
The Role of Sch9 and the V-ATPase in the Adaptation Response to Acetic Acid and the Consequences for Growth and Chronological Lifespan

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Growth Conditions
2.2. Lipidomics
2.3. Chronological Lifespan Analysis
2.4. Medium Analysis
2.5. Intracellular pH Measurements
2.6. Analysis of Acetic Acid Diffusion
3. Results
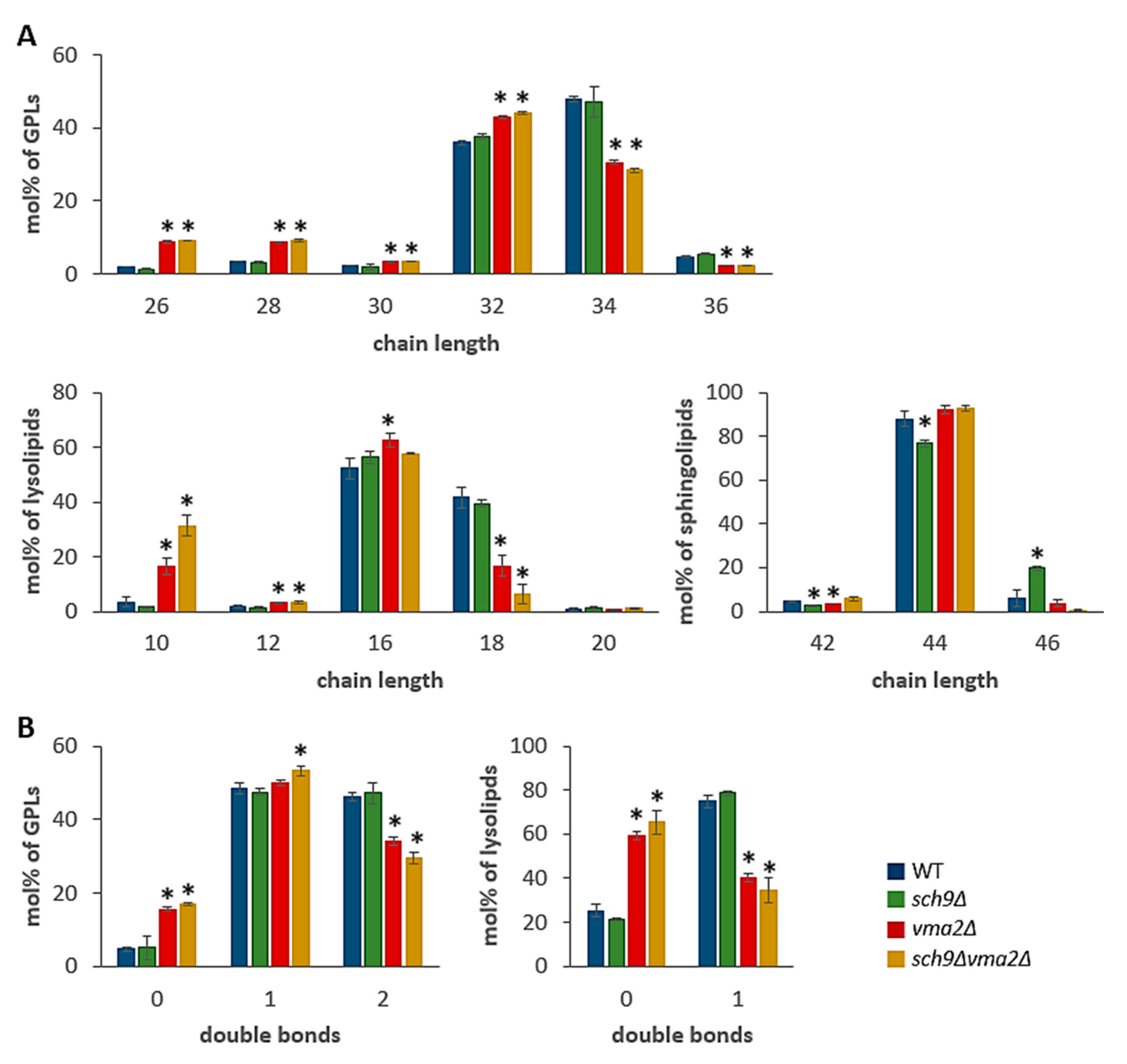
3.1. Loss of Sch9 or the V-ATPase Impacts on Lipid Homeostasis
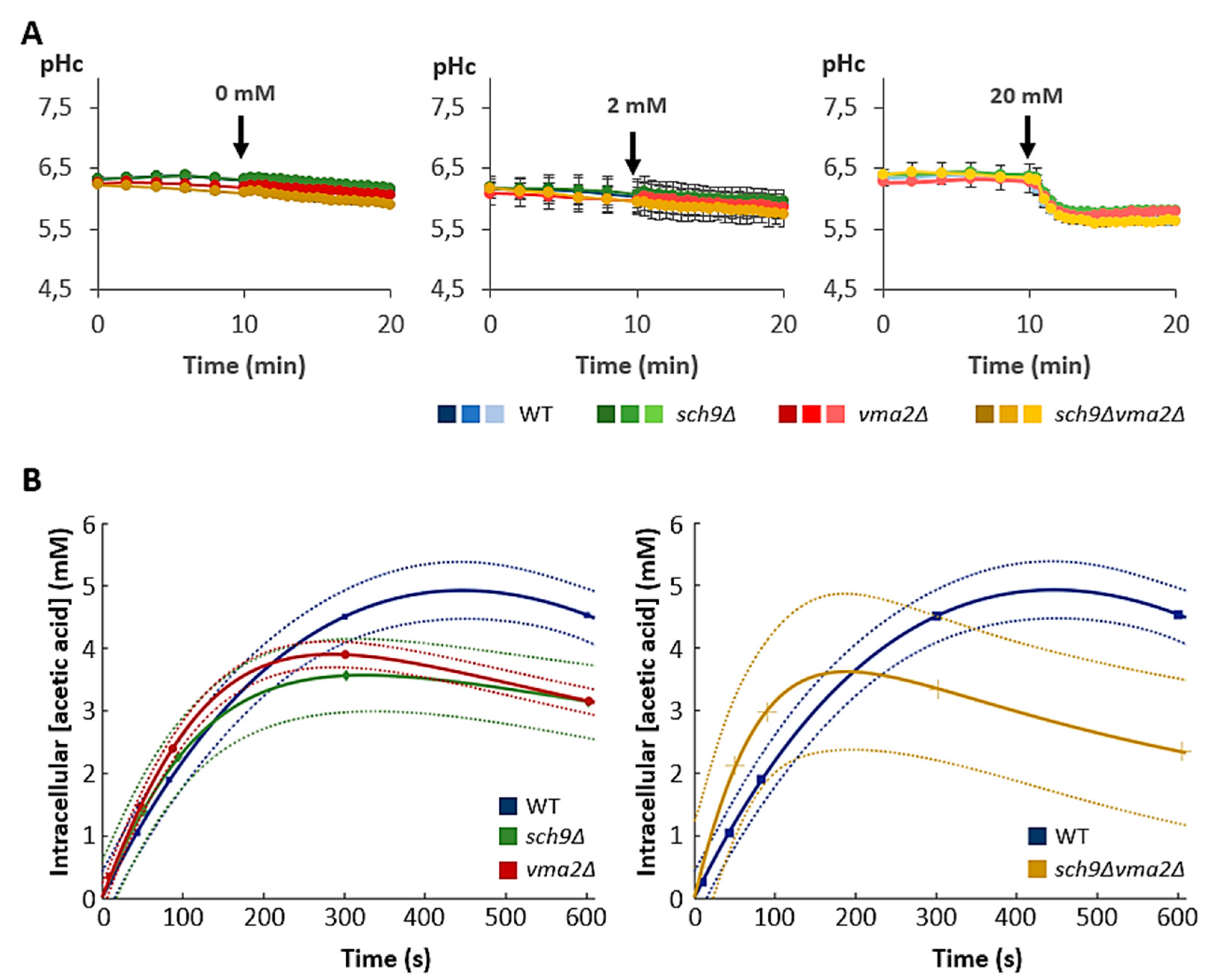
3.2. Loss of Sch9 or the V-ATPase Leads to Reduced Intracellular Acetic Acid Accumulation
3.3. A Functional V-ATPase Is Required to Adapt to Conditions of Extreme Acid Stress
3.4. The sch9Δ Strain Secretes More Acetic Acid
3.5. Cytoplasmic pH Correlates to Growth
3.6. The Addition of 20 mM Acetic Acid Increases Chronological Lifespan
4. Discussion
4.1. A Functional V-ATPase Is Required to Counteract Short-Term Effects of Acetic Acid Stress
4.2. The Long-Lived sch9Δ Strain Accumulates Acetic Acid and Triacylglycerol
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palma, M.; Guerreiro, J.F.; Sa-Correia, I. Adaptive Response and Tolerance to Acetic Acid in Saccharomyces cerevisiae and Zygosaccharomyces bailii: A Physiological Genomics Perspective. Front. Microbiol. 2018, 9, 274. [Google Scholar] [CrossRef]
- Casal, M.; Cardoso, H.; Leao, C. Mechanisms regulating the transport of acetic acid in Saccharomyces cerevisiae. Microbiology 1996, 142, 1385–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paiva, S.; Devaux, F.; Barbosa, S.; Jacq, C.; Casal, M. Ady2p is essential for the acetate permease activity in the yeast Saccharomyces cerevisiae. Yeast 2004, 21, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- dos Santos, M.M.; Gombert, A.K.; Christensen, B.; Olsson, L.; Nielsen, J. Identification of in vivo enzyme activities in the cometabolism of glucose and acetate by Saccharomyces cerevisiae by using 13C-labeled substrates. Eukaryot. Cell 2003, 2, 599–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warth, A.D. Effect of benzoic Acid on growth yield of yeasts differing in their resistance to preservatives. Appl. Environ. Microbiol. 1988, 54, 2091–2095. [Google Scholar] [CrossRef] [Green Version]
- Verduyn, C.; Postma, E.; Scheffers, W.A.; van Dijken, J.P. Energetics of Saccharomyces cerevisiae in anaerobic glucose-limited chemostat cultures. J. Gen. Microbiol. 1990, 136, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Pampulha, M.E.; Loureiro-Dias, M.C. Energetics of the effect of acetic acid on growth of Saccharomyces cerevisiae. FEMS Microbiol. Lett. 2000, 184, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Pampulha, M.E.; Loureiro-Dias, M.C. Combined effect of acetic acid, pH and ethanol on intracellular pH of fermenting yeast. Appl. Microbiol. Biotechnol. 1989, 31, 547–550. [Google Scholar] [CrossRef]
