
Comparison of In Vitro Killing Activity of Rezafungin, Anidulafungin, Caspofungin, and Micafungin against Four Candida auris Clades in RPMI-1640 in the Absence and Presence of Human Serum

 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolates
2.2. Antifungal Susceptibility Testing
2.3. Time-Kill Studies
2.4. Phase-Contrast Microscopy
3. Results
3.1. MIC Values of the Echinocandins against C. auris
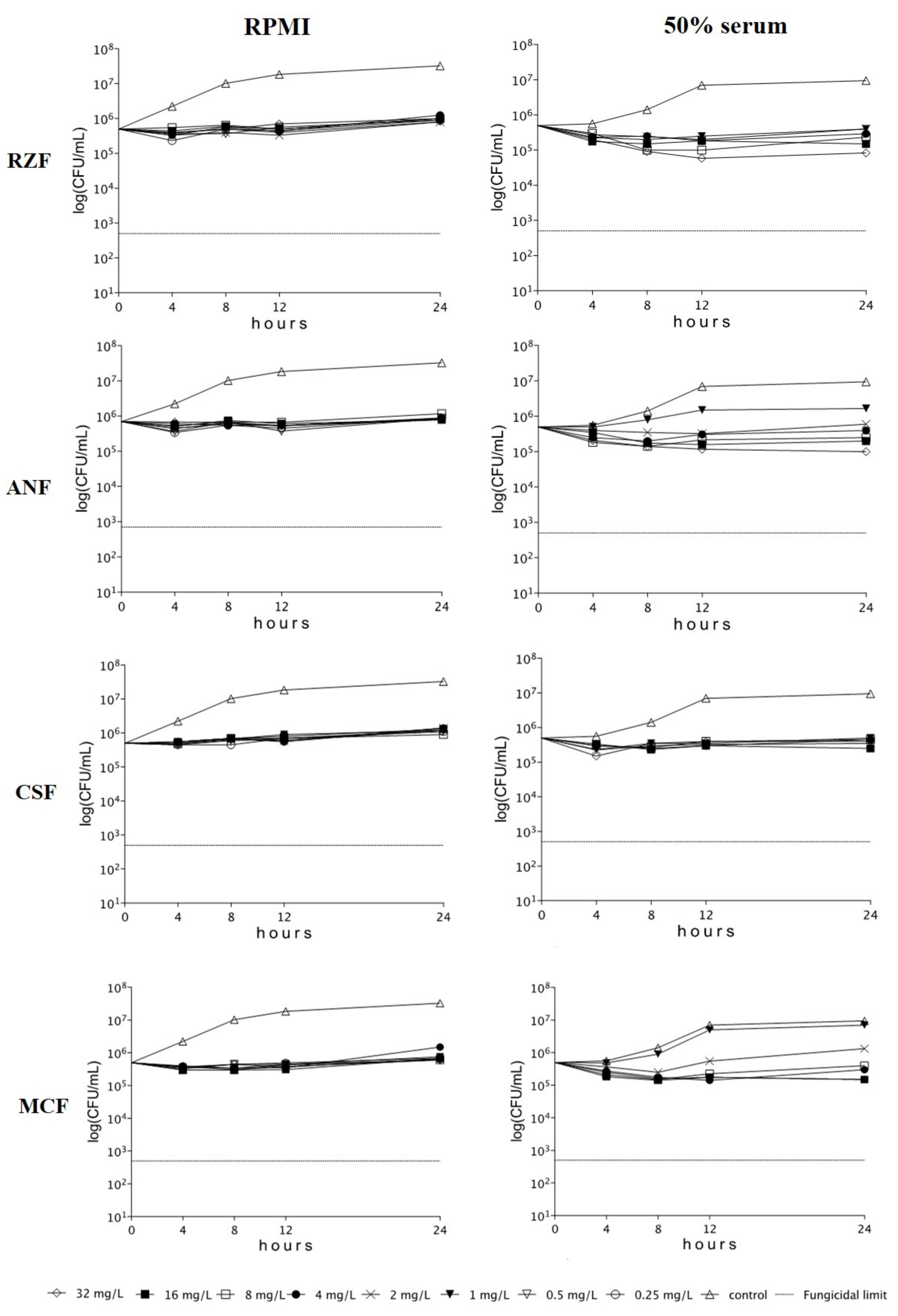
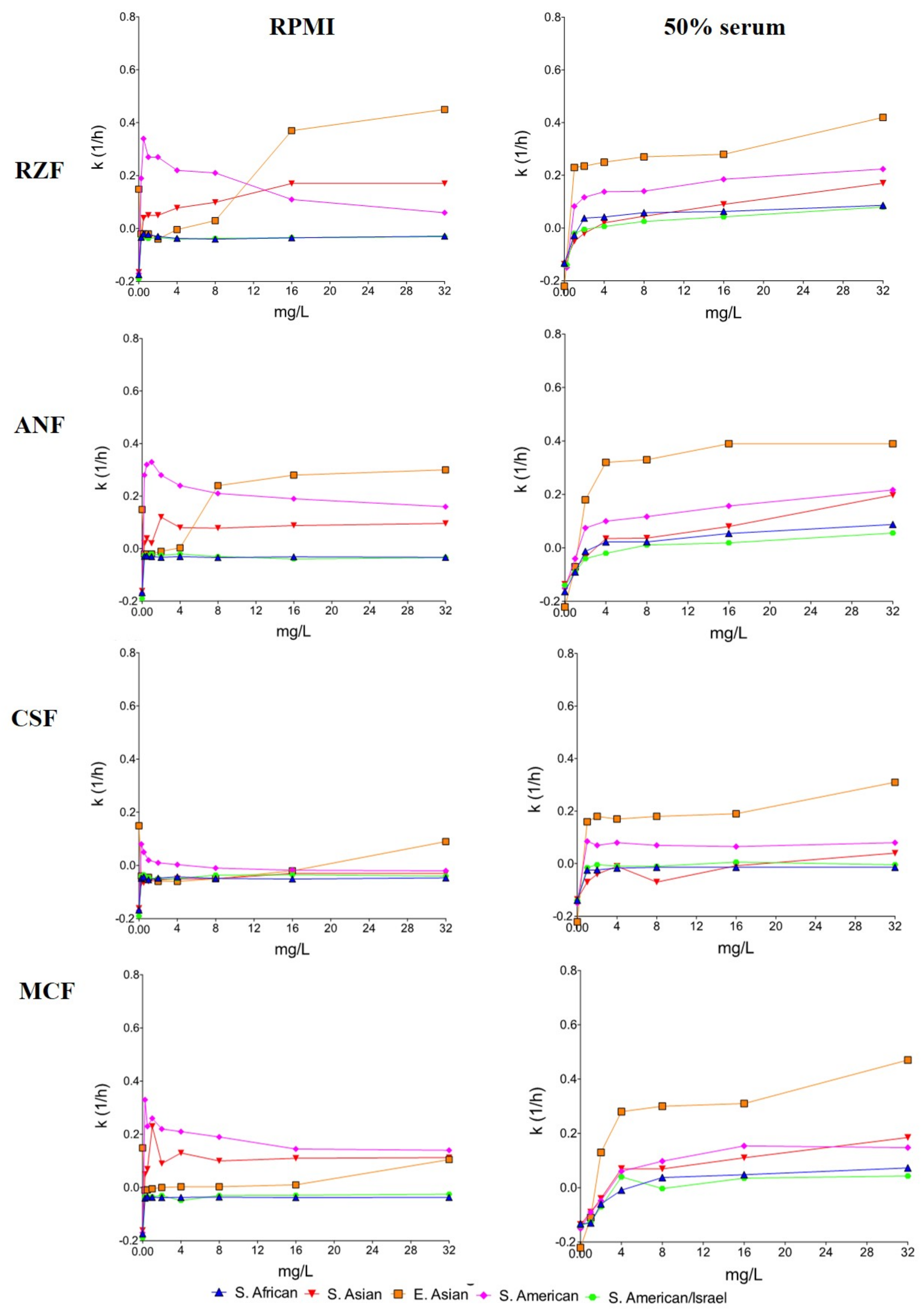
3.2. Time-Kill Studies
3.3. South Asian Clade
3.4. East Asian Clade
3.5. South African Clade
3.6. South American Clade
3.7. Phase-Contrast Microscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sekyere, J.O. Candida Auris: A Systematic Review and Meta-Analysis of Current Updates on an Emerging Multidrug-Resistant Pathogen. Microbiol. Open 2018, 7, e00578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A. Simultaneous Emergence of Multidrug-Resistant Candida Auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, N.A.; de Groot, T.; Badali, H.; Abastabar, M.; Chiller, T.M.; Meis, J.F. Potential fifth Clade of Candida Auris, Iran, 2018. Emerg. Infect. Dis. 2019, 25, 1780–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borman, A.M.; Szekely, A.; Johnson, E.M. Comparative Pathogenicity of United Kingdom Isolates of the Emerging Pathogen Candida auris and Other Key Pathogenic Candida Species. mSphere 2016, 1, e00189-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szekely, A.; Borman, A.M.; Johnson, E.M. Candida Auris Isolates of the Southern Asian and South African Lineages Exhibit Different Phenotypic and Antifungal Susceptibility Profiles In Vitro. J. Clin. Microbiol. 2019, 57, e02055-18. [Google Scholar] [CrossRef] [Green Version]
- Forgács, L.; Borman, A.; Prépost, E.; Tóth, Z.; Kardos, G.; Kovács, R.; Szekely, A.; Nagy, F.; Kovacs, I.; Majoros, L. Comparison of In Vivo Pathogenicity of Four Candida Auris Clades in a Neutropenic Bloodstream Infection Murine Model. Emerg. Microb. Infect. 2020, 9, 1160–1169. [Google Scholar] [CrossRef]
