
Endocrine Signals Altered by Heat Stress Impact Dairy Cow Mammary Cellular Processes at Different Stages of the Dry Period

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and methods
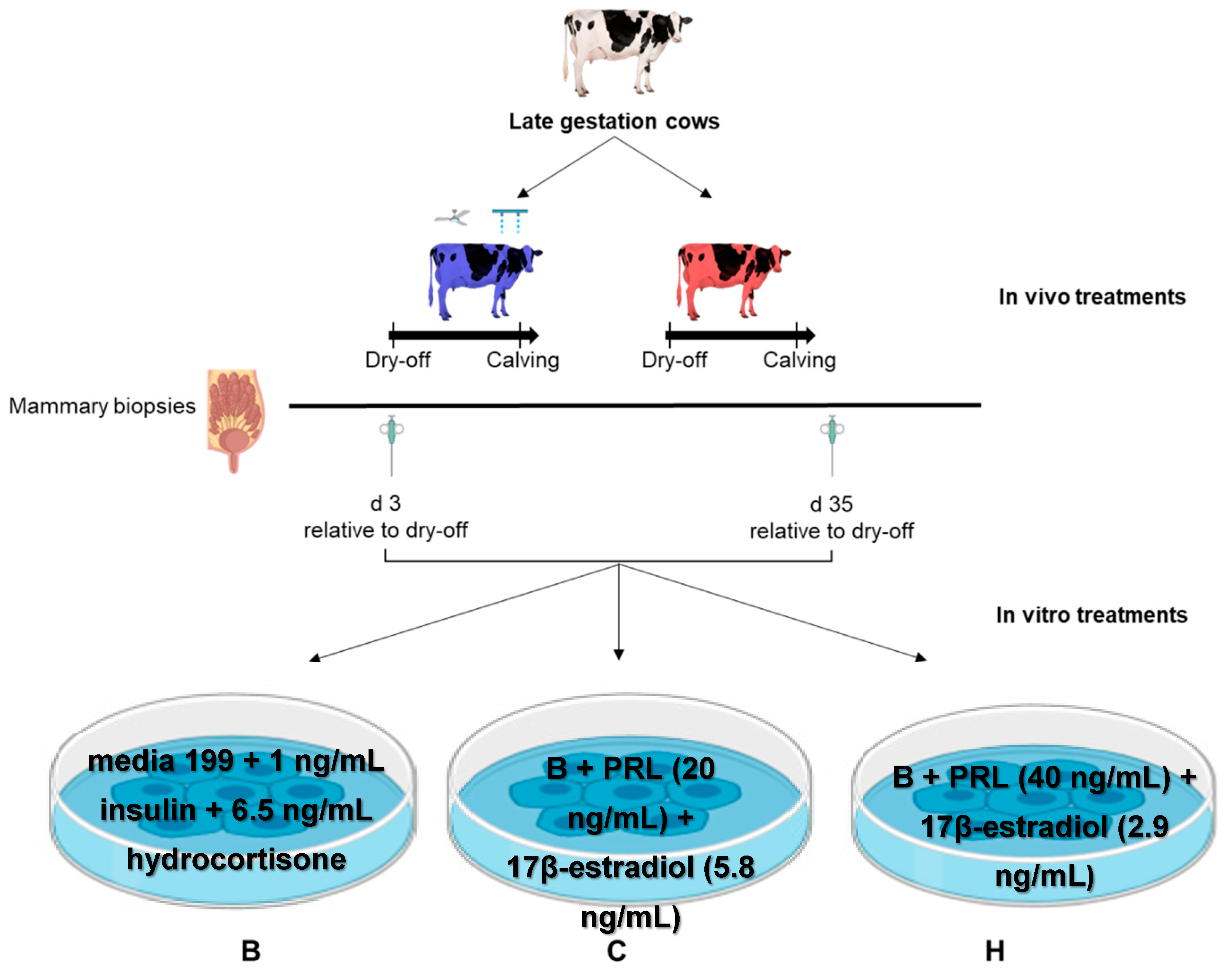
2.1. Animals and Experimental Design
2.2. Indicators of Heat Stress
2.3. Mammary Biopsies and Tissue Explant Processing
2.4. Lactate Dehydrogenase Activity Analysis
2.5. Explant RNA Extraction
2.6. Gene Expression Analysis
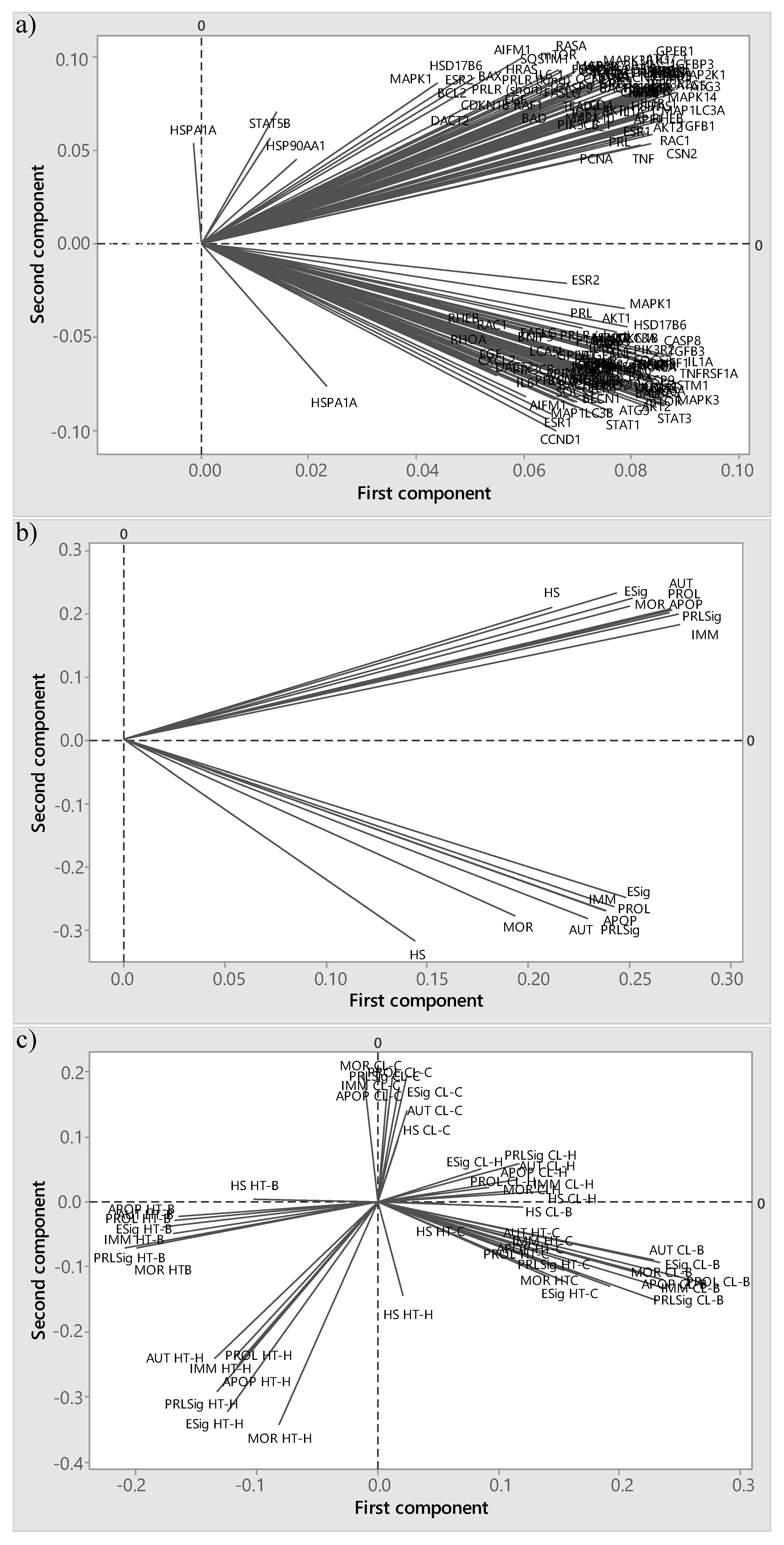
2.7. Principal Component Analysis
2.8. Statistical Analyses
3. Results
3.1. Thermal Environment and Physiological Parameters In Vivo
3.2. Lactate Dehydrogenase Activity In Vitro
3.3. Principal Component Analysis
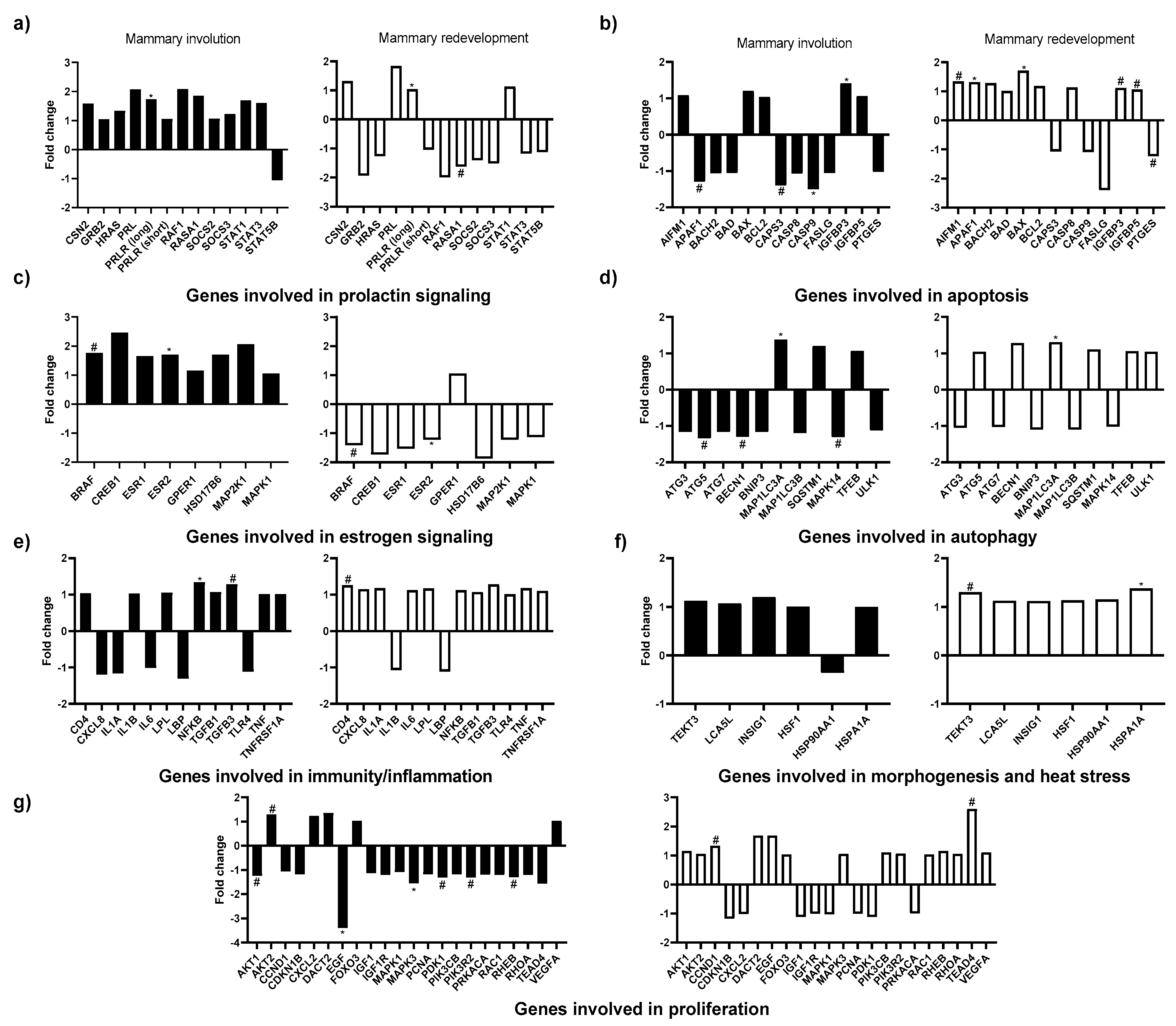
3.4. Effects of In Vivo Thermal Environment on Mammary Gland Gene Expression
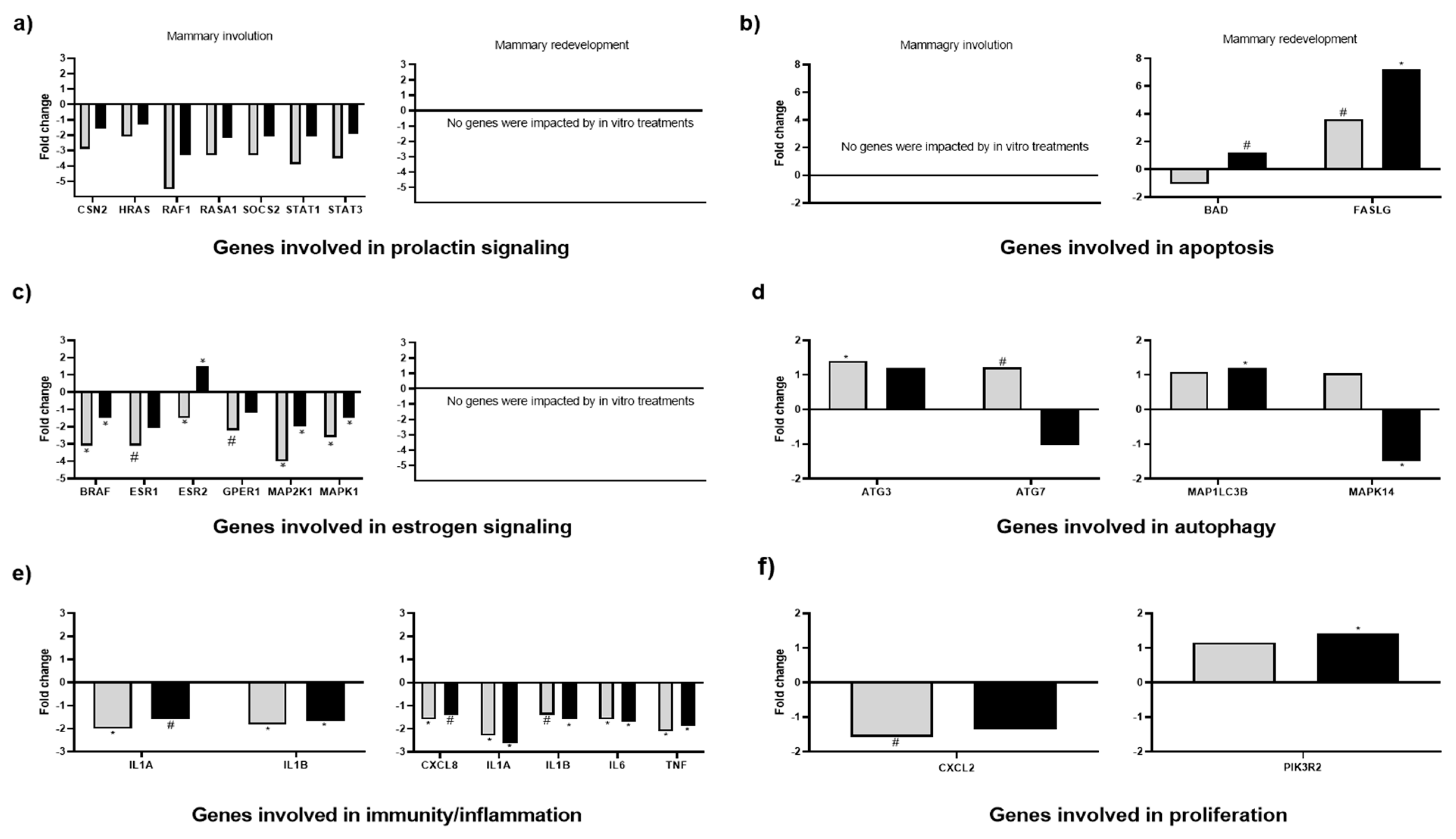
3.5. Effects of In Vitro Treatments on Mammary Gland Gene Expression
3.6. Interactions between In Vivo and In Vitro Treatments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Capuco, A.V.; Akers, R.M.; Smith, J.J. Mammary growth in Holstein cows during the dry period: Quantification of nucleic acids and histology. J. Dairy Sci. 1997, 80, 477–487. [Google Scholar] [CrossRef]
- Capuco, A.V.; Ellis, S.E.; Hale, S.A.; Long, E.; Erdman, R.A.; Zhao, X.; Paape, M.J. Lactation persistency: Insights from mammary cell proliferation studies. J. Anim. Sci. 2003, 81 (Suppl. 3), 18–31. [Google Scholar] [CrossRef] [Green Version]
- Hurley, W.L. Mammary Gland Function during Involution. J. Dairy Sci. 1989, 72, 1637–1646. [Google Scholar] [CrossRef]
- Wilde, C.J.; Quarrie, L.H.; Tonner, E.; Flint, D.J.; Peaker, M. Mammary apoptosis. Livest. Prod. Sci. 1997, 50, 29–37. [Google Scholar] [CrossRef]
