
Ex-Vivo Adhesion of Enterococcus faecalis and Enterococcus faecium to the Intestinal Mucosa of Healthy Beagles

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacteria and Growth Conditions
2.2. Duodenal Tissue Samples
2.3. Dye Preparation
2.4. Collecting and Labeling Bacteria
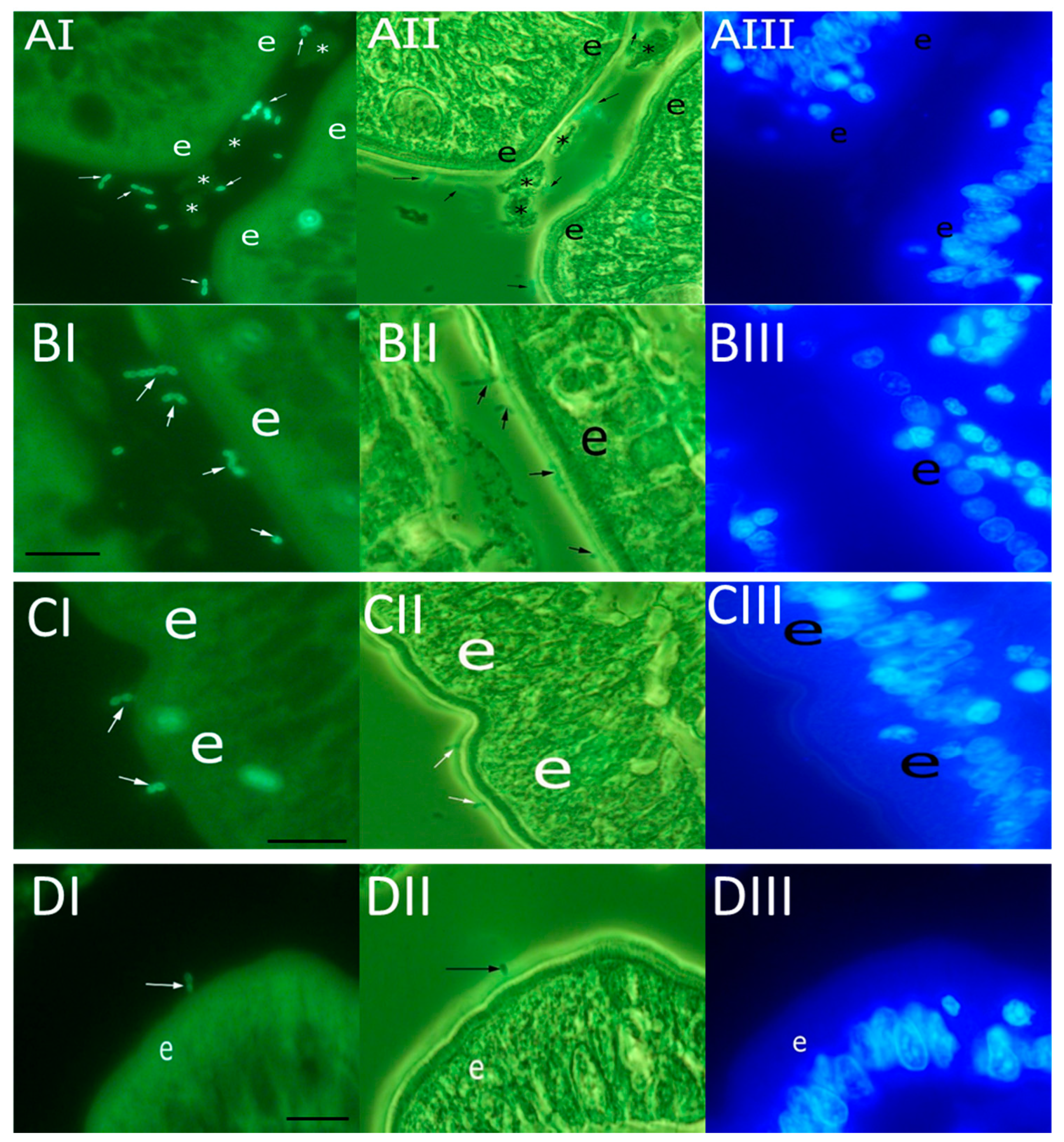
2.5. Bacterial Adhesion to Duodenal Mucosa
2.6. Measurement of Bacterial Hydrophobicity
2.7. Statistical Analyses
3. Results
3.1. Adherence of Enterococci to Canine Duodenal Mucosa
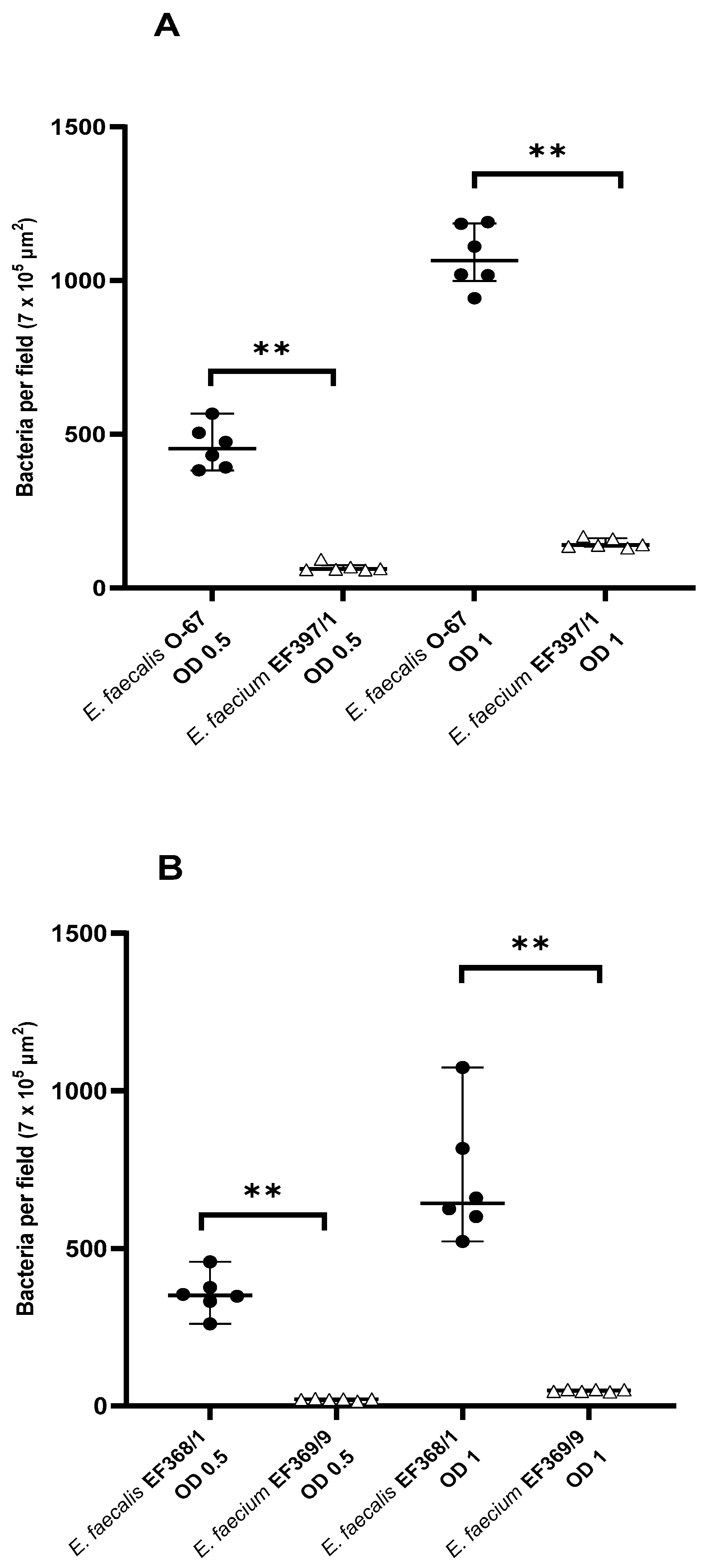
3.2. Intra-Host Species Comparison of Mucosal Adhesion of E. faecalis and E. faecium
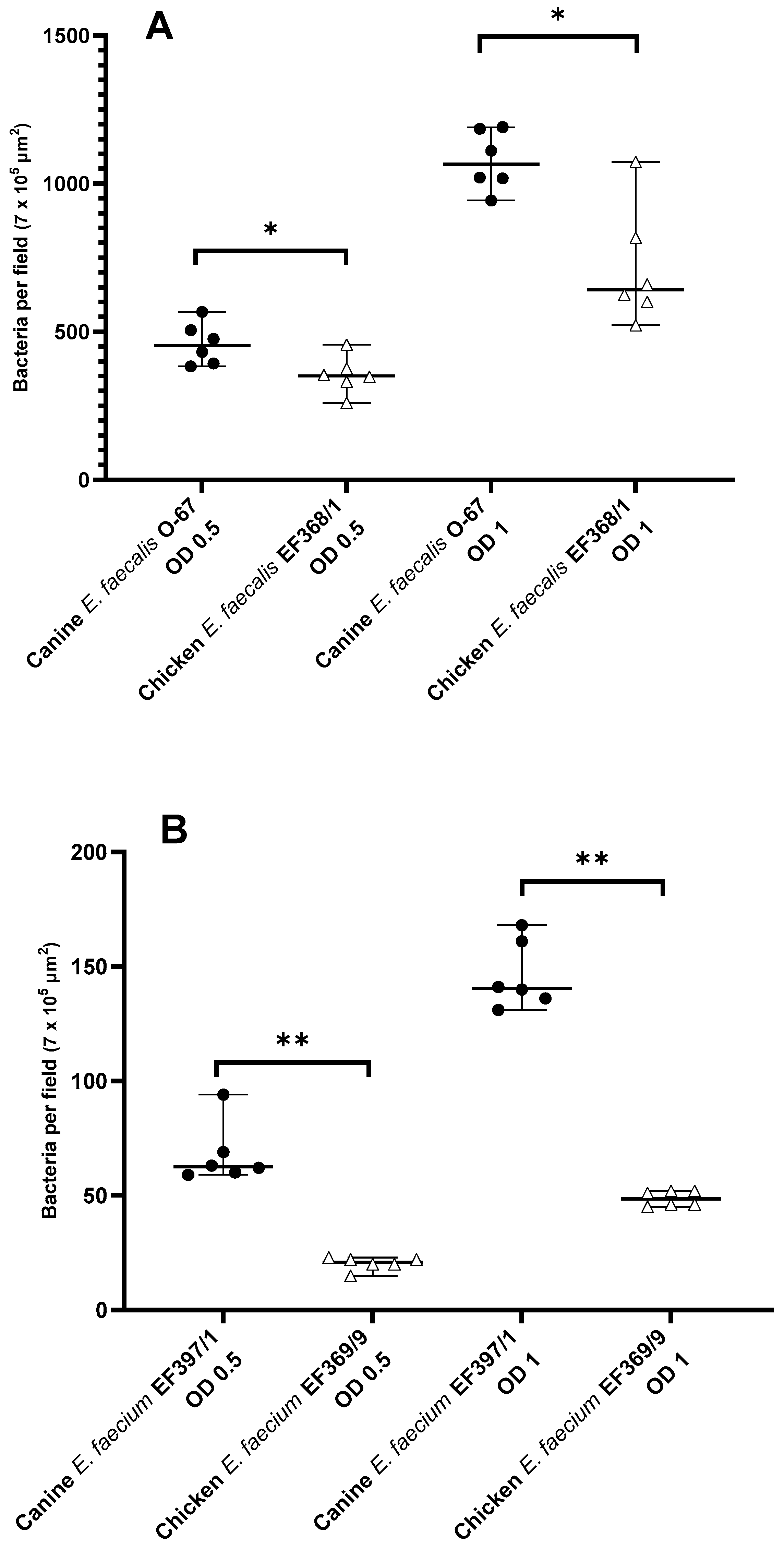
3.3. Inter-Host Species Comparison of Mucosal Adhesion of E. faecalis and E. faecium
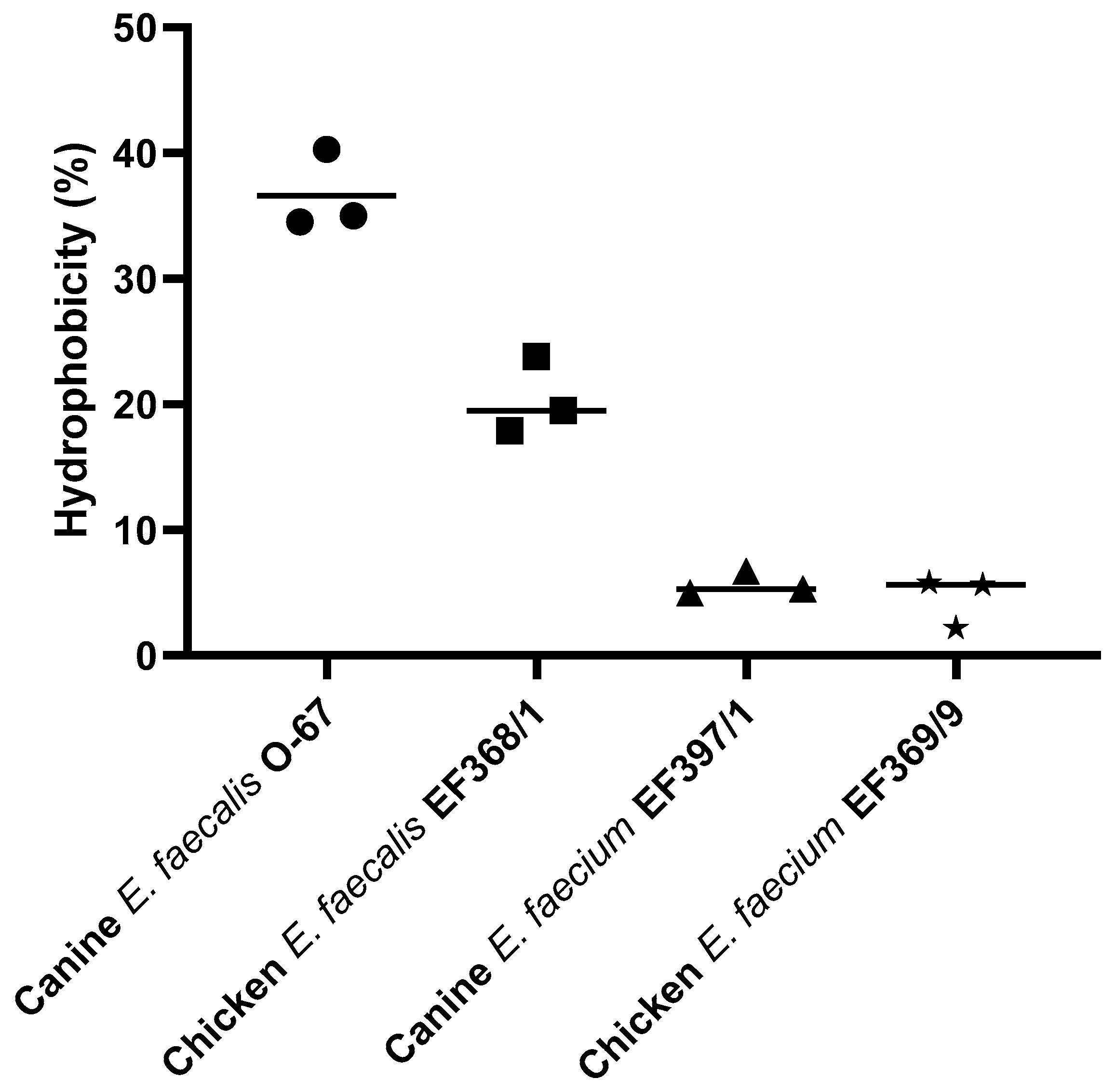
3.4. Hydrophobicity of E. faecalis and E. faecium
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gur, C.; Ibrahim, Y.; Isaacson, B.; Yamin, R.; Abed, J.; Gamliel, M.; Enk, J.; Bar-On, Y.; Stanietsky-Kaynan, N.; Coppenhagen-Glazer, S.; et al. Binding of the Fap2 protein of Fusobacterium nucleatum to human inhibitory receptor TIGIT protects tumors from immune cell attack. Immunity 2015, 42, 344–355. [Google Scholar] [CrossRef] [Green Version]
- Abed, J.; Emgard, J.E.; Zamir, G.; Faroja, M.; Almogy, G.; Grenov, A.; Sol, A.; Naor, R.; Pikarsky, E.; Atlan, K.A.; et al. Fap2 Mediates Fusobacterium nucleatum Colorectal Adenocarcinoma Enrichment by Binding to Tumor-Expressed Gal-GalNAc. Cell Host Microbe 2016, 20, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Isaacson, B.; Hadad, T.; Glasner, A.; Gur, C.; Granot, Z.; Bachrach, G.; Mandelboim, O. Stromal Cell-Derived Factor 1 Mediates Immune Cell Attraction upon Urinary Tract Infection. Cell Rep. 2017, 20, 40–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juntunen, M.; Kirjavainen, P.V.; Ouwehand, A.C.; Salminen, S.J.; Isolauri, E. Adherence of probiotic bacteria to human intestinal mucus in healthy infants and during rotavirus infection. Clin. Diagn. Lab. Immunol. 2001, 8, 293–296. [Google Scholar] [CrossRef] [Green Version]