- Hernando-Ortiz, A.; Mateo, E.; Perez-Rodriguez, A.; de Groot, P.W.J.; Quindós, G.; Eraso, E. Virulence of Candida Auris from Different Clinical Origins in Caenorhab-Ditis Elegans and Galleria Mellonella Host Models. Virulence 2021, 12, 1063–1075. [Google Scholar] [CrossRef]
- Chowdhary, A.; Tarai, B.; Singh, A.; Sharma, A. Multidrug-Resistant Candida auris Infections in Critically Ill Coronavirus Disease Patients, India, April–July 2020. Emerg. Infect. Dis. 2020, 26, 2694–2696. [Google Scholar] [CrossRef]
- CDC. Available online: https://www.cdc.gov/fungal/candida-auris/c-auris-antifungal.html (accessed on 22 March 2021).
- Perlin, D.S. Echinocandin Resistance, Susceptibility Testing and Prophylaxis: Implications for Patient Management. Drugs 2014, 74, 1573–1585. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Perez, W.B.; Jiménez-Ortigosa, C.; Hough, G.; Locke, J.B.; Ong, V.; Bartizal, K.; Perlin, D.S. CD101: A Novel Long-Acting Echinocandin. Cell. Microbiol. 2016, 18, 1308–1316. [Google Scholar] [CrossRef] [Green Version]
- Sandison, T.; Ong, V.; Lee, J.; Thye, D. Safety and Pharmacokinetics of CD101 IV, a Novel Echinocandin, in Healthy Adults. Antimicrob. Agents Chemother. 2017, 61, 01627-16. [Google Scholar] [CrossRef] [Green Version]
- Sofjan, A.K.; Mitchell, A.; Shah, D.N.; Nguyen, T.; Sim, M.; Trojcak, A.; Beyda, N.D.; Garey, K.W. Rezafungin (CD101), a next-generation echinocandin: A systematic literature review and assessment of possible place in therapy. J. Glob. Antimicrob. Resist. 2018, 14, 58–64. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Carvalhaes, C.; Messer, S.A.; Rhomberg, P.R.; Castanheira, M. Activity of a Long-Acting Echinocandin, Rezafungin, and Comparator Antifungal Agents Tested against Contemporary Invasive Fungal Isolates (SENTRY Program, 2016 to 2018). Antimicrob. Agents Chemother. 2020, 64, 00099-20. [Google Scholar] [CrossRef] [Green Version]
- Tóth, Z.; Forgács, L.; Locke, J.B.; Kardos, G.; Nagy, F.; Kovács, R.; Szekely, A.; Borman, A.; Majoros, L. In vitro activity of rezafungin against common and rare Candida species and Saccharomyces cerevisiae. J. Antimicrob. Chemother. 2019, 74, 3505–3510. [Google Scholar] [CrossRef] [Green Version]