- Sorensen, M.T.; Orgaard, J.V.N.; Theil, P.K.; Vestergaard, M.; Sejrsen, K. Cell turnover and activity in mammary tissue during lactation and dry period in dairy cows. J. Dairy Sci. 2006, 89, 4632–4639. [Google Scholar] [CrossRef]
- Zarzyńska, J.; Gajkowska, B.; Wojewodzka, U.; Dymnicki, E.; Motyl, T. Apoptosis and autophagy in involuting bovine mammary gland is accompanied by up-regulation of TGF-beta1 and suppression of somatotropic pathway. Pol. J. Vet. Sci. 2007, 10, 1–9. [Google Scholar] [PubMed]
- Tao, S.; Bubolz, J.W.; do Amaral, B.C.; Thompson, I.M.; Hayen, M.J.; Johnson, S.E.; Dahl, G.E. Effect of heat stress during the dry period on mammary gland development. J. Dairy Sci. 2011, 94, 5976–5986. [Google Scholar] [CrossRef]
- Tao, S.; Thompson, I.M.; Monteiro, A.P.A.; Hayen, M.J.; Young, L.J.; Dahl, G.E. Effect of cooling heat-stressed dairy cows during the dry period on insulin response. J. Dairy Sci. 2012, 95, 5035–5046. [Google Scholar] [CrossRef] [PubMed]
- Ouellet, V.; Laporta, J.; Dahl, G.E. Late gestation heat stress in dairy cows: Effects on dam and daughter. Theriogenology 2020, 150, 471–479. [Google Scholar] [CrossRef]
- Beardsley, G.L.; Muller, L.D.; Garverick, H.C.; Ludens, F.C.; Tucker, W.L. Initiation of parturition in dairy cows with dexame-thasone. II. Response to dexamethasone in combination with estradiol benzoate. J. Dairy Sci. 1976, 59, 241–247. [Google Scholar] [CrossRef]
- Bremmer, D.R.; Christensen, J.O.; Grummer, R.R.; Rasmussen, F.E.; Wiltbank, M.C. Effects of induced parturition and estradiol on feed intake, liver triglyceride concentration, and plasma metabolites of transition dairy cows. J. Dairy Sci. 1999, 82, 1440–1448. [Google Scholar] [CrossRef]
