M16 proteases, while CpINS-6 has one of the domains missing. Little is known of the impact of the absence of a domain on the function of INS. Therefore, we compared the biological functions of CpINS-4 and CpINS-6 in C. parvum infection.
2. Materials and Methods
2.1. Parasite, Host Cells and Cell Culture
Oocysts of the
C. parvum IOWA isolate were purchased from Waterborne, Inc. (New Orleans, LA, USA). Prior to use, they were treated on ice with 0.5% sodium hypochlorite for 10 min, washed three times with PBS and excysted at 37 °C for 30 min in the presence of 0.25% trypsin and 0.75% taurocholic acid as described [
19]. The sporozoites generated were collected by centrifugation at 5000×
g and 4 °C for 10 min, washed three times with PBS at 5,000×
g and 4 °C for 3 min, and resuspended in RPMI 1640 medium for in vitro infection.
Human ileocecal adenocarcinoma (HCT-8) cells were purchased from the Chinese Academy of Sciences, seeded into 12-well plates (5 × 105), and cultured to ~80% confluence. The culture was inoculated with 5 × 105 sporozoites of C. parvum in RPMI 1640 containing 10% fetal bovine serum (FBS) and 50 U/mL penicillin G and 50 U/mL streptomycin.
2.2. Identification and Analysis of cgd2_930 and cgd2_4270
The full-length sequences of
cgd2_930 and
cgd2_4270 genes were obtained from CryptoDB database (
https://cryptodb.org/cryptodb/app, accessed on 20 December 2020). Signal peptide and transmembrane domains (TMHMM) in them were predicted using SignalP-5.0 server (
http://www.cbs.dtu.dk/services/SignalP/, accessed on 20 December 2020) and TMHMM Server v. 2.0 (
http://www.cbs.dtu.dk/services/TMHMM/, accessed on 20 December 2020), respectively. Pfam 32.0 (
http://pfam.xfam.org/, accessed on 20 December 2020) was used to identify possible functional domains present in them. The tertiary structures of CpINS-4 and CpINS-6 were predicted using Protein Homology/analogY Recognition Engine V 2.0 (Phyre