- Ong, V.; Hough, G.; Schlosser, M.; Bartizal, K.; Balkovec, J.M.; James, K.D.; Krishnan, B.R. Preclinical Evaluation of the Stability, Safety, and Efficacy of CD101, a Novel Echinocandin. Antimicrob. Agents Chemother. 2016, 60, 6872–6879. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts. Approved Standard-Third Edition M27-A3; Clinical and Laboratory Standard Institute: Wayne, PA, USA, 2008. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antifungal Susceptibility Testing of Yeasts, 1st ed.; M60Ed1; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Domán, M.; Kovács, R.L.; Perlin, D.S.; Kardos, G.; Gesztelyi, R.; Juhász, B.; Bozó, A.; Majoros, L. Dose Escalation Studies with Caspofungin against Candida Glabrata. J. Med Microbiol. 2015, 64, 998–1007. [Google Scholar] [CrossRef]
- Brüggemann, R.J.M.; Middel-Baars, V.; De Lange, D.W.; Colbers, A.; Girbes, A.R.J.; Pickkers, P.; Swart, E.L. Pharmacokinetics of Anidulafungin in Critically Ill Intensive Care Unit Patients with Suspected or Proven Invasive Fungal Infections. Antimicrob. Agents Chemother. 2016, 61, 01894-16. [Google Scholar] [CrossRef] [Green Version]
- Muilwijk, E.W.; Schouten, J.A.; Van Leeuwen, H.J.; Van Zanten, A.R.H.; De Lange, D.W.; Colbers, A.; Verweij, P.E.; Burger, D.M.; Pickkers, P.; Brüggemann, R.J.M. Pharmacokinetics of caspofungin in ICU patients. J. Antimicrob. Chemother. 2014, 69, 3294–3299. [Google Scholar] [CrossRef] [Green Version]
- Boonstra, J.M.; Van Der Elst, K.C.; Veringa, A.; Jongedijk, E.M.; Brüggemann, R.J.; Koster, R.A.; Kampinga, G.A.; Kosterink, J.G.; Van Der Werf, T.S.; Zijlstra, J.G.; et al. Pharmacokinetic Properties of Micafungin in Critically Ill Patients Diagnosed with Invasive Candidiasis. Antimicrob. Agents Chemother. 2017, 61, e01398-17. [Google Scholar] [CrossRef] [Green Version]
- Dudiuk, C.; Berrio, I.; Leonardelli, F.; Morales-Lopez, S.; Theill, L.; Macedo, D.; Yesid-Rodriguez, J.; Salcedo, S.; Marin, A.; Gamarra, S.; et al. Antifungal activity and killing kinetics of anidulafungin, caspofungin and amphotericin B against Candida auris. J. Antimicrob. Chemother. 2019, 74, 2295–2302. [Google Scholar] [CrossRef]
