
Diverse Genotypes and Species of Cryptosporidium in Wild Rodent Species from the West Coast of the USA and Implications for Raw Produce Safety and Microbial Water Quality

Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Cryptosporidium Sequences of Wild Rodents from GenBank
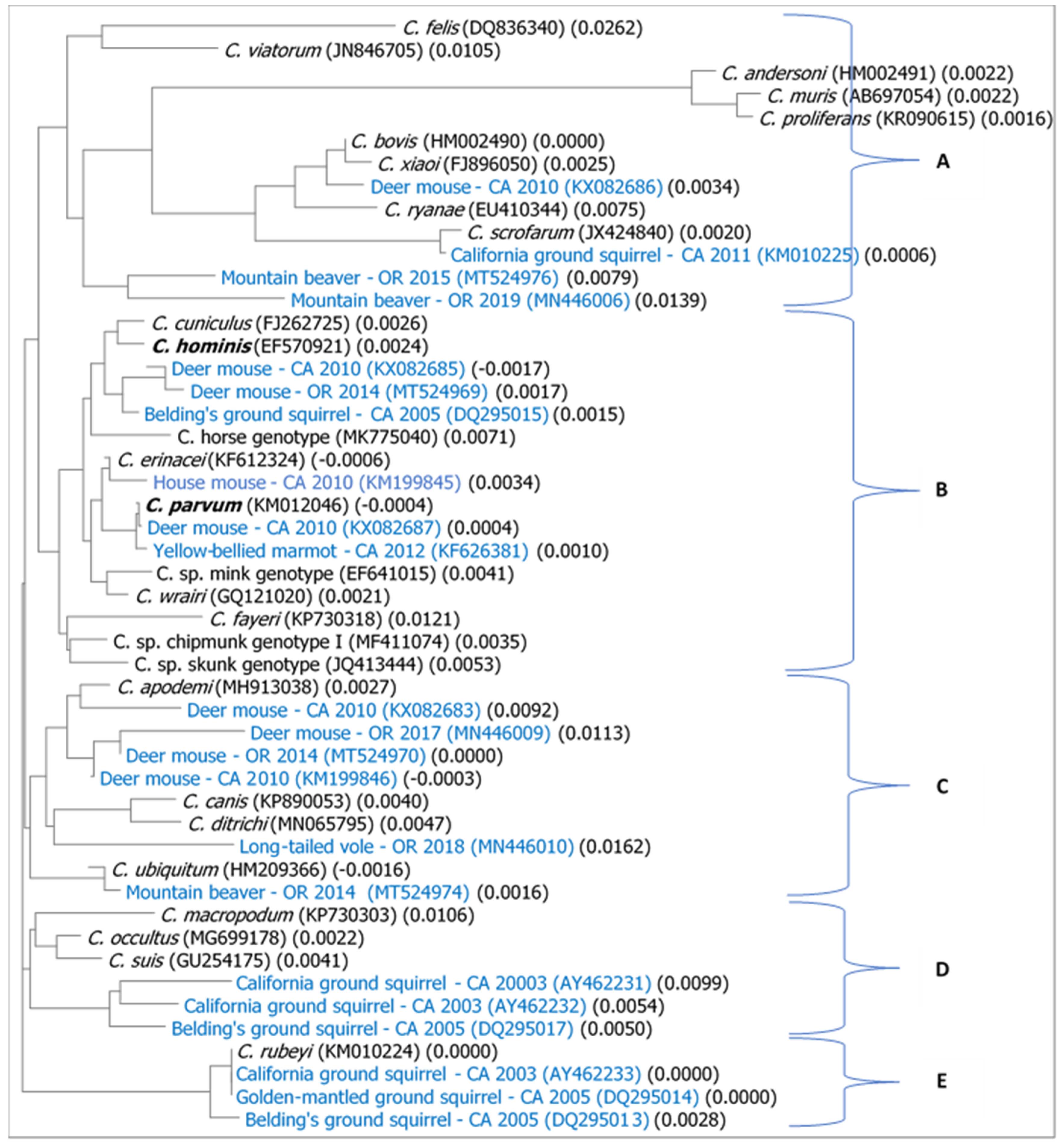
2.2. Phylogenetic Analysis
2.3. BLAST Analysis
3. Results
3.1. Diverse Genotypes of Relatively Recent Cryptosporidium Isolates from Wild Rodents throughout the West Coast, US
3.2. Phylogenetic Relationships between Relatively Recent Cryptosporidium Isolates from Wild Rodents from the West Coast and Representative Cryptosporidium Species and Zoonotic Genotypes
3.3. Comparison of Cryptosporidium from Wild Rodents with Cryptosporidium Species and Genotypes Deposited in GenBank
4. Discussion
4.1. The Selection of Representative Sequences of Cryptosporidium from Rodents
4.2. Diversity of Cryptosporidium in Wild Rodents on the West Coast, US
4.3. Water Quality and Produce Safety Implications from Zoonotic Cryptosporidium in Rodents
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fayer, R. Taxonomy and Species Delimitation in Cryptosporidium. Exp. Parasitol. 2010, 124, 90–97. [Google Scholar] [CrossRef]
- Garcia-R, J.C.; Hayman, D.T.S. Origin of a Major Infectious Disease in Vertebrates: The Timing of Cryptosporidium Evolution and Its Hosts. Parasitology 2016, 143, 1683–1690. [Google Scholar] [CrossRef] [Green Version]
- Jezkova, J.; Horcickova, M.; Hlaskova, L.; Sak, B.; Kvetonova, D.; Novak, J.; Hofmannova, L.; McEvoy, J.; Kvac, M. Cryptosporidium Testudinis Sp. n., Cryptosporidium Ducismarci Traversa, 2010 and Cryptosporidium Tortoise Genotype III (Apicomplexa: Cryptosporidiidae) in Tortoises. Folia Parasitol. 2016, 63. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, A.; Paparini, A.; Jian, F.; Robertson, I.; Ryan, U. Public Health Significance of Zoonotic Cryptosporidium Species in Wildlife: Critical Insights into Better Drinking Water Management. Int. J. Parasitol. Parasites Wildl. 2016, 5, 88–109. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, A.; Monis, P.; Aucote, S.; King, B.; Paparini, A.; Jian, F.; Yang, R.; Oskam, C.; Ball, A.; Robertson, I.; et al. Zoonotic Cryptosporidium Species in Animals Inhabiting Sydney Water Catchments. PLoS ONE 2016, 11, e0168169. [Google Scholar] [CrossRef] [Green Version]
