
Spatiotemporal Overlap between the European Brown Hare and Its Potential Predators and Competitors

 ,
,  , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
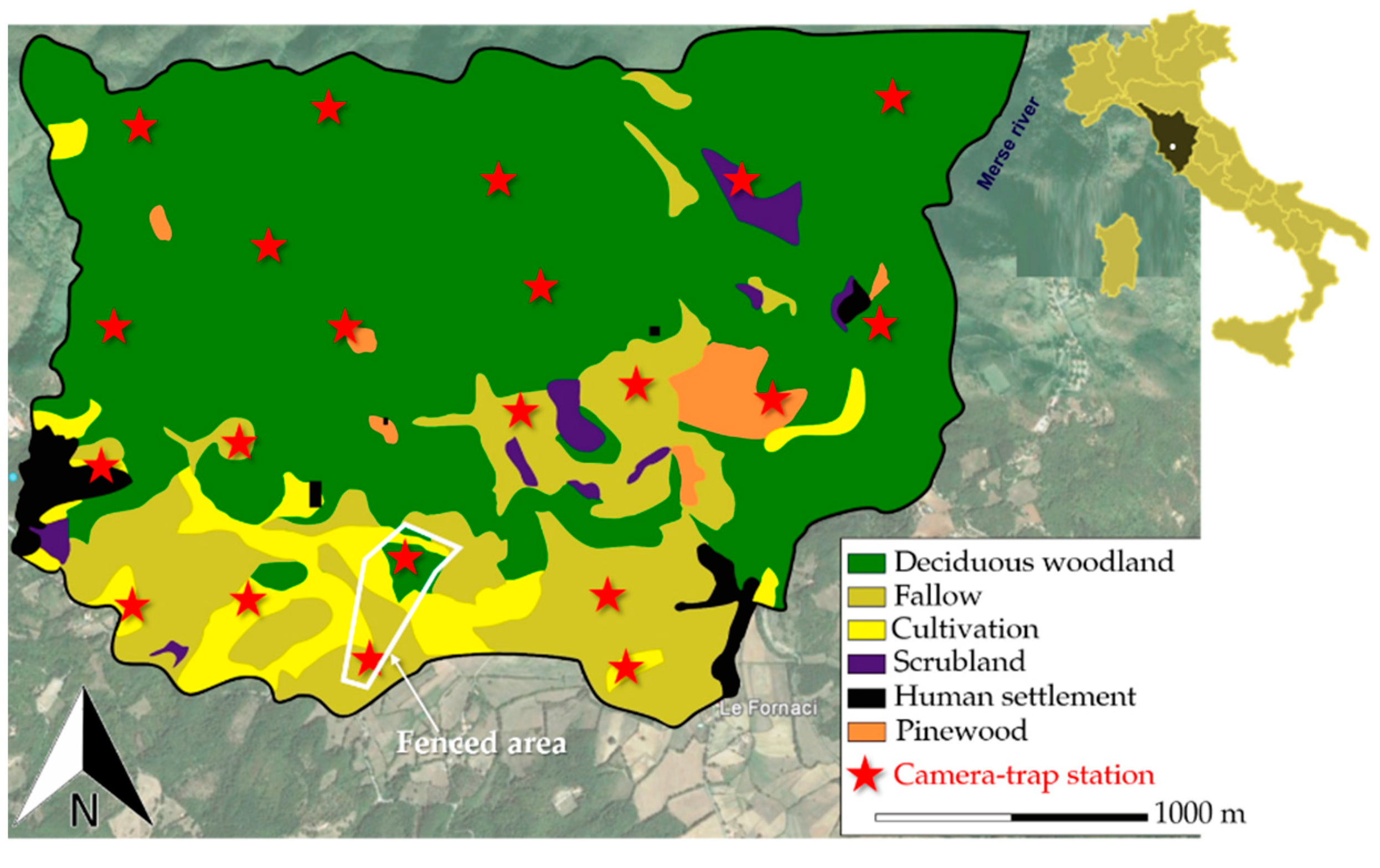
2.1. Study Area
2.2. Camera-Trap Sampling Design
2.3. Analysis of Spatiotemporal Overlap
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef] [Green Version]
- Finke, D.L.; Snyder, W.E. Niche partitioning increases resource exploitation by diverse communities. Science 2008, 321, 1488–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torretta, E.; Serafini, M.; Puopolo, F.; Schenone, L. Spatial and temporal adjustments allowing the coexistence among carnivores in Liguria (NW Italy). Acta Ethol. 2016, 19, 123–132. [Google Scholar] [CrossRef]
- Anile, S.; Devillard, S.; Ragni, B.; Rovero, F.; Mattucci, F.; Valvo, M.L. Habitat fragmentation and anthropogenic factors affect wildcat Felis silvestris silvestris occupancy and detectability on Mt Etna. Wildl. Biol. 2019, 1, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Houngbégnon, F.G.; Cornelis, D.; Vermeulen, C.; Sonké, B.; Ntie, S.; Fayolle, A.; Fonteyn, D.; Lhoest, S.; Ervard, Q.; Yapi, F.; et al. Daily activity patterns and co-occurrence of duikers revealed by an intensive camera trap survey across central african rainforests. Animals 2020, 10, 2200. [Google Scholar]
- Trolle, M.; Noss, A.J.; Cordeiro, J.L.P.; Oliveira, L.F.B. Brazilian tapir density in the Pantanal: A comparison of systematic camera-trapping and line-transect surveys. Biotropica 2008, 40, 211–217. [Google Scholar] [CrossRef]
- Chen, Y.; Xiao, Z.; Zhang, L.; Wang, X.; Li, M.; Xiang, Z. Activity rhythms of coexisting red serow and Chineseserow at Mt. Gaoligong as identified by camera traps. Animals 2019, 9, 1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, E.; Andreoni, A.; Cecere, F.; Magi, M.; Lazzeri, L. Patterns of activity rhythms of invasive coypus Myocastor coypus inferred through camera-trapping. Mammal. Biol. 2020, 100, 591–599. [Google Scholar] [CrossRef]
- Greco, I.; Fedele, E.; Salvatori, M.; Giampaoli Rustichelli, M.; Mercuri, F.; Santini, G.; Rovero, F.; Lazzaro, L.; Foggi, B.; Massolo, A.; et al. Guest or pest? Spatio-temporal occurrence and efforts on soil and vegetation of the wild boar on the Elba Island. Mammal. Biol. 2020. [Google Scholar] [CrossRef]
- Lashley, M.A.; Cove, M.V.; Chitwood, M.C.; Penido, G.; Gardner, B.; DePerno, C.S.; Moorman, C.E. Estimating wildlife activity curves: Comparison of methods and sample size. Sci. Rep. 2018, 8, 1–11. [Google Scholar]
- Cagnacci, F.; Cardini, A.; Ciucci, P.; Ferrari, N.; Mortelliti, A.; Preatoni, D.G.; Russo, D.; Scandura, M.; Wauters, L.A.; Amori, G. Less is more: Researcher survival guide in times of economic crisis. Hystrix 2012, 23, 1–7. [Google Scholar]
