
Whole-Genome Sequencing (WGS) of Carbapenem-Resistant K. pneumoniae Isolated in Long-Term Care Facilities in the Northern Italian Region

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Setting
2.2. Strains Selection
2.3. DNA Extraction and Whole-Genome Sequencing (WGS)
2.4. Bioinformatic Analysis
3. Results
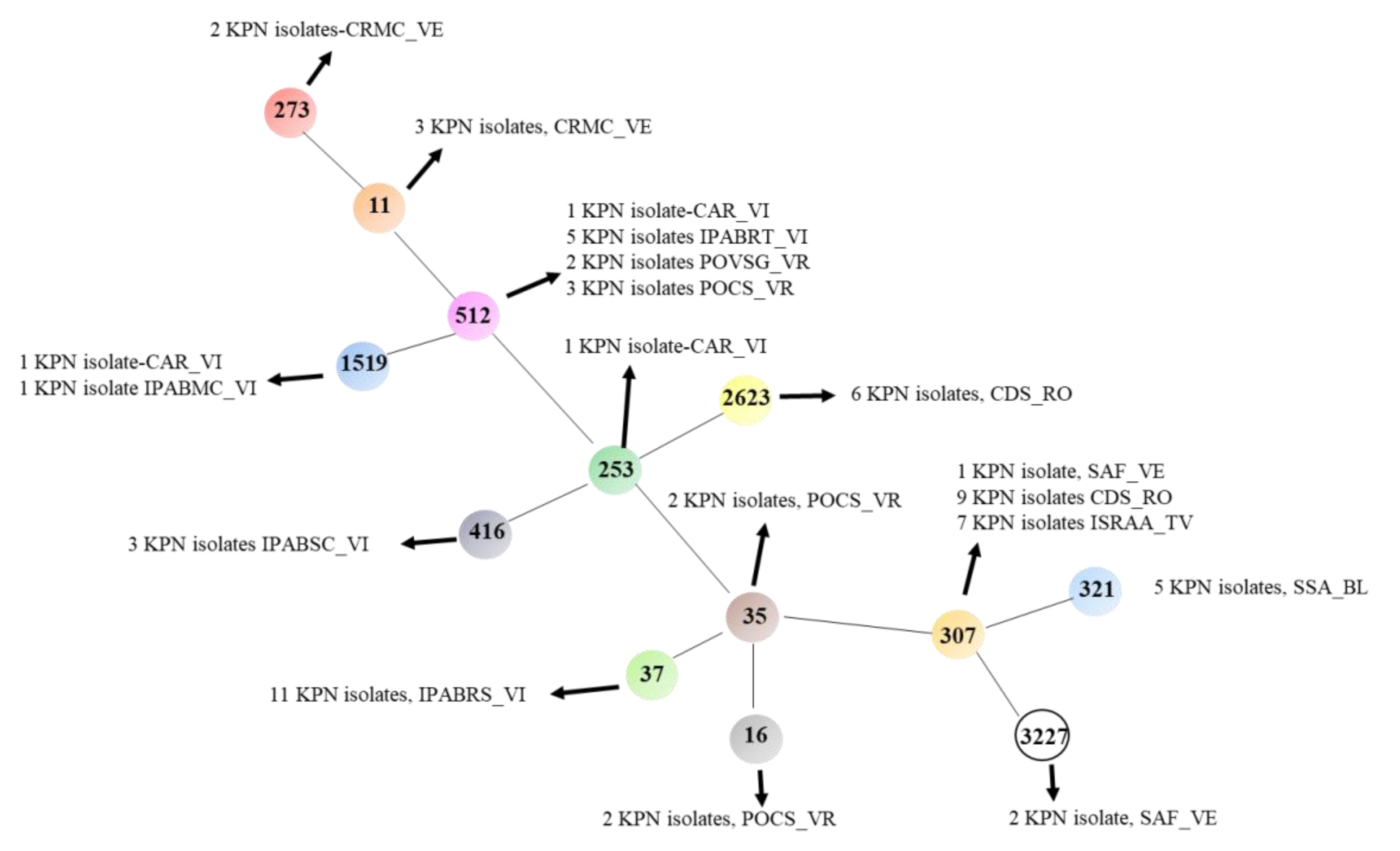
3.1. WGS of K. pneumoniae
3.2. K. pneumoniae MLST
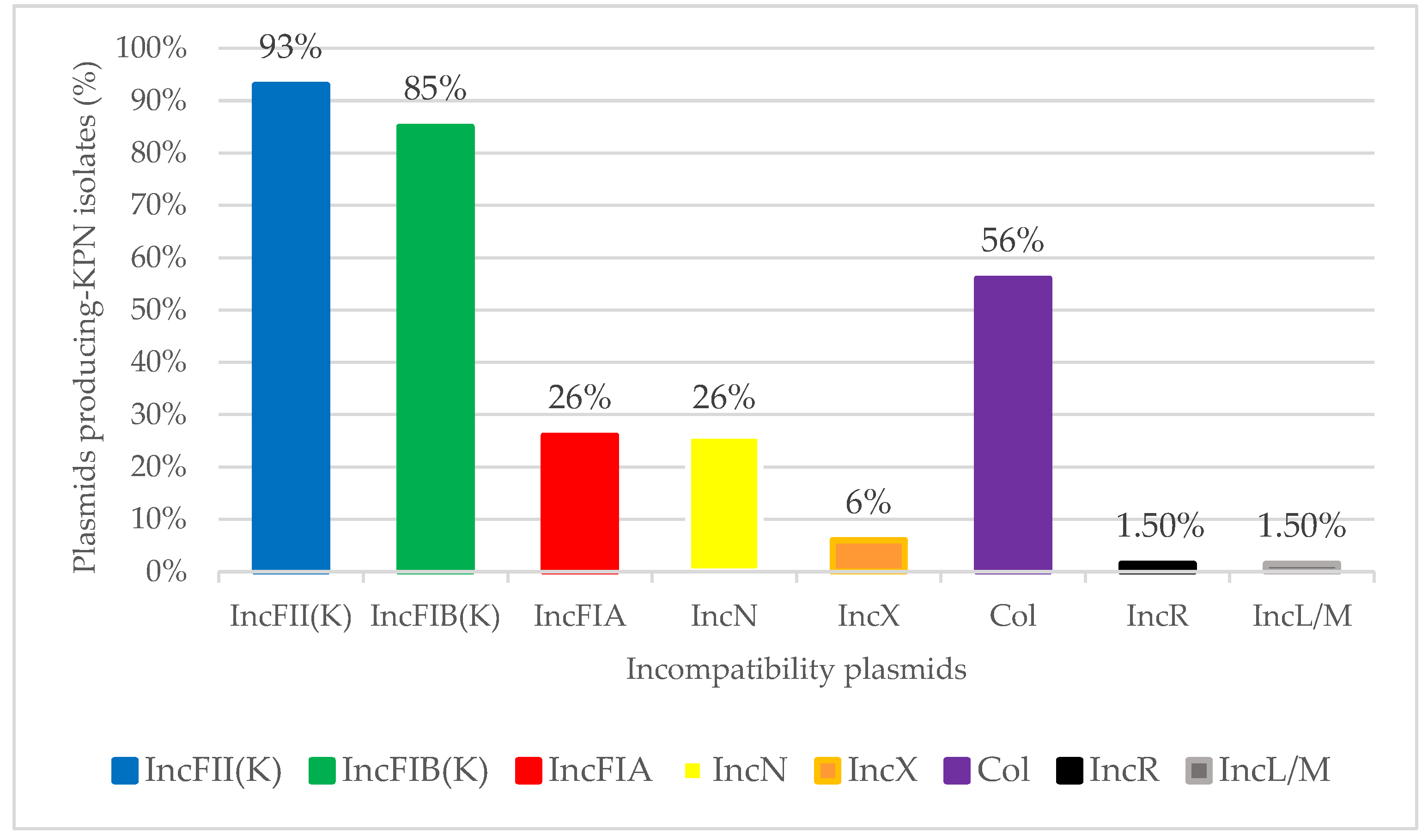
3.3. Plasmids Replicons and pMLST
3.4. β-Lactam Resistance Genes
3.5. Fluoroquinolone Resistance Genes
3.6. Other Antimicrobial Resistance Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bush, K.; Bradford, P.A. β-Lactams and β-lactamase inhibitors: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef]
- King, D.T.; Sobhanifar, S.; Strynadka, N. The mechanisms of resistance to β-lactam antibiotics. In Handbook of Antimicrobial Resistance; Gotte, M., Berghuis, A., Eds.; Springer: New York, NY, USA, 2014; pp. 1–22. [Google Scholar]
- Bush, K.; Bradford, P.A. Epidemiology of β-Lactamase-Producing Pathogens. Clin. Microbiol. Rev. 2020, 33, e00047-19. [Google Scholar] [CrossRef]
- Naas, T.; Vandel, L.; Sougakoff, W.; Livermore, D.M.; Nordmann, P. Cloning and sequence analysis of the gene for a carbapenem-hydrolyzing class A beta-lactamase, Sme-1, from Serratia marcescens S6. Antimicrob. Agents Chemother. 1994, 38, 1262–1270. [Google Scholar] [CrossRef] [Green Version]
- Bush, K. Past and Present Perspectives on β-Lactamases. Antimicrob. Agents Chemother. 2018, 62, e01076-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Centre for Disease Prevention and Control. Antimicrobial Resistance in the EU/EEA (EARS-Net)-Annual Epidemiological Report 2019; ECDC: Stockholm, Sweden, 2020. [Google Scholar]
- World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. 2017. Available online: https://www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf (accessed on 25 February 2017).