2) (
http://www.sbg.bio.ic.ac.uk/phyre2/html/, accessed on 20 December 2020). The crystal structures of hIDE (PDB ID: 2jbu_B) and hIDE with copurified peptides [
20] were selected as the template for the homology modeling.
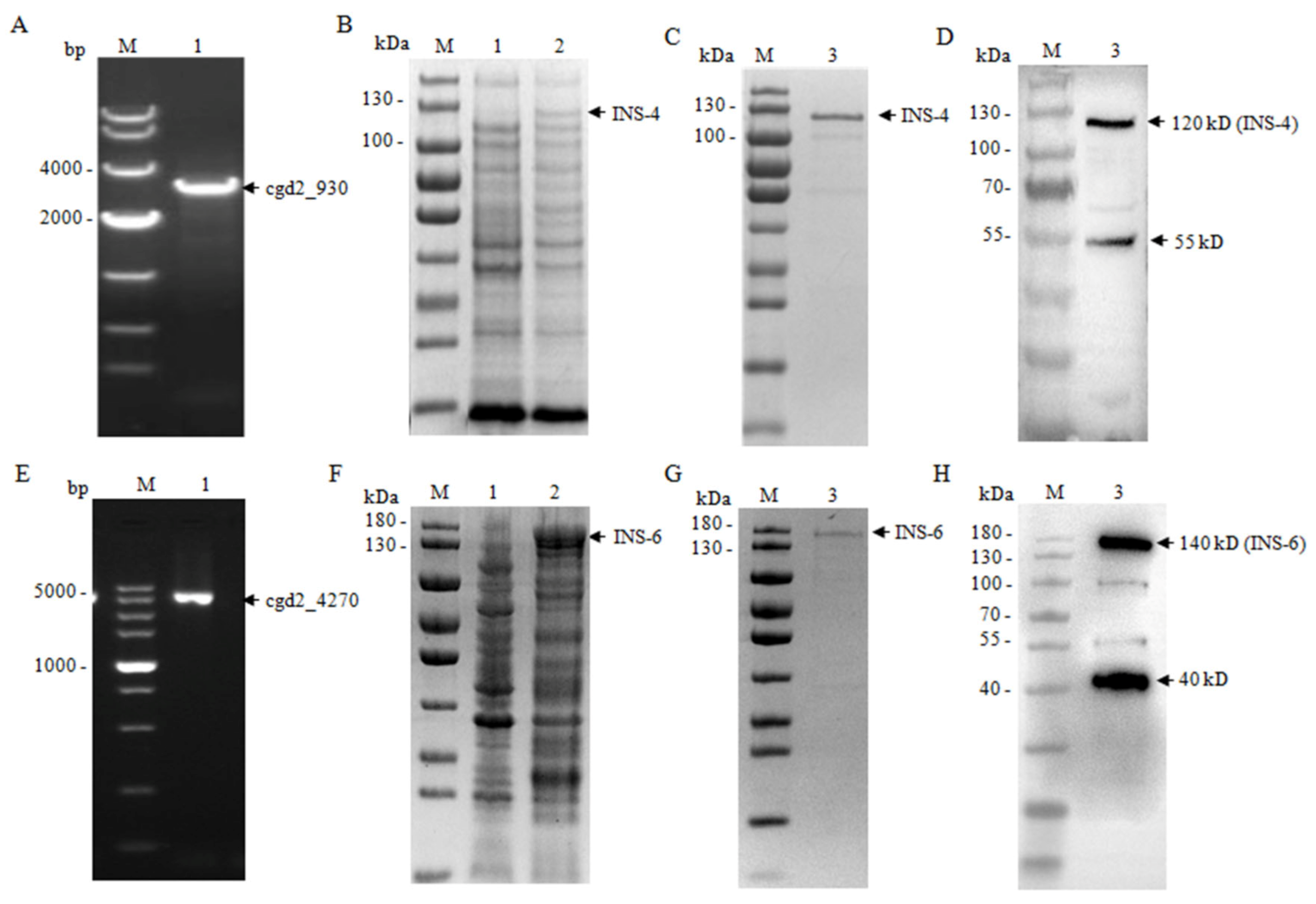
The cgd2_930 gene (amplicon = 3042 bp) was amplified from genomic DNA of the C. parvum IOWA isolate using primers 5′-CGGGATCCATGACAGAAATAA-3′ (the BamH1 restriction site underlined) and 5′-CCGCTCGAGTATAGTTATGTTAAG-3′ (the Xhol restriction site underlined), while the coding sequence of the cgd2_4270 gene without the putative signal peptide (amplicon = 3672 bp) was amplified using primers 5′-GCGTCGACAGTATTTGGGACCGAATTC-3′ (the Sal1 restriction site underlined) and 5′-TTGCGGCCGCAAGCTTGCTATGATCGCAGGA-3′ (the Not1 restriction site underlined). The PCR was performed in a T100TM Thermal Cycler (Bio-Rad, California, USA) under the following cycling conditions: 98 °C for 30 s; 30 cycles of 98 °C for 10 s, 52 °C 10 s, and 72 °C 2 min; and 72 °C for 5 min. The PCR products were inserted into the pET28a vector (Novagen, Madison, WI, USA), which was transformed into Escherichia coli DH5α cells. Positive colonies were identified by PCR, with the accuracy of cloned genes being verified using DNA sequence analysis.
2.3. Expression and Purification of CpINS-4 and CpINS-6
The recombinant plasmid containing the correct sequence of the cgd2_930 or cgd2_4270 gene was transformed into BL21-Star (DE3) pLysS Competent cells (Weidi Biotechnology, Shanghai, China), which were cultured in LB medium containing 100 mg/mL kanamycin. The expression of recombinant proteins was induced at 25 °C for 12 h by adding 0.5 mM isopropyl-β-D-thiogalactoside (IPTG).
