
RTG Signaling Sustains Mitochondrial Respiratory Capacity in HOG1-Dependent Osmoadaptation

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Growth Conditions
2.2. Micro- and Batch-Culture Growth Assays
2.3. Spotting Assay
2.4. Quantitative PCR (qPCR)
2.5. Oxygen Consumption Measurements
2.6. Statistical Analysis
3. Results
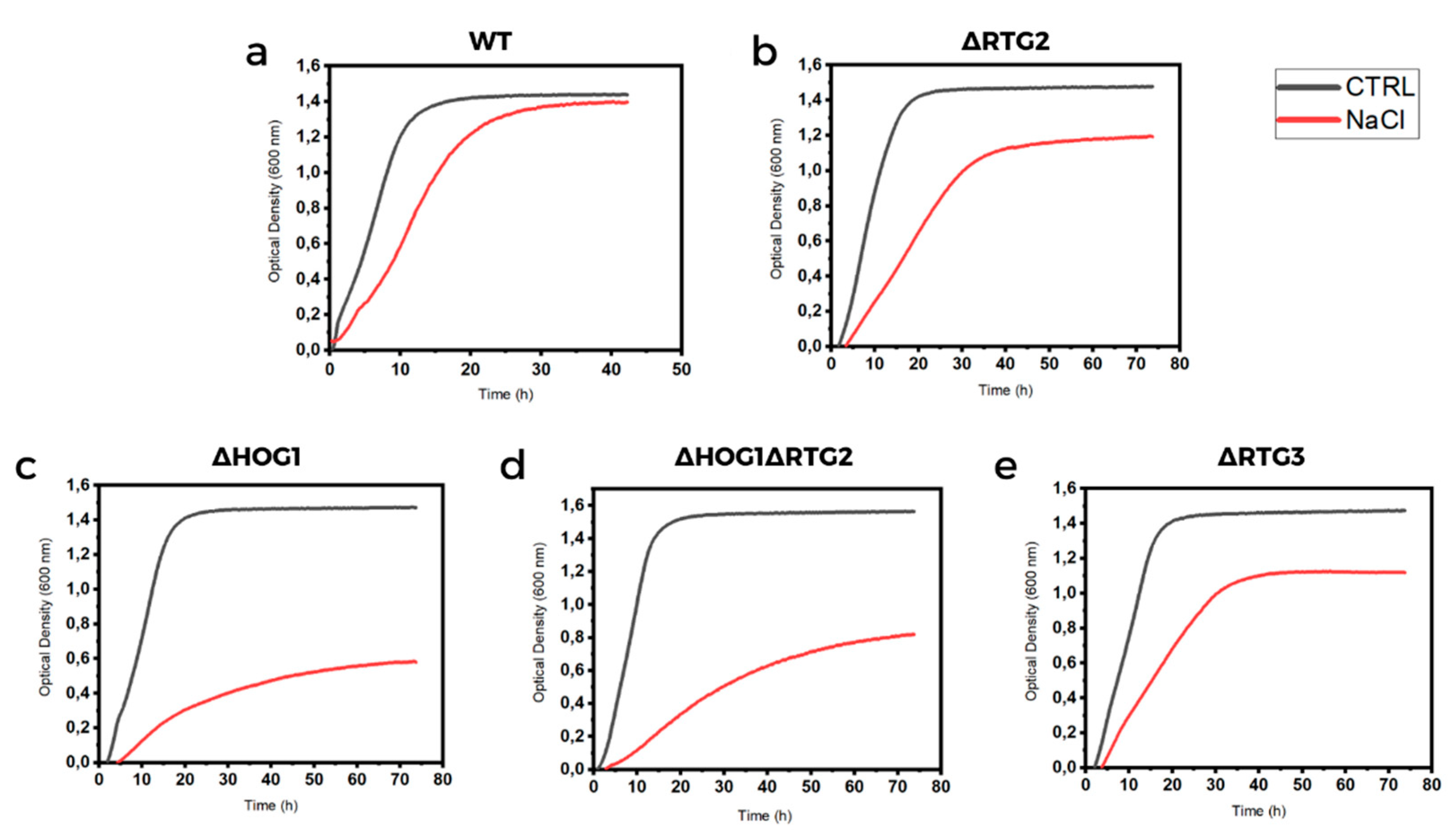
3.1. Cell Sensitivity to Osmotic Stress Due to NaCl Treatment
3.2. Impairment of RTG Signaling Sensitizes Cells to High Osmostress in an HOG1-Dependent Manner
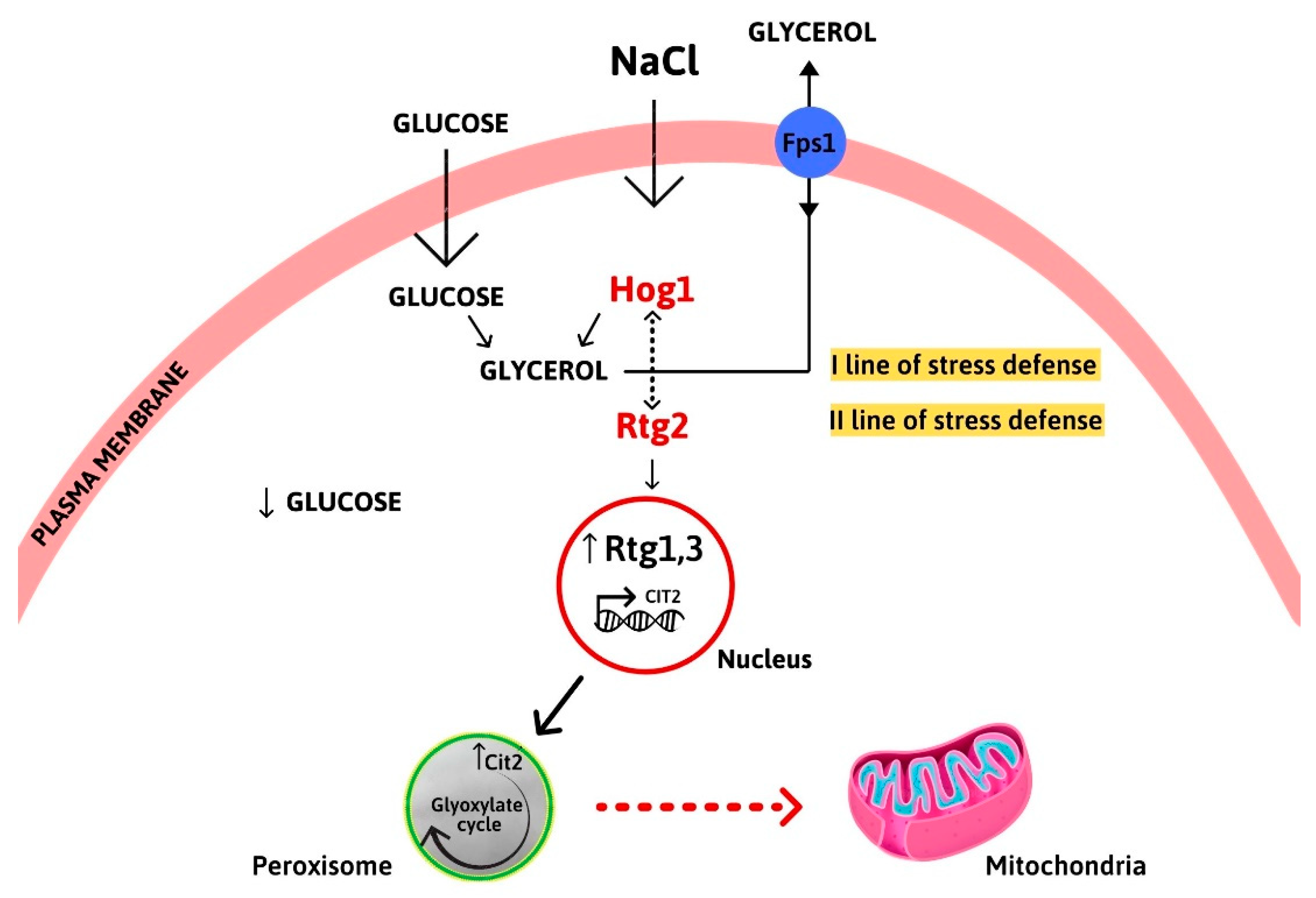
3.3. RTG Signaling Is Activated in Osmoadaptation
3.4. RTG Mutants Show Decreased Respiratory Capacity under Osmostress
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Z.; Butow, R.A. Mitochondrial retrograde signaling. Annu. Rev. Genet. 2006, 40, 159–185. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Butow, R.A. A transcriptional switch in the expression of yeast tricarboxylic acid cycle genes in response to a reduction or loss of respiratory function. Mol. Cell Biol. 1999, 19, 6720–6728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guaragnella, N.; Ždralević, M.; Palková, Z.; Giannattasio, S. Analysis of mitochondrial retrograde signaling in yeast model systems. Methods Mol. Biol. 2021, 2276, 87–102. [Google Scholar] [CrossRef]
- Guaragnella, N.; Ždralević, M.; Lattanzio, P.; Marzulli, D.; Pracheil, T.; Liu, Z.; Passarella, S.; Marra, E.; Giannattasio, S. Yeast growth in raffinose results in resistance to acetic-acid induced programmed cell death mostly due to the activation of the mitochondrial retrograde pathway. Biochim. Biophys. Acta 2013, 1833, 2765–2774. [Google Scholar] [CrossRef] [Green Version]
- Guaragnella, N.; Stirpe, M.; Marzulli, D.; Mazzoni, C.; Giannattasio, S. Acid stress triggers resistance to acetic acid-induced regulated cell death through Hog1 activation which requires RTG2 in yeast. Oxid. Med. Cell Longev. 2019, 2019, 4651062. [Google Scholar] [CrossRef] [Green Version]
- Hijazi, I.; Knupp, J.; Chang, A. Retrograde signaling mediates an adaptive survival response to endoplasmic reticulum stress in Saccharomyces cerevisiae. J. Cell Sci. 2020, 133, jcs241539. [Google Scholar] [CrossRef]