- David, S.; Reuter, S.; Harris, S.R.; Glasner, C.; Feltwell, T.; Argimon, S.; Abudahab, K.; Goater, R.; Giani, T.; Errico, G.; et al. Epidemic of carbapenem-resistant Klebsiella pneumoniae in Europe is driven by nosocomial spread. Nat. Microbiol. 2019, 4, 1919–1929. [Google Scholar] [CrossRef]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile β-Lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madni, O.; Amoako, D.G.; Abia, A.L.K.; Rout, J.; Essack, S.Y. Genomic investigation of carbapenem-resistant Klebsiella pneumoniae colonization in an intensive care unit in South Africa. Genes 2021, 12, 951. [Google Scholar] [CrossRef]
- Zhan, Q.; Xu, Y.; Wang, B.; Yu, J.; Shen, X.; Liu, L.; Cao, X.; Guo, Y.; Yu, F. Distribution of fluoroquinolone resistance determinants in carbapenem-resistant Klebsiella pneumoniae clinical isolates associated with bloodstream infections in China. BMC Microbiol. 2021, 21, 164. [Google Scholar] [CrossRef]
- Ahmed, M.A.E.E.; Yang, Y.; Yang, Y.; Yan, B.; Chen, G.; Hassan, R.M.; Zhong, L.L.; Chen, Y.; Roberts, A.P.; Wu, Y.; et al. Emergence of iypervirulent carbapenem-resistant Klebsiella pneumoniae coharboring a bla(NDM-1)-carrying virulent plasmid and a blaKPC-2)-carrying plasmid in an Egyptian Hospital. mSphere. 2021, 6, e00088-21. [Google Scholar] [CrossRef]
- Barbarini, D.; Russello, G.; Brovarone, F.; Capatti, C.; Colla, R.; Perilli, M.; Moro, M.L.; Carretto, E. Evaluation of carbapenem-resistant Enterobacteriaceae in an Italian setting: Report from the trench. Infect. Genet. Evol. 2015, 30, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Brescini, L.; Morroni, G.; Valeriani, C.; Castelletti, S.; Mingoia, M.; Simoni, S.; Masucci, A.; Montalti, R.; Vivarelli, M.; Giacometti, A.; et al. Clinical and epidemiological characteristics of KPC-producing Klebsiella pneumoniae from bloodstream infections in a tertiary referral center in Italy. BMC Infect. Dis. 2019, 19, 611. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.; Phan, H.T.T.; Findlay, J.; Stoesser, N.; Pankhurst, L.; Navickaite, I.; De Maio, N.; Eyre, D.W.; Toogood, G.; Orsi, N.M.; et al. Covert dissemination of carbapenemase-producing Klebsiella pneumoniae (KPC) in a successfully controlled outbreak: Long- and short-read whole-genome sequencing demonstrate multiple genetic modes of transmission. J. Antimicrob. Chemother. 2017, 72, 3025–3034. [Google Scholar] [CrossRef]
- Naas, T.; Cuzon, G.; Villegas, M.V.; Lartigue, M.F.; Quinn, J.P.; Nordmann, P. Genetic structures at the origin of acquisition of the β-lactamase blaKPC gene. Antimicrob. Agents. Chemother. 2008, 52, 1257–1263. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Fernandez, S.; Villa, L.; Carta, C.; Venditti, C.; Giordano, A.; Venditti, M.; Mancini, C.; Carattoli, A. Klebsiella pneumoniae ST258 producing KPC-3 identified in Italy carries novel plasmids and OmpK36/OmpK35 porin variants. Antimicrob. Agents. Chemother. 2012, 56, 2143–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Chavda, K.D.; Melano, R.G.; Jacobs, M.R.; Levi, M.H.; Bonomo, R.A.; Kreiswirth, B.N. Complete sequence of a bla(KPC-2)-harboring IncFII(K1) plasmid from a Klebsiella pneumoniae sequence type 258 strain, Antimicrob. Agents. Chemother. 2013; 57, 1542–1545. Agents. Chemother. 2013, 57, 1542–1545. [Google Scholar] [CrossRef] [Green Version]
- Cerdeira, L.T.; Cunha, M.P.V.; Francisco, G.R.; Bueno, M.F.C.; Araujo, B.F.; Ribas, R.M.; Gontijo-Filho, P.P.; Knöbl, T.; de Oliveira Garcia, D.; Lincopan, N. IncX3 plasmid harboring a non-Tn4401 genetic element (NTEKPC) in a hospital-associated clone of KPC-2-producing Klebsiella pneumoniae ST340/CG258. Diagn. Microbiol. Infect. Dis. 2017, 89, 164–167. [Google Scholar] [CrossRef]