For the purification of the recombinant protein, the cultured E. coli cells were harvested by centrifugation and resuspended in buffer A (50 mM sodium phosphate, 300 mM NaCI, pH7.4). The cell suspension was frozen-and-thawed three times and lysed by sonication at 4 °C. The lysate was centrifuged and the pellet containing the recombinant protein was dissolved in buffer B (50 mM sodium phosphate, 8 M urea, 40 mM imidazole, 300 mM NaCI, pH7.4) supplemented with a protease inhibitor cocktail (Sigma-Aldrich, St. Louis, MS, USA). The supernatant was collected by centrifugation and filtered through a 0.45-µm filter, and loaded onto histidine-tagged protein purification resins (Ni Sepharose 6 Fast Flow, GE Healthcare, Madison, WI, USA) at 4 °C and 70 rpm for 1 h. The resins were washed with 10 volumes buffer B and eluted with buffer C (50 mM sodium phosphate, 8 M urea, 200 mM imidazole, 300 mM NaCI, and pH7.4). The urea was removed by dialyzed against buffer A, and concentrated by using Amicon® Ultra-15 30 K Centrifugal Filter Devices (Millipore, Billerica, MA, USA). The purity of the recombinant protein was evaluated using SDS-PAGE. The INS identity of the recombinant proteins was verified using MALDI-TOF-MS analysis (Sangon Biotech, Shanghai, China).
2.4. Preparation of Polyclonal Antibodies against CpINS-4 and CpINS-6
Polyclonal antibodies against CpINS-4 and CpINS-6 were raised in specific pathogen-free rabbits as described previously [
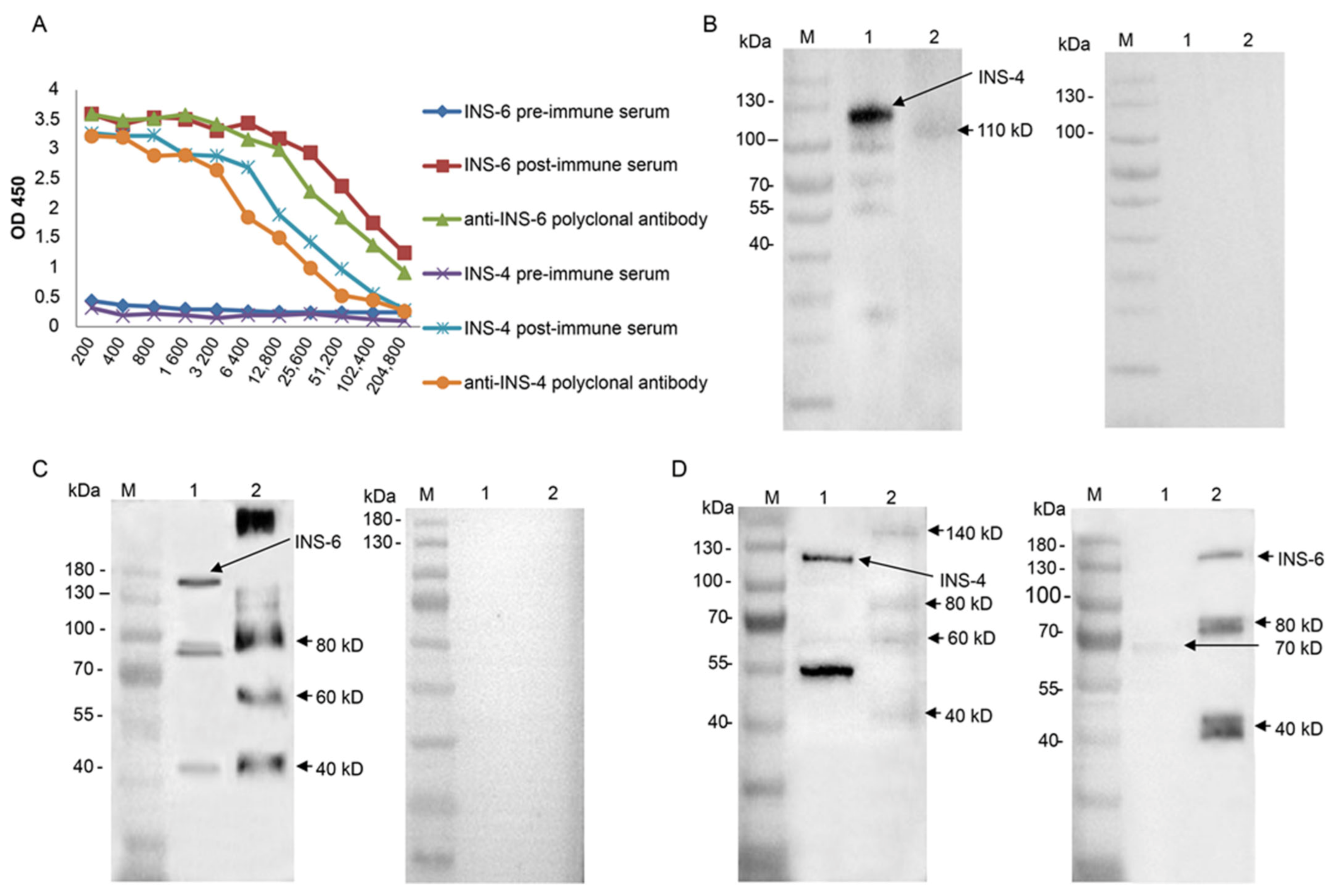