- Čondlová, Š.; Horčičková, M.; Sak, B.; Květoňová, D.; Hlásková, L.; Konečný, R.; Stanko, M.; McEvoy, J.; Kváč, M. Cryptosporidium Apodemi Sp. n. and Cryptosporidium Ditrichi Sp. n. (Apicomplexa: Cryptosporidiidae) in Apodemus Spp. Eur. J. Protistol. 2018, 63, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kváč, M.; Vlnatá, G.; Ježková, J.; Horčičková, M.; Konečný, R.; Hlásková, L.; McEvoy, J.; Sak, B. Cryptosporidium Occultus Sp. n. (Apicomplexa: Cryptosporidiidae) in Rats. Eur. J. Protistol. 2018, 63, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Holubová, N.; Zikmundová, V.; Limpouchová, Z.; Sak, B.; Konečný, R.; Hlásková, L.; Rajský, D.; Kopacz, Z.; McEvoy, J.; Kváč, M. Cryptosporidium Proventriculi Sp. n. (Apicomplexa: Cryptosporidiidae) in Psittaciformes Birds. Eur. J. Protistol. 2019, 69, 70–87. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Hijjawi, N. New Developments in Cryptosporidium Research. Int. J. Parasitol. 2015, 45, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Ryan, U.; Zahedi, A.; Paparini, A. Cryptosporidium in Humans and Animals-a One Health Approach to Prophylaxis. Parasite Immunol. 2016, 38, 535–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Fayer, R.; Ryan, U.; Upton, S.J. Cryptosporidium Taxonomy: Recent Advances and Implications for Public Health. Clin. Microbiol. Rev. 2004, 17, 72–97. [Google Scholar] [CrossRef] [Green Version]
- Capizzi, D.; Bertolino, S.; Mortelliti, A. Rating the Rat: Global Patterns and Research Priorities in Impacts and Management of Rodent Pests: Rating the Rat. Mammal. Rev. 2014, 44, 148–162. [Google Scholar] [CrossRef]
- Catzeflis, F.M. North American Rodents. Status Survey and Conservation Action Plan. Biol. Conserv. 2000, 95, 119. [Google Scholar] [CrossRef]
- Jones, J.K.; Hall, E.R. The Mammals of North America. 2nd Ed. John Wiley and Sons, New York, 1:Xv + 1-600 + 90 and 2:Vi + 601-1181 + 90, Illustrated. 1981. J. Mammal. 1982, 63, 717. [Google Scholar] [CrossRef]
- Kilonzo, C.; Li, X.; Vivas, E.J.; Jay-Russell, M.T.; Fernandez, K.L.; Atwill, E.R. Fecal Shedding of Zoonotic Food-Borne Pathogens by Wild Rodents in a Major Agricultural Region of the Central California Coast. Appl. Environ. Microbiol. 2013, 79, 6337–6344. [Google Scholar] [CrossRef] [Green Version]
- Meerburg, B.G.; Singleton, G.R.; Kijlstra, A. Rodent-Borne Diseases and Their Risks for Public Health. Crit. Rev. Microbiol. 2009, 35, 221–270. [Google Scholar] [CrossRef] [PubMed]
- Luis, A.D.; Hayman, D.T.S.; O’Shea, T.J.; Cryan, P.M.; Gilbert, A.T.; Pulliam, J.R.C.; Mills, J.N.; Timonin, M.E.; Willis, C.K.R.; Cunningham, A.A.; et al. A Comparison of Bats and Rodents as Reservoirs of Zoonotic Viruses: Are Bats Special? Proc. Biol. Sci. 2013, 280, 20122753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckle, A.; Smith, R. (Eds.) Rodent Pests and Their Control, 2nd ed.; CABI: Wallingford, UK, 2014; ISBN 978-1-84593-817-8. [Google Scholar]