- Saisamorn, A.; Duengkae, P.; Pattanavibool, A.; Duangchantrasiri, S.; Simcharoen, A.; Smith, J.L. Spatial and temporal analysis of leopards (Panthera pardus), their prey and tigers (Panthera tigris) in Huai Kha Khaeng Wildlife Sanctuary, Thailand. Folia Oecol. 2019, 46, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Pinzon, K.; Reyna-Hurtado, R.; Meyer, N.F. Moon light and the activity patterns of Baird’s tapir in the Calakmul region, Southern México. Therya 2020, 11, 137–142. [Google Scholar] [CrossRef]
- Tsunoda, H.; Newman, C.; Peeva, S.; Raichev, E.; Buesching, C.D.; Kaneko, Y. Spatio-temporal partitioning facilitates mesocarnivore sympatry in the Stara Planina Mountains, Bulgaria. J. Zool. 2020, 141, 125801. [Google Scholar] [CrossRef]
- Zhao, G.; Yang, H.; Xie, B.; Gong, Y.; Ge, J.; Feng, L. Spatio-temporal coexistence of sympatric mesocarnivores with a single apex carnivore in a fine-scale landscape. Glob. Ecol. Cons. 2020, 21, e00897. [Google Scholar] [CrossRef]
- Mori, E.; Bagnato, S.; Serroni, P.; Sangiuliano, A.; Rotondaro, F.; Marchianò, V.; Cascini, V.; Poerio, L.; Ferretti, F. Spatiotemporal mechanisms of coexistence in an European mammal community in a protected area of southern Italy. J. Zool. 2020, 310, 232–245. [Google Scholar] [CrossRef]
- Havmøller, R.W.; Jacobsen, N.S.; Scharff, N.; Rovero, F.; Zimmermann, F. Assessing the activity pattern overlap among leopards (Panthera pardus), potential prey and competitors in a complex landscape in Tanzania. J. Zool. 2020, 311, 175–182. [Google Scholar] [CrossRef]
- Tang, X.; Tang, S.; Li, X.; Menghe, D.; Bao, W.; Xiang, C.; Gao, F.; Bao, W. A study of population size and activity patterns and their relationship to the prey species of the Eurasian lynx using a camera trapping approach. Animals 2019, 9, 864. [Google Scholar] [CrossRef] [Green Version]
- Mori, E.; Menchetti, M. Living with roommates in a shared den: Spatial and temporal segregation among semifossorial mammals. Behav. Processes 2019, 164, 48–53. [Google Scholar] [CrossRef]
- Yang, W.; Chen, Y.; Deng, Y.; Wang, X.; Chen, L.; Hu, D.; Luo, X.; Song, D.; Xiao, Z. Preliminary surveys of mammals and birds by infrared camera traps in the Sichuan Baishuihe National Nature Reserve. Biodiv. Sci. 2019, 27, 1012. [Google Scholar]
- Croose, E.; Bled, F.; Fowler, N.L.; Beyer, D.E., Jr.; Belant, J.L. American marten and fisher do not segregate in space and time during winter in a mixed-forest system. Ecol. Evol. 2019, 9, 4906–4916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanni, M.; Brivio, F.; Grignolio, S.; Apollonio, M. Estimation of spatial and temporal overlap in three ungulate species in a Mediterranean environment. Mammal Res. 2020. [Google Scholar] [CrossRef]
- Monterroso, P.; Alves, P.C.; Ferreras, P. Plasticity in circadian activity patterns of mesocarnivores in Southwestern Europe: Implications for species coexistence. Behav. Ecol. Sociobiol. 2014, 68, 1403–1417. [Google Scholar] [CrossRef] [Green Version]