16]. Briefly, rabbits were immunized subcutaneously with 250 μg of the recombinant protein. Preimmune sera prior to the primary immunization were collected as negative controls. Boost immunizations were conducted twice at 2-week intervals using 250 μg purified protein. Postimmune sera were collected from the immunized animals seven days after the last immunization. Immunoglobulin G (IgG) was isolated from pooled immune sera using protein G affinity chromatography (GE Healthcare, Madison, WI, USA). The titer and specificity of polyclonal antibodies were evaluated using ELISA and Western blot, respectively.
2.5. Assessment of Cross-Reactivity of Anti-CpINS-4 and Anti-CpINS-6 Antibodies
To assess the cross-reactivity of anti-CpINS-4 and anti-CpINS-6 antibodies, recombinant CpINS-4 and CpINS-6 were separated by SDS-PAGE and transferred onto PVDF membranes. After blocking with 5% nonfat milk-PBST at 4 °C overnight, the membranes were incubated with anti-CpINS-4 or anti-CpINS-6 antibodies (1:1000) for 1 h, with goat-anti-rabbit IgG Antibody (Sigma-Aldrich) (1:5000) being used as the secondary antibody. The membranes were washed three times with PBST, treated with High-sig ECL Western Blotting Substrate (Tanon, Shanghai, China), and analyzed with a Tanon 5200 chemiluminescence imaging system.
2.6. Western Blot Analysis of Native CpINS-4 and CpINS-6
To identify native CpINS-4 and CpINS-6, hypochlorite-treated oocysts (1 × 107) were excysted as described above. The sporozoites were collected and resuspended in the RIPA Lysis and Extraction Buffer (ThermoFisher, Waltham, MA, USA) with Protease Inhibitor Cocktail (Sigma-Aldrich). After incubation at 4 °C overnight, the supernatant was collected, mixed with protein-loading buffer, and boiled for 10 min. The lysates from sporozoites were separated by SDS-PAGE and transferred onto a PVDF membrane. The membrane was processed further with antiserum (1:1000) and preimmune serum (1:1000) as described above.