- Pampulha, M.E.; Loureiro-Dias, M.C. Activity of glycolytic enzymes of Saccharomyces cerevisiae in the presence of acetic acid. Appl. Microbiol. Biotechnol. 1990, 34, 375–380. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, Z.; Wang, M.; He, Q.; Zhang, H. The inhibition of Saccharomyces cerevisiae cells by acetic acid quantified by electrochemistry and fluorescence. Bioelectrochemistry 2008, 72, 117–121. [Google Scholar] [CrossRef]
- Burtner, C.R.; Murakami, C.J.; Kennedy, B.K.; Kaeberlein, M. A molecular mechanism of chronological aging in yeast. Cell Cycle 2009, 8, 1256–1270. [Google Scholar] [CrossRef] [Green Version]
- Mira, N.P.; Teixeira, M.C.; Sa-Correia, I. Adaptive response and tolerance to weak acids in Saccharomyces cerevisiae: A genome-wide view. OMICS 2010, 14, 525–540. [Google Scholar] [CrossRef] [Green Version]
- Giannattasio, S.; Guaragnella, N.; Zdralevic, M.; Marra, E. Molecular mechanisms of Saccharomyces cerevisiae stress adaptation and programmed cell death in response to acetic acid. Front. Microbiol. 2013, 4, 33. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A.; Orij, R.; Brul, S.; Smits, G.J. Quantitative analysis of the modes of growth inhibition by weak organic acids in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2012, 78, 8377–8387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stratford, M.; Nebe-von-Caron, G.; Steels, H.; Novodvorska, M.; Ueckert, J.; Archer, D.B. Weak-acid preservatives: pH and proton movements in the yeast Saccharomyces cerevisiae. Int. J. Food Microbiol. 2013, 161, 164–171. [Google Scholar] [CrossRef]
- Carmelo, V.; Santos, H.; Sa-Correia, I. Effect of extracellular acidification on the activity of plasma membrane ATPase and on the cytosolic and vacuolar pH of Saccharomyces cerevisiae. Biochim. Biophys. Acta. 1997, 1325, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Kawahata, M.; Masaki, K.; Fujii, T.; Iefuji, H. Yeast genes involved in response to lactic acid and acetic acid: Acidic conditions caused by the organic acids in Saccharomyces cerevisiae cultures induce expression of intracellular metal metabolism genes regulated by Aft1p. FEMS Yeast Res. 2006, 6, 924–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mira, N.P.; Becker, J.D.; Sa-Correia, I. Genomic expression program involving the Haa1p-regulon in Saccharomyces cerevisiae response to acetic acid. OMICS 2010, 14, 587–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Nino, M.; Marquina, M.; Swinnen, S.; Rodriguez-Porrata, B.; Nevoigt, E.; Arino, J. The Cytosolic pH of Individual Saccharomyces cerevisiae Cells Is a Key Factor in Acetic Acid Tolerance. Appl. Environ. Microbiol. 