- Endimiani, A.; Depasquale, J.M.; Forero, S.; Perez, F.; Hujer, A.M.; Roberts-Pollack, D.; Fiorella, P.D.; Pickens, N.; Kitchel, B.; Casiano-Colón, A.E.; et al. Emergence of blaKPC-containing Klebsiella pneumoniae in a long-term acute care hospital: A new challenge to our healthcare system. J. Antimicrob. Chemother. 2009, 64, 1102–1110. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, D.; Masarwa, S.; Navon-Venezia, S.; Mishali, H.; Fridental, I.; Rubinovitch, B.; Smollan, G.; Carmeli, Y.; Schwaber, M.J.; Israel PACF CRKP (Post-Acute-Care Facility Carbapenem-Resistant Klebsiella pneumoniae) Working Group. Carbapenem-resistant Klebsiella pneumonia in post-acute-care facilities in Israel. Infect. Control Hosp. Epidemiol. 2011, 32, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Giani, T.; Antonelli, A.; Caltagirone, M.; Mauri, C.; Nicchi, J.; Arena, F.; Nucleo, E.; Bracco, S.; Pantosti, A.; AMCLI-CoSA survey participants; et al. Evolving β-lactamase epidemiology in Enterobacteriaceae from Italian nationwide surveillance, October 2013: KPC-carbapenemase spreading among outpatients. Euro. Surveill. 2017, 22, 30583. [Google Scholar] [CrossRef]
- Chen, H.Y.; Jean, S.S.; Lee, Y.L.; Lu, M.C.; Ko, W.C.; Liu, P.Y.; Hsueh, P.R. Carbapenem-Resistant Enterobacterales in Long-Term Care Facilities: A Global and Narrative Review. Front. Cell Infect. Microbiol. 2021, 11, 601968. [Google Scholar] [CrossRef]
- Lapp, Z.; Crawford, R.; Miles-Jay, A.; Pirani, A.; Trick, W.E.; Weinstein, R.A.; Hayden, M.K.; Snitkin, E.S.; Lin, M.Y. Regional spread of blaNDM-1-containing Klebsiella pneumoniae ST147 in post-acute care facilities. Clin. Infect. Dis. 2021, ciab457. [Google Scholar] [CrossRef] [PubMed]
- Ambretti, S.; Bassetti, M.; Clerici, P.; Petrosillo, N.; Tumietto, F.; Viale, P.; Rossolini, G.M. Screening for carriage of carbapenem-resistant Enterobacteriaceae in settings of high endemicity: A position paper from an Italian working group on CRE infections. Antimicrob. Resist. Infect. Control 2019, 8, 136. [Google Scholar] [CrossRef]
- Arena, F.; Vannetti, F.; Di Pilato, V.; Fabbri, L.; Colavecchio, O.L.; Giani, T.; Marraccini, C.; Pupillo, R.; Macchi, C.; Converti, F.; et al. Diversity of the epidemiology of carbapenemase-producing Enterobacteriaceae in long-term acute care rehabilitation settings from an area of hyperendemicity, and evaluation of an intervention bundle. J. Hosp. Infect. 2018, 100, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Bonura, C.; Giuffre, M.; Aleo, A.; Fasciana, T.; Di Bernardo, F.; Stampone, T.; Giammanco, A.; Palma, D.M.; Mammina, C.; MDR-GN Working Group. An update of the evolving epidemic of blaKPC carrying Klebsiella pneumoniae in Sicily, Italy, 2014: Emergence of multiple non-ST258 clones. PLoS ONE 2015, 10, e0132936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arena, F.; Di Pilato, V.; Vannetti, F.; Fabbri, L.; Antonelli, A.; Coppi, M.; Pupillo, R.; Macchi, C.; Rossolini, G.M. Population structure of KPC carbapenemase-producing Klebsiella pneumoniae in a long-term acute-care rehabilitation facility: Identification of a new lineage of clonal group 101, associated with local hyperendemicity. Microb. Genom. 2020, 6, e000308. [Google Scholar] [CrossRef] [PubMed]
- Zerbino, D.R. Using the Velvet de novo assembler for short-read sequencing technologies. Curr. Protoc. Bioinformatics 2010, 11, 11.5.1–11.5.12. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.V.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Pontén, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of methods for genomic taxonomy. J. Clin. Microbiol. 2014, 52, 1529–1539. [Google Scholar] [CrossRef] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents. Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yang, Y.; Chen, G.; Lin, M.; Chen, Y.; He, R.; Galvão, K.N.; El-Gawad El-Sayed Ahmed, M.A.; Roberts, A.P.; Wu, Y.; et al. Molecular characterization of carbapenem-resistant and virulent plasmids in Klebsiella pneumoniae from patients with bloodstream infections in China. Emerg. Microbes. Infect. 2021, 10, 700–709. [Google Scholar] [CrossRef]