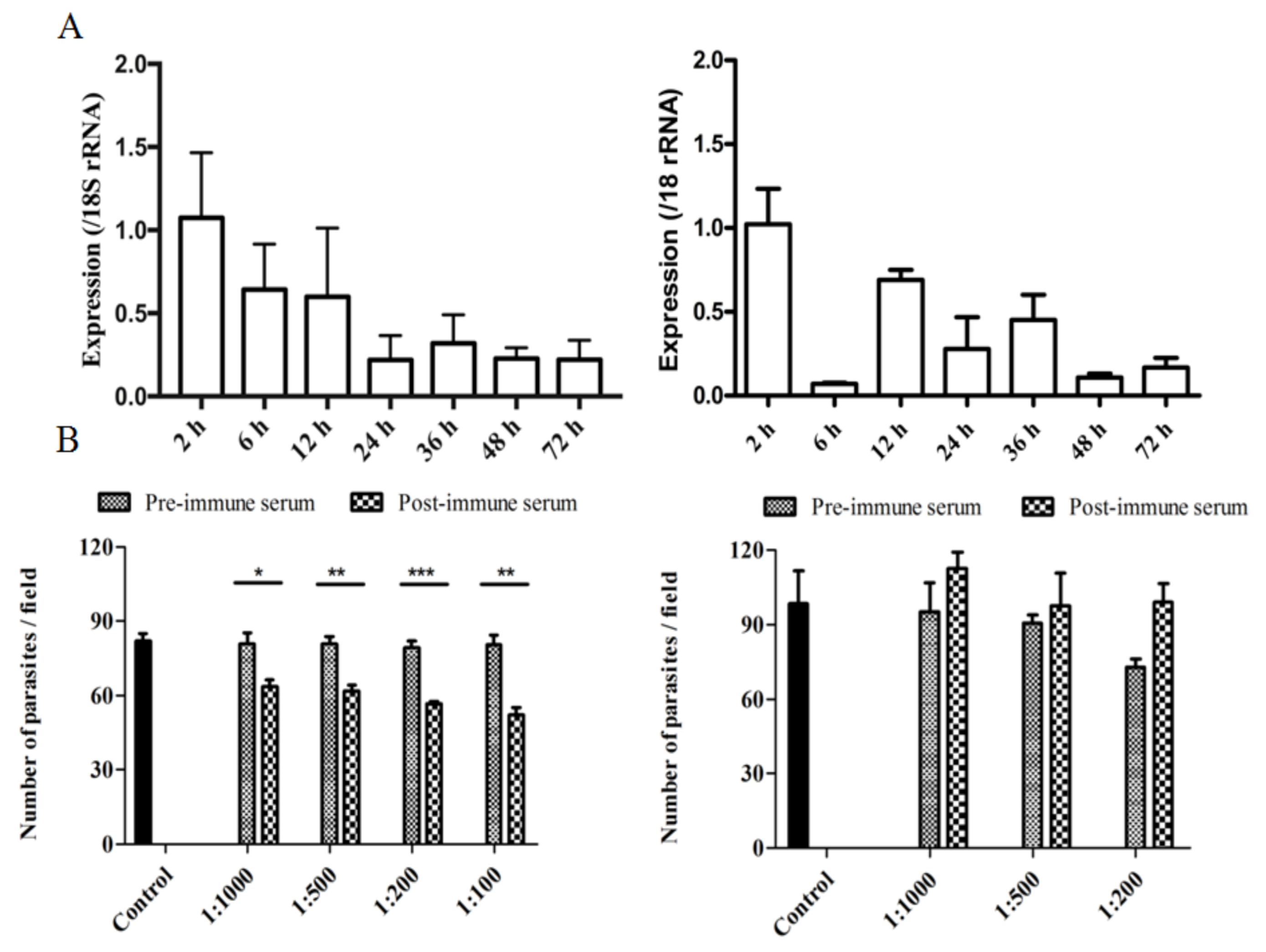
2.7. Assessment of cgd2_930 and cgd2_4270 Gene Expression in C. parvum
HCT-8 monolayers were inoculated with hypochlorite-treated oocysts and RNA was isolated from the infected HCT-8 cultures at 2–72 h using the RNeasy Mini Kit (QIAGEN, Germantown, Maryland, USA). Afterwards, cDNA was synthesized from the RNA using the GoScript Reverse Transcription System (Promega, Beijing, China). The expression of the
cgd2_930 or
cgd2_4270 gene was assessed using qPCR, with data from the 18S rRNA gene being used in data normalization. The qPCR was performed in a 20-μL reaction containing 0.1 mM primers, 1 µL of cDNA and 10 µL of SYBR Green PCR Mix (TOYOBO, Osaka, Japan) in a Light Cycler 480 (Roche, Basel, Switzerland), with the following cycling condition: 95 °C for 3 min and 45 cycles of 95 °C for 30 s, 58 °C for 30 s, and 72 °C for 30 s. The primers used included 5′-TCCAGAAGATGGTGCTCTTG-3′ and 5′-CTGGCCCTTCATGTCCTAAA-3′ for the
cgd2_930 gene, 5′-GCTCACTTCCTAACTCCACCAG-3′ and 5′-ACCATCTTGCCACTCTGTTCTT-3′ for the
cgd2_4270 gene, and 5′-CTCCACCAACTAAGAACGGCC-3′ and 5′-TAGAGATTGGAGGTTGTTCCT-3′ for the
18s rRNA gene [
15]. The relative expression levels of
cgd2_930 and
cgd2_4270 genes in various developmental stages were calculated using the delta-delta method [
21].