- Kainulainen, V.; Tang, Y.; Spillmann, T.; Kilpinen, S.; Reunanen, J.; Saris, P.E.J.; Satokari, R. The canine isolate Lactobacillus acidophilus LAB20 adheres to intestinal epithelium and attenuates LPS-induced IL-8 secretion of enterocytes in vitro. BMC Microbiol. 2015, 15, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alp, D.; Kuleasan, H. Adhesion mechanisms of lactic acid bacteria: Conventional and novel approaches for testing. World J. Microbiol. Biotechnol. 2019, 35, 156. [Google Scholar] [CrossRef]
- McNaught, C.E.; MacFie, J. Probiotics in clinical practice: A critical review of the evidence. Nutr. Res. 2001, 21, 343–353. [Google Scholar] [CrossRef]
- Schmitz, S.; Suchodolski, J. Understanding the canine intestinal microbiota and its modification by pro-, pre- and synbiotics—what is the evidence? Vet. Med. Sci. 2016, 2, 71–94. [Google Scholar] [CrossRef]
- Dowarah, R.; Verma, A.K.; Agarwal, N.; Singh, P.; Singh, B.R. Selection and characterization of probiotic lactic acid bacteria and its impact on growth, nutrient digestibility, health and antioxidant status in weaned piglets. PLoS ONE 2018, 13, e0192978. [Google Scholar] [CrossRef]
- Kailasapathy, K.; Chin, J. Survival and therapeutic potential of probiotic organisms with reference to Lactobacillus acidophilus and Bifidobacterium spp. Immunol. Cell Biol. 2000, 78, 80–88. [Google Scholar] [CrossRef]
- Grzeskowiak, L.; Endo, A.; Beasley, S.; Salminen, S. Microbiota and probiotics in canine and feline welfare. Anaerobe 2015, 34, 14–23. [Google Scholar] [CrossRef]
- Pilla, R.; Suchodolski, J.S. The Role of the Canine Gut Microbiome and Metabolome in Health and Gastrointestinal Disease. Front. Vet. Sci. 2020, 6, 498. [Google Scholar] [CrossRef] [Green Version]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The Genus Enterococcus: Between Probiotic Potential and Safety Concerns—An Update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef]