- Do Amaral, B.C.; Connor, E.E.; Tao, S.; Hayen, M.J.; Bubolz, J.W.; Dahl, G.E. Heat-stress abatement during the dry period: Does cooling improve transition into lactation? J. Dairy Sci. 2009, 92, 5988–5999. [Google Scholar] [CrossRef]
- Fabris, T.F.; Laporta, J.; Skibiel, A.L.; Corra, F.N.; Senn, B.; Wohlgemuth, S.E.; Dahl, G.E. Effect of heat stress during early, late, and entire dry period on dairy cattle. J. Dairy Sci. 2019, 02, 5647–5656. [Google Scholar] [CrossRef]
- Lough, D.S.; Beede, D.L.; Wilcox, C.J. Effects of feed intake and thermal stress on mammary blood flow and other physiological measurements in lactating dairy cows. J. Dairy Sci. 1990, 3, 325–332. [Google Scholar] [CrossRef]
- Collier, R.J.; Doelger, S.G.; Head, H.H.; Thatcher, W.W.; Wilcox, C.J. Effects of Heat Stress during Pregnancy on Maternal Hormone Concentrations, Calf Birth Weight and Postpartum Milk Yield of Holstein Cows. J. Anim. Sci. 1982, 54, 309–319. [Google Scholar] [CrossRef]
- Do Amaral, B.C.; Connor, E.E.; Tao, S.; Hayen, M.J.; Bubolz, J.W.; Dahl, G.E. Heat stress abatement during the dry period influences prolactin signaling in lymphocytes. Domest. Anim. Endocrinol. 2010, 38, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Do Amaral, B.C.; Connor, E.E.; Tao, S.; Hayen, M.J.; Bubolz, J.W.; Dahl, G.E. Heat stress abatement during the dry period influences metabolic gene expression and improves immune status in the transition period of dairy cows. J. Dairy Sci. 2011, 94, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Accorsi, P.; Pacioni, B.; Pezzi, C.; Forni, M.; Flint, D.; Seren, E. Role of Prolactin, Growth Hormone and Insulin-Like Growth Factor 1 in Mammary Gland Involution in the Dairy Cow. J. Dairy Sci. 2002, 85, 507–513. [Google Scholar] [CrossRef]