2015, 81, 7813–7821. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.R.; Mira, N.P.; Vargas, R.C.; Canelhas, I.; Sa-Correia, I. Saccharomyces cerevisiae adaptation to weak acids involves the transcription factor Haa1p and Haa1p-regulated genes. Biochem. Biophys. Res. Commun. 2005, 337, 95–103. [Google Scholar] [CrossRef]
- Mira, N.P.; Lourenco, A.B.; Fernandes, A.R.; Becker, J.D.; Sa-Correia, I. The RIM101 pathway has a role in Saccharomyces cerevisiae adaptive response and resistance to propionic acid and other weak acids. FEMS Yeast Res. 2009, 9, 202–216. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Ishii, Y.; Ogawa, J.; Shima, J. Enhancement of acetic acid tolerance in Saccharomyces cerevisiae by overexpression of the HAA1 gene, encoding a transcriptional activator. Appl. Environ. Microbiol. 2012, 78, 8161–8163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenreiro, S.; Rosa, P.C.; Viegas, C.A.; Sa-Correia, I. Expression of the AZR1 gene (ORF YGR224w), encoding a plasma membrane transporter of the major facilitator superfamily, is required for adaptation to acetic acid and resistance to azoles in Saccharomyces cerevisiae. Yeast 2000, 16, 1469–1481. [Google Scholar] [CrossRef]
- Tenreiro, S.; Nunes, P.A.; Viegas, C.A.; Neves, M.S.; Teixeira, M.C.; Cabral, M.G.; Sá-Correia, I. AQR1 gene (ORF YNL065w) encodes a plasma membrane transporter of the major facilitator superfamily that confers resistance to short-chain monocarboxylic acids and quinidine in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2002, 292, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Almeida, B.; Ohlmeier, S.; Almeida, A.J.; Madeo, F.; Leão, C.; Rodrigues, F.; Ludovico, P. Yeast protein expression profile during acetic acid-induced apoptosis indicates causal involvement of the TOR pathway. Proteomics 2009, 9, 720–732. [Google Scholar] [CrossRef]