- Baccouri, O.; Boukerb, A.M.; Farhat, L.B.; Zébré, A.; Zimmermann, K.; Domann, E.; Cambronel, M.; Barreau, M.; Maillot, O.; Rincé, I.; et al. Probiotic Potential and Safety Evaluation of Enterococcus faecalis OB14 and OB15, Isolated from Traditional Tunisian Testouri Cheese and Rigouta, Using Physiological and Genomic Analysis. Front. Microbiol. 2019, 10, 881. [Google Scholar] [CrossRef]
- Franz, C.M.; Huch, M.; Abriouel, H.; Holzapfel, W.; Galvez, A. Enterococci as probiotics and their implications in food safety. Int. J. Food Microbiol. 2011, 151, 125–140. [Google Scholar] [CrossRef] [Green Version]
- Nixon, S.L.; Rose, L.; Muller, A.T. Efficacy of an orally administered anti-diarrheal probiotic paste (Pro-Kolin Advanced) in dogs with acute diarrhea: A randomized, placebo-controlled, double-blinded clinical study. J. Vet. Intern. Med. 2019, 33, 1286–1294. [Google Scholar] [CrossRef] [Green Version]
- Bybee, S.N.; Scorza, A.V.; Lappin, M.R. Effect of the probiotic Enterococcus faecium SF68 on presence of diarrhea in cats and dogs housed in an animal shelter. J. Vet. Intern. Med. 2011, 25, 856–860. [Google Scholar] [CrossRef]
- Cerquetella, M.; Laus, F.; Speranzini, F.; Carnevali, C.; Spaterna, A.; Battaglia, E.; Bassotti, G. Efficacy of an enterovaccine in recurrent episodes of diarrhea in the dog: A pilot study. Rev. Esp. Sick Dig. 2012, 104, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Rinkinen, M.; Jalava, K.; Westermarck, E.; Salminen, S.; Ouwehand, A.C. Interaction between probiotic lactic acid bacteria and canine enteric pathogens: A risk factor for intestinal Enterococcus faecium colonization? Vet. Microbiol. 2003, 92, 111–119. [Google Scholar] [CrossRef]
- Rinkinen, M.; Mättö, J.; Salminen, S.; Westermarck, E.; Ouwehand, A.C. In vitro adhesion of lactic acid bacteria to canine small intestinal mucus. J. Anim. Physiol. Anim. Nutr. 2000, 84, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Rinkinen, M.; Westermarck, E.; Salminen, S.; Ouwehand, A.C. Absence of host specificity for in vitro adhesion of probiotic lactic acid bacteria to intestinal mucus. Vet. Microbiol. 2003, 97, 55–61. [Google Scholar] [CrossRef]
- Lauková, A.; Strompfová, V.; Ouwehand, A. Adhesion Properties of Enterococci to Intestinal Mucus of Different Hosts. Vet. Res. Commun. 2004, 28, 647–655. [Google Scholar] [CrossRef]
