
Comprehensive Profiling of Microbiologically Induced CaCO3 Precipitation by Ureolytic Bacillus Isolates from Alkaline Soils

 ,
,  ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
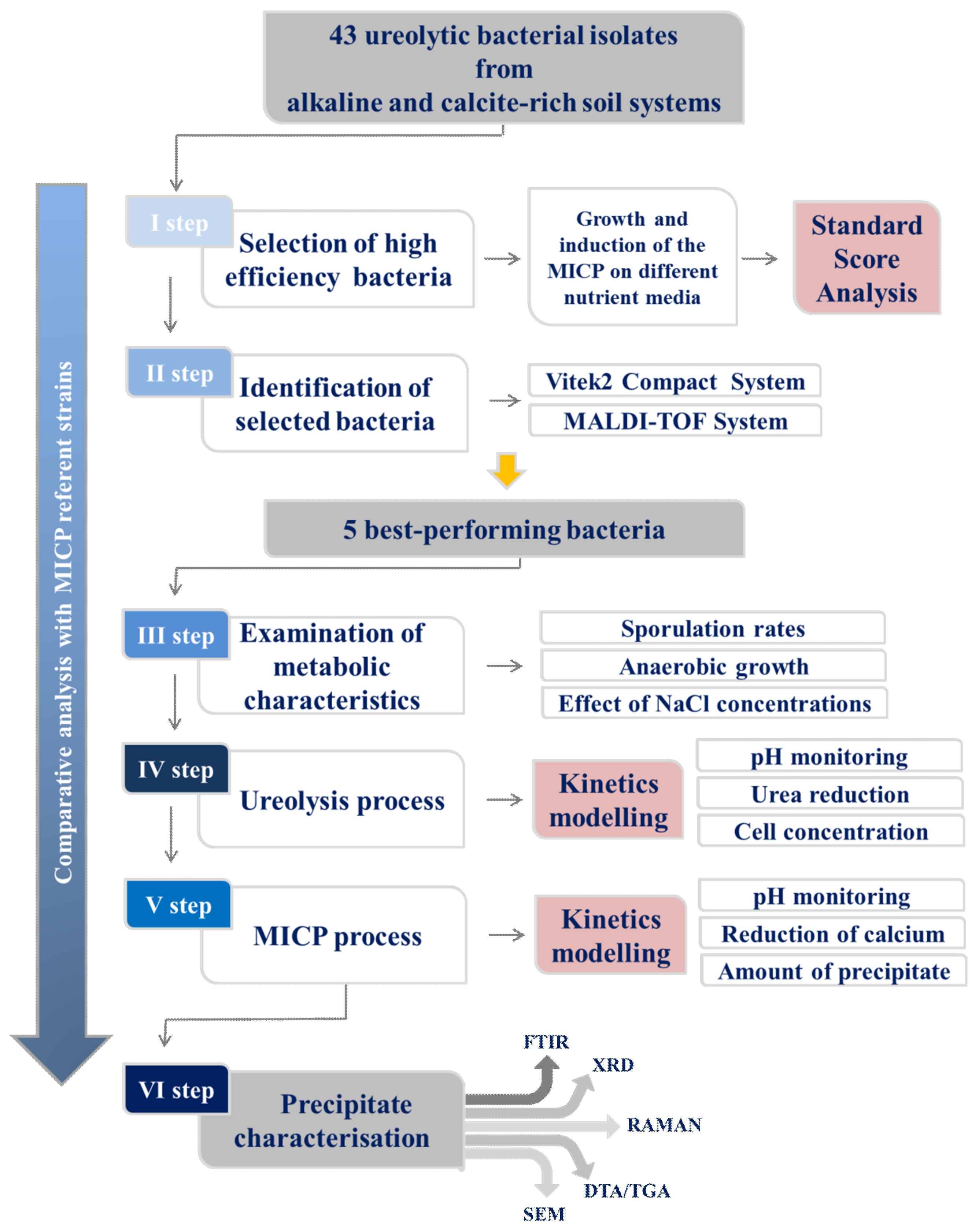
2. Materials and Methods
2.1. The First Step: Selection of the Bacterial Isolates
2.2. The Second Step: Bacterial Strain Identification
2.3. The Third Step: Examination of Metabolic Activities Relevant to the MICP
2.4. The Fourth Step: Kinetics of Ureolysis and pH Monitoring
2.5. The Fifth Step: Evaluation of the MICP Efficiency
2.5.1. Precipitation Rate Kinetics
2.5.2. Determination of Calcium Concentration
2.6. The Sixth Step: Characterization of Precipitates
3. Results and Discussion
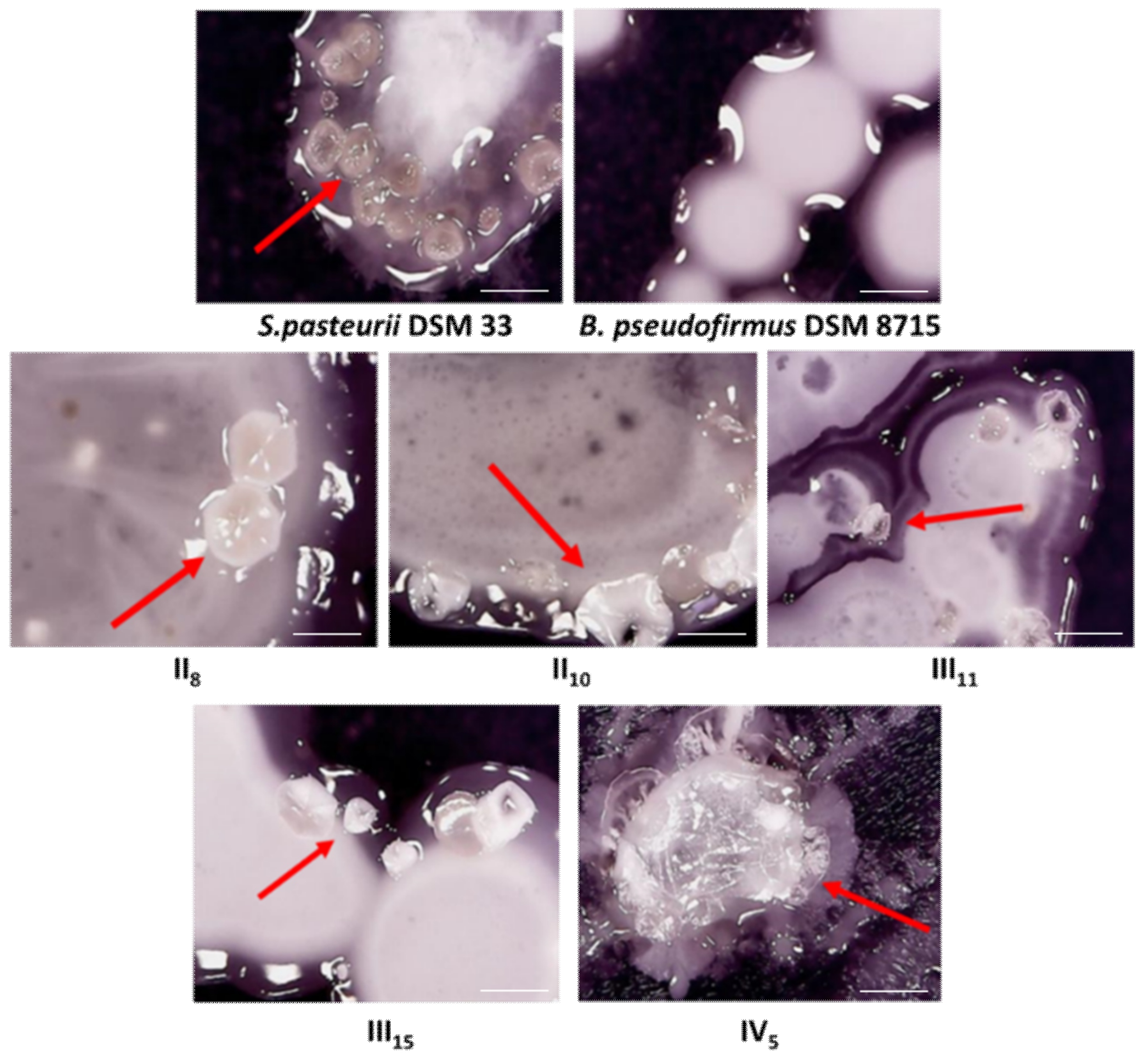
3.1. Selection of the Bacterial Isolates
3.2. Bacterial Strain Identification
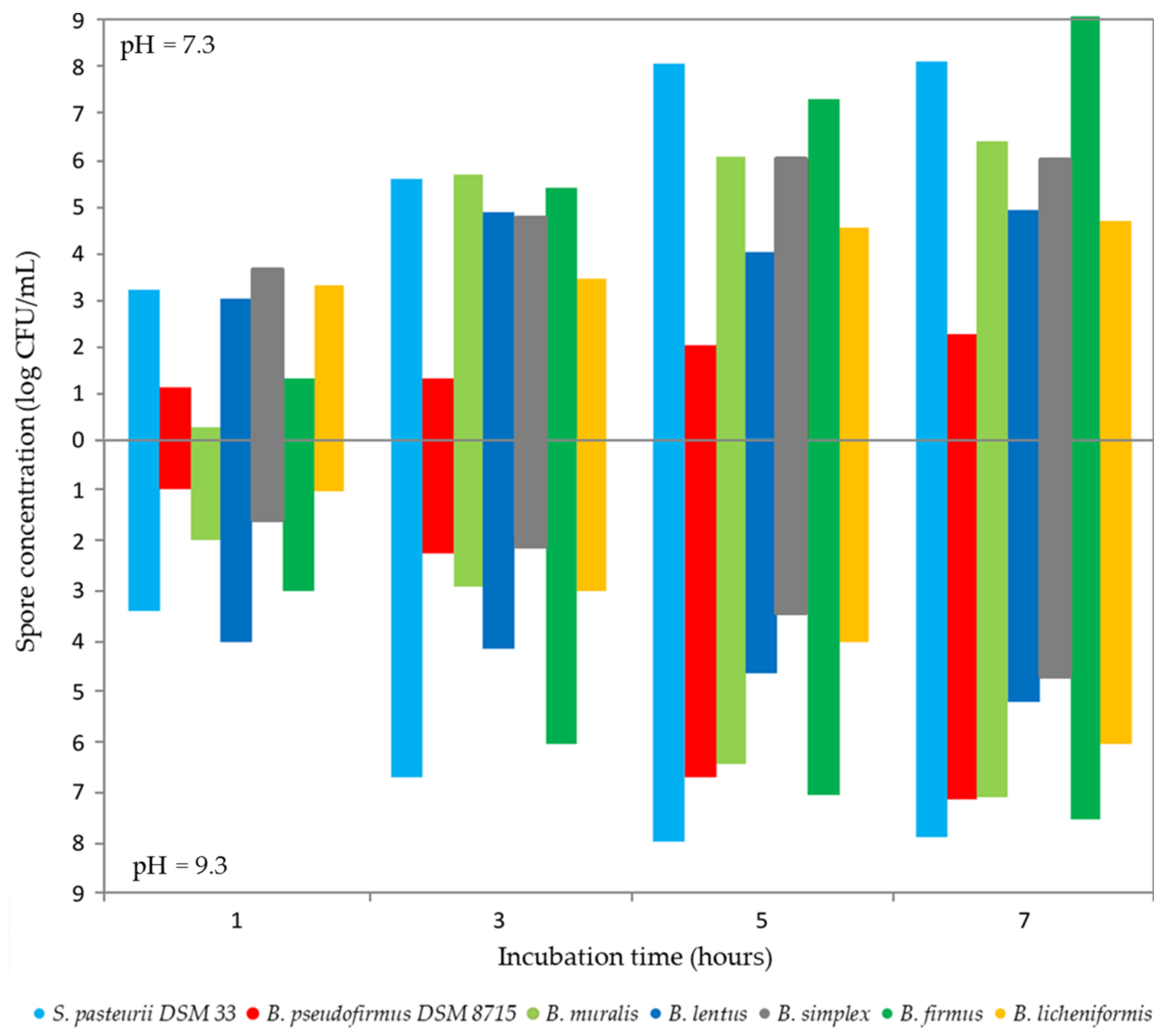
3.3. Examination of Metabolic Activities Relevant to the MICP Process
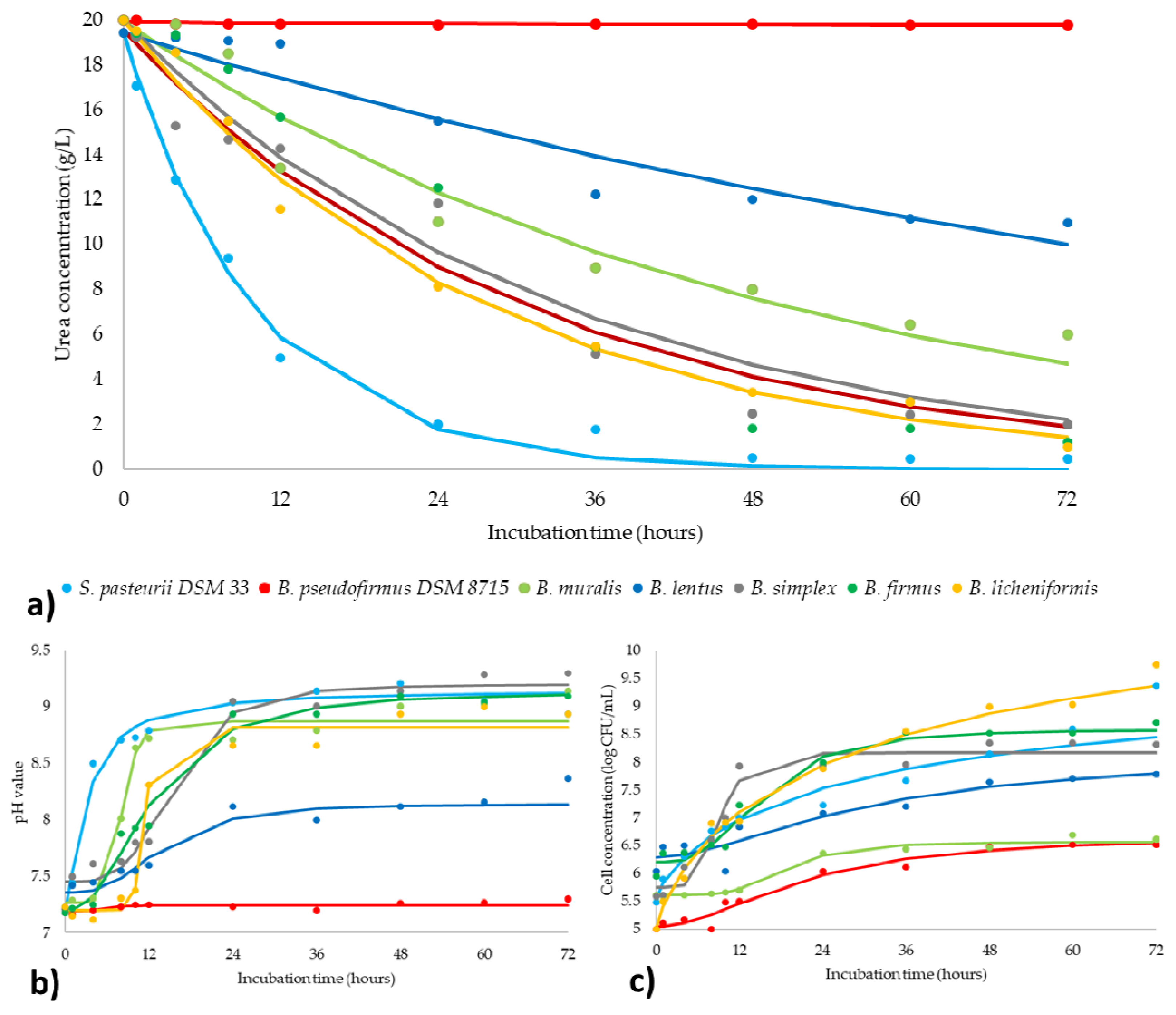
3.4. Kinetics of Ureolysis and pH Monitoring
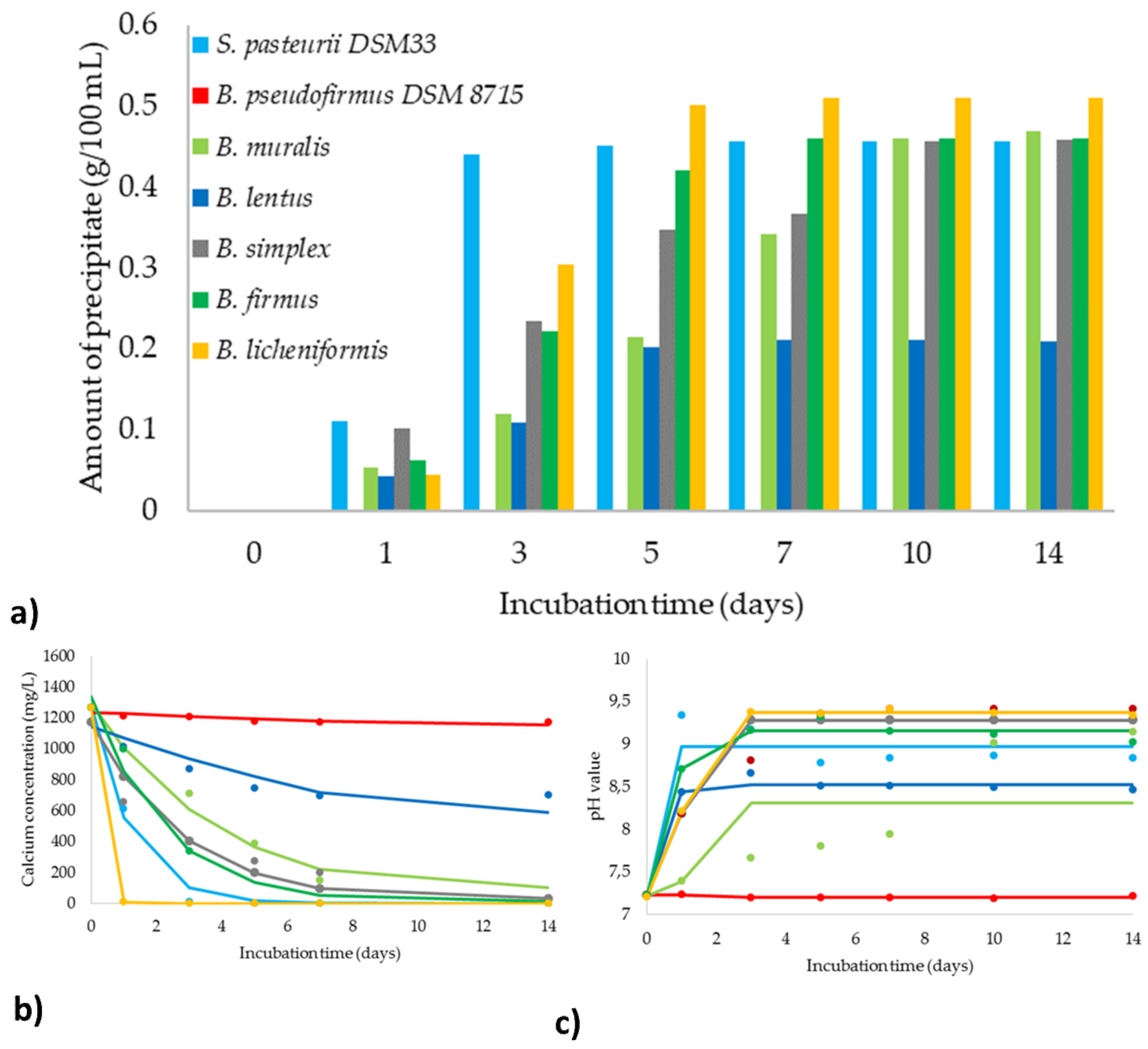
3.5. Evaluation of the MICP Efficiency
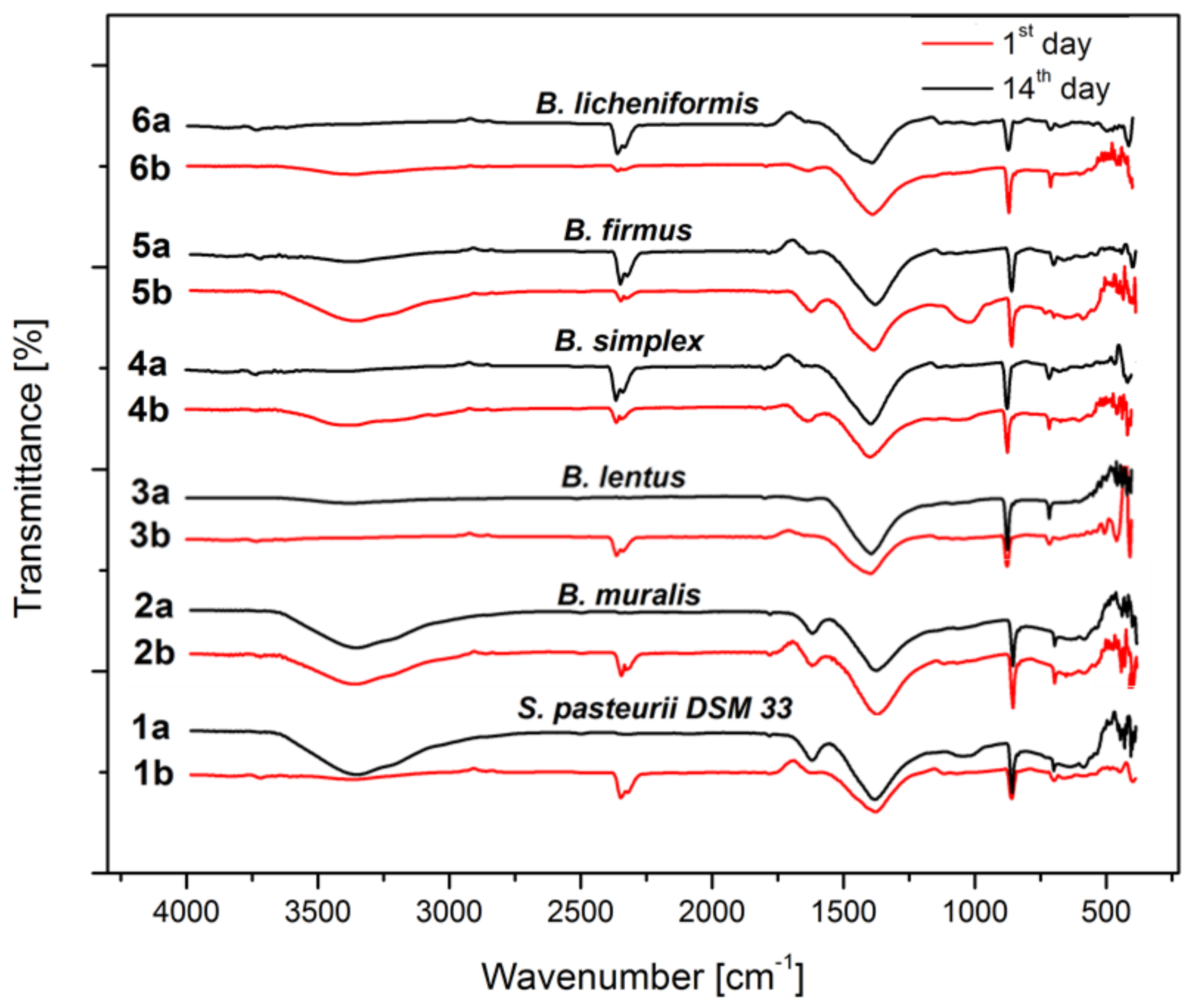
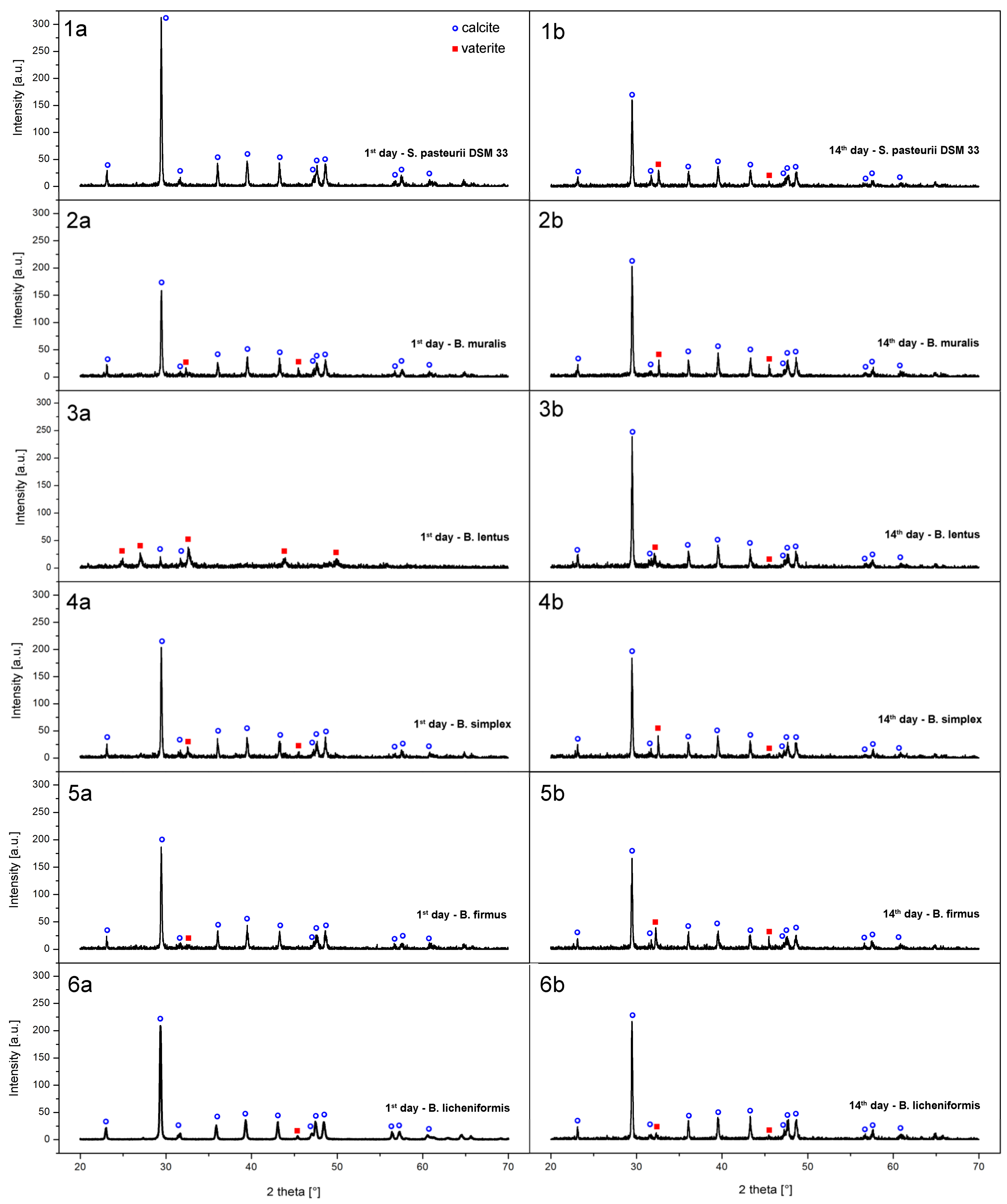
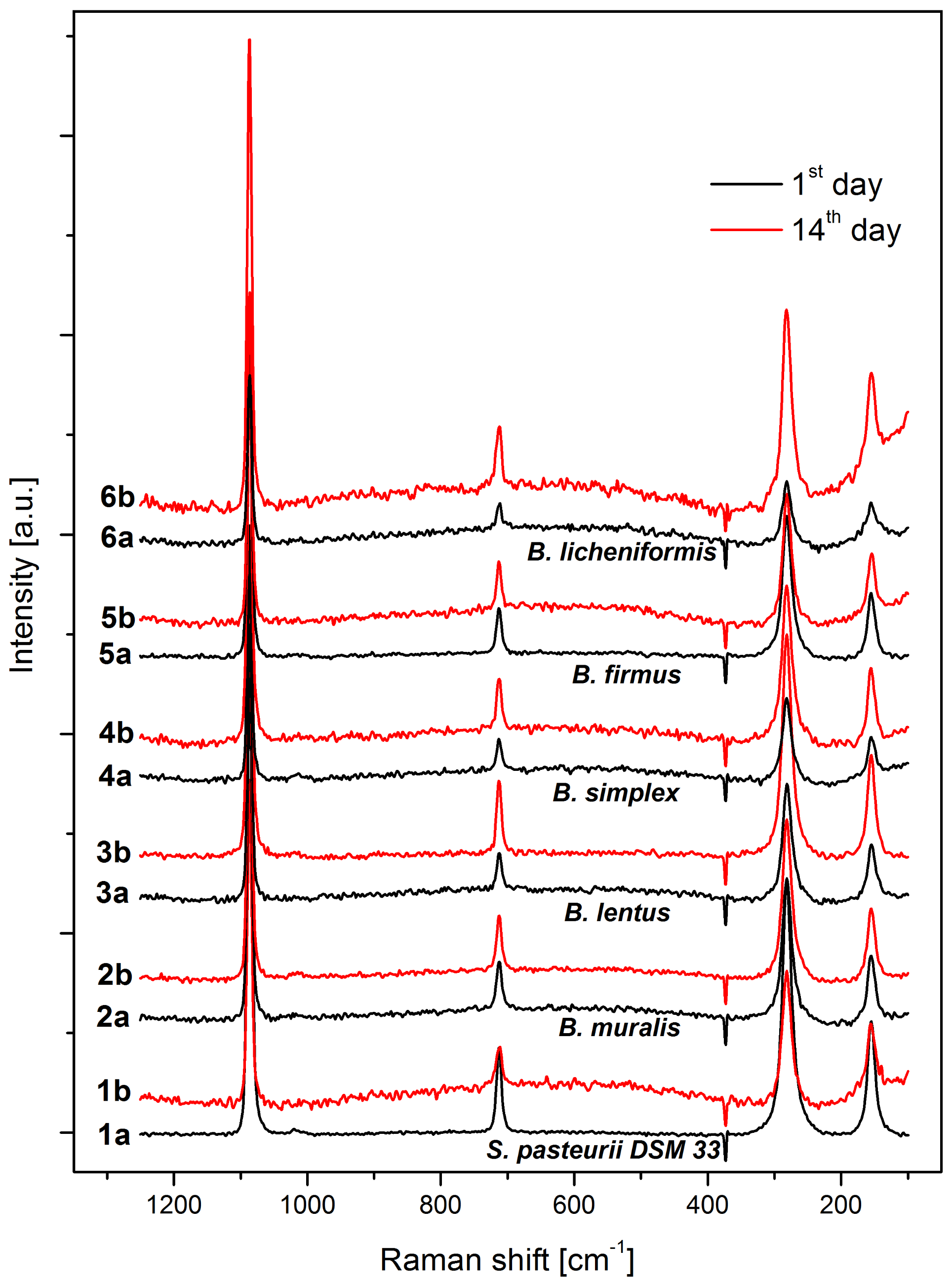
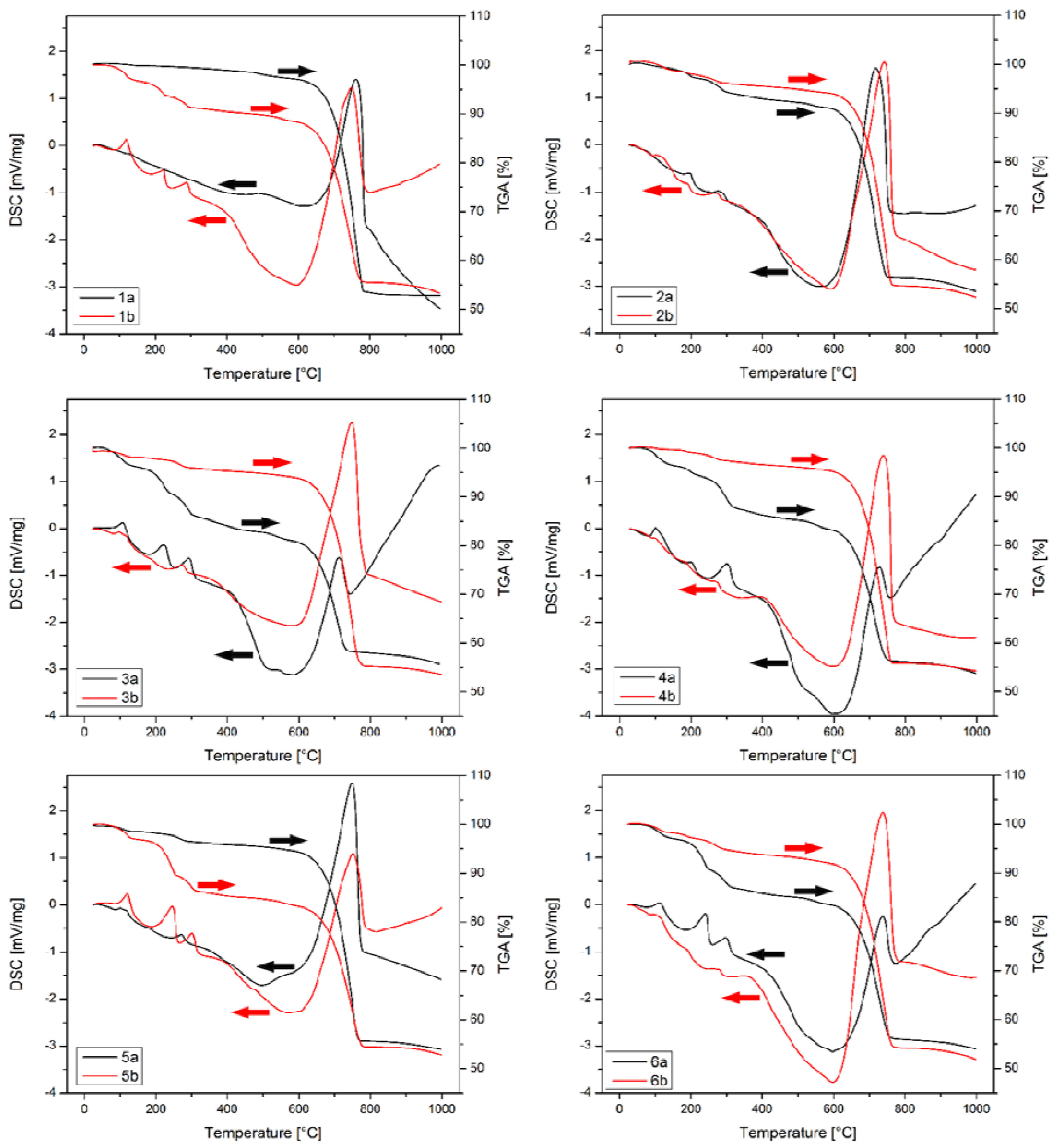
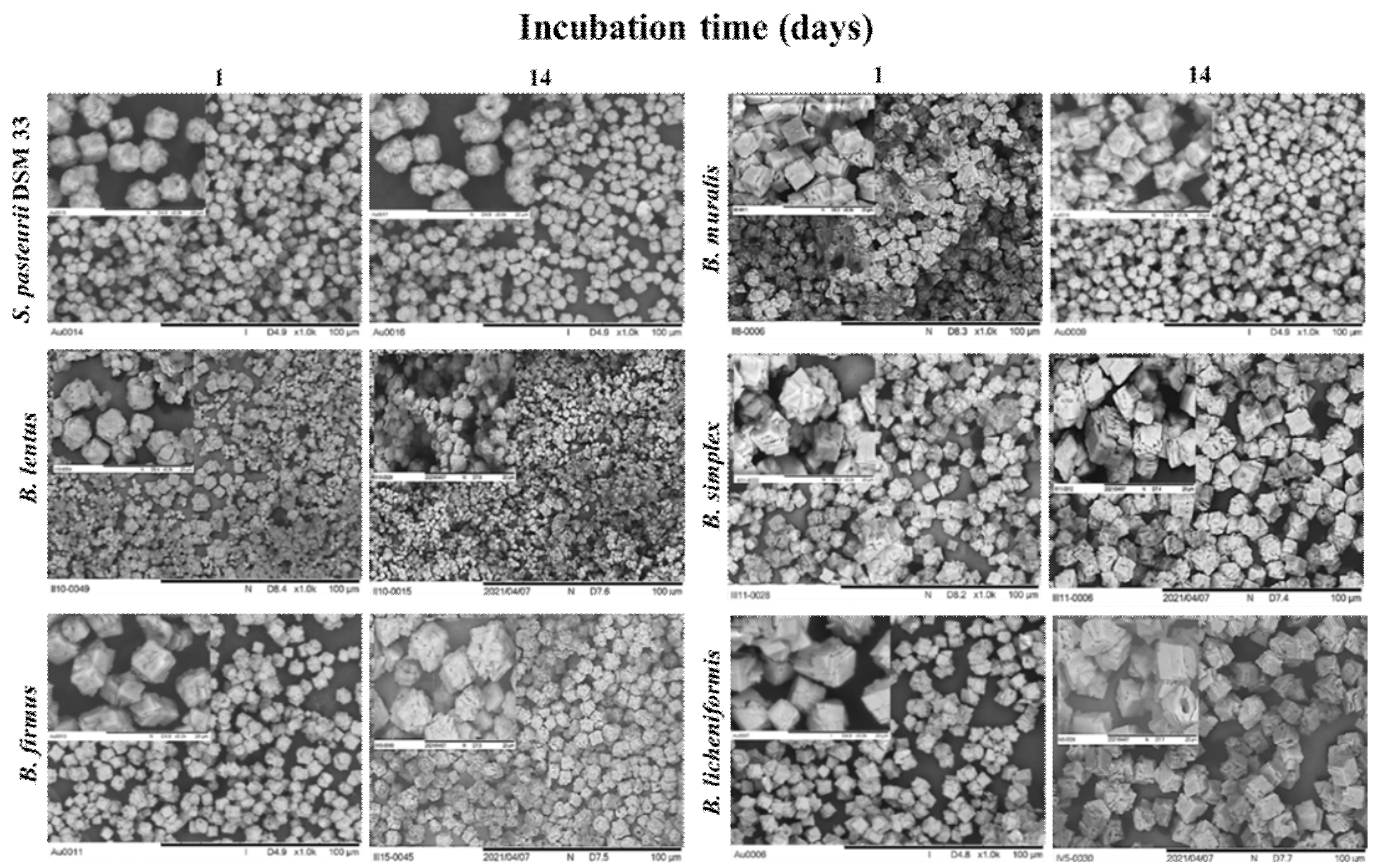
3.6. Precipitate Characterization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cacchio, P.; Ercole, C.; Cappuccio, G.; Lepidi, A. Calcium carbonate precipitation by bacterial strains isolated from a limestone cave and from a loamy soil. Geomicrobiol. J. 2003, 20, 85–98. [Google Scholar] [CrossRef]
- Jorquera, M.A.; Graether, S.P.; Maruyama, F. Bioprospecting and biotechnology of extremophiles. Front. Bioeng. Biotechnol. 2019, 7, 106. [Google Scholar] [CrossRef]