- Connor, E.; Meyer, M.; Li, R.; Van Amburgh, M.; Boisclair, Y.; Capuco, A. Regulation of Gene Expression in the Bovine Mammary Gland by Ovarian Steroids. J. Dairy Sci. 2007, 90 (Suppl. 1), E55–E65. [Google Scholar] [CrossRef] [Green Version]
- Sobolewska, A.; Gajewska, M.; Zarzyńska, J.; Gajkowska, B.; Motyl, T. IGF-I, EGF, and sex steroids regulate autophagy in bovine mammary epithelial cells via the mTOR pathway. Eur. J. Cell Biol. 2009, 88, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Wohlgemuth, S.E.; Ramirez-Lee, Y.; Tao, S.; Monteiro, A.P.A.; Ahmed, B.M.; Dahl, G.E. Short communication: Effect of heat stress on mammary gland autophagy during the dry period. J. Dairy Sci. 2016, 99, 4875–4880. [Google Scholar] [CrossRef] [PubMed]
- Fabris, T.F.; Skibiel, A.L.; Laporta, J.; McLean, D.J.; Kirk, D.J.; Chapman, J.D.; Dahl, G.E. Heat stress and OmniGen-AF alter mammary gland gene expression and endocrine responses in the dry period. J. Dairy Sci. 2018, 101 (Suppl. 2), 69. [Google Scholar]
- Fabris, T.F.; Laporta, J.; Skibiel, A.L.; Senn, B.; Wohlgemuth, S.E.; Dahl, G.E. Effect of heat stress during the early and late dry period on mammary gland development of Holstein dairy cattle. J. Dairy Sci. 2020, 103, 8576–8586. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Dairy Cattle, 7th ed.; National Academies Press: Washington, DC, USA, 2001. [Google Scholar]
- NRC. A Guide to Environmental Research on Animals; National Academy of Sciences: Washington, DC, USA, 1971. [Google Scholar]
- Dikmen, S.; Hansen, P. Is the temperature-humidity index the best indicator of heat stress in lactating dairy cows in a subtropical environment? J. Dairy Sci. 2009, 92, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dado-Senn, B.; Vega Acosta, L.; Torre Riviera, M.; Field, S.L.; Marrero, M.G.; Davison, B.D.; Tao, S.; Fabris, T.F.; Ortiz-Colón, G.; Dahl, G.E.; et al. Pre- and postnatal heat stress abatement affects dairy calf thermoregulation and performance. J. Dairy Sci. 2020, 103, 4822–4837. [Google Scholar] [CrossRef]