- Isaacson, B.; Hadad, T.; Bachrach, G.; Mandelboim, O. Quantification of Bacterial Attachment to Tissue Sections. Bio-Protoc. 2018, 8, e2741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Wouters, T.; Jans, C.; Niederberger, T.; Fischer, P.; Rühs, P.A. Adhesion Potential of Intestinal Microbes Predicted by Physico-Chemical Characterization Methods. PLoS ONE 2015, 10, e0136437. [Google Scholar] [CrossRef]
- Al Atya, A.K.; Drider-Hadiouche, K.; Ravallec, R.; Silvain, A.; Vachee, A.; Drider, D. Probiotic potential of Enterococcus faecalis strains isolated from meconium. Front. Microbiol. 2015, 6, 227. [Google Scholar] [CrossRef]
- Rosenberg, M.; Gutnick, D.; Rosenberg, E. Adherence of bacteria to hydrocarbons: A simple method for measuring cell-surface hydrophobicity. FEMS Microbiol. Lett. 1980, 9, 29–33. [Google Scholar] [CrossRef]
- Ocaña, V.S.; Nader-Macías, M.E. Vaginal lactobacilli: Self- and co-aggregating ability. Br. J. Biomed. Sci. 2002, 59, 183–190. [Google Scholar] [CrossRef]
- Pilla, R.; Guard, B.C.; Steiner, J.M.; Gaschen, F.P.; Olson, E.; Werling, D.; Allenspach, K.; Salavati Schmitz, S.; Suchodolski, J.S. Administration of a Synbiotic Containing Enterococcus faecium Does Not Significantly Alter Fecal Microbiota Richness or Diversity in Dogs With and Without Food-Responsive Chronic Enteropathy. Front. Vet. Sci. 2019, 6, 277. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, S.; Glanemann, B.; Garden, O.A.; Brooks, H.; Chang, Y.M.; Werling, D.; Allenspach, K. A Prospective, Randomized, Blinded, Placebo-Controlled Pilot Study on the Effect of Enterococcus faecium on Clinical Activity and Intestinal Gene Expression in Canine Food-Responsive Chronic Enteropathy. J. Vet. Intern. Med. 2015, 29, 533–543. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Niemi, P.; Salminen, S.J. The normal faecal microflora does not affect the adhesion of probiotic bacteria in vitro. FEMS Microbiol. Lett. 1999, 177, 35–38. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Salminen, S. In vitro adhesion assays for probiotics and their in vivo relevance: A review. Microb. Ecol. Health Dis. 2003, 15, 175–184. [Google Scholar] [CrossRef]