- Hueso, G.; Aparicio-Sanchis, R.; Montesinos, C.; Lorenz, S.; Murguia, J.R.; Serrano, R. A novel role for protein kinase Gcn2 in yeast tolerance to intracellular acid stress. Biochem. J. 2012, 441, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Guerreiro, J.F.; Mira, N.P.; Santos, A.X.S.; Riezman, H.; Sa-Correia, I. Membrane Phosphoproteomics of Yeast Early Response to Acetic Acid: Role of Hrk1 Kinase and Lipid Biosynthetic Pathways, in Particular Sphingolipids. Front. Microbiol. 2017, 8, 1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godinho, C.P.; Prata, C.S.; Pinto, S.N.; Cardoso, C.; Bandarra, N.M.; Fernandes, F.; Sá-Correia, I. Pdr18 is involved in yeast response to acetic acid stress counteracting the decrease of plasma membrane ergosterol content and order. Sci. Rep. 2018, 8, 7860. [Google Scholar] [CrossRef]
- Guo, Z.P.; Khoomrung, S.; Nielsen, J.; Olsson, L. Changes in lipid metabolism convey acid tolerance in Saccharomyces cerevisiae. Biotechnol. Biofuels 2018, 11, 297. [Google Scholar] [CrossRef] [Green Version]
- Lindberg, L.; Santos, A.X.; Riezman, H.; Olsson, L.; Bettiga, M. Lipidomic profiling of Saccharomyces cerevisiae and Zygosaccharomyces bailii reveals critical changes in lipid composition in response to acetic acid stress. PLoS ONE 2013, 8, e73936. [Google Scholar] [CrossRef] [Green Version]
- Ludovico, P.; Rodrigues, F.; Almeida, A.; Silva, M.T.; Barrientos, A.; Corte-Real, M. Cytochrome c release and mitochondria involvement in programmed cell death induced by acetic acid in Saccharomyces cerevisiae. Mol. Biol. Cell 2002, 13, 2598–2606. [Google Scholar] [CrossRef] [Green Version]
- Giannattasio, S.; Guaragnella, N.; Corte-Real, M.; Passarella, S.; Marra, E. Acid stress adaptation protects Saccharomyces cerevisiae from acetic acid-induced programmed cell death. Gene 2005, 354, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Guaragnella, N.; Antonacci, L.; Passarella, S.; Marra, E.; Giannattasio, S. Hydrogen peroxide and superoxide anion production during acetic acid-induced yeast programmed cell death. Folia. Microbiol. (Praha) 2007, 52, 237–240. [Google Scholar] [CrossRef]
- Guaragnella, N.; Antonacci, L.; Giannattasio, S.; Marra, E.; Passarella, S. Catalase T and Cu, Zn-superoxide dismutase in the acetic acid-induced programmed cell death in Saccharomyces cerevisiae. FEBS Lett. 2008, 582, 210–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zdralevic, M.; Guaragnella, N.; Antonacci, L.; Marra, E.; Giannattasio, S. Yeast as a tool to study signaling pathways in mitochondrial stress response and cytoprotection. Sci. World J. 2012, 912147. [Google Scholar] [CrossRef] [Green Version]