- Skibiel, A.L.; Dado-Senn, B.; Fabris, T.F.; Dahl, G.E.; Laporta, J. In utero exposure to thermal stress has long-term effects on mammary gland microstructure and function in dairy cattle. PLoS ONE 2018, 13, e0206046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vries, L.; Casey, T.; Dover, H.; VandeHaar, M.; Plaut, K. Effects of transforming growth factor-β on mammary remodeling during the dry period of dairy cows. J. Dairy Sci. 2011, 94, 6036–6046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, S.; Monteiro, A.; Thompson, I.; Hayen, M.; Dahl, G. Effect of late-gestation maternal heat stress on growth and immune function of dairy calves. J. Dairy Sci. 2012, 95, 7128–7136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrero, M.G.; Field, S.L.; Skibiel, A.L.; Dado-Senn, B.; Driver, J.P.; Laporta, J. Increasing serotonin bioavailability alters gene expression in peripheral leukocytes and lymphoid tissues of dairy calves. Sci. Rep. 2020, 10, 9712. [Google Scholar] [CrossRef] [PubMed]
- Macciotta, N.; Gaspa, G.; Steri, R.; Nicolazzi, E.; DiMauro, C.; Pieramati, C.; Cappio-Borlino, A. Using eigenvalues as variance priors in the prediction of genomic breeding values by principal component analysis. J. Dairy Sci. 2010, 93, 2765–2774. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2011, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dado-Senn, B.; Skibiel, A.L.; Fabris, T.F.; Zhang, Y.; Dahl, G.E.; Peñagaricano, F.; LaPorta, J. RNA-Seq reveals novel genes and pathways involved in bovine mammary involution during the dry period and under environmental heat stress. Sci. Rep. 2018, 8, 11096. [Google Scholar] [CrossRef]