2.8. Immunofluorescence Assay (IFA)
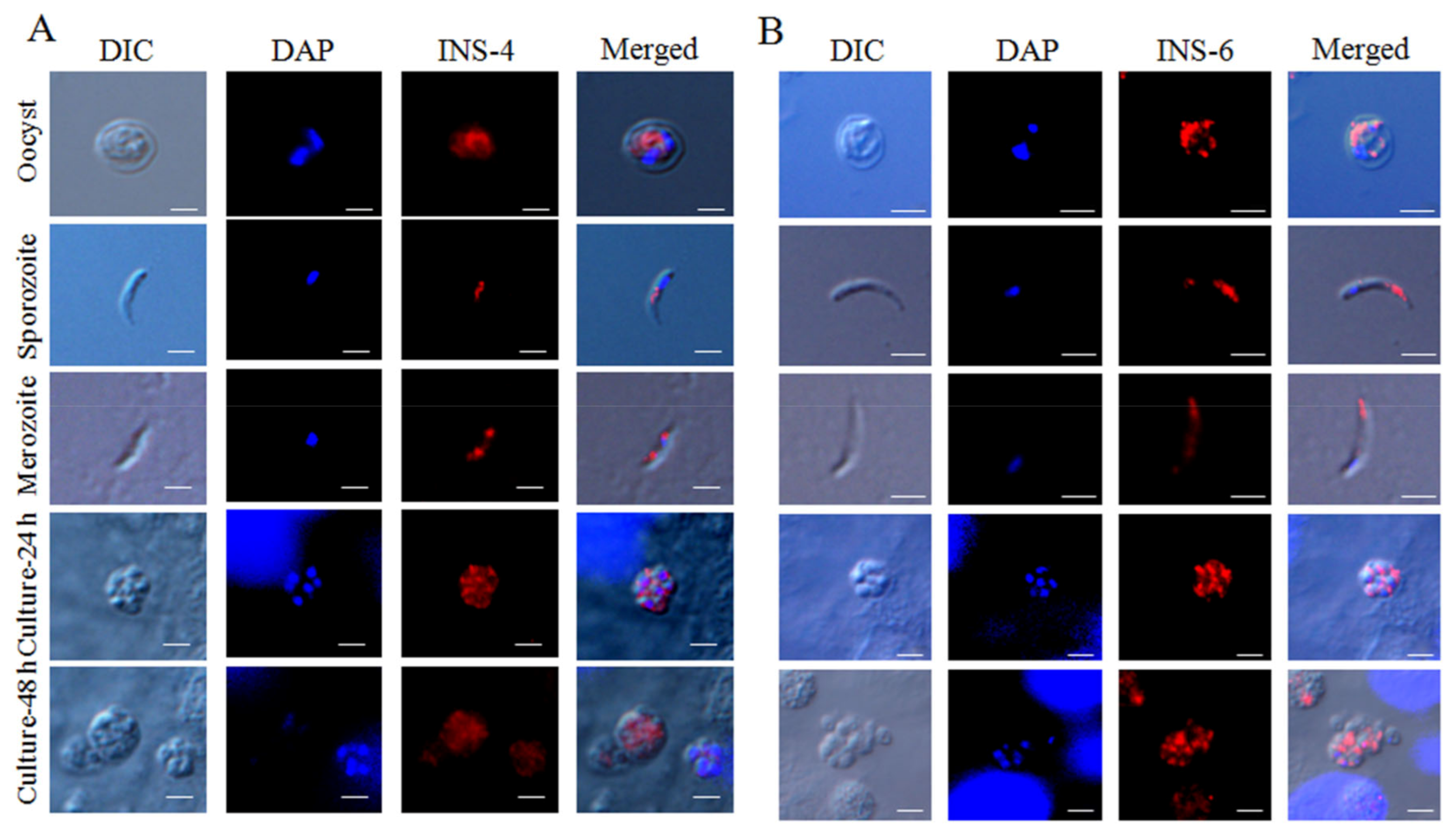
For the assessment of CpINS-4 and CpINS-6 expression in developmental stages of C. parvum, oocysts and sporozoites were suspended in PBS and dried onto poly-L-lysine-treated microscope slides (Waterborne, New Orleans, LA, USA), whereas HCT-8 cells infected with sporozoites were grown on coverslips for 24 and 48 h, fixed with methanol for 15 min, and permeabilized with 0.5% Triton X-100 for 30 min. After blocking with 5% BSA at room temperature for 1 h, oocysts, sporozoites and infected HCT-8 cells were probed overnight with the polyclonal antibodies (1:500) in 5% BSA/PBS (weight/volume), followed by staining with Alexa Fluor 594-conjugated goat-anti-rabbit IgG (Cell Signaling Technology, Danvers, MA, USA) and nuclear staining with 4′, 6-diamidino-2-phenylindole (DAPI, Sigma-Aldrich). After three washes with PBS, the slides and coverslips were mounted with Antifade Mounting Medium (Booster, Wuhan, China) and examined using differential interference contrast (DIC) and fluorescence microscopy on a BX53 microscope (Olympus, Tokyo, Japan).