- Wei, X.; Hou, S.; Pan, X.; Xu, C.; Li, J.; Yu, H.; Chase, J.; Atwill, E.R.; Li, X.; Chen, K.; et al. Microbiological Contamination of Strawberries from U-Pick Farms in Guangzhou, China. Int. J. Environ. Res. Public. Health 2019, 16, 4910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilonzo, C.; Li, X.; Vodoz, T.; Xiao, C.; Chase, J.A.; Jay-Russell, M.T.; Vivas, E.J.; Atwill, E.R. Quantitative Shedding of Multiple Genotypes of Cryptosporidium and Giardia by Deer Mice (Peromyscus Maniculatus) in a Major Agricultural Region on the California Central Coast. J. Food Prot. 2017, 819–828. [Google Scholar] [CrossRef]
- Dahmana, H.; Granjon, L.; Diagne, C.; Davoust, B.; Fenollar, F.; Mediannikov, O. Rodents as Hosts of Pathogens and Related Zoonotic Disease Risk. Pathogens 2020, 9, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atwill, E.R.; Camargo, S.M.; Phillips, R.; Alonso, L.H.; Tate, K.W.; Jensen, W.A.; Bennet, J.; Little, S.; Salmon, T.P. Quantitative Shedding of Two Genotypes of Cryptosporidium Parvum in California Ground Squirrels (Spermophilus Beecheyi). Appl. Environ. Microbiol. 2001, 67, 2840–2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das Pereira, M.G.C.; Li, X.; McCowan, B.; Phillips, R.L.; Atwill, E.R. Multiple Unique Cryptosporidium Isolates from Three Species of Ground Squirrels (Spermophilus Beecheyi, S. Beldingi, and S. Lateralis) in California. Appl. Environ. Microbiol. 2010, 76, 8269–8276. [Google Scholar] [CrossRef] [Green Version]
- Atwill, E.R.; Phillips, R.; Pereira, M.D.G.C.; Li, X.; McCowan, B. Seasonal Shedding of Multiple Cryptosporidium Genotypes in California Ground Squirrels (Spermophilus Beecheyi). Appl. Environ. Microbiol. 2004, 70, 6748–6752. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; das Pereira, M.G.C.; Larsen, R.; Xiao, C.; Phillips, R.; Striby, K.; McCowan, B.; Atwill, E.R. Cryptosporidium Rubeyi n. Sp. (Apicomplexa: Cryptosporidiidae) in Multiple Spermophilus Ground Squirrel Species. Int. J. Parasitol. Parasites Wildl. 2015, 4, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montecino-Latorre, D.; Li, X.; Xiao, C.; Atwill, E.R. Elevation and Vegetation Determine Cryptosporidium Oocyst Shedding by Yellow-Bellied Marmots (Marmota Flaviventris) in the Sierra Nevada Mountains. Int. J. Parasitol. Parasites Wildl. 2015, 4, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Nguyen, T.; Xiao, C.; Levy, A.; Akagi, Y.; Silkie, S.; Atwill, E.R. Prevalence and Genotypes of Cryptosporidium in Wildlife Populations Co-Located in a Protected Watershed in the Pacific Northwest, 2013 to 2016. Microorganisms 2020, 8, 914. [Google Scholar] [CrossRef] [PubMed]
- Gaydos, J.K.; Miller, W.A.; Gilardi, K.V.K.; Melli, A.; Schwantje, H.; Engelstoft, C.; Fritz, H.; Conrad, P.A. Cryptosporidium and Giardia in Marine-Foraging River Otters (Lontra Canadensis) from the Puget Sound Georgia Basin Ecosystem. J. Parasitol. 2007, 93, 198–202. [Google Scholar] [CrossRef]