- Anile, S.; Devillard, S. Camera-trapping provides insights into adult sex ratio variability in felids. Mammal Rev. 2018, 48, 168–179. [Google Scholar] [CrossRef]
- Ossi, F.; Ranc, N.; Moorcroft, P.; Bonanni, P.; Cagnacci, F. Ecological and behavioral drivers of supplemental feeding use by roe deer Capreolus capreolus in a peri-urban context. Animals 2020, 10, 2088. [Google Scholar] [CrossRef]
- Lioy, S.; Mori, E.; Wauters, L.A.; Bertolino, S. Weight operated see-saw feeding hoppers are not selective for red squirrels when greys are present. Mammal. Biol. 2016, 81, 365–371. [Google Scholar] [CrossRef]
- Mori, E.; Paniccia, C.; Munkhstog, B.; Cicero, M.; Augugliaro, C. Temporal overlap among small-mammals in a grassland and a forest-alpine meadow of Central Asia. Mammal. Biol. 2021. [Google Scholar] [CrossRef]
- Oberosler, V.; Groff, C.; Iemma, A.; Pedrini, P.; Rovero, F. The influence of human disturbance on occupancy and activity patterns of mammals in the Italian Alps from systematic camera trapping. Mammal. Biol. 2017, 87, 50–61. [Google Scholar] [CrossRef]
- Riga, F.; Trocchi, V.; Randi, E.; Toso, S. Morphometric differentiation between the Italian hare (Lepus corsicanus De Winton, 1898) and the European brown hare (Lepus europaeus Pallas, 1778). J. Zool. 2001, 253, 241–252. [Google Scholar] [CrossRef]
- Fattorini, N.; Burrini, L.; Morao, G.; Ferretti, F.; Romeo, G.; Mori, E. Splitting hairs: How to tell hair of hares apart for predator diet studies. Mammal. Biol. 2018, 89, 84–89. [Google Scholar] [CrossRef]
- Viganò, M.; Ancillotto, L.; Agnelli, P.; Ficetola, G.F.; Mori, E. Frequency of Occurrence and Ingested Biomass of Different Prey of the Barn Owl Tyto alba in an Island Ecosystem. Birds 2020, 1, 2. [Google Scholar] [CrossRef]
- Böhm, C.; Landmann, A. Carnivory in the Alpine marmot (Marmota marmota): An underestimated phenomenon? Ethol. Ecol. Evol. 2020. [Google Scholar] [CrossRef]
- Santilli, F.; Bagliacca, M.; Paci, G. Density and habitat use of sympatric Brown hares and European rabbits in a Mediterranean farmland area of Tuscany (Central Italy). Ethol. Ecol. Evol. 2015, 27, 233–243. [Google Scholar] [CrossRef]
- Barbar, F.; Lambertucci, S.A. The roles of leporid species that have been translocated: A review of their ecosystem effects as native and exotic species. Mammal Rev. 2018, 48, 245–260. [Google Scholar] [CrossRef]
- Gortazar, C.; Millán, J.; Acevedo, P.; Escudero, M.A.; Marco, J.; De Luco, D.F. A large-scale survey of brown hare Lepus europaeus and Iberian hare L. granatensis populations at the limit of their ranges. Wildl. Biol. 2007, 13, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Edwards, P.J.; Fletcher, M.R.; Berny, P. Review of the factors affecting the decline of the European brown hare, Lepus europaeus (Pallas, 1778) and the use of wildlife incident data to evaluate the significance of paraquat. Agric. Ecosyst. Environ. 2000, 79, 95–103. [Google Scholar] [CrossRef]