2.9. Invasion Neutralization Assay
The effect of polyclonal antibodies against CpINS-4 and CpINS-6 on
C. parvum infection of HCT-8 cells was examined using an in vitro neutralization assay. Hypochlorite-treated oocysts were preincubated at 37 °C in antiserum or preimmune serum diluted with culture medium for 15 min, with the culture medium only as the negative control. They were inoculated into HCT-8 cell culture grown in coverslips in 24-well plates at ~90% confluence as described above. After 2-h incubation, residual oocysts were washed off the monolayers with fresh culture medium. After additional cultivation for 24 h, the monolayers were fixed with methanol and stained with Cy3-labeled Sporo-Glo antibodies (Waterborne, New Orleans, LA, USA). The coverslips were examined by immunofluorescence microscopy. Images of 50 random microscope fields per coverslip were captured under 200× magnification and analyzed using ImageJ (
https://imagej.nih.gov/ij/download.html, accessed on 20 December 2020). The percent of invasion inhibition was calculated using the following formula: (1 − [No. of parasites after postimmune serum treatment/No. of parasites after preimmune serum treatment]) × 100%. Three independent experiments were conducted for the determination of percent invasion inhibition by the antibodies alone or in combination.
4. Discussion
The data from the present study suggest that CpINS-4 and CpINS-6 have very different structures and are expressed in different organelles of
C. parvum, thus may play different roles in the invasion and growth of the pathogen. CpINS-4 has all four conserved domains in functional M16 proteases, while a peptidase M16 inactive domain is absent in CpINS-6. Furthermore, there is only 15.2% identity in amino acids between CpINS-4 and CpINS-6. The differences in functional domains and amino acid sequences have probably led to completely different tertiary structures of the two INS. Like other M16A proteases subfamily, CpINS-4 and CpINS-6 both adopt a clamshell structure, with the N- and C-terminal domains forming a hydrolytically active central cavity (
Figure 1B). The opening and closing of the N- and C-terminal domains are required to facilitate the access of the substrate to the hydrolytically active site and represent a rate-limiting step within the INS catalytic mechanism [
10], while the absence of a peptidase M16 inactive domain in C-terminal domains of CpINS-6 may directly affect the function of the C-terminal domains and clamshell structure switch, thereby changing the catalytic activity on the substrate.