- Kordalewska, M.; Lee, A.; Park, S.; Berrio, I.; Chowdhary, A.; Zhao, Y.; Perlin, D.S. Understanding Echinocandin Resistance in the Emerging Pathogen Candida auris. Antimicrob. Agents Chemother. 2018, 62, e00238-18. [Google Scholar] [CrossRef] [Green Version]
- Tóth, Z.; Forgács, L.; Kardos, T.; Kovács, R.; Locke, J.B.; Kardos, G.; Nagy, F.; Borman, A.M.; Adnan, A.; Majoros, L. Relative Frequency of Paradoxical Growth and Trailing Effect with Caspofungin, Micafungin, Anidulafungin, and the Novel Echinocandin Rezafungin against Candida Species. J. Fungi 2020, 6, 136. [Google Scholar] [CrossRef]
- Perlin, D.S. Cell Wall-Modifying Antifungal Drugs. Curr. Top. Microbiol. Immunol. 2019, 425, 255–275. [Google Scholar] [CrossRef]
- Navarro-Arias, M.J.; Hernández-Chávez, M.J.; Garcia-Carnero, L.C.; Amezcua-Hernández, D.G.; Lozoya-Pérez, N.E.; Estrada-Mata, E.; Martínez-Duncker, I.; Franco, B.; Mora-Montes, H.M. Differential Recognition of Candida Tropicalis, Candida Guilliermondii, Candida Krusei, and Candida Auris by Human Innate Immune Cells. Infect. Drug Resist. 2019, 12, 783–794. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Paul, R.A.; Chakrabarti, A.; Bhattacharya, S.; Soman, R.; Shankarnarayan, S.A.; Chavan, D.; Singh, S.; Das, P.; Kaur, H.; et al. Caspofungin Resistance in Candia Auris due to Mutations in Fks1 with Adjunctive Role of Chitin and Key Cell Wall Stress Response Pathway Genes. bioRxiv 2020, 196600. [Google Scholar] [CrossRef]
- Lepak, A.J.; Zhao, M.; Berkow, E.L.; Lockhart, S.R.; Andes, D.R. Pharmacodynamic Optimization for Treatment of Invasive Candida auris Infection. Antimicrob. Agents Chemother. 2017, 61, 00791-17. [Google Scholar] [CrossRef] [Green Version]
- Lepak, A.J.; Zhao, M.; Andes, D.R. Pharmacodynamic Evaluation of Rezafungin (CD101) against Candida auris in the Neutropenic Mouse Invasive Candidiasis Model. Antimicrob. Agents Chemother. 2018, 62, 01572-18. [Google Scholar] [CrossRef] [Green Version]