- Bertolino, S.; Perrone, A.; Gola, L.; Viterbi, R. Population density and habitat use of the introduced Eastern cottontail (Sylvilagus floridanus) compared to the native European hare (Lepus europaeus). Zool. Stud. 2011, 50, 315–326. [Google Scholar]
- Santilli, F.; Galardi, L. Factors affecting brown hare (Lepus europaeus) hunting bags in Tuscany region (central Italy). Hystrix 2007, 17, 143–153. [Google Scholar]
- Santilli, F.; Galardi, L. Effect of habitat structure and type of farming on European hare (Lepus europaeus) abundance. Hystrix 2016, 27, 120–122. [Google Scholar]
- Sliwinski, K.; Ronnenberg, K.; Jung, K.; Strauß, E.; Siebert, U. Habitat requirements of the European brown hare (Lepus europaeus Pallas 1778) in an intensively used agriculture region (Lower Saxony, Germany). BMC Ecol. 2019, 19, 31. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.K.; Jennings, N.V.; Robinson, A.; Harris, S. Conservation of European hares Lepus europaeus in Britain: Is increasing habitat heterogeneity in farmland the answer? J. Appl. Ecol. 2004, 41, 1092–1102. [Google Scholar] [CrossRef]
- Vaughan, N.; Lucas, E.A.; Harris, S.; White, P.C. Habitat associations of European hares Lepus europaeus in England and Wales: Implications for farmland management. J. Appl. Ecol. 2003, 40, 163–175. [Google Scholar] [CrossRef]
- Weber, D.; Roth, T.; Kohli, L. Increasing brown hare (Lepus europaeus) densities in farmland without predator culling: Results of a field experiment in Switzerland. Eur. J. Wildl. Res. 2019, 65, 75. [Google Scholar] [CrossRef]
- Soyumert, A.; Ertürk, A.; Tavşanoğlu, Ç. The importance of lagomorphs for the Eurasian lynx in Western Asia: Results from a large scale camera-trapping survey in Turkey. Mammal. Biol. 2019, 95, 18–25. [Google Scholar] [CrossRef]
- Weterings, M.J.; Zaccaroni, M.; van der Koore, N.; Zijlstra, L.M.; Kuipers, H.J.; van Langevelde, F.; van Wieren, S.E. Strong reactive movement response of the medium-sized European hare to elevated predation risk in short vegetation. Anim. Behav. 2016, 115, 107–114. [Google Scholar] [CrossRef]
- Ullmann, W.; Fischer, C.; Kramer-Schadt, S.; Pirhofer-Walzl, K.; Glemnitz, M.; Blaum, N. How do agricultural practices affect the movement behaviour of European brown hares (Lepus europaeus)? Agric. Ecosyst. Environ. 2020, 292, 106819. [Google Scholar] [CrossRef]
- Mayer, M.; Fog Bjerre, D.H.; Sunde, P. Better safe than sorry: The response to a simulated predator and unfamiliar scent by the European hare. Ethology 2020, 126, 704–715. [Google Scholar] [CrossRef]