- Bibi, S.; Oualha, M.; Ashfaq, M.Y.; Suleiman, M.; Zouari, N. Isolation, differentiation and biodiversity of ureolytic bacteria of Qatari soil and their potential in microbially induced calcite precipitation (MICP) for soil stabilization. RSC Adv. 2018, 8, 5854–5863. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Cheng, X.; Guo, H. Heavy metal removal by biomineralization of urease producing bacteria isolated from soil. Int. Biodeter. Biodegrad. 2013, 76, 81–85. [Google Scholar] [CrossRef]
- Tiano, P. Biodeterioration of stone monuments a worldwide issue. Open Conf. Proc. J. 2016, 7, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Tiano, P.; Biagiotti, L.; Mastromei, G. Bacterial bio-mediated calcite precipitation for monumental stones conservation: Methods of evaluation. J. Microbiol. Meth. 2019, 36, 139–145. [Google Scholar] [CrossRef]
- Ersan, Y.; De Belie, N.; Boon, N. Resilient denitrifiers wink at microbial self-healing concrete. Inter. J. Environ. Eng. 2015, 2, 28–32. [Google Scholar]
- Achal, V.; Mukherjee, A.; Basu, P.; Reddy, M. Strain improvement of Sporosarcina pasteurii for enhanced urease and calcite production. J. Ind. Microb. Biotech. 2009, 7, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Anbu, P.; Kang, C.H.; Shin, Y.J.; So, J.S. Formations of calcium carbonate minerals by bacteria and its multiple applications. SpringerPlus 2016, 5, 2–26. [Google Scholar] [CrossRef] [Green Version]
- Lauchnor, E.G.; Topp, D.; Parker, A.; Gerlach, R. Whole-cell kinetics of ureolysis by Sporosarcina pasteurii. J. Appl. Microb. 2015, 118, 1321–1332. [Google Scholar] [CrossRef] [Green Version]
- Krajewska, B.; Ureases, I. Functional, catalytic and kinetic properties: A review. J. Mol. Catal. B Enzym. 2009, 59, 9–21. [Google Scholar] [CrossRef]
- Whiffin, V.S.; van Paassen, L.A.; Harkes, M.P. Microbial carbonate precipitation as a soil improvement technique. Geomicrobiol. J. 2007, 24, 417–423. [Google Scholar] [CrossRef]
- Dhami, N.K.; Reddy, M.S.; Mukherjee, A. Synergistic role of bacterial urease and carbonic anhydrase in carbonate mineralization. Appl. Biochem. Biotechnol. 2014, 172, 2552–2561. [Google Scholar] [CrossRef]