- Stępień-Pyśniak, D.; Hauschild, T.; Kosikowska, U.; Dec, M.; Urban-Chmiel, R. Biofilm formation capacity and presence of virulence factors among commensal Enterococcus spp. from wild birds. Sci. Rep. 2019, 9, 11204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favaro, L.; Basaglia, M.; Casella, S.; Hue, I.; Dousset, X.; de Melo Franco, B.D.G.; Todorov, S.D. Bacteriocinogenic potential and safety evaluation of non-starter Enterococcus faecium strains isolated from home made white brine cheese. Food Microbiol. 2014, 38, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, K.M.; Vieira, A.D.; Salles, H.O.; Oliveira Jda, S.; Rocha, C.R.; Borges Mde, F.; Bruno, L.M.; Franco, B.D.; Todorov, S.D. Safety, beneficial and technological properties of Enterococcus faecium isolated from Brazilian cheeses. Braz. J. Microbiol. 2015, 46, 237–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorov, S.D.; Botes, M.; Guigas, C.; Schillinger, U.; Wiid, I.; Wachsman, M.B.; Holzapfel, W.H.; Dicks, L.M.T. Boza, a natural source of probiotic lactic acid bacteria. J. Appl. Microbiol. 2008, 104, 465–477. [Google Scholar] [CrossRef]
- Roos, S.; Jonsson, H. A high-molecular-mass cell-surface protein from Lactobacillus reuteri 1063 adheres to mucus components. Microbiology 2002, 148, 433–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granato, D.; Perotti, F.; Masserey, I.; Rouvet, M.; Golliard, M.; Servin, A.; Brassart, D. Cell surface-associated lipoteichoic acid acts as an adhesion factor for attachment of Lactobacillus johnsonii La1 to human enterocyte-like Caco-2 cells. Appl. Environ. Microbiol. 1999, 65, 1071–1077. [Google Scholar] [CrossRef] [Green Version]
