
Straw Incorporation with Nitrogen Amendment Shapes Bacterial Community Structure in an Iron-Rich Paddy Soil by Altering Nitrogen Reserves

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design and Soil Sampling
2.3. Soil Physicochemical Analysis
2.4. DNA Extraction, PCR Amplification, and Illumina MiSeq Sequencing
2.5. Data Analysis
3. Results
3.1. Soil Physicochemical Properties
3.2. Soil Bacterial Community Diversity
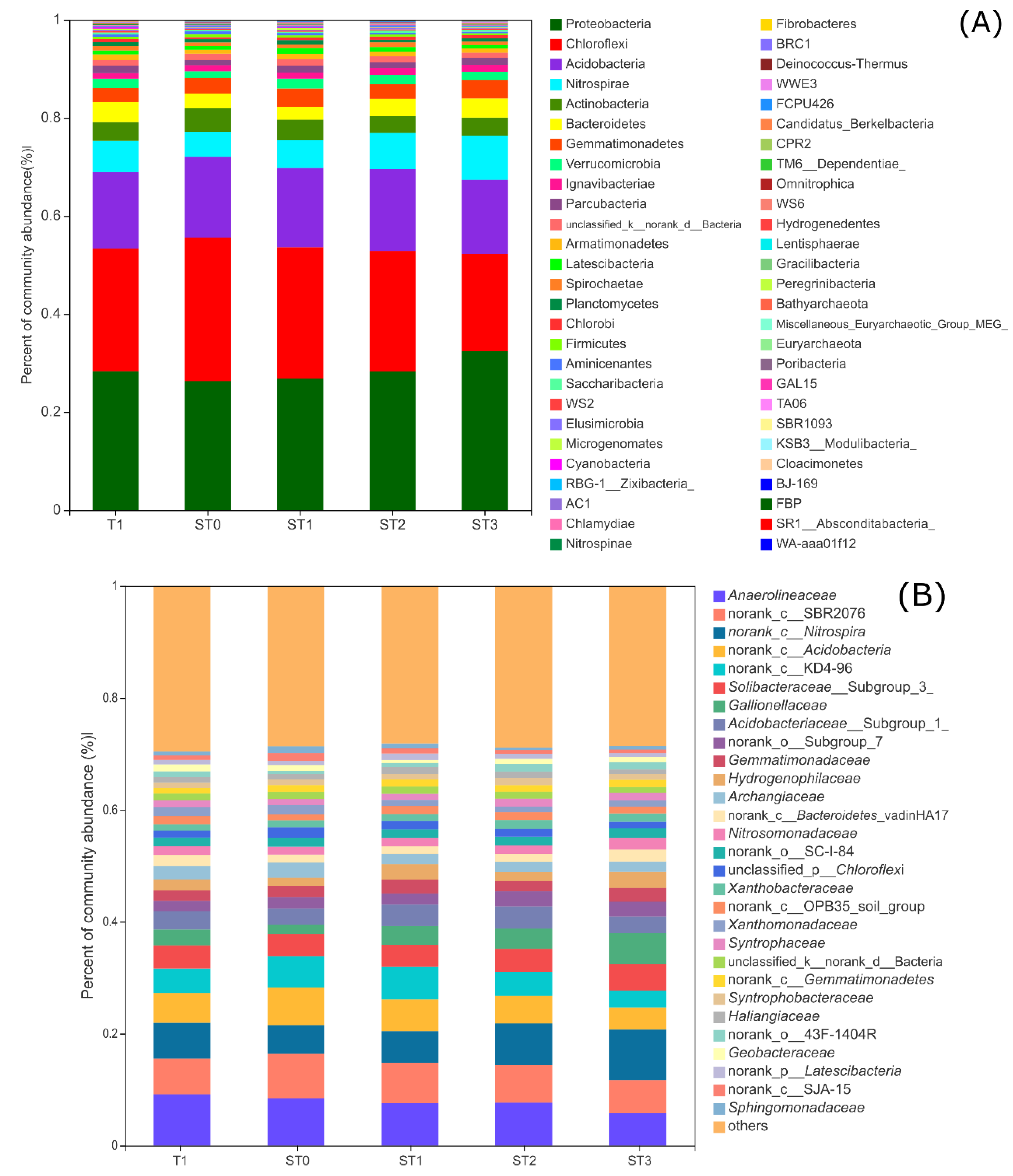
3.3. Taxonomic Composition of Bacterial Communities
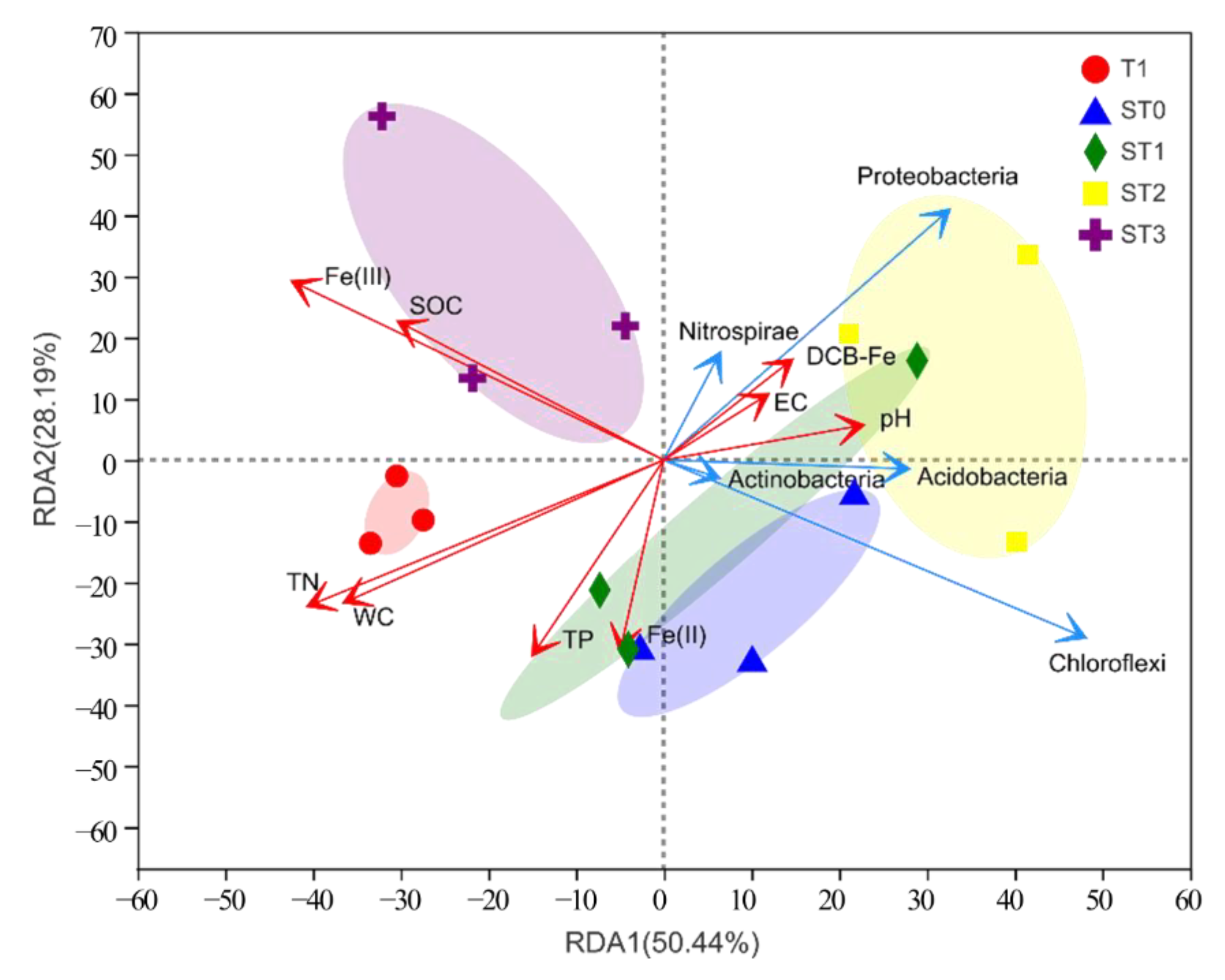
3.4. Influence of Environmental Factors on Bacterial Community Structure
3.5. Predicted Functions of Bacterial Communities
4. Discussion
4.1. Effects of Straw Incorporation with N Amendment on Paddy Soil Fertility
4.2. Effects of Straw Incorporation with N Amendment on Bacterial Community Structure
4.3. Relationship between Bacterial Community Structure and Soil Environment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cook, H.F.; Valdes, G.S.B.; Lee, H.C. Mulch effects on rainfall interception, soil physical characteristics and temperature under Zea mays L. Soil Tillage Res. 2006, 91, 227–235. [Google Scholar] [CrossRef]
- Lal, R. Beyond Copenhagen: Mitigating climate change and achieving food security through soil carbon sequestration. Glob. Food Sec. 2010, 2, 169–177. [Google Scholar] [CrossRef]
- Feng, J.; Wang, B.; Zhang, D.; Chu, S.H.; Zhi, Y.; Hayat, K.; Wang, J.C.; Chen, X.F.; Hui, N.; Zhou, P. Streptomyces griseorubens JSD-1 promotes rice straw composting efficiency in industrial-scale fermenter: Evaluation of change in physicochemical properties and microbial community. Bioresour. Technol. 2021, 321, 124465. [Google Scholar] [CrossRef] [PubMed]
- Medina, J.; Monreal, C.; Barea, J.M.; Arriagada, C.; Borie, F.; Cornejo, P. Crop residue stabilization and application to agricultural and degraded soils: A review. Waste Manag. 2015, 42, 41–54. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, J.B.; Zhao, B.Z.; Yan, P.; Zhou, G.X.; Xin, X.L. Effects of straw amendment and moisture on microbial communities in Chinese fluvo-aquic soil. J. Soils Sediments 2014, 14, 1829–1840. [Google Scholar] [CrossRef]
- Li, H.; Dai, M.W.; Dai, S.L.; Dong, X.J. Current status and environment impact of direct straw return in China’s cropland-A review. Ecotoxicol. Environ. Saf. 2018, 159, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.L.; Cheng, H.; Liu, Z.F.; Ren, W.W. Experimental warming on the rice-wheat rotation agro-ecosystem. Plant Sci. J. 2013, 31, 49–56. [Google Scholar] [CrossRef]