- Hammes, F.; Boon, N.; Clement, G.; de Villiers, J.; Siciliano, S.D.; Verstraete, W. Molecular, biochemical and ecological characterization of a bio-catalytic calcification reactor. Appl. Microb. Biotech. 2003, 62, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Achal, V.; Siddique, R.; Reddy, M.; Mukherjee, A. Improvement in the compressive strength of cement mortar by the use of a microorganism—Bacillus megaterium. In Proceedings of the International Conference, Concrete Construction; Excellence in Concrete Construction through Innovation, London, UK, 9–10 September 2009; pp. 27–30. [Google Scholar]
- Stabnikov, V.; Chu, J.; Ivanov, V.; Li, Y. Halotolerant, alkaliphilic urease-producing bacteria from different climate zones and their application for biocementation of sand. World J. Microbiol. Biotechnol. 2013, 29, 1453–1460. [Google Scholar] [CrossRef]
- Al-Thavadi, S. High Strength In-Situ Biocementation of Soil by Calcite Precipitating Locally Isolated Ureolytic Bacteria. Ph.D. Thesis, Murdoch University, Perth, Australia, 2008. [Google Scholar]
- Šovljanski, O.; Tomić, A.; Lato, L.; Markov, M. Temperature and pH growth profile prediction of newly isolated bacterial strains from alkaline soils. J. Sci. Food Agric. 2019, 100, 1155–1163. [Google Scholar] [CrossRef]
- Brlek, T.; Pezo, L.; Voća, N.; Krička, T.; Vukmirović, Đ.; Čolović, R.; Bodroža-Solarov, M. Chemometric approach for assessing the quality of olive cake pellets. Fuel Process. Technol. 2013, 116, 250–256. [Google Scholar] [CrossRef]
- Shukla, A.; Cameotra, S.S. A new method for the screening of ureolytic bacteria inducing calcium carbonate precipitation. J. Biotechnol. Biomater. 2016, 6, 248–251. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.C.; Ferris, F.G. The coprecipitation of Sr into calcite precipitates induced by bacterial ureolysis in artificial groundwater: Temperature and kinetic dependence. Geochim. Cosmochim. Acta 2005, 69, 4199–4210. [Google Scholar] [CrossRef]