- Semchyshyn, H.M.; Abrat, O.B.; Miedzobrodzki, J.; Inoue, Y.; Lushchak, V.I. Acetate but not propionate induces oxidative stress in bakers’ yeast Saccharomyces cerevisiae. Redox. Rep. 2011, 16, 15–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guaragnella, N.; Stirpe, M.; Marzulli, D.; Mazzoni, C.; Giannattasio, S. Acid Stress Triggers Resistance to Acetic Acid-Induced Regulated Cell Death through Hog1 Activation Which Requires RTG2 in Yeast. Oxid. Med. Cell Longev. 2019, 4651062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Wei, M.; Mirzaei, H.; Madia, F.; Mirisola, M.; Amparo, C.; Chagoury, S.; Kennedy, B.; Longo, V.D. Tor-Sch9 deficiency activates catabolism of the ketone body-like acetic acid to promote trehalose accumulation and longevity. Aging Cell 2014, 13, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Orlandi, I.; Ronzulli, R.; Casatta, N.; Vai, M. Ethanol and acetate acting as carbon/energy sources negatively affect yeast chronological aging. Oxid. Med. Cell Longev. 2013, 802870. [Google Scholar] [CrossRef] [Green Version]
- Yucel, E.B.; Eraslan, S.; Ulgen, K.O. The impact of medium acidity on the chronological life span of Saccharomyces cerevisiae-lipids, signaling cascades, mitochondrial and vacuolar functions. FEBS J. 2014, 281, 1281–1303. [Google Scholar] [CrossRef] [PubMed]
- Wilms, T.; Swinnen, E.; Eskes, E.; Dolz-Edo, L.; Uwineza, A.; Van Essche, R.; Rosseels, J.; Zabrocki, P.; Cameroni, E.; Franssens, V.; et al. The yeast protein kinase Sch9 adjusts V- ATPase assembly/ disassembly to control pH homeostasis and longevity in response to glucose availability. PLOS Genet. 2017, 13, e1006835. [Google Scholar] [CrossRef]
- Casatta, N.; Porro, A.; Orlandi, I.; Brambilla, L.; Vai, M. Lack of Sir2 increases acetate consumption and decreases extracellular pro-aging factors. Biochim. Biophys. Acta. 2013, 1833, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Swinnen, E.; Wilms, T.; Idkowiak-Baldys, J.; Smets, B.; De Snijder, P.; Accardo, S.; Ghillebert, R.; Thevissen, K.; Cammue, B.; De Vos, D.; et al. The protein kinase Sch9 is a key regulator of sphingolipid metabolism in Saccharomyces cerevisiae. Mol. Biol. Cell 2014, 25, 196–211. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.H.; Lester, R.L.; Dickson, R.C. Sphingolipid requirement for generation of a functional v1 component of the vacuolar ATPase. J. Biol. Chem. 2013, 278, 28872–28881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finnigan, G.C.; Ryan, M.; Stevens, T.H. A genome-wide enhancer screen implicates sphingolipid composition in vacuolar ATPase function in Saccharomyces cerevisiae. Genetics 2011, 187, 771–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tani, M.; Toume, M. Alteration of complex sphingolipid composition and its physiological significance in yeast Saccharomyces cerevisiae lacking vacuolar ATPase. Microbiology 2015, 161, 2369–2383. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Q.; Gamarra, S.; Garcia-Effron, G.; Park, S.; Perlin, D.S.; Rao, R. Requirement for ergosterol in V-ATPase function underlies antifungal activity of azole drugs. PLoS Pathog. 2010, 6, e1000939. [Google Scholar] [CrossRef] [PubMed]