- Zhu, L.Q.; Hu, N.J.; Yang, M.F.; Zhan, X.H.; Zhang, Z.W. Effects of different tillage and straw return on soil organic carbon in a rice-wheat rotation system. PLoS ONE 2014, 9, e0088900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, T.; Feiner, H.; Bettina, J.; Helfrich, M.; Ludwig, B. Organic matter in density fractions of water-stable aggregates in silty soils: Effect of land use. Soil Biol. Biochem. 2006, 38, 3222–3234. [Google Scholar] [CrossRef]
- Yang, H.S.; Yang, B.; Dai, Y.J.; Xu, M.M.; Koide, R.T.; Wang, X.H.; Liu, J.; Bian, X.M. Soil nitrogen retention is increased by ditch-buried straw return in a rice-wheat rotation system. Eur. J. Agron. 2015, 69, 52–58. [Google Scholar] [CrossRef]
- Wu, J. Carbon accumulation in paddy ecosystems in subtropical China: Evidence from landscape studies. Eur. J. Soil Sci. 2011, 62, 29–34. [Google Scholar] [CrossRef]
- Majumder, B.; Mandal, B.; Bandyopadhyay, P.; Gangopadhyay, A.; Mani, P.; Kundu, A.; Mazumdar, D. Organic amendments influence soil organic carbon pools and rice-wheat productivity. Soil Sci. Soc. Am. J. 2008, 72, 775–785. [Google Scholar] [CrossRef]
- Potthoff, M.; Dyckmans, J.; Flessa, H.; Muhs, A.; Beese, F.; Joergensen, R.G. Dynamics of maize (Zea mays L.) leaf straw mineralization as affected by the presence of soil and the availability of nitrogen. Soil Biol. Biochem. 2005, 37, 1259–1266. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Xu, M.; Feng, G.; Zhang, W.; Lu, C. Crop yield and soil organic matter after long-term straw return to soil in China. Nutr. Cycl. Agroecosyst. 2015, 102, 371–381. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Wang, M.Y.; Hu, S.J.; Zhang, X.D.; Ouyang, Z.; Zhang, G.L.; Huang, B.; Zhao, S.W.; Wu, J.S.; Xie, D.T.; et al. Economics-and policy-driven organic carbon input enhancement dominates soil organic carbon accumulation in Chinese croplands. Proc. Natl. Acad. Sci. USA 2018, 115, 4045–4050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.M.; He, L.; Zhang, Z.D.; Wang, H.B.; Zhao, L.P. Simulation of accumulation and mineralization (CO2 release) of organic carbon in chernozem under different straw return ways after corn harvesting. Soil Tillage Res. 2016, 156, 148–154. [Google Scholar] [CrossRef]
- Ryan, M.; Terry, J.; Thomas, W. Bacterial metataxonomic profile and putative functional behavior associated with C and N cycle processes remain altered for decades after forest harvest. Soil Biol. Biochem. 2018, 119, 184–193. [Google Scholar]
- Olk, D.C.; Samson, M.I.; Gapas, P. Inhibition of nitrogen mineralization in young humic fractions by anaerobic decomposition of rice crop residues. Eur. J. Soil Sci. 2007, 58, 270–281. [Google Scholar] [CrossRef]