- Sava, I.G.; Zhang, F.; Toma, I.; Theilacker, C.; Li, B.; Baumert, T.F.; Holst, O.; Linhardt, R.J.; Huebner, J. Novel interactions of glycosaminoglycans and bacterial glycolipids mediate binding of enterococci to human cells. J. Biol. Chem. 2009, 284, 18194–18201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willems, R.J.L.; Top, J.; Braak van den, N.; van Belkum, A.; Endtz, H.; Mevius, D.; Stobberingh, E.; van den Bogaard, A.; van Embden, J.D.A. Host Specificity of Vancomycin-Resistant Enterococcus faecium. J. Infect. Dis. 2000, 182, 816–823. [Google Scholar] [CrossRef] [Green Version]
- Ahn, Y.T.; Lim, K.L.; Ryu, J.C.; Kang, D.K.; Ham, J.S.; Jang, Y.H.; Kim, H.U. Characterization of Lactobacillus acidophilus Isolated from Piglets and Chicken. Asian-Australas. J. Anim. Sci. 2002, 15, 1790–1797. [Google Scholar] [CrossRef]
- Van Tassell, M.L.; Miller, M.J. Lactobacillus adhesion to mucus. Nutrients 2011, 3, 613–636. [Google Scholar] [CrossRef] [Green Version]
- Aquilina, G.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; de Knecht, J.; Dierick, N.A.; Gralak, M.A.; Gropp, J.; Halle, I.; Hogstrand, C.; et al. Guidance on the safety assessment of Enterococcus faecium in animal nutrition. EFSA J. 2012, 10, 2682. [Google Scholar] [CrossRef]
- Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Dusemund, B.; Kouba, M.; Kos Durjava, M.; López-Alonso, M.; López Puente, S.; Marcon, F.; et al. Assessment of the application for renewal of authorisation of Bonvital® (Enterococcus faecium DSM 7134) as a feed additive for weaned piglets and pigs for fattening. EFSA J. 2019, 17, e05650. [Google Scholar] [CrossRef] [PubMed]
- Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Dusemund, B.; Kouba, M.; Fašmon Durjava, M.; López-Alonso, M.; López Puente, S.; Marcon, F.; et al. Assessment of the feed additive consisting of Enterococcus faecium DSM 7134 (Bonvital®) for chickens for fattening for the renewal of its authorisation (Lactosan GmbH & Co. KG). EFSA J. 2021, 19, e06451. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Weber, J.; Loening-Baucke, V.; Hale, L.P.; Lochs, H. Spatial organization and composition of the mucosal flora in patients with inflammatory bowel disease. J. Clin. Microbiol. 2005, 43, 3380–3389. [Google Scholar] [CrossRef] [Green Version]