- Klose, C.; Surma, M.A.; Gerl, M.J.; Meyenhofer, F.; Shevchenko, A.; Simons, K. Flexibility of a eukaryotic lipidome--insights from yeast lipidomics. PLoS ONE 2012, 7, e35063. [Google Scholar] [CrossRef] [Green Version]
- Ejsing, C.S.; Sampaio, J.L.; Surendranath, V.; Duchoslav, E.; Ekroos, K.; Klemm, R.W.; Simons, K.; Shevchenko, A. Global analysis of the yeast lipidome by quantitative shotgun mass spectrometry. Proc. Natl. Acad. Sci. USA 2009, 106, 2136–2141. [Google Scholar] [CrossRef] [Green Version]
- Surma, M.A.; Herzog, R.; Vasilj, A.; Klose, C.; Christinat, N.; Morin-Rivron, D.; Simons, K.; Masoodi, M.; Sampaio, J.L. An automated shotgun lipidomics platform for high throughput, comprehensive, and quantitative analysis of blood plasma intact lipids. Eur. J. Lipid. Sci. Technol. 2015, 117, 1540–1549. [Google Scholar] [CrossRef] [Green Version]
- Herzog, R.; Schuhmann, K.; Schwudke, D.; Sampaio, J.L.; Bornstein, S.R.; Schroeder, M.; Shevchenko, A. LipidXplorer: A software for consensual cross-platform lipidomics. PLoS ONE 2012, 7, e29851. [Google Scholar] [CrossRef] [Green Version]
- Brett, C.L.; Kallay, L.; Hua, Z.; Green, R.; Chyou, A.; Zhang, Y.; Graham, T.R.; Donowitz, M.; Rao, R. Genome-wide analysis reveals the vacuolar pH-stat of Saccharomyces cerevisiae. PLoS ONE 2011, 6, e17619. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, L.; Genheden, S.; Faria-Oliveira, F.; Allard, S.; Eriksson, L.A.; Olsson, L.; Bettiga, M. Alcohols enhance the rate of acetic acid diffusion in S. cerevisiae: Biophysical mechanisms and implications for acetic acid tolerance. Microb. Cell 2017, 5, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, L.; Genheden, S.; Eriksson, L.A.; Olsson, L.; Bettiga, M. Sphingolipids contribute to acetic acid resistance in Zygosaccharomyces bailii. Biotechnol. Bioeng. 2016, 113, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Xiang, T.X.; Anderson, B.D. Permeability of acetic acid across gel and liquid-crystalline lipid bilayers conforms to free-surface-area theory. Biophys. J. 1997, 72, 223–237. [Google Scholar] [CrossRef] [Green Version]
- Serrano, R.; Kielland-Brandt, M.C.; Fink, G.R. Yeast plasma membrane ATPase is essential for growth and has homology with (Na+ + K+), K+- and Ca2+-ATPases. Nature 1986, 319, 689–693. [Google Scholar] [CrossRef]