- Caballero, U.; Kim, S.; Eraso, E.; Quindós, G.; Vozmediano, V.; Schmidt, S.; Jauregizar, N. In Vitro Synergistic Interactions of Isavuconazole and Echinocandins against Candida auris. Antibiotics 2021, 10, 355. [Google Scholar] [CrossRef]
- Nagy, F.; Tóth, Z.; Nyikos, F.; Forgács, L.; Jakab, Á.; Borman, A.M.; Majoros, L.; Kovács, R. In Vitro and In Vivo Interaction of Caspofungin with Isavuconazole against Candida Auris Planktonic Cells and Biofilms. bioRxiv 2021, 434267. [Google Scholar] [CrossRef]
- Villanueva-Lozano, H.; Treviño-Rangel, R.D.J.; González, G.M.; Ramírez-Elizondo, M.T.; Lara-Medrano, R.; Aleman-Bocanegra, M.C.; Guajardo-Lara, C.E.; Gaona-Chávez, N.; Castilleja-Leal, F.; Torre-Amione, G.; et al. Outbreak of Candida auris infection in a COVID-19 hospital in Mexico. Clin. Microbiol. Infect. 2021. [Google Scholar] [CrossRef]
- Flanagan, S.; Goodman, D.B.; Jandourek, A.; O’Reilly, T.; Sandison, T. Lack of Effect of Rezafungin on QT/QTc Interval in Healthy Subjects. Clin. Pharmacol. Drug Dev. 2019, 9, 456–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Isolates Number | Clade | Body Site | MIC Values in RPMI/RPMI + 50% Serum (mg/L) | |||
|---|---|---|---|---|---|---|
| RZF | ANF | CSF | MCF | |||
| 12 | South Asian | unknown | 0.03–0.06/0.5 | 0.12/2 | 1/0.5–1 | 0.12–0.25/2 |
| 20 (NCPF 8985) | South Asian | Wound | 0.25/0.5 | 0.25/1 | 1/2 | 0.25/2 |
| 27 (NCPF 89891) | South Asian | Pleural fluid | 0.12/1 | 0.12/2 | 0.5/2 | 0.25/2 |
| Type strain (NCPF 13029 = CBS 10913) | East Asian | External ear | 0.06/0.5 | 0.03/0.5–1 | 0.25/0.5 | 0.12/2–4 |
| 15 (NCPF 8984) | East Asian | External ear | 0.03/0.25–0.5 | 0.03–0.06/2 | 0.5–1/1 | 0.12/2 |
| 12372 (CBS 12372) | East Asian | Blood | 0.06/0.5 | 0.03/0.5–1 | 0.12/0.5 | 0.03/2 |
| 204 | South African | Tracheostomy | 0.06/0.25-0.5 | 0.03/2 | 0.25/0.5–1 | 0.12–0.25/2 |
| 2 (NCPF 8977) | South African | Cerebrospinal fluid | 0.12/1 | 0.03/2 | 0.5/1 | 0.25/2–4 |
| 185 | South African | Blood | 0.12/1 | 0.25/2 | 0.25/1 | 0.25/4 |
| I-24 | South American (Israel) | Blood | 0.25/0.5 | 0.06/4 | 0.25/1 | 0.12/4 |
| I-172 | South American (Israel) | Blood | 0.25/0.5 | 0.06/2 | 0.25/1 | 0.12/2 |