- Torelli, N.Q.; Ferreira-Júnior, J.R.; Kowaltowski, A.J.; da Cunha, F.M. RTG1- and RTG2-dependent retrograde signaling controls mitochondrial activity and stress resistance in Saccharomyces cerevisiae. Free Radic Biol. Med. 2015, 81, 30–37. [Google Scholar] [CrossRef]
- Ruiz-Roig, C.; Noriega, N.; Duch, A.; Posas, F.; de Nadal, E. The Hog1 SAPK controls the Rtg1/Rtg3 transcriptional complex activity by multiple regulatory mechanisms. Mol. Biol. Cell 2012, 23, 4286–4296. [Google Scholar] [CrossRef]
- Guaragnella, N.; Coyne, L.P.; Chen, X.J.; Giannattasio, S. Mitochondria-cytosol-nucleus crosstalk: Learning from Saccharomyces cerevisiae. FEMS Yeast Res. 2018, 18. [Google Scholar] [CrossRef]
- Saito, H.; Posas, F. Response to hyperosmotic stress. Genetics 2012, 192, 289–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor, M.M.; Proft, M.; Pascual-Ahuir, A. Mitochondrial function is an inducible determinant of osmotic stress adaptation in yeast. J. Biol. Chem. 2009, 284, 30307–30317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohmann, S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parmar, J.H.; Bhartiya, S.; Venkatesh, K.V. Characterization of the adaptive response and growth upon hyperosmotic shock in Saccharomyces cerevisiae. Mol. Biosyst. 2011, 7, 1138–1148. [Google Scholar] [CrossRef] [PubMed]
- Brewster, J.L.; Gustin, M.C. Hog1: 20 years of discovery and impact. Sci. Signal. 2014, 7, re7. [Google Scholar] [CrossRef] [PubMed]
- Klipp, E.; Nordlander, B.; Krüger, R.; Gennemark, P.; Hohmann, S. Integrative model of the response of yeast to osmotic shock. Nat. Biotechnol. 2005, 23, 975–982. [Google Scholar] [CrossRef]
- Lee, B.; Jeong, S.-G.; Jin, S.H.; Mishra, R.; Peter, M.; Lee, C.-S.; Lee, S.S. Quantitative analysis of yeast MAPK signaling networks and crosstalk using a microfluidic device. Lab Chip 2020, 20, 2646–2655. [Google Scholar] [CrossRef]
- Toussaint, M.; Conconi, A. High-throughput and sensitive assay to measure yeast cell growth: A bench protocol for testing genotoxic agents. Nat. Protoc. 2006, 1, 1922–1928. [Google Scholar] [CrossRef]
- Manzanares-Estreder, S.; Espí-Bardisa, J.; Alarcón, B.; Pascual-Ahuir, A.; Proft, M. Multilayered control of peroxisomal activity upon salt stress in Saccharomyces cerevisiae. Mol. Microbiol. 2017, 104, 851–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarcia, P.; Agrimi, G.; Germinario, L.; Ibrahim, A.; Rottensteiner, H.; Palmieri, F.; Palmieri, L. In Saccharomyces cerevisiae grown in synthetic minimal medium supplemented with non-fermentable carbon sources glutamate is synthesized within mitochondria. Rend. Fis. Acc. Lincei 2018, 29, 483–490. [Google Scholar] [CrossRef]