- Schai-Braun, S.C.; Rödel, H.G.; Hackländer, K. The influence of daylight regime on diurnal locomotor activity patterns of the European hare (Lepus europaeus) during summer. Mammal. Biol. 2012, 77, 434–440. [Google Scholar] [CrossRef]
- Pepin, D.; Cargnelutti, B. Individual variations of daily activity patterns in radiotracked European hares during winter. Acta Theriol. 1994, 39, 399. [Google Scholar] [CrossRef] [Green Version]
- Zaccaroni, M.; Biliotti, N.; Buccianti, A.; Calieri, S.; Ferretti, M.; Genghini, M.; Riga, F.; Trocchi, V.; Dessì-Fulgheri, F. Winter locomotor activity patterns of European hares (Lepus europaeus). Mammal. Biol. 2013, 78, 482–485. [Google Scholar] [CrossRef]
- Homolka, M. Daily activity pattern of the European hare (Lepus europaeus). Folia Zool. 1986, 35, 33–42. [Google Scholar]
- Panek, M. Factors affecting predation of red foxes Vulpes vulpes on brown hares Lepus europaeus during the breeding season in Poland. Wildl. Biol. 2009, 15, 345–349. [Google Scholar] [CrossRef]
- Balestrieri, A.; Remonti, L.; Ruiz-González, A.; Vergara, M.; Capelli, E.; Gómez-Moliner, B.J.; Prigioni, C. Food habits of genetically identified pine marten (Martes martes) expanding in agricultural lowlands (NW Italy). Acta Theriol. 2011, 56, 199–207. [Google Scholar] [CrossRef]
- Hummel, S.; Meyer, L.; Hackländer, K.; Weber, D. Activity of potential predators of European hare (Lepus europaeus) leverets and ground-nesting birds in wildflower strips. Eur. J. Wildl. Res. 2017, 63, 102. [Google Scholar] [CrossRef]
- Liberg, O. Food habits and prey impact by feral and house-based domestic cats in a rural area in southern Sweden. J. Mammal. 1984, 65, 424–432. [Google Scholar] [CrossRef]
- Sangiuliano, A.; Lovari, S.; Ferretti, F. Dietary partitioning between European roe deer and European brown hare. Eur. J. Wildl. Res. 2016, 62, 527–535. [Google Scholar] [CrossRef]
- Mori, E.; Ferretti, F.; Fattorini, N. Alien war: Ectoparasite load, diet and temporal niche partitioning in a multi-species assembly of small rodents. Biol. Invasions 2019, 21, 3305–3318. [Google Scholar] [CrossRef]
- Mori, E.; Ferretti, F.; Lagrotteria, A.; La Greca, L.; Solano, E.; Fattorini, N. Impact of wild boar on small forest-dwelling rodents. Ecol. Res. 2020, 35, 675–681. [Google Scholar] [CrossRef]
- Mori, E.; Menchetti, M.; Dondini, G.; Biosa, D.; Vergari, S. Theriofauna of Site of Community Importance Poggi di Prata (Grosseto, Central Italy): Terrestrial mammals and preliminary data on Chiroptera. Check List 2014, 10, 718–723. [Google Scholar] [CrossRef] [Green Version]
- Sogliani, D.; Mori, E. “The Fox and the Cat”: Sometimes they do not agree. Mammal. Biol. 2019, 95, 150–154. [Google Scholar] [CrossRef]
- Franchini, M.; Fazzi, P.; Lucchesi, M.; Mori, E. Diet of adult and juvenile wildcats in Southern Tuscany (Central Italy). Folia Zool. 2017, 66, 147–151. [Google Scholar] [CrossRef]