| 13108 (CDC B-13108) | South American (Colombia) | Hospital environment | 0.12/0.5 | 0.06/2 | 0.25/1 | 0.12/4 |
| 16565 (CDC B-16565) | South American (Colombia) | Hospital environment | 0.12/0.25 | 0.015/0.5 | 0.25/0.5 | 0.06/2 |
| 50% Serum | Clade | Drug | Time (Hours) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 32 mg/L | 16 mg/L | 8 mg/L | 4 mg/L | 2 mg/L | 1 mg/L | 0.5 mg/L | 0.25 mg/L | |||
| (−) | South Asian (3 isolates) | RZF | 1.77 | 1.77 | 3 | 3.75 | 1.36 | 1.09 | 1.23 | 1.19 |
| ANF | 3.1 | 3.4 | 3.4 | 2.14 | 2.14 | 3.51 | 1.22 | 1.82 | ||
| MCF | 1.54 | 1.62 | 1.56 | 1.27 | 1.58 | 0.75 | 2 | 2.06 | ||
| CSF | NA | NA | NA | NA | NA | NA | NA | NA | ||
| East Asian (2 isolates) | RZF | 0.67 | 0.81 | 10 | 23 | NA | 15.5 | 15.3 | 16.4 | |
| ANF | 1 | 1.07 | 1.25 | 30.1 | 24.6 | 19.3 | 21.6 | 13.8 | ||
| MCF | 1.3 | 6.95 | 1.56 | 13.6 | 18 | 31.6 | 69.7 | NA | ||
| CSF | 1.38 | NA | NA | NA | NA | NA | NA | NA | ||
| South African (3 isolates) | RZF | NA | NA | NA | NA | NA | NA | NA | NA | |
| ANF | NA | NA | NA | NA | NA | NA | NA | NA | ||
| MCF | NA | NA | NA | NA | NA | NA | NA | NA | ||
| CSF | NA | NA | NA | NA | NA | NA | NA | NA | ||
| South American (2 isolates from Israel) | RZF | NA | NA | NA | NA | NA | NA | NA | NA | |
| ANF | NA | NA | NA | NA | NA | NA | NA | NA | ||
| MCF | NA | NA | NA | NA | NA | NA | NA | NA | ||
| CSF | NA | NA | NA | NA | NA | NA | NA | NA | ||
| South American (2 isolates from Colombia) | RZF | 5 | 2.7 | 1.43 | 1.36 | 1.11 | 1.11 | 0.88 | 1.58 | |
| ANF | 1.58 | 1.42 | 1.15 | 1.08 | 1.5 | 0.91 | 0.71 | 1.25 | ||
| MCF | 2.14 | 2.07 | 1.58 | 1.43 | 1.36 | 1.15 | 1.3 | 0.91 | ||
| CSF | NA | NA | NA | 100.3 | 29.4 | 15 | 6 | 3.75 | ||
| (+) | South Asian (3 isolates) | RZF | 1.77 | 3.34 | 6.69 | 15.1 | 75.3 | NA | ND | ND |
| ANF | 1.5 | 3.75 | 7.5 | 7.5 | NA | NA | ND | ND | ||
| MCF | 1.58 | 2.73 | 4.29 | 4.29 | NA | NA | ND | ND | ||
| CSF | 3.75 | 75 | NA | NA | NA | NA | ND | ND | ||
| East Asian (2 isolates) | RZF | 0.71 | 1.08 | 1.11 | 1.2 | 1.25 | 1.3 | ND | ND | |
| ANF | 0.77 | 0.77 | 0.91 | 0.94 | 1.67 | NA | ND | ND | ||
| MCF | 0.64 | 0.97 | 1 | 1.07 | 0.97 | NA | ND | ND | ||