- Magi, G.; Tontarelli, F.; Caucci, S.; Sante, L.D.; Brenciani, A.; Morroni, G.; Giovanetti, E.; Menzo, S.; Mingoia, M. High prevalence of carbapenem-resistant Klebsiella pneumoniae ST307 recovered from fecal samples in an Italian hospital. Future Microbiol. 2021, 16, 703–711. [Google Scholar] [CrossRef]
- Giani, T.; Pini, B.; Arena, F.; Conte, V.; Bracco, S.; Migliavacca, R.; AMCLI-CRE Survey Participants; Pantosti, A.; Pagani, L.; Luzzaro, F.; et al. Epidemic diffusion of KPC carbapenemase-producing Klebsiella pneumoniae in Italy: Results of the first countrywide survey, 15 May to 30 June 2011. Euro. Surveill. 2013, 18, 20489. [Google Scholar] [CrossRef]
- Conte, V.; Monaco, M.; Giani, T.; D’Ancona, F.; Moro, M.L.; Arena, F.; D’Andrea, M.M.; Rossolini, G.M.; Pantosti, A.; AR-ISS Study Group on Carbapenemase-Producing K. pneumonia. Molecular epidemiology of KPC-producing Klebsiella pneumoniae from invasive infections in Italy: Increasing diversity with predominance of the ST512 clade II sublineage. J. Antimicrob. Chemother. 2016, 71, 3386–3391. [Google Scholar]
- Peirano, G.; Chen, L.; Kreiswirth, B.N.; Pitout, J.D.D. Emerging Antimicrobial-Resistant High-Risk Klebsiella pneumoniae Clones ST307 and ST147. Antimicrob. Agents. Chemother. 2020, 64, e01148-20. [Google Scholar] [CrossRef] [PubMed]
- Baek, E.H.; Kim, S.E.; Kim, S.; Lee, S.; Cho, O.H.; In Hong, S.; Shin, J.H.; Hwang, I. Successful control of an extended-spectrum β-lactamase-producing Klebsiella pneumoniae ST307 outbreak in a neonatal intensive care unit. BMC Infect. Dis. 2020, 20, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonstra, M.B.; Spijkerman, D.C.M.; Voor In ‘t Holt, A.F.; van der Laan, R.J.; Bode, L.G.M.; van Vianen, W.; Klaassen, C.H.W.; Vos, M.C.; Severin, J.A. An outbreak of ST307 extended-spectrum beta-lactamase (ESBL)-producing Klebsiella pneumoniae in a rehabilitation center: An unusual source and route of transmission. Infect. Control. Hosp. Epidemiol. 2020, 41, 31–36. [Google Scholar] [CrossRef]
- Giufre, M.; Accogli, M.; Ricchizzi, E.; Barbanti, F.; Farina, C.; Fazii, P.; Mattei, R.; Sarti, M.; Barozzi, A.; Buttazzi, R.; et al. Multidrug-resistant infections in long-term care facilities: Extended-spectrum beta-lactamase-producing Enterobacteriaceae and hypervirulent antibiotic resistant Clostridium difficile. Diagn. Microbiol. Infect. Dis. 2018, 91, 275–281. [Google Scholar] [CrossRef]
- Wyres, K.L.; Hawkey, J.; Hetland, M.A.K.; Fostervold, A.; Wick, R.R.; Judd, L.M.; Hamidian, M.; Howden, B.P.; Lohr, I.H.; Holt, K.E. Emergence and rapid global dissemination of CTX-M-15-associated Klebsiella pneumoniae strain ST307. J. Antimicrob. Chemother. 2019, 74, 577–581. [Google Scholar] [CrossRef] [Green Version]
- Piccirilli, A.; Perilli, M.; Piccirilli, V.; Segatore, B.; Amicosante, G.; Maccacaro, L.; Bazaj, A.; Naso, L.; Lo Cascio, G.; Cornaglia, G. Molecular characterization of carbapenem-resistant Klebsiella pneumoniae ST14 and ST512 causing bloodstream infections in ICU and surgery wards of a tertiary university hospital of Verona (northern Italy): Co-production of KPC-3, OXA-48, and CTX-M-15 β-lactamases. Diagn. Microbiol. Infect. Dis. 2020, 96, 114968. [Google Scholar]
- Perilli, M.; Bottoni, C.; Pontieri, E.; Segatore, B.; Celenza, G.; Setacci, D.; Bellio, P.; Strom, R.; Amicosante, G. Emergence of blaKPC-3-Tn4401a in Klebsiella pneumoniae ST512 in the municipal wastewater treatment plant and in the university hospital of a town in central Italy. J. Glob. Antimicrob. Resist. 2013, 1, 217–220. [Google Scholar] [CrossRef]