- Meredith, M.; Ridout, M. Overview of the Overlap Package. 2014. Available online: http://cran.cs.wwu.edu/web/packages/overlap/vignettes/overlap.pdf (accessed on 12 December 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; ISBN 3-900051-07-0. Available online: http://www.Rproject.org/ (accessed on 22 January 2021).
- Landler, L.; Ruxton, G.D.; Malkemper, E.P. The Hermans–Rasson test as a powerful alternative to the Rayleigh test for circular statistics in biology. BMC Ecol. 2019, 19, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Viviano, A.; Amori, G.; Luiselli, L.; Oebel, H.; Bahleman, F.; Mori, E. Blessing the rains down in Africa: Spatiotemporal behaviour of the crested porcupine Hystrix cristata (Mammalia: Rodentia) in the rainy and dry seasons, in the African savannah. Trop. Zool. 2020, 33, 113–124. [Google Scholar] [CrossRef]
- Pianka, E.R. Niche overlap and diffuse competition. Proc. Natl. Acad. Sci. USA 1974, 71, 2141–2145. [Google Scholar] [CrossRef] [Green Version]
- Graham, I.M.; Redpath, S.M.; Thirgood, S.J. The diet and breeding density of Common Buzzards Buteo buteo in relation to indices of prey abundance. Bird Study 1995, 42, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Butterworth, A.; Turner, K.M.; Jennings, N. Minimising orphaning in the brown hare Lepus europaeus in England and Wales: Should a close season be introduced? Wildl. Biol. 2017, 1, 4. [Google Scholar]
- Canova, L.; Gazzola, A.; Pollini, L.; Balestrieri, A. Surveillance and habitat diversity affect European brown hare (Lepus europaeus) density in protected breeding areas. Eur. J. Wildl. Res. 2020, 66, 1–10. [Google Scholar] [CrossRef]
- Mori, E.; Menchetti, M.; Camporesi, A.; Cavigioli, L.; Tabarelli de Fatis, K.; Girardello, M. License to Kill? Domestic Cats Affect a Wide Range of Native Fauna in a Highly Biodiverse Mediterranean Country. Front. Ecol. Evol. 2019, 7, 477. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, F.; Pacini, G.; Belardi, I.; ten Cate, B.; Sensi, M.; Oliveira, R.; Rossa, M.; Burrini, L.; Lovari, S. Recolonizing wolves and opportunistic foxes: Interference or facilitation? Biol. J. Linn Soc. 2021, 132, 196–210. [Google Scholar] [CrossRef]
- Cavallini, P.; Volpi, T. Variation in the diet of the red fox in a Mediterranean area. Revue Ecol. 1996, 51, 173–189. [Google Scholar]
- Goszczyński, J.; Wasilewski, M. Predation of foxes on a hare population in central Poland. Acta Theriol. 1992, 37, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Cozzi, G.; Broekhuis, F.; McNutt, J.W.; Turnbull, L.A.; Macdonald, D.W.; Schmid, B. Fear of the dark or dinner by moonlight? Reduced temporal partitioning among Africa’s large carnivores. Ecology 2012, 93, 2590–2599. [Google Scholar] [CrossRef]
- Lima, S.L.; Dill, L.M. Behavioural decision made under the risk of predation: A review and prospectus. Can. J. Zool. 1990, 68, 619–640. [Google Scholar] [CrossRef]