- Di Noia, M.A.; Todisco, S.; Cirigliano, A.; Rinaldi, T.; Agrimi, G.; Iacobazzi, V.; Palmieri, F. The human SLC25A33 and SLC25A36 genes of solute carrier family 25 encode two mitochondrial pyrimidine nucleotide transporters. J. Biol. Chem. 2014, 289, 33137–33148. [Google Scholar] [CrossRef] [Green Version]
- Di Bartolomeo, F.; Malina, C.; Campbell, K.; Mormino, M.; Fuchs, J.; Vorontsov, E.; Gustafsson, C.M.; Nielsen, J. Absolute yeast mitochondrial proteome quantification reveals trade-off between biosynthesis and energy generation during diauxic shift. Proc. Natl. Acad. Sci. USA 2020, 117, 7524–7535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costanzo, M.; Kuzmin, E.; van Leeuwen, J.; Mair, B.; Moffat, J.; Boone, C.; Andrews, B. Global genetic networks and the genotype-to-phenotype relationship. Cell 2019, 177, 85–100. [Google Scholar] [CrossRef] [Green Version]
- DeRisi, J.L.; Iyer, V.R.; Brown, P.O. Exploring the metabolic and genetic control of gene expression on a genomic scale. Science 1997, 278, 680–686. [Google Scholar] [CrossRef] [Green Version]
- Rajvanshi, P.K.; Arya, M.; Rajasekharan, R. The stress-regulatory transcription factors Msn2 and Msn4 regulate fatty acid oxidation in budding yeast. J. Biol. Chem. 2017, 292, 18628–18643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laera, L.; Guaragnella, N.; Ždralević, M.; Marzulli, D.; Liu, Z.; Giannattasio, S. The transcription factors ADR1 or CAT8 are required for RTG pathway activation and evasion from yeast acetic acid-induced programmed cell death in raffinose. Microb. Cell 2016, 3, 621–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trendeleva, T.A.; Zvyagilskaya, R.A. Retrograde signaling as a mechanism of yeast adaptation to unfavorable factors. Biochemistry 2018, 83, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Knupp, J.; Arvan, P.; Chang, A. Increased mitochondrial respiration promotes survival from endoplasmic reticulum stress. Cell Death Differ. 2019, 26, 487–501. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Pastor, M.; Proft, M.; Pascual-Ahuir, A. Adaptive changes of the yeast mitochondrial proteome in response to salt stress. Omics 2010, 14, 541–552. [Google Scholar] [CrossRef]