- Cucu, M.A.; Pullicino, D.; Maurino, V.; Bonifacio, E.; Romani, M.; Celi, L. Influence of redox conditions and rice straw incorporation on nitrogen availability in fertilized paddy soils. Biol. Fertil. Soils 2014, 50, 75–764. [Google Scholar] [CrossRef]
- Xia, L.L.; Lam, S.K.; Wolf, B.; Kiese, R.; Chen, D.L.; Butterbach-Bahl, K. Trade-offs between soil carbon sequestration and reactive nitrogen losses under straw return in global agroecosystems. Glob. Chang. Biol. 2018, 24, 5919–5932. [Google Scholar] [CrossRef]
- Breulmann, M.; Schulz, E.; Weißhuhn, K.; Buscot, F. Impact of the plant community composition on labile soil organic carbon, soil microbial activity and community structure in semi-natural grassland ecosystems of different productivity. Plant Soil 2012, 352, 253–265. [Google Scholar] [CrossRef]
- Xia, L.L.; Lam, S.K.; Yan, X.Y.; Chen, D.L. How Does Recycling of Livestock Manure in Agroecosystems Affect Crop Productivity, Reactive Nitrogen Losses, and Soil Carbon Balance? Environ. Sci. Technol. 2017, 51, 7450–7457. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Caspari, T.; Gonzalez, M.R.; Batjes, N.H.; Toth, Z. Effects of agricultural management practices on soil quality. Agric. Ecosyst. Environ. 2018, 265, 1–7. [Google Scholar] [CrossRef]
- Tardy, V.; Chabbi, A.; Charrier, X.; Berranger, C.D.; Reignier, T.; Dequiedt, S.; Faivre-Primot, C.; Terrat, S.; Ranjard, L.; Maron, P. Land use history shifts in situ fungal and bacterial successions following wheat straw input into the soil. PLoS ONE 2015, 10, e0130672. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.L.; Xin, L.; Liu, J.T.; Yuan, M.Z.; Liu, S.T.; Jiang, W.; Chen, J.P. Changes in bacterial community of soil induced by long-term straw returning. Sci. Agric. 2017, 74, 349–356. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, S.Q.; Xing, G.X. Nitrification, acidification, and nitrogen leaching from subtropical cropland soils as affected by rice straw-based biochar: Laboratory incubation and column leaching studies. J. Soils Sediments 2014, 14, 471–482. [Google Scholar] [CrossRef]
- Huang, X.Z.; Wang, C.; Liu, Q.; Zhu, Z.K.; Lynn, T.M.; Shen, J.L.; Whiteley, A.S.; Kumaresan, D.; Ge, T.D.; Wu, J.S. Abundance of microbial CO2-fixing genes during the late rice season in a long-term management paddy field amended with straw and straw-derived biochar. Can. J. Soil Sci. 2018, 98, 306–316. [Google Scholar] [CrossRef]
- Bu, R.Y.; Ren, T.; Lei, M.J.; Liu, B.; Li, X.K.; Cong, R.H.; Zhang, Y.Y.; Lu, J.W. Tillage and straw-returning practices effect on soil dissolved organic matter, aggregate fraction and bacteria community under rice-rice-rapeseed rotation system. Agric. Ecosyst. Environ. 2020, 287. [Google Scholar] [CrossRef]