- Giaretta, P.R.; Suchodolski, J.S.; Jergens, A.E.; Steiner, J.M.; Lidbury, J.A.; Cook, A.K.; Hanifeh, M.; Spillmann, T.; Kilpinen, S.; Syrjä, P.; et al. Bacterial Biogeography of the Colon in Dogs With Chronic Inflammatory Enteropathy. Vet. Pathol. 2020, 57, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Cassmann, E.; White, R.; Atherly, T.; Wang, C.; Sun, Y.; Khoda, S.; Mosher, C.; Ackermann, M.; Jergens, A. Alterations of the Ileal and Colonic Mucosal Microbiota in Canine Chronic Enteropathies. PLoS ONE 2016, 11, e0147321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janeczko, S.; Atwater, D.; Bogel, E.; Greiter-Wilke, A.; Gerold, A.; Baumgart, M.; Bender, H.; McDonough, P.L.; McDonough, S.P.; Goldstein, R.E.; et al. The relationship of mucosal bacteria to duodenal histopathology, cytokine mRNA, and clinical disease activity in cats with inflammatory bowel disease. Vet. Microbiol. 2008, 128, 178–193. [Google Scholar] [CrossRef] [PubMed]
- Flecknell, P. Replacement, reduction and refinement. Altex 2002, 19, 73–78. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Origin | |
|---|---|---|
| Enterococcus faecalis | O-67 | Dog feces; isolated at the Central Laboratory of the Department of Equine and Small Animal Medicine, University of Helsinki, Helsinki, Finland |
| Enterococcus faecalis | EF368/1 | Chicken cecum; isolated at the Microbiome Laboratory, Orion Corporation, Orion Pharma, R&D, Turku, Finland. |
| Enterococcus faecium | EF397/1 | Dog feces; isolated at the Microbiome Laboratory, Orion Corporation, Orion Pharma, R&D, Turku, Finland. |
| Enterococcus faecium | EF369/9 | Chicken cecum; isolated at the Microbiome Laboratory, Orion Corporation, Orion Pharma, R&D, Turku, Finland. |
| Adhesion Model | Description | Advantages | Disadvantages |
|---|---|---|---|
| Cell culture | Polar monolayer of enterocytes resembling intestinal tissue | Provides conditions more similar to in vivo environment | Derived from cancer cells, could differ from healthy tissue. Not representative of cell-type ratios in mucosal epithelial tissues |
| Caco-2/HT29 | Caco-2 and HT29 carcinoma cell lines | Simple, well-established in literature | Does not account for mucus presence |