- Perilli, M.; Bottoni, C.; Grimaldi, A.; Segatore, B.; Celenza, G.; Mariani, M.; Bellio, P.; Frascaria, P.; Amicosante, G. Carbapenem-resistant Klebsiella pneumoniae harbouring blaKPC-3 and blaVIM-2 from central Italy. Diagn. Microbiol. Infect. Dis. 2013, 75, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Geraci, D.M.; Bonura, C.; Giuffrè, M.; Saporito, L.; Graziano, G.; Aleo, A.; Fasciana, T.; Di Bernardo, F.; Stampone, T.; Palma, D.M.; et al. Is the monoclonal spread of the ST258, KPC-3-producing clone being replaced in southern Italy by the dissemination of multiple clones of carbapenem-nonsusceptible, KPC-3-producing Klebsiella pneumoniae? Clin. Microbiol. Infect. 2015, 21, e15–e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcìa-Fernandez, A.; Miriagou, V.; Papagiannitsis, C.C.; Giordano, A.; Venditti, M.; Mancini, C.; Carattoli, A. An ertapenem-resistant extended-spectrum-β-lactamase-producing Klebsiella pneumoniae clone carries a novel OmpK36 porin variant. Antimicrob. Agents. Chemother. 2010, 54, 4178–4184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orsi, G.B.; Bencardino, A.; Vena, A.; Carattoli, A.; Venditti, C.; Falcone, M.; Giordano, A.; Venditti, M. Patient risk factors for outer membrane permeability and KPC-producing carbapenem-resistant Klebsiella pneumoniae isolation: Results of a double case-control study. Infection 2013, 41, 61–67. [Google Scholar] [CrossRef]
- Oliveira, É.M.; Beltrão, E.M.B.; Scavuzzi, A.M.L.; Barros, J.F.; Lopes, A.C.S. High plasmid variability, and the presence of IncFIB, IncQ, IncA/C, IncHI1B, and IncL/M in clinical isolates of Klebsiella pneumoniae with blaKPC and blaNDM from patients at a public hospital in Brazil. Rev. Soc. Bras. Med. Trop. 2020, 53, e20200397. [Google Scholar] [CrossRef] [PubMed]
- Villa, L.; García-Fernández, A.; Fortini, D.; Carattoli, A. Replicon sequence typing of IncF plasmids carrying virulence and resistance determinants. J. Antimicrob. Chemother. 2010, 65, 2518–2529. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, M.S.; Bonomo, R.A.; Tolmasky, M.E. Carbapenemases: Transforming Acinetobacter baumannii into a yet more dangerous menace. Biomolecules 2020, 10, 720. [Google Scholar] [CrossRef]
- Arabacı, Ç.; Dal, T.; Başyiğit, T.; Genişel, N.; Durmaz, R. Investigation of carbapenemase and mcr-1 genes in carbapenem-resistant Klebsiella pneumoniae isolates. J. Infect. Dev. Ctries. 2019, 13, 504–509. [Google Scholar] [CrossRef]
- Chen, L.; Chavda, K.D.; Melano, R.G.; Hong, T.; Rojtman, A.D.; Jacobs, M.R.; Bonomo, R.A.; Kreiswirth, B.N. Molecular survey of the dissemination of two blaKPC-harboring IncFIA plasmids in New Jersey and New York hospitals. Antimicrob. Agents. Chemother. 2014, 58, 2289–2294. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| LTCFs | No. Isolates | ST | Resistance Profile |
|---|---|---|---|
| CDS_RO | 15 | 307, 2623 | AMX, TZP, CTX, CAZ, ERT, MEM, CIP |
| POCS_VR | 2 | 16 | AMX, TZP, CTX, CAZ, ERT, MEM, CIP, SXT |
| 2 | 35 | AMX, TZP, CTX, CAZ, CIP, MEM, SXT | |
| 3 | 512 | AMX, TZP, CTX, CAZ, ERT, MEM, CIP | |
| POVSG_VR | 2 | 512 | AMX, TZP, CTX, CAZ, ERT, MEM, CIP |
| IPABMC_VI | 2 | 253, 1519 | AMX, TZP, CTX, CAZ, ERT, MEM, CIP, SXT |
| CAR_VI | 2 | 512, 1519 | AMX, TZP, CTX, CAZ, ERT, MEM, CIP, SXT |
| IPABRS_VI | 11 | 37 | AMX, TZP, CTX, CAZ, ERT, MEM, CIP, SXT |
| IPABRT_VI | 5 | 512 | AMX, TZP, CTX, CAZ, ERT, MEM, CIP, SXT |
| IPABSC_VI | 3 | 416 | AMX, TZP, CTX, CAZ, MEM, CIP, SXT |
| SSA_BL | 5 | 321 | AMX, TZP, CTX, CAZ, MEM, CIP, SXT |
| ISRAA_TV | 7 | 307 | AMX, TZP, CTX, CAZ, ERT, MEM, CIP, SXT |
| CRMC_VE | 5 | 11, 273 | AMX, TZP, CTX, CAZ, ERT, MEM, CIP, SXT |
| SAF_VE | 4 | 307 | AMX, TZP, CTX, CAZ, ERT, MEM, CIP, SXT |
| LTCFs | No. Isolates | Genome Size (bp) | MLST | Plasmid Replicons/pMLST | β-lactams Resistance Genes | Other Antibiotics Resistance Genes |
|---|---|---|---|---|---|---|
| SSA_BL | 5 | 5,736,085 | ST321 | IncFII(K), IncFIB(K), IncFIA(HI1), IncN/fii_k1 fia_19 | blaVIM-1, blaSHV-99 | aadA1, aph(3′)-XV, aacA4, fosA, mph(A), aac(6′)Ib-cr,oqxA, oqxB, qnrS1, sul1 dfrA14, catB2 |
| ISRAA_TV | 2 | 5,497,503 | ST307 | IncFIB(K), IncFII(K)/fii_k7 | blaSHV-28, blaKPC-2, blaOXA-9, blaTEM-1A, blaCTX-M-15 | fosA, dfrA14, qnrB66 |
| 2 | 5,164,710 | ST307 | IncFIB(K), IncFII(K)/fii_k7 | blaKPC-2, blaTEM-1A, blaOXA-9, blaCTX-M-15, blaSHV-28, blaOXA-1 | aac(6′)-Ib-cr, fosA, dfrA14, qnrB66, catB4 | |