- Miao, S.J.; Zhao, H.F.; Pan, S.Q.; Zhang, N.; Qiao, Y.F. Elevated CO2 weakens the shift in bacterial community structure in response to 8-year soybean straw return in the same experiment. Int. J. Phytoremed. 2020, 1–6. [Google Scholar] [CrossRef]
- Pronk, G.J.; Heister, K.; Kögel-Knabner, I. Iron oxides as major available interface component in loamy arable topsoils. Soil Sci. Soc. Am. J. 2011, 75, 2158–2168. [Google Scholar] [CrossRef]
- Wang, J.; Muyzer, G.; Bodelier, P.L.E.; Laanbroek, H.J. Diversity of iron oxidizers in wetland soils revealed by novel 16S rRNA primers targeting Gallionella-related bacteria. ISME J. 2009, 3, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Hartland, A.; Andersen, M.S.; Hamilton, D.P. Phosphorus and arsenic distributions in a seasonally stratified, iron- and manganese-rich lake: Microbiological and geochemical controls. Environ. Chem. 2015, 12, 708–724. [Google Scholar] [CrossRef]
- Pan, W.; Kan, J.; Inamdar, S.; Chen, C.; Sparks, D. Dissimilatory microbial iron reduction release doc (dissolved organic carbon) from carbon-ferrihydrite association. Soil Biol. Biochem. 2016, 103, 232–240. [Google Scholar] [CrossRef]
- Wang, N.; Yu, J.G.; Zhao, Y.H.; Chang, Z.Z.; Shi, X.X.; Ma, L.Q.; Li, H.B. Straw enhanced CO2 and CH4 but decreased N2O emissions from flooded paddy soils: Changes in microbial community compositions. Atmos. Environ. 2018, 174, 171–179. [Google Scholar] [CrossRef]
- Hu, J.; Wu, H.; Sun, Z.; Peng, Q.A.; Hu, R. Ferrous iron (Fe2+) addition decreases methane emissions induced by rice straw in flooded paddy soils. ACS Earth Space Chem. 2020, 4, 843–853. [Google Scholar] [CrossRef]
- Violante, A.; Barberis, E.; Pigna, M.; Boero, V. Factors affecting the formation, nature, and properties of iron precipitation products at the soil–root interface. J. Plant Nutr. 2003, 26, 1889–1908. [Google Scholar] [CrossRef]
- Xu, X.W.; Wang, P.; Zhang, J.; Chen, C.; Wang, Z.P.; Kopittke, P.M.; Kretzschmar, R.; Zhao, F.J. Microbial sulfate reduction decreases arsenic mobilization in flooded paddy soils with high potential for microbial Fe reduction. Environ. Pollut. 2019, 251, 952–960. [Google Scholar] [CrossRef]
- Weber, K.A.; Achenbach, L.A.; Coates, J.D. Microorganisms pumping iron: Anaerobic microbialiron oxidation and reduction. Nat. Rev. Microbiol. 2006, 4, 752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Su, J.Q.; Yang, X.R.; Zhou, G.W.; Lassen, S.B.; Zhu, Y.G. RNA stable isotope probing of potential Feammox population in paddy soil. EST 2019, 53, 4841–4849. [Google Scholar] [CrossRef]