- Monterroso, P.; Alves, P.C.; Ferreras, P. Catch me if you can: Diel activity patterns of Mammalian prey and predators. Ethology 2016, 119, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Fattorini, N.; Pokheral, C.P. Activity and habitat selection of the Indian crested porcupine. Ethol. Ecol. Evol. 2012, 24, 377–387. [Google Scholar] [CrossRef]
- Mori, E.; Lovari, S.; Cozzi, F.; Gabbrielli, C.; Giari, C.; Torniai, L.; Romeo, G.; Ferretti, F.; Fattorini, N. Safety or satiety? Spatiotemporal behaviour of a threatened herbivore. Mammal. Biol. 2020, 100, 49–61. [Google Scholar] [CrossRef]
- Penteriani, V.; Kuparinen, A.; del Mar Delgado, M.; Palomares, F.; López-Bao, J.V.; Fedriani, J.M.; Calzada, J.; Moreno, S.; Villafuerte, R.; Campioni, L.; et al. Responses of a top and a meso predator and their prey to moon phases. Oecologia 2013, 173, 753–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilman, D. The importance of the mechanisms of interspecific competition. Am. Nat. 1987, 129, 769–774. [Google Scholar] [CrossRef]
- Telfer, E.S. Forage yield and browse utilization on logged areas in New Brunswick. Can. J. For. Res. 1972, 2, 346–350. [Google Scholar] [CrossRef]
- Belovsky, G.E. Moose and snowshoe hare competition and a mechanistic explanation from foraging theory. Oecologia 1984, 61, 150–159. [Google Scholar] [CrossRef]
- Hulbert, I.A.; Andersen, R. Food competition between a large ruminant and a small hindgut fermentor: The case of the roe deer and mountain hare. Oecologia 2001, 128, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Weterings, M.J.; Moonen, S.; Prins, H.H.; van Wieren, S.E.; van Langevelde, F. Food quality and quantity are more important in explaining foraging of an intermediate-sized mammalian herbivore than predation risk or competition. Ecol. Evol. 2018, 8, 8419–8432. [Google Scholar] [CrossRef] [PubMed]
- Weterings, M.J.; Ewert, S.P.; Peereboom, J.N.; Kuipers, H.J.; Kuijper, D.P.; Prins, H.H.; Jansen, P.A.; van Langevelde, F.; van Wieren, S.E. Implications of shared predation for space use in two sympatric leporids. Ecol. Evol. 2019, 9, 3457–3469. [Google Scholar] [CrossRef] [Green Version]
- Loy, A.; Aloise, G.; Ancillotto, L.; Angelici, F.M.; Bertolino, S.; Capizzi, D.; Castiglia, R.; Colangelo, P.; Contoli, L.; Cozzi, B.; et al. Mammals of Italy: An annotated checklist. Hystrix 2019, 30, 87–106. [Google Scholar]
- Amori, G.; Contoli, L.; Nappi, A. Mammalia II: Erinaceomorpha, Soricomorpha, Lagomorpha, Rodentia (Series “La Fauna d’Italia”); Calderini Editions: Bologna, Italy, 2008. [Google Scholar]
- Mayer, M.; Ullmann, W.; Sunde, P.; Fischer, C.; Blaum, N. Habitat selection by the European hare in arable landscapes: The importance of small-scale habitat structure for conservation. Ecol. Evol. 2018, 8, 11619–11633. [Google Scholar] [CrossRef]