- Petelenz-Kurdziel, E.; Kuehn, C.; Nordlander, B.; Klein, D.; Hong, K.-K.; Jacobson, T.; Dahl, P.; Schaber, J.; Nielsen, J.; Hohmann, S.; et al. Quantitative analysis of glycerol accumulation, glycolysis and growth under hyper osmotic stress. PLoS Comput. Biol. 2013, 9, e1003084. [Google Scholar] [CrossRef]
- You, T.; Ingram, P.; Jacobsen, M.D.; Cook, E.; McDonagh, A.; Thorne, T.; Lenardon, M.D.; de Moura, A.P.S.; Romano, M.C.; Thiel, M.; et al. A systems biology analysis of long and short-term memories of osmotic stress adaptation in fungi. BMC Res. Notes 2012, 5, 258. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast Strains | Specific Growth Rate (µmax h−1) | Doubling Time (h) | Growth Efficiency (%) |
|---|---|---|---|
| WT | 0.38 ± 0.02 | 2.0 ± 0.32 | - |
| WT + NaCl | 0.17 ± 0.01 | 3.9 ± 0.71 | 87 ± 2 |
| ∆RTG2 | 0.39 ± 0.03 | 2.2 ± 0.07 | - |
| ∆RTG2 + NaCl | 0.12 ± 0.01 | 5.9 ± 0.12 | 80 ± 4 |
| ∆HOG1 | 0.32 ± 0.01 | 2.1 ± 0.4 | - |
| ∆HOG1 + NaCl | 0.08 ± 0.02 | 10.3 ± 0.2 | 44 ± 9 |
| ∆HOG1∆RTG2 | 0.29 ± 0.09 | 2.5 ± 0.78 | - |
| ∆HOG1∆RTG2 + NaCl | 0.12 ± 0.01 | 7.2 ± 0.2 | 56 ± 12 |
| ∆RTG3 | 0.37 ± 0.04 | 2.5 ± 0.14 | - |
| ∆RTG3 + NaCl | 0.12 ± 0.01 | 5.4 ± 0.71 | 80 ± 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guaragnella, N.; Agrimi, G.; Scarcia, P.; Suriano, C.; Pisano, I.; Bobba, A.; Mazzoni, C.; Palmieri, L.; Giannattasio, S. RTG Signaling Sustains Mitochondrial Respiratory Capacity in HOG1-Dependent Osmoadaptation. Microorganisms 2021, 9, 1894. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091894
Guaragnella N, Agrimi G, Scarcia P, Suriano C, Pisano I, Bobba A, Mazzoni C, Palmieri L, Giannattasio S. RTG Signaling Sustains Mitochondrial Respiratory Capacity in HOG1-Dependent Osmoadaptation. Microorganisms. 2021; 9(9):1894. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091894
Chicago/Turabian StyleGuaragnella, Nicoletta, Gennaro Agrimi, Pasquale Scarcia, Clelia Suriano, Isabella Pisano, Antonella Bobba, Cristina Mazzoni, Luigi Palmieri, and Sergio Giannattasio. 2021. "RTG Signaling Sustains Mitochondrial Respiratory Capacity in HOG1-Dependent Osmoadaptation" Microorganisms 9, no. 9: 1894. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091894