- Ding, B.J.; Zhang, H.; Luo, W.Q.; Sun, S.Y.; Cheng, F.; Li, Z.K. Nitrogen loss through denitrification, anammox and Feammox in a paddy soil. Sci. Total Environ. 2021, 145601. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Xie, G.J.; Nie, W.B.; Xing, D.F.; Liu, B.F.; Ding, J.; Ren, N.Q. Fe(III)-mediated anaerobic ammonium oxidation: A novel microbial nitrogen cycle pathway and potential applications. Crit. Rev. Environ. Sci. Technol. 2021, 1–33. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Duan, G.L.; Chen, B.D.; Peng, X.H.; Chen, Z.L.; Sun, G.X. Mineral weathering and element cycling in soil-microorganism-plant system. Sci. China Earth Sci. 2014, 57, 888–896. [Google Scholar] [CrossRef]
- Xia, L.L.; Lam, S.K.; Wang, S.W.; Zhou, W.; Chen, D.L.; Yan, X.Y. Optimizing nitrogen fertilization rate to enhance soil carbon storage and decrease nitrogen pollution in paddy ecosystems with simultaneous straw incorporation. Agric. Ecosyst. Environ. 2020, 298. [Google Scholar] [CrossRef]
- Huang, T.; Yang, H.; Huang, C.; Ju, X. Effects of nitrogen management and straw return on soil organic carbon sequestration and aggregate-associated carbon. Eur. J. Soil Sci. 2018, 69, 913–923. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agro-chemistrical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2007; pp. 268–270, 389–391. [Google Scholar]
- Kostka, J.E.; Iii, G.W.L. Partitioning and speciation of solid phase iron in saltmarsh sediments. Geochim. Cosmochim. Acta 1994, 58, 1701–1710. [Google Scholar] [CrossRef]
- Mehra, O.P.; Jackson, M.L. Iron oxide removal from soils and clays by a dithionite-citrate system buffered with sodium bicarbonate. Clays Clay Miner 1960, 7, 317–327. [Google Scholar] [CrossRef]
- Dennis, K.L.; Wang, Y.; Blatner, N.R.; Wang, S.; Abdulrahman, S.; Trudeau, E.; Roers, A.; Weaver, C.T.; Lee, J.J.; Gilbert, J.A. Adenomatous polyps are driven by microbe-instigated focal inflammation and are controlled by IL-10-producing T cells. Cancer Res. 2013, 73, 5905–5913. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Stackebrandt, E.; Goebel, B. Taxonomic note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Goh, K.M. Nitrogen release from crop residues and organic amendments as affected by biochemical composition. Commun. Soil Sci. Plan. 2003, 34, 2441–2460. [Google Scholar] [CrossRef]
- Wang, S.C.; Zhao, Y.W.; Wang, J.Z.; Zhu, P.; Cui, X.; Han, X.Z.; Xu, M.G.; Lu, C.A. The efficiency of long-term straw return to sequester organic carbon in Northeast China’s cropland. J. Integr. Agric. 2018, 17, 436–448. [Google Scholar] [CrossRef]
- Li, S.; Li, M.; Wang, G.; Sun, X.; Hu, Z. Compositional and chemical characteristics of dissolved organic matter in various types of cropped and natural Chinese soils. Chem. Biol. Technol. Agric. 2019, 6, 1–11. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, K.; Li, K.; Zheng, C.; Li, B. Simulating the effects of long-term discontinuous and continuous fertilization with straw return on crop yields and soil organic carbon dynamics using the DNDC model. Soil Tillage Res. 2017, 165, 302–314. [Google Scholar] [CrossRef]