| 2 | 5,578,258 | ST307 | IncFIB(K), IncFII(K)/fii_k7 | blaKPC-2, blaTEM-1A, blaOXA-9, blaCTX-M-15, blaSHV-28, blaOXA-1 | aac(6′)-Ib-cr, fosA, dfrA14, qnrB66, catB4, oqxA, oqxB | |
| 1 | 5,505,674 | ST307 | IncFIB(K), IncFII(K)/fii_k7 | blaKPC-2, blaTEM-1A, blaOXA-9, blaCTX-M-15, blaSHV-28, blaOXA-1 | aac(6′)-Ib-cr, fosA, dfrA14, catB4, oqxA, oqxB | |
| SAF_VE | 1 | 5,545,376 | ST307 | IncFIB(K), IncFII(K)/fii_k7 | blaKPC-2, blaTEM-1A, blaOXA-9, blaCTX-M-15, blaSHV-28, blaOXA-1 | aac(6′)-Ib-cr, fosA, dfrA14, qnrB66, catB4, oqxA, oqxB |
| 1 | 5,330,546 | ST3227 | IncFIB(pQil), IncFIB(K)/fii_k7 | blaKPC-9, blaTEM-1B, blaOXA-9, blaLEN-12, blaCTX-M-15, blaOXA-1 | aph(3′)-IIa, aac(6′)-Ib-cr, fosA, strA, strB, sul2, dfrA14, qnrB66, catB4 | |
| 1 | 5,545,568 | ST3227 | IncFIB(pQil), IncFII(K), IncFIB(K)/fii_k9 | blaOXA-9, blaTEM-1A, blaKPC-3, blaLEN-12, blaLEN-7, blaOXA-1 | aac(6′)-Ib-cr, aph(3′)-IIa, fosA, dfrA14, oqxB, qnrB66, catB4 | |
| 1 | 5,540,988 | ST307 | IncFII(K), IncFIB(K), IncR, IncL/M (pOXA-48)/fii_k5 | blaTEM-1A, blaSHV-11 | aph(3′)-Ia, fosA, qnrB19, oqxB | |
| CRMC_VE | 3 | 5,531,831 | ST11 | IncFII(pKPX1), ColpVC, Col(MG828)/fii_k7 | blaSHV-11, blaCTX-M-15, blaNDM-1 | aac(6′)-Ib-cr, fosA, oqxB, oqxA |
| 2 | 5,387,384 | ST273 | IncFII(K), IncFIB(K), Col(MG828)/fii_k7 | blaSHV-11, blaTEM-1B, blaOXA-1, blaCTX-M-15 | aac(3′)-IIa, aac(6′)-Ib-cr, strA, strB, fosA, sul2, dfrA14, oqxA, oqxB, catB4 | |
| CDS_RO | 3 | 5,424,562 | ST2623 | IncFII(K), IncFIB(K), IncFIA(HI1), ColRNAI/fii_k1 fia_10 | blaCTX-M-15, blaSHV-1, blaTEM-1B, blaOXA-1 | aac(6′)-Ib-cr, strA, strB, sul2 oqxA, oqxB, qnrB66, tet(D), catB4 |
| 3 | 5,666,417 | ST 2623 | IncFIB(pQil), IncFII(K), IncFIB(K), IncFIA(HI1), ColRNAI/fii_k1 fia_10 | blaCTX-M-15, blaOXA-9, blaTEM-1B, blaOXA-1, blaKPC-3 | aac(6′)-Ib-cr, fosA, strA, strB, sul2, oqxA, oqxB, qnrB66, tet(D), catB4 | |
| 2 | 5,607,404 | ST307 | IncFIB(pQil), IncFII(K), IncFIA(HI1)/fii_k2 fia_19 | blaTEM-1A, blaOXA-9, blaSHV-28, blaCTX-M-15, blaOXA-1, blaKPC-3 | aac(6′)-Ib-cr, strA, strB, fosA, sul2, oqxA, oqxB, qnrB66, catB4 | |
| 37 | 5,496,232 | ST307 | IncFIB(pQil), IncFII(K)/fii_k2 | blaCTX-M-15, blaTEM-1A, blaOXA-9, blaSHV-28, blaOXA-1, blaKPC-3 | aac(6′)-Ib-cr, fosA, sul2, oqxA, oqxB, qnrB66, catB4 | |
| 2 | 5,566,824 | ST307 | IncFIB(pQil), IncFII(K), IncFIA(HI1)/fii_k2 fia_19 | blaCTX-M-15, blaTEM-1A, blaOXA-9, blaSHV-28, blaOXA-1, blaKPC-3 | aac(6′)-Ib-cr, strA, strB, fosA, sul2, oqxA, oqxB qnrB66, catB4 | |
| 2 | 5,634,072 | ST307 | IncFIB(pQil), IncFII(K), IncFIB(K)/fii_k2 | blaCTX-M-15, blaTEM-1A, blaOXA-9, blaSHV-28, blaOXA-1, blaKPC-3 | aac(6′)-Ib-cr, aac(3)-IIa, strA, strB, fosA, sul2, dfrA14, oqxA, oqxB, qnrB66, catB4 | |
| CAR_VI | 1 | 5,649,046 | ST512 | IncX3, IncFIB(K), IncFIB(pQil) IncFII(K)/fii_k2 | blaKPC-3, blaTEM-1A, blaOXA-9, blaSHV-11 | aadA2, aph(3′)-Ia, fosA, mph(A), sul1, dfrA12, oqxA, oqxB, catA1 |
| 1 | 5,487,176 | ST1519 | ColRNAI, IncFIB(pQil), IncFII(K)/fii_k2 | blaKPC-3, blaOXA-9, blaSHV-11, blaTEM-1A | aadA2, aac(6′)-Ib-cr, fosA, sul1, oqxA, oqxB | |
| IPABMC_VI | 1 | 5,311,196 | ST1519 | ColRNAI, IncFIB(pQil), IncFII(K), IncFIB(K)/fii_k2 | blaSHV-11, blaKPC-3 | aac(6′)-Ib-cr, aadA2, fosA, mph(A), sul1, dfrA12, oqxB, catA1 |
| 1 | 5,651,358 | ST253 | ColRNAI, IncFIB(pQil), IncFII(K), IncFIB(K), IncN/ fii_k5/ST7 | blaVIM-1, blaSHV-36 | aac(6′)-Ib-cr, aadA1, aacA4, fosA, sul1, dfrA14, qnrS1, catB2 | |
| IPABRS_VI | 2 | 5,167,604 | ST37 | ColRNAI, IncFII(K), IncFIB(K)/fii_k7 | blaCTX-M-15, blaSHV-11, blaOXA-1, blaTEM-1B | aac(6′)-Ib-cr, strA, strB, fosA, sul2, dfrA14, oqxA, oqxB, catB4 |
| 4 | 5,451,323 | ST37 | ColRNAI, IncFIB(pQil), IncFII(K), IncFIB(K), IncN/ fii_k7 | blaSHV-11, blaOXA-1, blaKPC-2 | aac(6′)-Ib-cr, fosA, dfrA14, dfrA30, oqxB, oqxA, catB4 | |
| 3 | 5,424,901 | ST37 | ColRNAI, IncFIB(pQil), IncFII(K), IncFIB(K), IncN/ IncF: fii_k7 IncN: unknown | blaSHV-11, blaOXA-1, blaKPC-2 | aac(6′)-Ib-cr, fosA, dfrA30, dfrA14, oqxA, oqxB, catB4 | |