| HT29-MTX/FU | HT29 culture treated with methotrexate or fluoruracil to secrete various types of mucus | Accounts for presence of mucus | May not represent appropriate MUC gene expression |
| Co-cultures | Mixed culture of secreting and mucus-secreting cells | Better represents cell-type ratio of mucosal epithelial tissues | Little literature on use in adhesion studies |
| Immobilized mucus | Mucus preparations immobilized, usually in microtitre wells | Fast, isolates mucus-microbe interactions from other in vivo conditions | Difficult to separate mucus, specifically from hydrophobic interactions |
| Whole tissue | Whole, intact, or excised tissue | Provides in vitro conditions most similar to in vivo environment | Costly, difficult to obtain |
| Resected tissue | Fragments of tissue excised from host | Mucus, epithelial tissue, and commensal organisms accounted for in model | Only small fragments at a time available from living hosts |
| Organ culture | Whole organs maintained in vitro | Better maintains the architecture of the tissue | Prohibitively expensive, may not function in same manner as in vivo |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanifeh, M.; Spillmann, T.; Huhtinen, M.; Sclivagnotis, Y.S.; Grönthal, T.; Hynönen, U. Ex-Vivo Adhesion of Enterococcus faecalis and Enterococcus faecium to the Intestinal Mucosa of Healthy Beagles. Animals 2021, 11, 3283. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11113283
Hanifeh M, Spillmann T, Huhtinen M, Sclivagnotis YS, Grönthal T, Hynönen U. Ex-Vivo Adhesion of Enterococcus faecalis and Enterococcus faecium to the Intestinal Mucosa of Healthy Beagles. Animals. 2021; 11(11):3283. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11113283
Chicago/Turabian StyleHanifeh, Mohsen, Thomas Spillmann, Mirja Huhtinen, Yannes S. Sclivagnotis, Thomas Grönthal, and Ulla Hynönen. 2021. "Ex-Vivo Adhesion of Enterococcus faecalis and Enterococcus faecium to the Intestinal Mucosa of Healthy Beagles" Animals 11, no. 11: 3283. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11113283