- Xu, X.; Pang, D.W.; Chen, J.; Luo, Y.L.; Zheng, M.J.; Yin, Y.P.; Li, Y.X.; Li, Y.; Wang, Z.L. Straw return accompany with low nitrogen moderately promoted deep root. Field Crops Res. 2018, 221, 71–80. [Google Scholar] [CrossRef]
- Zeng, X.N.; Song, Q.L.; Yu, S.; Zhou, M.; Feng, Y.J.; Lai, Y.C.; Wang, Q. Effects of fertilization and straw incorporation on bacterial communities in black soil, Northeastern China. Appl. Ecol. Environ. Res. 2020, 18, 7535–7550. [Google Scholar] [CrossRef]
- Fierer, N.; Christian, L.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [Green Version]
- Geisseler, D.; Scow, K.M. Long-term effects of mineral fertilizers on soil microorganisms—A review. Soil Biol. Biochem. 2014, 75, 54–63. [Google Scholar] [CrossRef]
- Suding, K.N.; Collins, S.L.; Gough, L.; Clark, C.; Cleland, E.E.; Gross, K.L.; Milchunas, D.G.; Pennings, S.; Chapin, F.S. Functional-and abundance-based mechanisms explain diversity loss due to N fertilization. Proc. Natl. Acad. Sci. USA 2005, 102, 4387–4392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes. Glob. Chang. Biol. 2012, 18, 1918–1927. [Google Scholar] [CrossRef]
- Zhu, J.; Peng, H.; Ji, X.H.; Li, C.J.; Li, S.N. Effects of reduced inorganic fertilization and rice straw recovery on soil enzyme activities and bacterial community in double-rice paddy soils. Eur. J. Soil Biol. 2019, 94. [Google Scholar] [CrossRef]
- Liu, L.; Ding, M.J.; Zhou, L.K.; Chen, Y.; Li, H.P.; Zhang, F.M.; Li, G.; Zhou, Z.F.; Zhang, Y.; Zhou, X.X. Effects of different rice straw on soil microbial community structure. Agron. J. 2020. [Google Scholar] [CrossRef]
- Zhou, G.P.; Gao, S.J.; Chang, D.N.; Rees, R.M.; Cao, W.D. Using milk vetch (Astragalus sinicus L.) to promote rice straw decomposition by regulating enzyme activity and bacterial community. Bioresour. Technol. 2021, 319, 124–215. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ge, Y.; Song, J.; Christopher, R. Assembly of root-associated microbial community of typical rice cultivars in different soil types. Biol. Fert. Soils 2020, 56, 1–12. [Google Scholar] [CrossRef]
- Bastida, F.; Torres, I.F.; Hernández, T.; García, C. The impacts of organic amendments: Do they confer stability against drought on the soil microbial community? Soil Biol. Biochem. 2017, 113, 173–183. [Google Scholar] [CrossRef]
- Liu, J.J.; Yao, Q.; Li, Y.S.; Zhang, W.; Wang, G.H. Continuous cropping of soybean alters the bulk and rhizospheric soil fungal communities in a Mollisol of Northeast PR China. Land Degrad. Dev. 2019, 30, 725–1738. [Google Scholar] [CrossRef]