The recombinant CpINS-4 and CpINS-6 appear to be processed proteolytically. Multiple products were observed in
E. coli-expressed recombinant INS proteins and native CpINS-4 and CpINS-6. It is possible that both CpINS-4 and CpINS-6 sequences contain proteolytic cleavage sites. In sequence analysis, CpINS-4 has eight putative SΦX(E/D) cleavage sites with the sequences SPED (265–268), SFSD (543–546), SALD (973–976), SIME (261–264), SVIE (361–364), SLDE (401–404), SIPE (454–457), and SFFE (659–662), which in theory can lead to the hydrolysis of the full-CpINS-4 into several fragments. Similarly, CpINS-6 has four putative SΦX (E/D) cleavage sites, with the sequences SIWD (23–26), SVSE (197–200), SIFE (334–337), SVSD (401–404). These cleavage sites may be responsible for the proteolytic processing of CpINS-4 and CpINS-6 in a way similar to the post-translational processing of rhoptry proteins in
T. gondii. In
T. gondii, the N-terminal prodomain of ROP1 is processed at its SΦXE cleavage site [
22], while the proteolytical processing of TLN1 occurs at the cleavage site SΦXD [
13]. TLN1 is a rhoptry metalloprotease and targeted to the rhoptry via an N-terminal prodomain, which is processed at SΦX (E/D) cleavage sites. After proteolytic processing and removing the organelle-targeting sequences in toxolysins, the C-terminal domain is associated tightly with the N-terminal domain, forming homo-oligomers. The proteolytic processing may be an evolutionary adaptation in order to alter the substrate of TLN1 [
13]. Two other rhoptry proteins that are involved in virulence of
T. gondii, ROP17 and ROP18, are also predicted to have several SΦX (E/D) cleavage sites [
23,
24].
CpINS-6 may be present in a high molecular weight complex in
C. parvum. In the analysis of native CpINS-4 and CpINS-6, the polyclonal antibodies against CpINS-6 reacted with a protein with a higher molecular weight than 180 kDa in lysates of
C. parvum. As a secreted protein, CpINS-6 could initially exist as a large precursor, which is processed into multiple fragments within the secretory system of the parasite. In
T. gondii, TLN4 is initially synthesized as a large (~260 kDa) precursor, and subsequently processed into multiple fragments that remain associated with each other in a large molecular complex [
12]. The large protein could also be a mixed dimer. Insulin proteases are known to be active as a homodimer containing one active subunit and another subunit that binds substrate with normal affinity [
25]. In addition, the C-terminal domain is also required for dimerization [
9], suggesting that C-terminal domain might play an important role in the formation of a high molecular weight complex of CpINS-6.
CpINS-4 and CpINS-6 may have classic INS activities. In the INS of C. parvum, CpINS-4 has four conserved domains, with the N-terminal domain containing the inverted Zn2+-binding motif HFLEH in amino acids 69–73. Although CpINS-6 may be not a classic insulinlike protease with all four conserved domains, its N-terminal domain does contain the inverted Zn2+-binding motif HLMEH in amino acids 126–130. The inverted Zn2+-binding motif is a key feature of M16 proteases, is required for catalytic activity, and affects substrate binding. Further studies should be conducted to determine whether CpINS-4 and CpINS-6 both have proteolytic activities associated with classic insulinaselike protease.
CpINS-4 could be involved in the invasion process of
C. parvum. Various proteases and protein kinases are secreted by the secretory organelles of apicomplexans parasites to counteract the host immune responses [
6]. In agreement with its potential role in the invasion of host cells, CpINS-4 is expressed in both sporozoites and merozoites, the invasion stages of the parasites, mainly in the middle region. Previously, based on sequence homology, CpINS-4 was indicated as a putative rhoptry protein [
26]. While the protein appears to be processed in the same way as some rhoptry proteins in
T. gondii, its expression pattern in both sporozoites and merozoites revealed by immunofluorescence is more in line with a location in dense granules. The early expression level of the gene encoding it during the in vitro culture and the partial neutralization effect of antibodies against it support the role of CpINS-4 in host cell invasion. Compared with CpINS-4, the location of CpINS-6 expression appears different; its expression in both spozoites and merozoites is confined to apical end, more in line with a rhoptry or microneme location. While the
cgd2_4270 gene encoding CpINS-6 was the highest at 2 h of in vitro culture, antibodies against it failed in neutralizing the invasion of host cells by sporozoites.
In conclusion, the preliminary study represents our initial attempt in characterizing the functions of CpINS-4 and CpINS-6 in C. parvum. Although the two INS both have the active Zn-chelating “HXXEH” motif and similar post-transcriptional processing, they appear to be very different in structure, are expressed in different organelles, and play different roles in the invasion and growth of the parasites. Further studies using more advanced tools are needed to understand their precise locations and functions in Cryptosporidium spp.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