| CSF | 0.97 | 1.58 | 1.97 | 1.77 | 1.97 | 1.88 | ND | ND | ||
| South African (3 isolates) | RZF | 3.42 | 3.81 | 4.3 | 1.85 | 1.88 | 100 | ND | ND | |
| ANF | 2.04 | 3.01 | 2.08 | 2.48 | 6.53 | NA | ND | ND | ||
| MCF | 2.73 | 3.75 | 4.29 | 7.3 | NA | NA | ND | ND | ||
| CSF | 17.2 | 43.8 | NA | NA | 21.1 | NA | ND | ND | ||
| South American (2 isolates from Israel) | RZF | 3.76 | 7.07 | 10.03 | 12.57 | 38.2 | 39.2 | ND | ND | |
| ANF | 5.38 | 7.17 | 7.61 | 80.1 | NA | NA | ND | ND | ||
| MCF | 7 | 4.27 | 24.6 | 9.56 | NA | NA | ND | ND | ||
| CSF | NA | 17.3 | NA | NA | 67.3 | NA | ND | ND | ||
| South American (2 isolates from Colombia) | RZF | 1.37 | 1.63 | 2.14 | 2.17 | 2.57 | 3.64 | ND | ND | |
| ANF | 1.39 | 1.92 | 2.57 | 1.41 | 1.89 | 15.1 | ND | ND | ||
| MCF | 2.04 | 1.96 | 3.08 | 1.76 | 45.5 | NA | ND | ND | ||
| CSF | 1.81 | 2.15 | 1.95 | 1.78 | 2.01 | 1.74 | ND | ND | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovács, R.; Tóth, Z.; Locke, J.B.; Forgács, L.; Kardos, G.; Nagy, F.; Borman, A.M.; Majoros, L. Comparison of In Vitro Killing Activity of Rezafungin, Anidulafungin, Caspofungin, and Micafungin against Four Candida auris Clades in RPMI-1640 in the Absence and Presence of Human Serum. Microorganisms 2021, 9, 863. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040863
Kovács R, Tóth Z, Locke JB, Forgács L, Kardos G, Nagy F, Borman AM, Majoros L. Comparison of In Vitro Killing Activity of Rezafungin, Anidulafungin, Caspofungin, and Micafungin against Four Candida auris Clades in RPMI-1640 in the Absence and Presence of Human Serum. Microorganisms. 2021; 9(4):863. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040863
Chicago/Turabian StyleKovács, Renátó, Zoltán Tóth, Jeffrey B. Locke, Lajos Forgács, Gábor Kardos, Fruzsina Nagy, Andrew M. Borman, and László Majoros. 2021. "Comparison of In Vitro Killing Activity of Rezafungin, Anidulafungin, Caspofungin, and Micafungin against Four Candida auris Clades in RPMI-1640 in the Absence and Presence of Human Serum" Microorganisms 9, no. 4: 863. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040863