- Chapman, K.W.; Lawless, H.T.; Boor, K.J. Quantitative Descriptive Analysis and Principal Component Analysisfor Sensory Characterization of Ultrapasteurized Milk. J. Dairy Sci. 2001, 84, 12–20. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Rhoads, R. Effects of Heat Stress on Postabsorptive Metabolism and Energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bole-Feysot, C.; Goffin, V.; Edery, M.; Binart, N.; Kelly, P.A. Prolactin (PRL) and its receptor: Actions, signal transduction path-ways and phenotypes observed in PRL receptor knockout mice. Endocr. Rev. 1998, 19, 225–268. [Google Scholar] [CrossRef] [PubMed]
- Corazzin, M.; Saccà, E.; Lippe, G.; Romanzin, A.; Foletto, V.; Da Borso, F.; Piasentier, E. Effect of Heat Stress on Dairy Cow Performance and on Expression of Protein Metabolism Genes in Mammary Cells. Animals 2020, 10, 2124. [Google Scholar] [CrossRef] [PubMed]
- Sobolewska, A.; Elminowska-Wenda, G.; Bogucka, J.; Szpinda, M.; Walasik, K.; Bednarcyk, M.; Paruszewska-Actel, M. Myogenesis—Possibilities of its stimulation in chickens. Folia Biol. 2011, 59, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Lanneau, D.; De Thonel, A.; Maurel, S.; Didelot, C.; Garrido, C. Apoptosis Versus Cell Differentiation: Role of heat shock protein HSP90, HSP70 and HSP27. Prion 2007, 1, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Collier, R.J.; Dahl, G.E.; Van Baale, M.J. Major advances associated with environmental effects on dairy cattle. J. Dairy Sci. 2006, 89, 1244–1253. [Google Scholar] [CrossRef]