- Vallejo, C.G.; Serrano, R. Physiology of mutants with reduced expression of plasma membrane H+-ATPase. Yeast 1989, 5, 307–319. [Google Scholar] [CrossRef]
- Tarsio, M.; Zheng, H.; Smardon, A.M.; Martinez-Munoz, G.A.; Kane, P.M. Consequences of loss of Vph1 protein-containing vacuolar ATPases (V-ATPases) for overall cellular pH homeostasis. J. Biol. Chem. 2011, 286, 28089–28096. [Google Scholar] [CrossRef] [Green Version]
- Dechant, R.; Saad, S.; Ibanez, A.J.; Peter, M. Cytosolic pH Regulates Cell Growth through Distinct GTPases, Arf1 and Gtr1, to Promote Ras/PKA and TORC1 Activity. Mol. Cell 2014, 55, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Dechant, R.; Binda, M.; Lee, S.S.; Pelet, S.; Winderickx, J.; Peter, M. Cytosolic pH is a second messenger for glucose and regulates the PKA pathway through V-ATPase. Embo. J. 2010, 29, 2515–2526. [Google Scholar] [CrossRef] [Green Version]
- Orij, R.; Urbanus, M.L.; Vizeacoumar, F.J.; Giaever, G.; Boone, C.; Nislow, C.; Brul, S.; Smits, G.J. Genome-wide analysis of intracellular pH reveals quantitative control of cell division rate by pH(c) in Saccharomyces cerevisiae. Genome Biol. 2012, 13, 15. [Google Scholar] [CrossRef] [Green Version]
- Fabrizio, P.; Liou, L.L.; Moy, V.N.; Diaspro, A.; Valentine, J.S.; Gralla, E.B.; Longo, V.D. SOD2 functions downstream of Sch9 to extend longevity in yeast. Genetics 2003, 163, 35–46. [Google Scholar] [CrossRef]
- Deprez, M.A.; Eskes, E.; Wilms, T.; Ludovico, P.; Winderickx, J. pH homeostasis links the nutrient sensing PKA/TORC1/Sch9 menage-a-trois to stress tolerance and longevity. Microb. Cell 2018, 5, 119–136. [Google Scholar] [CrossRef]
- Holyoak, C.D.; Stratford, M.; McMullin, Z.; Cole, M.B.; Crimmins, K.; Brown, A.J.; Coote, P.J. Activity of the plasma membrane H(+)-ATPase and optimal glycolytic flux are required for rapid adaptation and growth of Saccharomyces cerevisiae in the presence of the weak-acid preservative sorbic acid. Appl. Environ. Microbiol. 1996, 62, 3158–3164. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Clapp, K.; Tarsio, M.; Kane, P.M. Interaction of the late endo-lysosomal lipid PI(3,5)P2 with the Vph1 isoform of yeast V-ATPase increases its activity and cellular stress tolerance. J. Biol. Chem. 2019, 294, 9161–9171. [Google Scholar] [CrossRef]
- Techo, T.; Jindarungrueng, S.; Tatip, S.; Limcharoensuk, T.; Pokethitiyook, P.; Kruatrachue, M.; Auesukaree, C. Vacuolar H(+) -ATPase is involved in preventing heavy metal-induced oxidative stress in Saccharomyces cerevisiae. Environ. Microbiol. 2020, 22, 2403–2418. [Google Scholar] [CrossRef]