- Ruecker, N.J.; Hoffman, R.M.; Chalmers, R.M.; Neumann, N.F. Detection and Resolution of Cryptosporidium Species and Species Mixtures by Genus-Specific Nested PCR-Restriction Fragment Length Polymorphism Analysis, Direct Sequencing, and Cloning. Appl. Environ. Microbiol. 2011, 77, 3998–4007. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Yang, C.; Xiao, L. PCR-Mediated Recombination between Cryptosporidium Spp. of Lizards and Snakes. J. Eukaryot. Microbiol. 2003, 50, 563–565. [Google Scholar] [CrossRef]
- Prystajecky, N.; Huck, P.M.; Schreier, H.; Isaac-Renton, J.L. Assessment of Giardia and Cryptosporidium Spp. as a Microbial Source Tracking Tool for Surface Water: Application in a Mixed-Use Watershed. Appl. Environ. Microbiol. 2014, 80, 2328–2336. [Google Scholar] [CrossRef] [Green Version]
- Mac Kenzie, W.R.; Hoxie, N.J.; Proctor, M.E.; Gradus, M.S.; Blair, K.A.; Peterson, D.E.; Kazmierczak, J.J.; Addiss, D.G.; Fox, K.R.; Rose, J.B. A Massive Outbreak in Milwaukee of Cryptosporidium Infection Transmitted through the Public Water Supply. N. Engl. J. Med. 1994, 331, 161–167. [Google Scholar] [CrossRef] [PubMed]
- DeSilva, M.B.; Schafer, S.; Kendall Scott, M.; Robinson, B.; Hills, A.; Buser, G.L.; Salis, K.; Gargano, J.; Yoder, J.; Hill, V.; et al. Communitywide Cryptosporidiosis Outbreak Associated with a Surface Water-Supplied Municipal Water System—Baker City, Oregon, 2013. Epidemiol. Infect. 2016, 144, 274–284. [Google Scholar] [CrossRef]
- Van Dyke, M.I.; Ong, C.S.L.; Prystajecky, N.A.; Isaac-Renton, J.L.; Huck, P.M. Identifying Host Sources, Human Health Risk and Indicators of Cryptosporidium and Giardia in a Canadian Watershed Influenced by Urban and Rural Activities. J. Water Health 2012, 10, 311–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, C.; Moorehead, W.; Ross, A.; Isaac-Renton, J. Studies of Giardia Spp. and Cryptosporidium Spp. in Two Adjacent Watersheds. Appl. Environ. Microbiol. 1996, 62, 2798–2805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnet, J.-B.; Penny, C.; Ogorzaly, L.; Cauchie, H.-M. Spatial and Temporal Distribution of Cryptosporidium and Giardia in a Drinking Water Resource: Implications for Monitoring and Risk Assessment. Sci. Total Environ. 2014, 472, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Lemarchand, K.; Lebaron, P. Occurrence of Salmonella Spp and Cryptosporidium Spp in a French Coastal Watershed: Relationship with Fecal Indicators. FEMS Microbiol. Lett. 2003, 218, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Chen, P.; Villegas, E.N.; Landy, R.B.; Kanetsky, C.; Cama, V.; Dearen, T.; Schultz, C.L.; Orndorff, K.G.; Prelewicz, G.J.; et al. Cryptosporidium Source Tracking in the Potomac River Watershed. Appl. Environ. Microbiol. 2008, 74, 6495–6504. [Google Scholar] [CrossRef] [Green Version]