- Paterson, E.; Osler, G.; Dawson, L.A.; Gebbing, T.; Sim, A.; Ord, B. Labile and recalcitrant plant fractions are utilised by distinct microbial communities in soil: Independent of the presence of roots and mycorrhizal fungi. Soil Biol. Biochem. 2008, 40, 1103–1111. [Google Scholar] [CrossRef]
- Baker, B.J.; Sheik, C.S.; Taylor, C.A.; Jain, S.; Bhasi, A.; Cavalcoli, J.D.; Dick, G.J. Community transcriptomic assembly reveals microbes that contribute to deep-sea carbon and nitrogen cycling. ISME J. 2013, 7, 1962–1973. [Google Scholar] [CrossRef] [Green Version]
- Harris, J. Soil microbial communities and restoration ecology: Facilitators or followers? Science 2009, 325, 573–574. [Google Scholar] [CrossRef]
- Jeong, S.; Hong, J.K.; Jho, E.H.; Nam, K. Interaction among soil physicochemical properties, bacterial community structure, and arsenic contamination: Clay-induced change in long-term arsenic contaminated soils. J. Hazard. Mater. 2019, 378, 120729. [Google Scholar] [CrossRef]
- Li, X.; Mou, S.; Chen, Y.; Liu, T.; Dong, J.; Li, F. Microaerobic Fe(II) oxidation coupled to carbon assimilation processes driven by microbes from paddy soil. Sci. China-Earth Sci. 2019, 62, 1719–1729. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Wheat Straw | Nitrogen in the Form of Urea | Phosphorus in the Form of Calcium Superphosphate | Potassium in the Form Potassium Chloride |
|---|---|---|---|---|
| T1 | 0 | 80 | 80 | 80 |
| ST0 | 6000 | 0 | 80 | 80 |
| ST1 | 6000 | 80 | 80 | 80 |
| ST2 | 6000 | 300 | 80 | 80 |
| ST3 | 6000 | 450 * | 80 | 80 |
| Treatments | pH | EC | SOM (g kg−1) | TP (g kg−1) | TN (g kg−1) | WC (%) | Fe(II) (g kg−1) | Fe(III) (g kg−1) | DCB-Fe (g kg−1) |
|---|---|---|---|---|---|---|---|---|---|
| T1 | 5.05 ± 0.08 a | 154.00 ± 7.00 b | 32.29 ± 2.61 a | 0.60 ± 0.03 a | 1.75 ± 0.15 a | 65.94 ± 9.31 a | 3.81 ± 0.17 a | 13.12 ± 1.34 a | 74.38 ± 1.08 a |
| ST0 | 5.06 ± 0.02 a | 164.67 ± 23.01 ab | 27.13 ± 1.79 b | 0.58 ± 0.26 a | 1.57 ± 0.26 ab | 58.16 ± 9.51 ab | 3.95 ± 1.66 a | 10.39 ± 1.58 a | 79.24 ± 7.96 a |
| ST1 | 5.11 ± 0.22 a | 188.33 ± 16.07 a | 27.13 ± 0.40 b | 0.57 ± 0.04 a | 1.46 ± 0.07 b | 47.71 ± 1.95 b | 3.53 ± 1.28 a | 11.84 ± 3.49 a | 77.18 ± 3.08 a |
| ST2 | 5.05 ± 0.09 a | 158.00 ± 14.11 ab | 30.21 ± 1.71 ab | 0.57 ± 0.02 a | 1.40 ± 0.01 b | 45.95 ± 4.36 b | 2.57 ± 0.17 a | 10.53 ± 3.17 a | 78.47 ± 11.07 a |
| ST3 | 5.00 ± 0.15 a | 179.67 ± 19.09ab | 30.99 ± 2.86 ab | 0.55 ± 0.00 a | 1.47 ± 0.08 b | 50.84 ± 6.84 b | 4.86 ± 1.89 a | 14.17 ± 2.34 a | 77.92 ± 4.44 a |
| Treatments | Sobs | ACE | Shannon | Coverage |
|---|---|---|---|---|
| T1 | 2841.3 ± 96.03 b | 3812.9 ± 47.71 ab | 6.86 ± 0.02 a | 0.959 |