- Kanninen, T.T.; Sisti, G.; Witkin, S.S. Induction of the 70 kDa heat shock protein stress response inhibits autophagy: Possible consequences for pregnancy outcome. J. Matern. Fetal Neonatal Med. 2016, 29, 159–162. [Google Scholar] [CrossRef]
- Thompson, I.M.; Dahl, G.E. Dry period seasonal effects on the subsequent lactation. Prof. Anim. Sci. 2012, 28, 628–631. [Google Scholar] [CrossRef]
- Montilla, S.I.R.; Johnson, T.P.; Pearce, S.C.; Gardan-Salmon, D.; Gabler, N.K.; Ross, J.W.; Rhoads, R.P.; Baumgard, L.H.; Lonergan, S.M.; Selsby, J.T. Heat stress causes oxidative stress but not inflammatory signaling in porcine skeletal muscle. Temperature 2014, 1, 42–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.; Vetharaniam, I.; Dobson, J.; Prewitz, M.; Oden, K.; Murney, R.; Swanson, K.; McDonald, R.; Henderson, H.; Stelwagen, K. Cell survival signaling in the bovine mammary gland during the transition from lactation to involution. J. Dairy Sci. 2016, 99, 7523–7543. [Google Scholar] [CrossRef] [Green Version]
- Tao, S.; Dahl, G. Invited review: Heat stress effects during late gestation on dry cows and their calves. J. Dairy Sci. 2013, 96, 4079–4093. [Google Scholar] [CrossRef] [PubMed]

 ), tissue explants cultured in media (
), tissue explants cultured in media ( ), and tissue explants incubated in Triton X-100 (
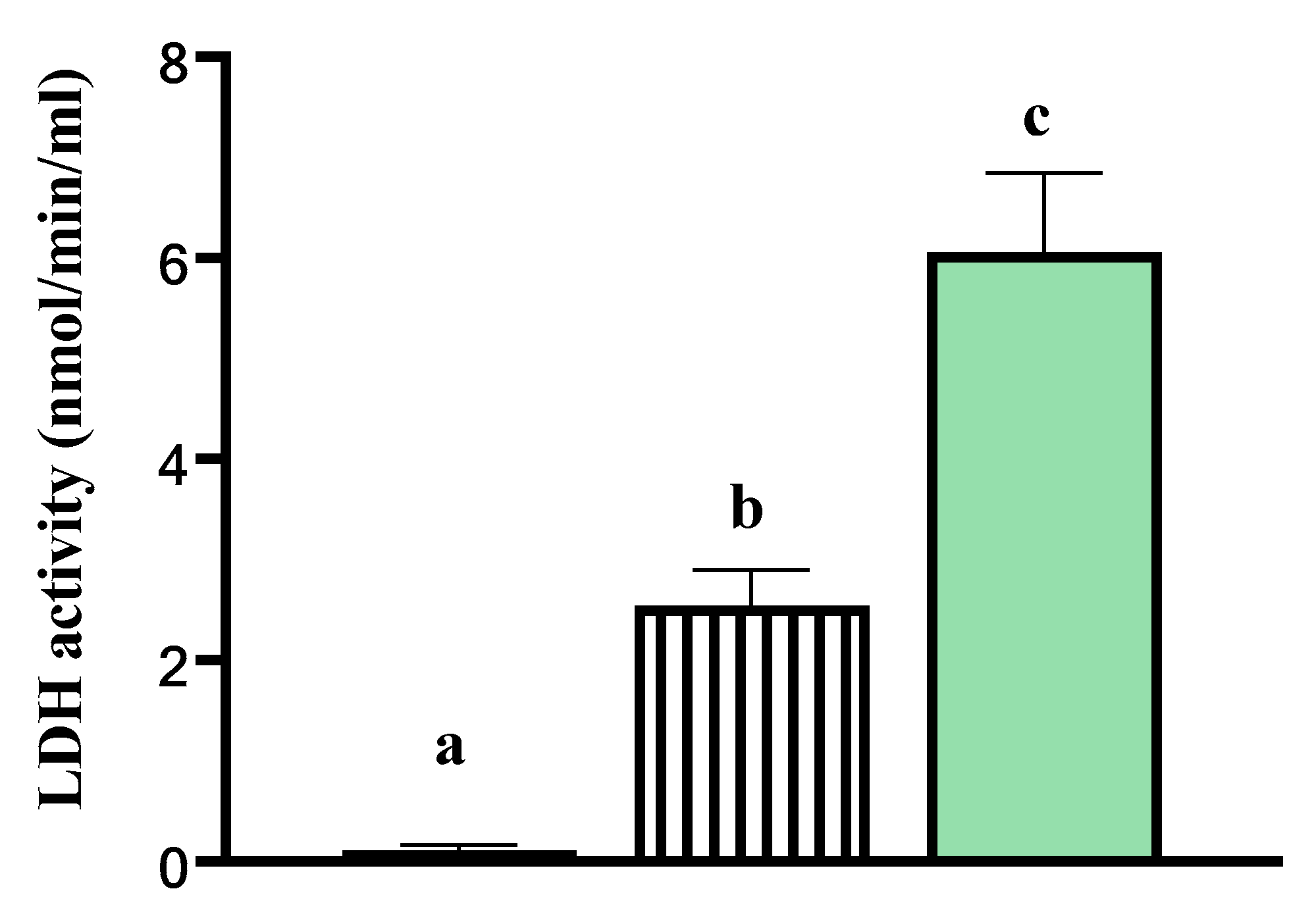
), and tissue explants incubated in Triton X-100 ( ). Data are presented as least square means ± standard error of the mean (SEM) Different letters a–c indicate p-value < 0.05. LDH activity was higher in the media of explants incubated in Triton X-100 at room temperature relative to both the media from tissue explants cultured in vitro and relative to the basal media deprived of tissue explants. This indicated that cells within the explants cultured in vitro for 24 h were viable.