- Jellison, K.L.; Hemond, H.F.; Schauer, D.B. Sources and Species of Cryptosporidium Oocysts in the Wachusett Reservoir Watershed. Appl. Environ. Microbiol. 2002, 68, 569–575. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Alderisio, K.A.; Xiao, L. Distribution of Cryptosporidium Genotypes in Storm Event Water Samples from Three Watersheds in New York. Appl. Environ. Microbiol. 2005, 71, 4446–4454. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Chase, J.A.; Bond, R.F.; Lor, P.; Fernandez, K.; Nguyen, T.H.; Partyka, M.L.; Thiptara, A.; Atwill, E.R. Microbiological Safety of Popular Recreation Swimming Sites in Central California. Environ. Monit. Assess. 2019, 191, 456. [Google Scholar] [CrossRef]
- Hansen, J.S.; Ongerth, J.E. Effects of Time and Watershed Characteristics on the Concentration of Cryptosporidium Oocysts in River Water. Appl. Environ. Microbiol. 1991, 57, 2790–2795. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, B.G.; Mazurek, J.M.; Hlavsa, M.; Park, J.; Tillapaw, M.; Parrish, M.; Salehi, E.; Franks, W.; Koch, E.; Smith, F.; et al. Cryptosporidiosis Associated with Ozonated Apple Cider. Emerg. Infect. Dis. 2006, 12, 684–686. [Google Scholar] [CrossRef]
- Robertson, L.J.; Temesgen, T.T.; Tysnes, K.R.; Eikås, J.E. An Apple a Day: An Outbreak of Cryptosporidiosis in Norway Associated with Self-Pressed Apple Juice. Epidemiol. Infect. 2019, 147, e139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millard, P.S.; Gensheimer, K.F.; Addiss, D.G.; Sosin, D.M.; Beckett, G.A.; Houck-Jankoski, A.; Hudson, A. An Outbreak of Cryptosporidiosis from Fresh-Pressed Apple Cider. JAMA 1994, 272, 1592–1596. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.P.; Caldart, E.T.; Freire, R.L.; Mitsuka-Breganó, R.; de Freitas, F.M.; Miura, A.C.; Mareze, M.; Martins, F.D.C.; Urbano, M.R.; Seifert, A.L.; et al. The Effect of Water Source and Soil Supplementation on Parasite Contamination in Organic Vegetable Gardens. Rev. Bras. Parasitol. Vet. 2018, 27, 327–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Shi, K.; Sun, F.; Li, T.; Wang, R.; Zhang, S.; Jian, F.; Ning, C.; Zhang, L. Identification of Human Pathogenic Enterocytozoon Bieneusi, Cyclospora Cayetanensis, and Cryptosporidium Parvum on the Surfaces of Vegetables and Fruits in Henan, China. Int. J. Food Microbiol. 2019, 307, 108292. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.; Carazo, M.; Arias, M.L.; Chaves, C.; Monge, R.; Chinchilla, M. Prevalence of Cyclospora sp., Cryptosporidium sp, microsporidia and fecal coliform determination in fresh fruit and vegetables consumed in Costa Rica. Arch. Latinoam. Nutr. 2004, 54, 428–432. [Google Scholar]