| ST0 | 2726.0 ± 85.58 ab | 3759.6 ± 188.9 a | 6.85 ± 0.07 ab | 0.966 |
| ST1 | 2771.3 ± 101.53 ab | 3712 ± 99.25 ab | 6.79 ± 0.04 ab | 0.968 |
| ST2 | 2970.7 ± 100.55 a | 3843.8 ± 99.4 a | 6.81 ± 0.14 ab | 0.972 |
| ST3 | 2667.0 ± 74.73 b | 3620.2 ± 39.83 b | 6.66 ± 0.09 b | 0.964 |
| Phylum | pH | EC | SOC | TP | TN | WC | Fe(II) | Fe(III) | DCB-Fe |
|---|---|---|---|---|---|---|---|---|---|
| Proteobacteria | 0.24 | 0.21 | 0.00 | −0.63 * | −0.78 *** | −0.58 * | −0.46 | −0.23 | 0.14 |
| Chloroflexi | 0.24 | 0.06 | −0.54 * | −0.02 | −0.57 * | −0.31 | −0.48 | −0.71 ** | −0.19 |
| Acidobacteria | 0.12 | 0.15 | −0.33 | −0.04 | −0.70 ** | −0.49 | −0.51 | −0.32 | −0.03 |
| Nitrospirae | −0.05 | 0.30 | 0.20 | −0.33 | −0.40 | −0.46 | −0.41 | 0.10 | 0.44 |
| Actinobacteria | 0.44 | 0.07 | −0.61 * | −0.29 | −0.61 * | −0.35 | −0.36 | −0.85 *** | −0.07 |
| Bacteroidetes | −0.31 | −0.36 | 0.51 | −0.40 | −0.27 | 0.19 | −0.14 | 0.07 | −0.12 |
| Gemmatimonadetes | 0.42 | 0.50 | −0.62 * | −0.45 | −0.74 ** | −0.80 *** | −0.24 | −0.41 | 0.20 |
| Verrucomicrobia | −0.53 * | 0.25 | 0.08 | 0.39 | 0.09 | −0.08 | 0.00 | 0.69 ** | 0.39 |
| Ignavibacteriae | 0.14 | 0.13 | −0.10 | −0.57 * | −0.80 *** | −0.36 | −0.33 | −0.12 | 0.05 |
| Parcubacteria | 0.17 | 0.48 * | −0.28 | 0.11 | −0.31 | −0.43 | −0.04 | 0.32 | 0.46 |
| Unclassified_k_norank_d_Bacteria | 0.15 | 0.50 * | −0.63 | 0.01 | −0.63 * | −0.63 * | −0.66 ** | −0.30 | 0.28 |
| Armatimonadetes | −0.04 | 0.28 | −0.11 | 0.45 | −0.03 | −0.26 | −0.03 | 0.43 | 0.33 |
| Latescibacteria | 0.32 | 0.58 * | −0.46 | 0.11 | −0.29 | −0.58 * | −0.50 | −0.13 | 0.14 |
| Spirochaetae | 0.18 | −0.34 | 0.04 | −0.54 * | −0.52 * | 0.02 | −0.27 | −0.44 | −0.55 * |
| Planctomycetes | 0.03 | 0.54 * | −0.54 * | 0.13 | −0.32 | −0.41 | 0.10 | 0.27 | 0.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Ma, Y.; Di, L.; Qian, X.; Wang, G. Straw Incorporation with Nitrogen Amendment Shapes Bacterial Community Structure in an Iron-Rich Paddy Soil by Altering Nitrogen Reserves. Microorganisms 2021, 9, 988. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9050988
Wang J, Ma Y, Di L, Qian X, Wang G. Straw Incorporation with Nitrogen Amendment Shapes Bacterial Community Structure in an Iron-Rich Paddy Soil by Altering Nitrogen Reserves. Microorganisms. 2021; 9(5):988. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9050988
Chicago/Turabian StyleWang, Juanjuan, Yao Ma, Lin Di, Xiaoqing Qian, and Guiliang Wang. 2021. "Straw Incorporation with Nitrogen Amendment Shapes Bacterial Community Structure in an Iron-Rich Paddy Soil by Altering Nitrogen Reserves" Microorganisms 9, no. 5: 988. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9050988