- Connolly, J.M.; Jackson, B.; Rothman, A.P.; Klapper, I.; Gerlach, R. Estimation of a biofilm-specific reaction rate: Kinetics of bacterial urea hydrolysis in a biofilm. npj Biofilms Microbiomes 2015, 1, 15014. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.C.; Espinosa-Ortiz, E.J.; Parks, S.L.; Phillips, A.J.; Cunninghan, A.B.; Gerlach, R. Kinetics of calcite precipitation by ureolytic bacteria under aerobic and anaerobic conditions. Biogeosciences 2019, 16, 2147–2161. [Google Scholar] [CrossRef] [Green Version]
- Romano, A.; Toraldo, G.; Cavella, S.; Masi, P. Description of leavening of bread dough with mathematical modelling. J. Food Process. 2007, 83, 142–148. [Google Scholar] [CrossRef]
- Teng, H.H.; Dove, P.M.; De Yoreo, J.J. Kinetics of calcite growth: Surface processes and relationships to macroscopic rate laws. Geochim. Cosmochim. Acta 2000, 64, 2255–2266. [Google Scholar] [CrossRef]
- Jonkers, H.; Schlangen, E. Development of a bacteria-based self-healing concrete. Tailor Made Concr. Struct. 2008, 1, 425–430. [Google Scholar]
- Omoregie, A.I.; Khoshdelnezamiha, G.; Seniana, N.; Ek Leong Ongb, D.; Nissoma, P.M. Experimental optimization of various cultural conditions on urease activity for isolated Sporosarcina pasteurii strains and evaluation of their biocement potentials. Ecol. Eng. 2017, 109, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Alazhari, M.; Sharma, T.; Heath, A.; Cooper, R.; Paine, K. Application of expanded perlite encapsulated bacteria and growth media for self-healing concrete. Constr. Build. Mater. 2017, 160, 610–619. [Google Scholar] [CrossRef]
- Xu, J.; Wang, X.; Wang, B. Biochemical process of ureolysis-based microbial CaCO3 precipitation and its application in self-healing concrete. Appl. Microbiol. Biotechnol. 2018, 102, 3121–3132. [Google Scholar] [CrossRef] [PubMed]
- Jonkers, H.M.; Thijssen, A.; Muyzer, G.; Copuroglu, O.; Schlangen, E. Application of bacteria as self-healing agent for the development of sustainable concrete. Ecol. Eng. 2010, 36, 230–235. [Google Scholar] [CrossRef]