- Marek, A.; Korona, R. Restricted pleiotropy facilitates mutational erosion of major life-history traits. Evolution 2013, 67, 3077–3086. [Google Scholar] [CrossRef]
- Burtner, C.R.; Murakami, C.J.; Olsen, B.; Kennedy, B.K.; Kaeberlein, M. A genomic analysis of chronological longevity factors in budding yeast. Cell Cycle 2011, 10, 1385–1396. [Google Scholar] [CrossRef] [Green Version]
- Handee, W.; Li, X.; Hall, K.W.; Deng, X.; Li, P.; Benning, C.; Williams, B.L.; Kuo, M.-H. An Energy-Independent Pro-longevity Function of Triacylglycerol in Yeast. PLoS Genet. 2016, 12, e1005878. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Kane, P.M. Direct interaction of the Golgi V-ATPase a-subunit isoform with PI(4)P drives localization of Golgi V-ATPases in yeast. Mol. Biol. Cell 2017, 28, 2518–2530. [Google Scholar] [CrossRef]
- Banerjee, S.; Kane, P.M. Regulation of V-ATPase Activity and Organelle pH by Phosphatidylinositol Phosphate Lipids. Front. Cell Dev. Biol. 2020, 8, 510. [Google Scholar] [CrossRef]
- Vasanthakumar, T.; Bueler, S.A.; Wu, D.; Beilsten-Edmands, V.; Robinson, C.V.; Rubinstein, J.L. Structural comparison of the vacuolar and Golgi V-ATPases from Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2019, 116, 7272–7277. [Google Scholar] [CrossRef] [Green Version]
- Dolz-Edo, L.; van der Deen, M.; Brul, S.; Smits, G.J. Caloric restriction controls stationary phase survival through Protein Kinase A (PKA) and cytosolic pH. Aging Cell 2019, 18, e12921. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deprez, M.-A.; Maertens, J.M.; Olsson, L.; Bettiga, M.; Winderickx, J. The Role of Sch9 and the V-ATPase in the Adaptation Response to Acetic Acid and the Consequences for Growth and Chronological Lifespan. Microorganisms 2021, 9, 1871. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091871
Deprez M-A, Maertens JM, Olsson L, Bettiga M, Winderickx J. The Role of Sch9 and the V-ATPase in the Adaptation Response to Acetic Acid and the Consequences for Growth and Chronological Lifespan. Microorganisms. 2021; 9(9):1871. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091871
Chicago/Turabian StyleDeprez, Marie-Anne, Jeroen M. Maertens, Lisbeth Olsson, Maurizio Bettiga, and Joris Winderickx. 2021. "The Role of Sch9 and the V-ATPase in the Adaptation Response to Acetic Acid and the Consequences for Growth and Chronological Lifespan" Microorganisms 9, no. 9: 1871. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091871