- Schai-Braun, S.C.; Ruf, T.; Klansek, E.; Arnold, W.; Hackländer, K. Positive effects of set-asides on European hare (Lepus europaeus) populations: Leverets benefit from an enhanced survival rate. Biol. Cons. 2020, 244, 108518. [Google Scholar] [CrossRef]
- Oldfield, T.E.E.; Smith, R.J.; Harrop, S.R.; Leader-Williams, N. Field sports and conservation in the United Kingdom. Nature 2003, 423, 531–533. [Google Scholar] [CrossRef]
- Reid, N.; Magee, C.; Montgomery, W.I. Integrating field sports, hare population management and conservation. Acta Theriol. 2010, 55, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Naldi, L.; Greco, I.; Ferretti, F.; Zaccaroni, M. Density estimates and habitat preferences of the European hare (Lepus europaeus) on mountainous areas in Italy. Mammal Study 2020, 45, 123–131. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Comparison | Overlap Coefficient | 95% CI | p |
|---|---|---|---|
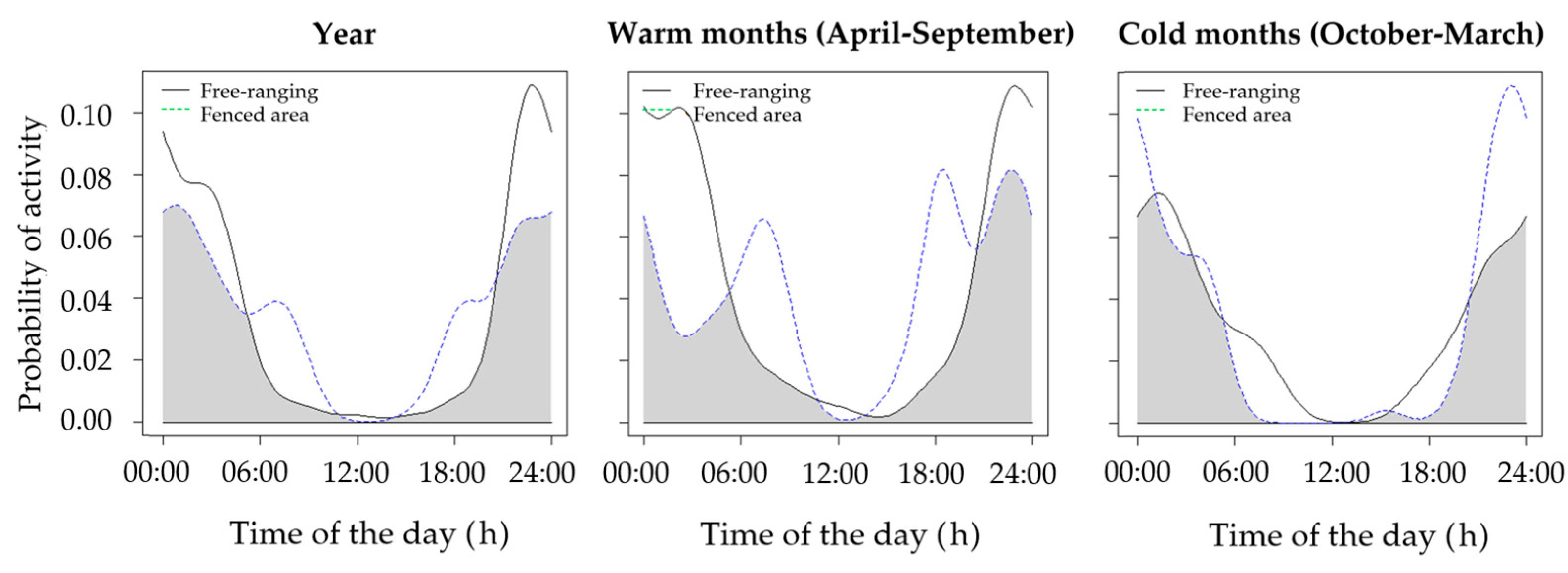
| Fenced vs. non-fenced brown hare (warm months) | 0.64 | 0.53–0.80 | 0.02 |
| Fenced vs. non-fenced brown hare (cold months) | 0.78 | 0.68–0.88 | 0.02 |
| Fenced vs. non-fenced brown hare (whole year) | 0.75 | 0.70–0.87 | <0.01 |
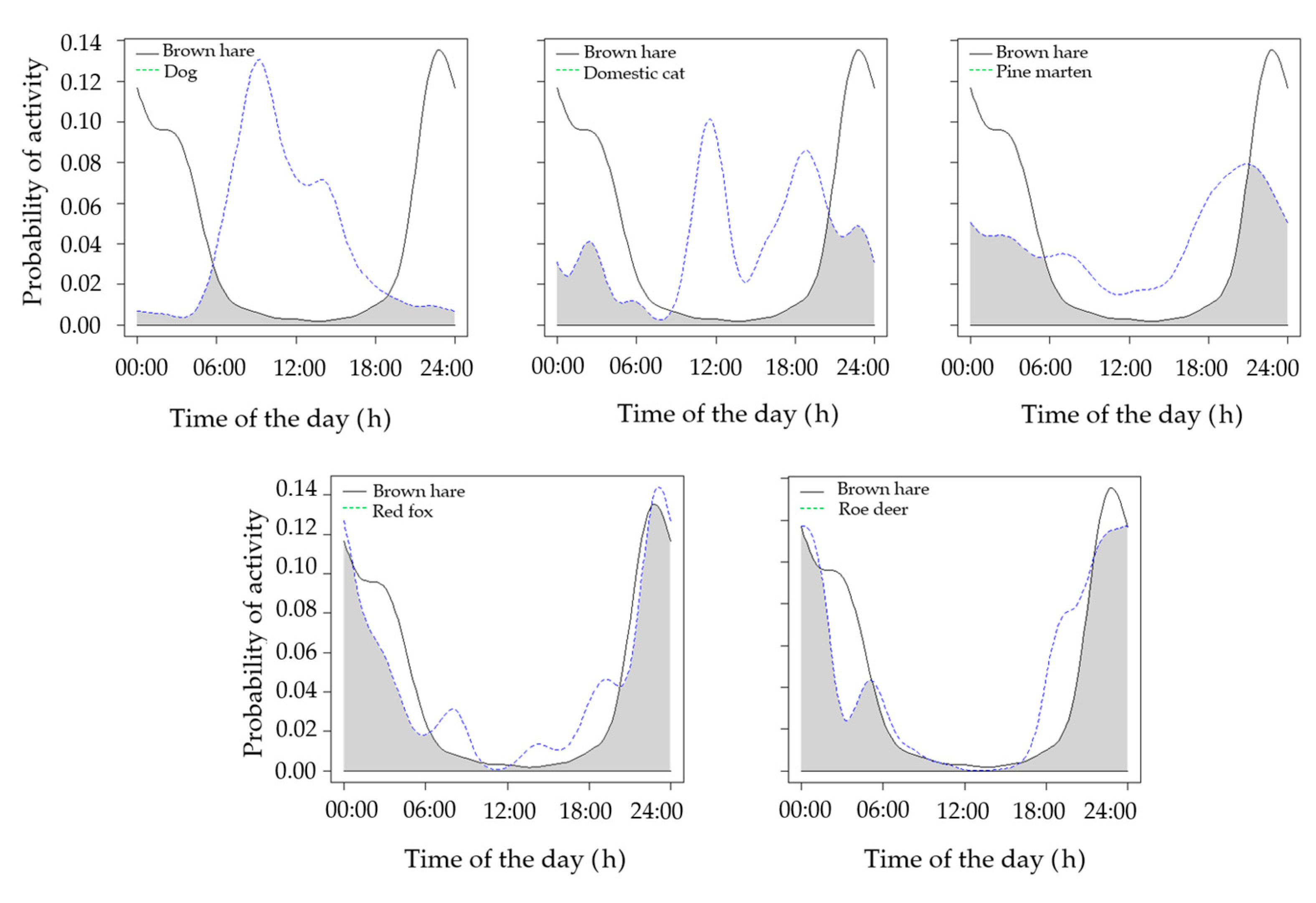
| Non-fenced brown hare vs. red fox | 0.82 | 0.74–0.90 | <0.01 |
| Non-fenced brown hare vs. domestic cat | 0.39 | 0.29–0.59 | <0.01 |
| Non-fenced brown hare vs. domestic dog | 0.15 | 0.13–0.27 | <0.01 |
| Non-fenced brown hare vs. pine marten | 0.59 | 0.51–0.75 | 0.02 |
| Non-fenced brown hare vs. roe deer | 0.81 | 0.77–0.88 | <0.01 |
| Comparison | Pianka Index |
|---|---|
| Brown hare vs roe deer | 0.15 |
| Brown hare vs domestic cat | 0.36 |
| Brown hare vs domestic dog | 0.77 |
| Brown hare vs pine marten | 0.18 |
| Brown hare vs red fox | 0.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viviano, A.; Mori, E.; Fattorini, N.; Mazza, G.; Lazzeri, L.; Panichi, A.; Strianese, L.; Mohamed, W.F. Spatiotemporal Overlap between the European Brown Hare and Its Potential Predators and Competitors. Animals 2021, 11, 562. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11020562
Viviano A, Mori E, Fattorini N, Mazza G, Lazzeri L, Panichi A, Strianese L, Mohamed WF. Spatiotemporal Overlap between the European Brown Hare and Its Potential Predators and Competitors. Animals. 2021; 11(2):562. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11020562
Chicago/Turabian StyleViviano, Andrea, Emiliano Mori, Niccolò Fattorini, Giuseppe Mazza, Lorenzo Lazzeri, Alessandra Panichi, Luigi Strianese, and Walid Fathy Mohamed. 2021. "Spatiotemporal Overlap between the European Brown Hare and Its Potential Predators and Competitors" Animals 11, no. 2: 562. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11020562