- Duedu, K.O.; Yarnie, E.A.; Tetteh-Quarcoo, P.B.; Attah, S.K.; Donkor, E.S.; Ayeh-Kumi, P.F. A Comparative Survey of the Prevalence of Human Parasites Found in Fresh Vegetables Sold in Supermarkets and Open-Aired Markets in Accra, Ghana. BMC Res. Notes 2014, 7, 836. [Google Scholar] [CrossRef] [Green Version]
- Kudah, C.; Sovoe, S.; Baiden, F. Parasitic Contamination of Commonly Consumed Vegetables in Two Markets in Ghana. Ghana Med. J. 2018, 52, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Utaaker, K.S.; Kumar, A.; Joshi, H.; Chaudhary, S.; Robertson, L.J. Checking the Detail in Retail: Occurrence of Cryptosporidium and Giardia on Vegetables Sold across Different Counters in Chandigarh, India. Int. J. Food Microbiol. 2017, 263, 1–8. [Google Scholar] [CrossRef]
- Ranjbar-Bahadori, S.; Mostoophi, A.; Shemshadi, B. Study on Cryptosporidium Contamination in Vegetable Farms around Tehran. Trop. Biomed. 2013, 30, 193–198. [Google Scholar]
- Caradonna, T.; Marangi, M.; Del Chierico, F.; Ferrari, N.; Reddel, S.; Bracaglia, G.; Normanno, G.; Putignani, L.; Giangaspero, A. Detection and Prevalence of Protozoan Parasites in Ready-to-Eat Packaged Salads on Sale in Italy. Food Microbiol. 2017, 67, 67–75. [Google Scholar] [CrossRef]
- Hong, S.; Kim, K.; Yoon, S.; Park, W.-Y.; Sim, S.; Yu, J.-R. Detection of Cryptosporidium Parvum in Environmental Soil and Vegetables. J. Korean Med. Sci. 2014, 29, 1367–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, S.; Won, J.; Kim, J.-W.; Kim, K.; Park, W.-Y.; Yu, J.-R. Simultaneous Molecular Detection of Cryptosporidium and Cyclospora from Raw Vegetables in Korea. Korean J. Parasitol. 2017, 55, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Robertson, L.J.; Gjerde, B. Occurrence of Parasites on Fruits and Vegetables in Norway. J. Food Prot. 2001, 64, 1793–1798. [Google Scholar] [CrossRef] [PubMed]
- Ortega, Y.R.; Roxas, C.R.; Gilman, R.H.; Miller, N.J.; Cabrera, L.; Taquiri, C.; Sterling, C.R. Isolation of Cryptosporidium Parvum and Cyclospora Cayetanensis from Vegetables Collected in Markets of an Endemic Region in Peru. Am. J. Trop. Med. Hyg. 1997, 57, 683–686. [Google Scholar] [CrossRef] [PubMed]
- Rzezutka, A.; Nichols, R.A.B.; Connelly, L.; Kaupke, A.; Kozyra, I.; Cook, N.; Birrell, S.; Smith, H.V. Cryptosporidium Oocysts on Fresh Produce from Areas of High Livestock Production in Poland. Int. J. Food Microbiol. 2010, 139, 96–101. [Google Scholar] [CrossRef]
- Amorós, I.; Alonso, J.L.; Cuesta, G. Cryptosporidium Oocysts and Giardia Cysts on Salad Products Irrigated with Contaminated Water. J. Food Prot. 2010, 73, 1138–1140. [Google Scholar] [CrossRef]