- Heyrman, J.; Logan, N.; Rodrıgue-Dıaz, M.; Scheldeman, P.; Lebbe, L.; Swings, J.; Heyndrickx, M.; De Vos, P. Study of mural painting isolates, leading to the transfer of ‘Bacillus maroccanus’ and ‘Bacillus carotarum’ to Bacillus simplex, emended description of Bacillus simplex, a re-examination of the strains previously attributed to ‘Bacillus macroides’ and description of Bacillus muralis sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 119–131. [Google Scholar]
- Reeksting, B.J.; Hoffmann, T.D.; Tan, L.; Paine, K.; Gebhard, S. In-depth profiling of calcite precipitation by environmental bacteria reveals fundamental mechanistic differences with relevance to application. Appl. Environ. Microb. 2020, 86, e02739-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vahabi, A.; Ramezanianpour, A.A.; Akbari Noghabi, K. A preliminary insight into the revolutionary new line in improving concrete properties using an indigenous bacterial strain Bacillus licheniformis AK01, as a healing agent. Eur. J. Environ. Civil. Eng. 2014, 19, 614–627. [Google Scholar] [CrossRef]
- Enyedi, N.T.; Makk, J.; Kótai, L.; Berényi, B.; Klébert, S.; Sebestyén, Z.; Németh, P. Cave bacteria-induced amorphous calcium carbonate formation. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Cui, H.; Jiang, Z.; Liu, H.; He, H.; Fang, N. Biomineralization processes of calcite induced by bacteria isolated from marine sediments. Braz. J. Microbiol. 2015, 46, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Ercole, C.; Bozzelli, P.; Altieri, F.; Cacchio, P.; Del Gallo, M. Calcium carbonate mineralization: Involvement of extracellular polymeric materials isolated from calcifying bacteria. Microscop. Microanal. 2012, 18, 829–839. [Google Scholar] [CrossRef]
- Timperio, A.M.; Gorrasi, S.; Zolla, L.; Fenice, M. Evaluation of MALDI-TOF mass spectrometry and MALDI BioTyper in comparison to 16S rDNA sequencing for the identification of bacteria isolated from Arctic seawater. PLoS ONE 2007, 12, e0181860. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Ye, L.; Zhao, Q.; Ma, Y.; Yang, J.; Luo, J. Comparative study of MALDI-TOF MS and VITEK 2 in bacteria identification. J. Thorac. Dis. 2014, 6, 534–538. [Google Scholar]