| 2 | 5,444,858 | ST37 | ColRNAI, IncFIB(pQil), IncFII(K), IncFIB(K), IncN/ IncF: fii_k7 | blaSHV-11, blaOXA-1, blaKPC-2 | aac(6′)-Ib-cr, fosA, dfrA30, dfrA14, oqxA, oqxB, catB4 | |
| IPABRT_VI | 5 | 5,406,932 | ST512 | ColRNAI, IncFIB(pQil), IncFII(K), IncFIB(K)/fii_k2 | blaLEN-12, blaTEM-1A, blaKPC-3, blaOXA-9 | aac(6′)-Ib-cr, aadA1, fosA, sul1, dfrA1, oqxB, tet(D) |
| IPABSC_VI | 1 | 5,634,812 | ST416 | IncFII(K), IncFIA(HI1), ColRNAI, IncN, IncFIB(K)/ IncF: fii_k4, fia_18 IncN: ST7 | blaVIM-1, blaSHV-14 | aadA1, aph(3′)-XV, aacA4, fosA, sul1, dfrA14, oqxA, oqxB, qnrS1, catB2 |
| 2 | 5,477,368 | ST416 | IncFII(K), IncFIB(Mar), ColRNAI, IncN, IncFIB(K)/ IncF: fii_k5 IncN: ST7 | blaVIM-1, blaOKP-B-3 | aadA1, aph(3′)-XV, aacA4, aac(6′)-Ib-cr, fosA, sul1, qnrS1, catB2, oqxB, oqxA | |
| POVSG_VR | 1 | 5,346,528 | ST512 | ColRNAI, IncFIB(K)/fii_k2 | blaLEN-7, blaOXA-9, blaTEM-1A | aph(3′)-IIa, aadA2, aac(6′)-Ib-cr, fosA, sul1 |
| 1 | 5,101,877 | ST512 | ColRNAI, IncFIB(pQil), IncFII(K), IncFIB(K)/fii_k2 | blaKPC-9, blaOXA-9, blaTEM-1A | aph(3′)-IIa, aadA2, aac(6′)-Ib-cr, fosA, sul1 | |
| POCS_VR | 1 | 5,548,552 | ST512 | IncFIB(pQil), IncFII(K), IncFIB(K), ColRNAI, IncX3/ fii_k2 | blaOXA-9, blaSHV-11, blaTEM-1A, blaKPC-3 | aadA2, aac(6′)-Ib-cr, mph(A), sul1, dfrA12, oqxB, oqxA, catA1 |
| 2 | 5,589,508 | ST512 | IncFII(K), IncFIA(HI1), ColRNAI, IncFIB(K), Col(MG828), FIA(pBK30683)/fii_k12 fia_10 | blaOXA-23, blaLEN-12, blaCTX-M-15, blaOXA-1 | aac(3)-IIa, aac(6′)-Ib-cr, aadA1, fosA, sul1, dfrA1, oqxB, qnrB6, tet(D), tet(B), catB4, catA1 | |
| 2 | 5,604,773 | ST16 | IncFIB(pQil), IncFII(K), IncFIB(K), IncX4/fii_k2 | blaKPC-3, blaCTX-M-15, blaTEM-1B, blaSHV-1, blaOXA-1 | strA, aac(6′)-Ib-cr, strB, fosA, sul2, dfrA14, oqxB, oqxA, qnrB66, tet(A), catB4 | |
| 2 | 5,589,102 | ST35 | IncFIB(pQil), IncFII(K), IncFIB(K)/fii_k2 | blaTEM-1A, blaOXA-9, blaKPC-3, blaSHV-33 | fosA, oqxB, oqxA |
| β-lactamases | Classes | Isolates No. (%) | ST |
|---|---|---|---|
| TEM-1 | A | 44 (65) | 16, 35, 37, 273, 307, 512, 1519, 2623, 3227 |
| KPC-2 | A | 17 (25) | 37, 307 |
| KPC-3 | A | 30 (44) | 16, 35, 307, 512, 1519, 2623, 3227 |
| KPC-9 | A | 2 (3) | 512, 3227 |
| SHV-1 | A | 5 (7) | 16, 2623 |
| SHV-11 | A | 21 (31) | 11, 37, 273, 307, 512, 1519 |
| SHV-14 | A | 1 (1.5) | 416 |
| SHV-28 | A | 17 (25) | 307 |
| SHV-33 | A | 2 (3) | 35 |
| SHV-36 | A | 1 (1.5) | 253 |
| SHV-99 | A | 5 (7) | 321 |
| CTX-M-15 | A | 35 (51) | 11, 16, 37, 273, 307, 512, 2623, 3227 |
| LEN-7 | A | 2 (3) | 512, 3227 |
| LEN-12 | A | 9 (13) | 512, 3227 |
| OKP-B3 | A | 2 (3) | 416 |
| VIM-1 | B | 9 (13) | 253, 321, 416 |
| NDM-1 | B | 3 (4.5) | 11 |
| OXA-1 | D | 40 (59) | 16, 37, 273, 307, 512, 2623, 3227 |
| OXA-9 | D | 33 (48) | 35, 307, 512, 1519, 2623, 3227 |
| OXA-23 | D | 2 (3) | 512 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piccirilli, A.; Cherubini, S.; Azzini, A.M.; Tacconelli, E.; Lo Cascio, G.; Maccacaro, L.; Bazaj, A.; Naso, L.; Amicosante, G.; LTCF-Veneto Working Group; et al. Whole-Genome Sequencing (WGS) of Carbapenem-Resistant K. pneumoniae Isolated in Long-Term Care Facilities in the Northern Italian Region. Microorganisms 2021, 9, 1985. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091985
Piccirilli A, Cherubini S, Azzini AM, Tacconelli E, Lo Cascio G, Maccacaro L, Bazaj A, Naso L, Amicosante G, LTCF-Veneto Working Group, et al. Whole-Genome Sequencing (WGS) of Carbapenem-Resistant K. pneumoniae Isolated in Long-Term Care Facilities in the Northern Italian Region. Microorganisms. 2021; 9(9):1985. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091985
Chicago/Turabian StylePiccirilli, Alessandra, Sabrina Cherubini, Anna Maria Azzini, Evelina Tacconelli, Giuliana Lo Cascio, Laura Maccacaro, Alda Bazaj, Laura Naso, Gianfranco Amicosante, LTCF-Veneto Working Group, and et al. 2021. "Whole-Genome Sequencing (WGS) of Carbapenem-Resistant K. pneumoniae Isolated in Long-Term Care Facilities in the Northern Italian Region" Microorganisms 9, no. 9: 1985. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091985