- McKerr, C.; Adak, G.K.; Nichols, G.; Gorton, R.; Chalmers, R.M.; Kafatos, G.; Cosford, P.; Charlett, A.; Reacher, M.; Pollock, K.G.; et al. An Outbreak of Cryptosporidium Parvum across England & Scotland Associated with Consumption of Fresh Pre-Cut Salad Leaves, May 2012. PLoS ONE 2015, 10, e0125955. [Google Scholar] [CrossRef] [Green Version]
- Åberg, R.; Sjöman, M.; Hemminki, K.; Pirnes, A.; Räsänen, S.; Kalanti, A.; Pohjanvirta, T.; Caccio, S.M.; Pihlajasaari, A.; Toikkanen, S.; et al. Cryptosporidium Parvum Caused a Large Outbreak Linked to Frisée Salad in Finland, 2012. Zoonoses Public Health 2015, 62, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Atwill, E.R.; Chase, J.A.; Oryang, D.; Bond, R.F.; Koike, S.T.; Cahn, M.D.; Anderson, M.; Mokhtari, A.; Dennis, S. Transfer of Escherichia Coli O157:H7 from Simulated Wildlife Scat onto Romaine Lettuce during Foliar Irrigation. J. Food Prot. 2015, 78, 240–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellers, L.A.; Long, R.F.; Jay-Russell, M.T.; Li, X.; Atwill, E.R.; Engeman, R.M.; Baldwin, R.A. Impact of Field-Edge Habitat on Mammalian Wildlife Abundance, Distribution, and Vectored Foodborne Pathogens in Adjacent Crops. Crop. Prot. 2018, 108, 1–11. [Google Scholar] [CrossRef]
- Li, X.; Atwill, E.R.; Dunbar, L.A.; Tate, K.W. Effect of Daily Temperature Fluctuation during the Cool Season on the Infectivity of Cryptosporidium Parvum. Appl. Environ. Microbiol. 2010, 76, 989–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Cryptosporidium in Rodents in the West Coast (California, Oregon) | Highly Similar Sequences in GenBank by BLAST Analysis (Last Access at 31 December 2020) | ||||
|---|---|---|---|---|---|
| Rodent Host Species | Location/Year | GenBank Accession No. | Cryptosporidium Species and Isolates | GenBank Accession No. | Max. Identity (%) |
| Deer mouse (Peromyscus maniculatus) | CA, 2010 | KX082687 | Cryptosporidium parvum isolate 438–723 | MT071829 | 99.88 |
| Deer mouse (Peromyscus maniculatus) | CA, 2010 | KX082686 | Cryptosporidium xiaoi isolate AH S1 | MH049731 | 99.75 |
| Deer mouse (Peromyscus maniculatus) | CA, 2010 | KX082685 | Cryptosporidium sp. isolate Deer mouse 2014_PNWR6C | MT524969 | 100 |
| Deer mouse (Peromyscus maniculatus) | CA, 2010 | KX082683 | Cryptosporidium sp. isolate 1848-Pero-NA | KY644646 | 99.15 |
| Deer mouse (Peromyscus maniculatus) | CA, 2010 | KM199846 | Cryptosporidium sp. deer mouse genotype IV (W3) isolate CRY1811 | JQ413348 | 99.75 |
| Deer mouse (Peromyscus maniculatus) | OR, 2014 | MT524969 | Cryptosporidium sp. ex Peromyscus maniculatus isolate 2951 | KX082685 | 100 |
| Deer mouse (Peromyscus maniculatus) | OR, 2017 | MN446009 | Cryptosporidium sp. isolate Deer mouse 2014_PNW1052 | MT524970 | 98.87 |
| Deer mouse (Peromyscus maniculatus) | OR, 2014 | MT524970 | Cryptosporidium sp. deer mouse genotype IV (W3) isolate CRY1811 | JQ413348 | 99.75 |