- Abdel Samad, R.; Al Disi, Z.; Mohammad Ashfaq, M.Y.; Wahib, S.M.; Zouari, N. The use of principle component analysis and MALDI-TOF MS for the differentiation of mineral forming Virgibacillus and Bacillus species isolated from sabkhas. RSC Adv. 2020, 10, 14606–14616. [Google Scholar] [CrossRef] [Green Version]
- Whiffin, V.S. Microbial CaCO3 Precipitation for the Production of Biocement. Ph.D. Thesis, Murdoch University, Murdoch, Australia, 2004. [Google Scholar]
- Nakbanpote, W.; Panitlurtumpai, N.; Sangdee, A.; Sakulpone, N.; Sirisom, P.; Pimthong, A. Salt-tolerant and plant growth-promoting bacteria isolated from Zn/Cd contaminated soil: Identification and effect on rice under saline conditions. J. Plant. Inter. 2014, 9, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Orhan, F.; Gulluce, M. Isolation and characterization of salt-tolerant bacterial strains in salt-affected soils of erzurum, Turkey. Geomicrobiol. J. 2014, 32, 521–529. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, L.; Huang, X.; Liang, Z.; Liu, B.; Han, N.; Xing, F.; Deng, X. Enhanced calcite precipitation for crack healing by bacteria isolated under low-nitrogen conditions. Appl. Microbiol. Biotechnol. 2019, 103, 7971–7982. [Google Scholar] [CrossRef]
- Kim, G.; Kim, J.; Joun, H. Effect of temperature, pH, and reaction duration on microbially induced calcite precipitation. Appl. Sci. 2018, 8, 1277. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Satyam, N.; Reddy, K.R. Investigation of various gram-positive bacteria for MICP in Narmada Sand, India. Int. J. Geotech. Eng. 2019, 15, 220–234. [Google Scholar] [CrossRef]
- Fujita, Y.; Ferris, F.G.; Lawson, R.D.; Colwell, F.S.; Smith, R.W. Calcium carbonate precipitation by ureolytic subsurface bacteria. Geomicrobiol. J. 2000, 17, 305–318. [Google Scholar] [CrossRef]