), tissue explants cultured in media (), and tissue explants incubated in Triton X-100 (). Data are presented as least square means ± standard error of the mean (SEM) Different letters a–c indicate p-value < 0.05. LDH activity was higher in the media of explants incubated in Triton X-100 at room temperature relative to both the media from tissue explants cultured in vitro and relative to the basal media deprived of tissue explants. This indicated that cells within the explants cultured in vitro for 24 h were viable.
). Data are presented as least square means ± standard error of the mean (SEM) Different letters a–c indicate p-value < 0.05. LDH activity was higher in the media of explants incubated in Triton X-100 at room temperature relative to both the media from tissue explants cultured in vitro and relative to the basal media deprived of tissue explants. This indicated that cells within the explants cultured in vitro for 24 h were viable.
), tissue explants cultured in media (), and tissue explants incubated in Triton X-100 (). Data are presented as least square means ± standard error of the mean (SEM) Different letters a–c indicate p-value < 0.05. LDH activity was higher in the media of explants incubated in Triton X-100 at room temperature relative to both the media from tissue explants cultured in vitro and relative to the basal media deprived of tissue explants. This indicated that cells within the explants cultured in vitro for 24 h were viable.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| In Vivo-Cooled Cows | In Vivo Heat-Stressed Cows | |||||
|---|---|---|---|---|---|---|
| Genes 1 | C-Fold Change | H-Fold Change | C-Fold Change | H-Fold Change | p Value | |
| Early involution | ||||||
| E signalling | ||||||
| BRAF | −1.13 | −1.10 | −9.32 | −2.39 | 0.03 | |
| ESR2 | 1.03 | 1.04 | −2.59 | 1.89 | 0.03 | |
| Mammary growth | ||||||
| E signalling | ||||||
| HSD17B6 | 1.80 | −2.33 | 1.95 | 14.93 | 0.14 | |
| Apoptosis | ||||||
| CASP8 | −1.38 | −1.06 | 1.36 | 1.24 | 0.06 | |
| Autophagy | ||||||
| MAP1LC3B | 1.12 | 1.82 | 1.13 | 1.03 | 0.08 | |
| Inflammation | ||||||
| LBP | −1.09 | 1.33 | 1.37 | −1.05 | 0.01 | |
| Morphology | ||||||
| VEGFA | 1.31 | 1.06 | −1.35 | 1.34 | 0.08 | |
| Proliferation | ||||||
| PIK3R2 | 1.12 | 1.82 | 1.18 | 1.11 | 0.05 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouellet, V.; Negrao, J.; Skibiel, A.L.; Lantigua, V.A.; Fabris, T.F.; Marrero, M.G.; Dado-Senn, B.; Laporta, J.; Dahl, G.E. Endocrine Signals Altered by Heat Stress Impact Dairy Cow Mammary Cellular Processes at Different Stages of the Dry Period. Animals 2021, 11, 563. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11020563
Ouellet V, Negrao J, Skibiel AL, Lantigua VA, Fabris TF, Marrero MG, Dado-Senn B, Laporta J, Dahl GE. Endocrine Signals Altered by Heat Stress Impact Dairy Cow Mammary Cellular Processes at Different Stages of the Dry Period. Animals. 2021; 11(2):563. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11020563
Chicago/Turabian StyleOuellet, Véronique, João Negrao, Amy L. Skibiel, Valerie A. Lantigua, Thiago F. Fabris, Marcela G. Marrero, Bethany Dado-Senn, Jimena Laporta, and Geoffrey E. Dahl. 2021. "Endocrine Signals Altered by Heat Stress Impact Dairy Cow Mammary Cellular Processes at Different Stages of the Dry Period" Animals 11, no. 2: 563. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11020563