| House mouse (Mus musculus) | CA, 2010 | KM199845 | Cryptosporidium mouse genotype isolate J46 | EU553589 | 100 |
| Yellow-bellied marmot (Marmota flaviventris) | CA, 2012 | KF626381 | Cryptosporidium parvum isolate Swec402 | KU892559 | 100 |
| Long-tailed vole (Microtus longicaudus) | OR, 2018 | MN446010 | Cryptosporidium sp. C5605-1st | KF994580 | 99.73 |
| Mountain beaver (Aplodontia rufa) | OR, 2014 | MT524974 | Cryptosporidium ubiquitum isolate A2 | KC608024 | 100 |
| Mountain beaver (Aplodontia rufa) | OR, 2015 | MT524976 | Cryptosporidium sp. isolate Bobcat 2014_PNW1108 | MT524975 | 98.02 |
| Mountain beaver (Aplodontia rufa) | OR, 2019 | MN446006 | Cryptosporidium ducismarci strain R17-999 | MF737079 | 97.95 |
| California ground squirrel (Otospermophilus beecheyi) | CA, 2003 | AY462233 | Cryptosporidium rubeyi | KM010224 | 100 |
| California ground squirrel (Otospermophilus beecheyi) | CA, 2003 | AY462231 | Cryptosporidium sp. Sbld05a | DQ295017 | 98.60 |
| California ground squirrel (Otospermophilus beecheyi) | CA, 2003 | AY462232 | Cryptosporidium sp. Sbld05a | DQ295017 | 97.85 |
| California ground squirrel (Otospermophilus beecheyi) | CA, 2011 | KM010225 | Cryptosporidium scrofarum isolate Henan KB | MH178036 | 99.75 |
| Belding’s ground squirrel (Urocitellus beldingi) | CA, 2005 | DQ295015 | Cryptosporidium sp. wild rodent strain isolate 1134 (deer mouse) | KM199844 | 99.76 |
| Belding’s ground squirrel (Urocitellus beldingi) | CA, 2005 | DQ295017 | Cryptosporidium sp. Sbey03b | AY462232 | 97.85 |
| Belding’s ground squirrel (Urocitellus beldingi) | CA, 2005 | DQ295013 | Cryptosporidium sp. Sltl05c | DQ295014 | 98.98 |
| Golden-mantled ground squirrel (Callospermophilus lateralis) | CA, 2005 | DQ295014 | Cryptosporidium sp. Sbey05c | DQ295012 | 99.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Atwill, E.R. Diverse Genotypes and Species of Cryptosporidium in Wild Rodent Species from the West Coast of the USA and Implications for Raw Produce Safety and Microbial Water Quality. Microorganisms 2021, 9, 867. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040867
Li X, Atwill ER. Diverse Genotypes and Species of Cryptosporidium in Wild Rodent Species from the West Coast of the USA and Implications for Raw Produce Safety and Microbial Water Quality. Microorganisms. 2021; 9(4):867. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040867
Chicago/Turabian StyleLi, Xunde, and Edward Robert Atwill. 2021. "Diverse Genotypes and Species of Cryptosporidium in Wild Rodent Species from the West Coast of the USA and Implications for Raw Produce Safety and Microbial Water Quality" Microorganisms 9, no. 4: 867. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040867