- Okwadha, O.; Li, J. Optimum conditions for microbial carbonate precipitation. Chemosphere 2010, 81, 1143–1148. [Google Scholar] [CrossRef] [PubMed]
- Henry, D.; Watson, J.S.; John, C.M. Assessing and calibrating the ATR-FTIR approach as a carbonate rock characterization tool. Sediment. Geol. 2017, 347, 36–52. [Google Scholar] [CrossRef] [Green Version]
- Tepe, M.; Arslan, S.; Koralay, T.; MercanDogan, N. Precipitation and characterization of CaCO3 of Bacillus amyloliquefaciens U17 strain producing urease and carbonic anhydrase. Turk. J. Biol. 2019, 43, 198–208. [Google Scholar] [CrossRef]
- Hsu, C.M.; Huang, Y.H.; Nimje, V.; Lee, W.C.; Chen, H.J.; Kuo, Y.H.; Chen, C.Y. Comparative study on the sand bioconsolidation through calcium carbonate precipitation by Sporosarcina pasteurii and Bacillus subtilis. Crystals 2018, 8, 189. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Liu, Z. Influence of temperature on microbially induced calcium carbonate precipitation for soil treatment. PLoS ONE 2019, 14, e0218396. [Google Scholar] [CrossRef]
- Heveran, C.M.; Liang, L.; Nagarajan, A.; Hubler, M.; Gill, R.; Cameron, J.C.; Cook, S.M.; Srubar, W. Engineered ureolytic microorganisms can tailor the morphology and nanomechanical properties of microbial-precipitated calcium carbonate. Sci. Rep. 2019, 9, 14721. [Google Scholar] [CrossRef]
- Golovkina, D.A.; Zhurishkina, E.V.; Ivanova, L.A.; Baranchikov, A.E.; Sokolov, A.Y.; Bobrov, K.S.; Kulminskaya, A.A. Calcifying bacteria flexibility in induction of CaCO3 mineralization. Life 2020, 10, 317. [Google Scholar] [CrossRef]
- Ivanova, L.; Golovkina, D.; Zhurishkina, E.; Garmay, Y.; Baranchikov, A.; Tsvigun, N.; Zabrodskaya, Y.; Yapryntsev, A.; Gorshkov, A.; Lebedev, K.; et al. The matrix is everywhere: CaCO3 biomineralization by the Bacillus licheniformis planktonic cells. BioRxiv 2020. Available online: https://www.biorxiv.org/content/10.1101/2020.10.22.351619v1.full (accessed on 1 June 2021).
- Saracho, C.A.; Haigh, S.K.; Hata, T.; Soga, K.; Farsang, S.; Redfern, S.A.T.; Marek, E. Characterisation of CaCO3 phases during strain-specific ureolytic precipitation. Sci. Rep. 2020, 10, 1–12. [Google Scholar]
- Sheng Han, Y.; Hadiko, G.; Fuji, M.; Takahashi, M. Influence of initial CaCl2 concentration on the phase and morphology of CaCO3 prepared by carbonation. J. Mater. Sci. 2006, 41, 4663–4667. [Google Scholar]
- Donnelly, F.C.; Purcell-Milton, F.; Framont, V.; Cleary, O.; Dunne, P.W.; Gunko, Y.K. Synthesis of CaCO3 nano- and micro-particles by dry ice carbonation. Chem. Commun. 2017, 53, 6657–6660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, M.; Zuo, Y.; Chen, Z.; Yin, X.; Liu, Y.; Yang, W.; Chen, Y. Crystallization of CaCO3 in aqueous solutions with extremely high concentrations of NaCl. Crystals 2019, 9, 647. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Yu, J.; Zhao, X. Controlled synthesis of crystalline calcium carbonate aggregates with unusual morphologies involving the phase transformation from amorphous calcium carbonate. Mater. Res. Bull. 2009, 44, 831–835. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinetics of ureolysis (Section 3.4) | ||||||||
| Parameters | Coefficient | Referent strain | Natural isolates | |||||
| S. pasteurii | B. pseudofirmus | B. muralis | B. lentus | B. simplex | B. firmus | B. licheniformis | ||
| Urea concentration | kUrea | 0.10 | 0.00 | 0.02 | 0.01 | 0.03 | 0.03 | 0.04 |
| cUrea | 19.45 | 19.878 | 19.95 | 19.42 | 19.55 | 19.99 | 19.99 | |
| pH value | d | 9.15 | 7.24 | 8.87 | 8.14 | 9.2 | 9.14 | 8.812 |
| a | 7.19 | 7.19 | 7.27 | 7.36 | 7.45 | 7.19 | 7.2 | |
| c | 3.07 | 5.54 | 8.13 | 13.85 | 15.35 | 12.33 | 11.40 | |
| b | 1.37 | 5.28 | 7.43 | 2.99 | 4.01 | 2.37 | 15.30 | |
| Cell concentration | d | 9.90 | 6.68 | 6.567 | 8.11 | 8.18 | 8.6 | 11.66 |
| a | 5.55 | 5.05 | 5.61 | 6.3 | 5.75 | 6.2 | 5.03 | |
| c | 30.00 | 20.89 | 18.71 | 30.00 | 9.16 | 15.20 | 32.15 | |
| b | 0.79 | 1.96 | 4.49 | 1.78 | 4.93 | 2.95 | 0.8 | |
| Kinetic of the MICP process (Section 3.5) | ||||||||
| Parameters | Coefficient | S. pasteurii | B. pseudofirmus | B. muralis | B. lentus | B. simplex | B. firmus | B. licheniformis |
| Amount of precipitate | d | 0.46 | 0.001 | 0.57 | 0.21 | 0.56 | 0.47 | 0.51 |
| a | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | |
| c | 1.32 | 5.78 | 5.88 | 2.98 | 3.69 | 3.04 | 2.82 | |
| b | 4.09 | 2.56 | 2.06 | 5.97 | 1.22 | 3.75 | 6.00 | |
| pH value | d | 8.97 | 7.20 | 8.31 | 8.53 | 9.28 | 9.15 | 9.37 |
| a | 7.23 | 7.23 | 7.22 | 7.23 | 7.23 | 7.23 | 7.21 | |
| c | 0.50 | 1.78 | 1.07 | 0.89 | 1.01 | 0.95 | 1.01 | |
| b | 19.17 | 19.06 | 24.99 | 22.89 | 19.00 | 25.00 | 24.55 | |
| Calcium concentration | kprecipitate | 0.84 | 0.01 | 0.251 | 0.07 | 0.36 | 0.45 | 4.84 |
| cCa2+ | 1282.34 | 1237.18 | 1294.89 | 1146.27 | 1171.96 | 1340.89 | 1266.00 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šovljanski, O.; Pezo, L.; Stanojev, J.; Bajac, B.; Kovač, S.; Tóth, E.; Ristić, I.; Tomić, A.; Ranitović, A.; Cvetković, D.; et al. Comprehensive Profiling of Microbiologically Induced CaCO3 Precipitation by Ureolytic Bacillus Isolates from Alkaline Soils. Microorganisms 2021, 9, 1691. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9081691
Šovljanski O, Pezo L, Stanojev J, Bajac B, Kovač S, Tóth E, Ristić I, Tomić A, Ranitović A, Cvetković D, et al. Comprehensive Profiling of Microbiologically Induced CaCO3 Precipitation by Ureolytic Bacillus Isolates from Alkaline Soils. Microorganisms. 2021; 9(8):1691. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9081691
Chicago/Turabian StyleŠovljanski, Olja, Lato Pezo, Jovana Stanojev, Branimir Bajac, Sabina Kovač, Elvira Tóth, Ivan Ristić, Ana Tomić, Aleksandra Ranitović, Dragoljub Cvetković, and et al. 2021. "Comprehensive Profiling of Microbiologically Induced CaCO3 Precipitation by Ureolytic Bacillus Isolates from Alkaline Soils" Microorganisms 9, no. 8: 1691. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9081691