
Characterization of Longitudinal Testosterone, Cortisol, and Musth in Male Asian Elephants (Elephas maximus), Effects of Aging, and Adrenal Responses to Social Changes and Health Events

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. Hormone Analysis
2.3. Determination of Musth State
2.3.1. Physical and Behavioral Signs of Musth
2.3.2. Criteria for Defining Musth
2.3.3. Calculation of Individual Musth Thresholds
2.4. Demographics and Social Life Events
2.5. Tuberculosis Diagnosis and Treatment
2.6. Health Declines
2.7. Data Analysis
3. Results
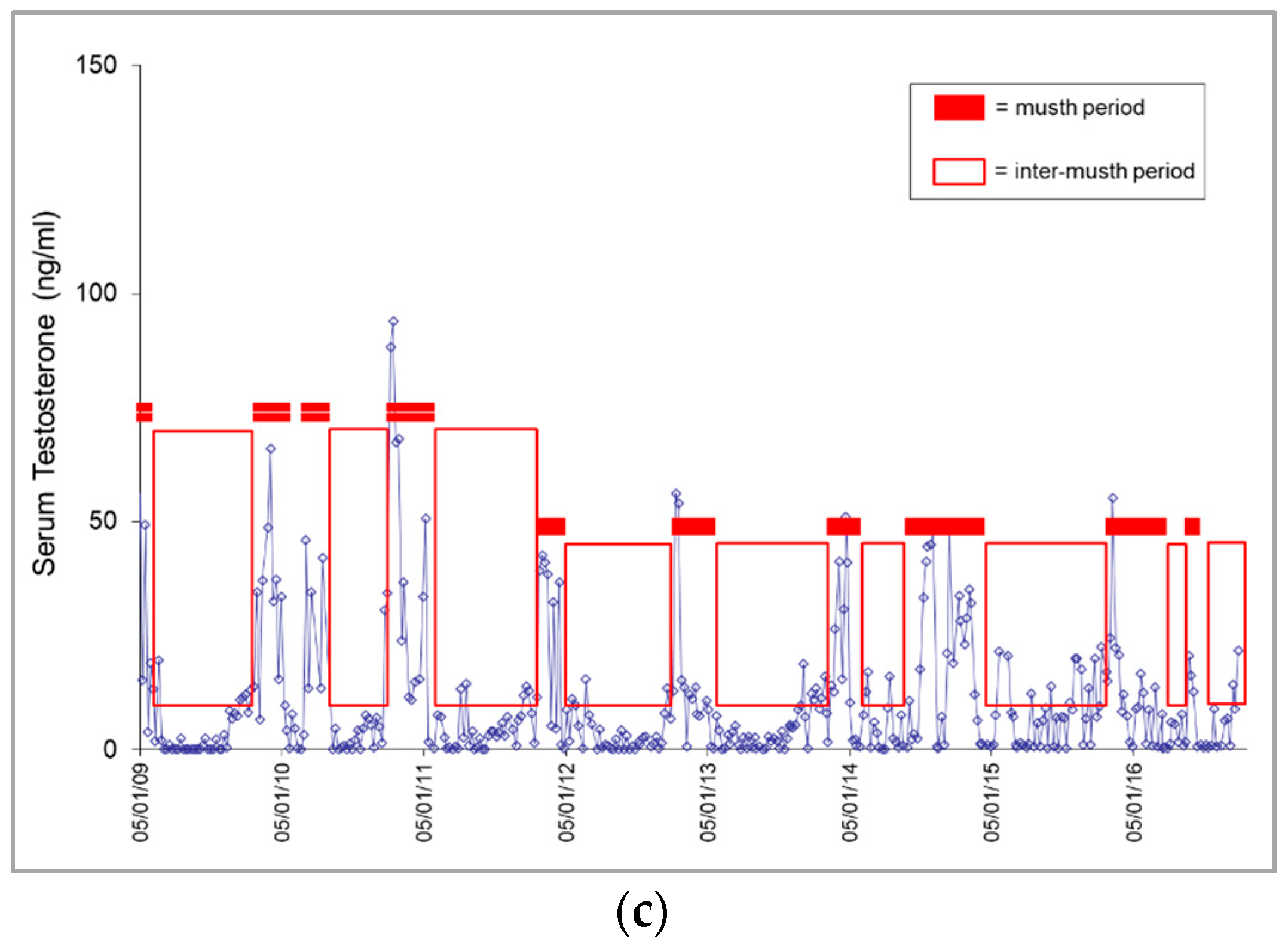
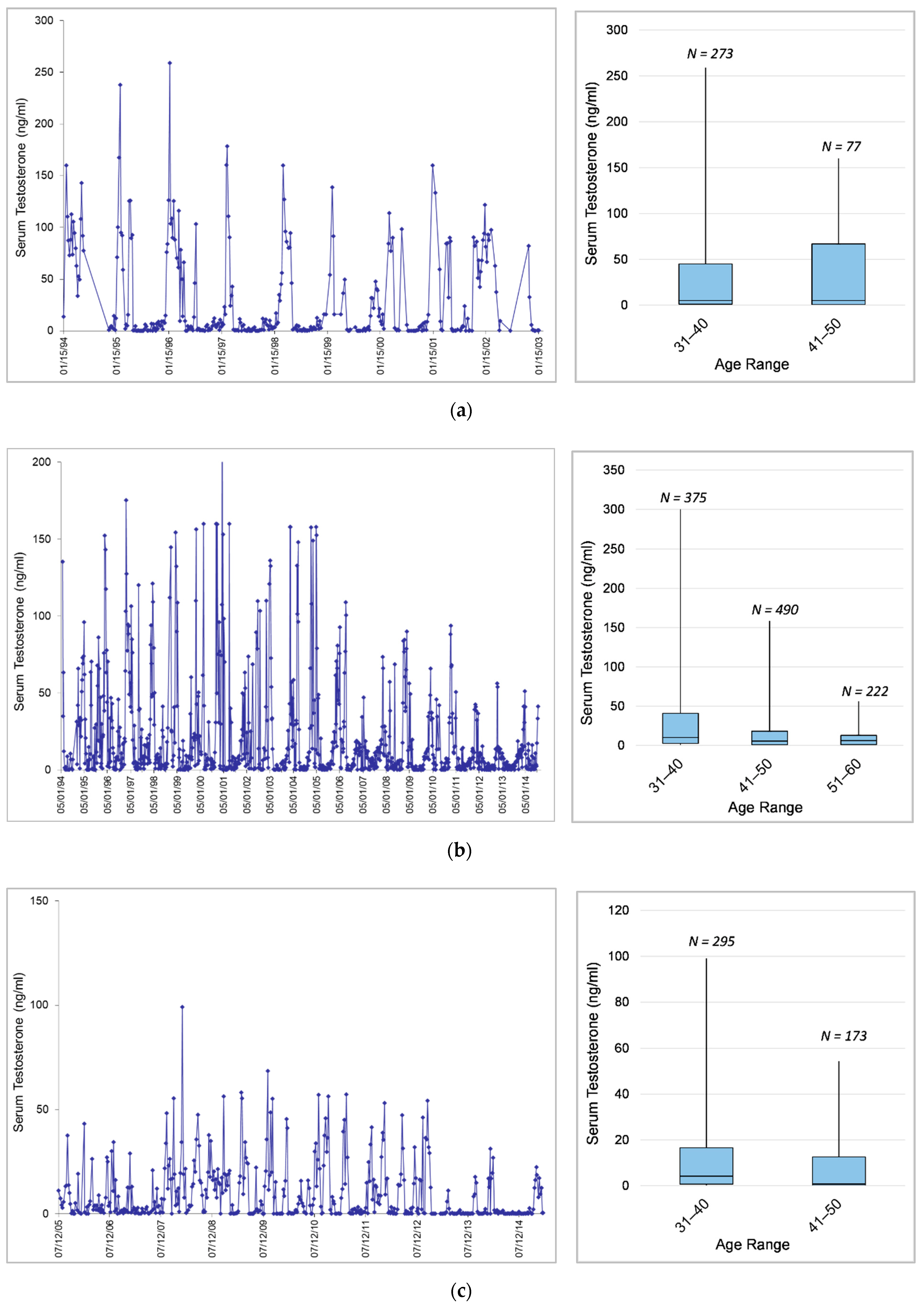
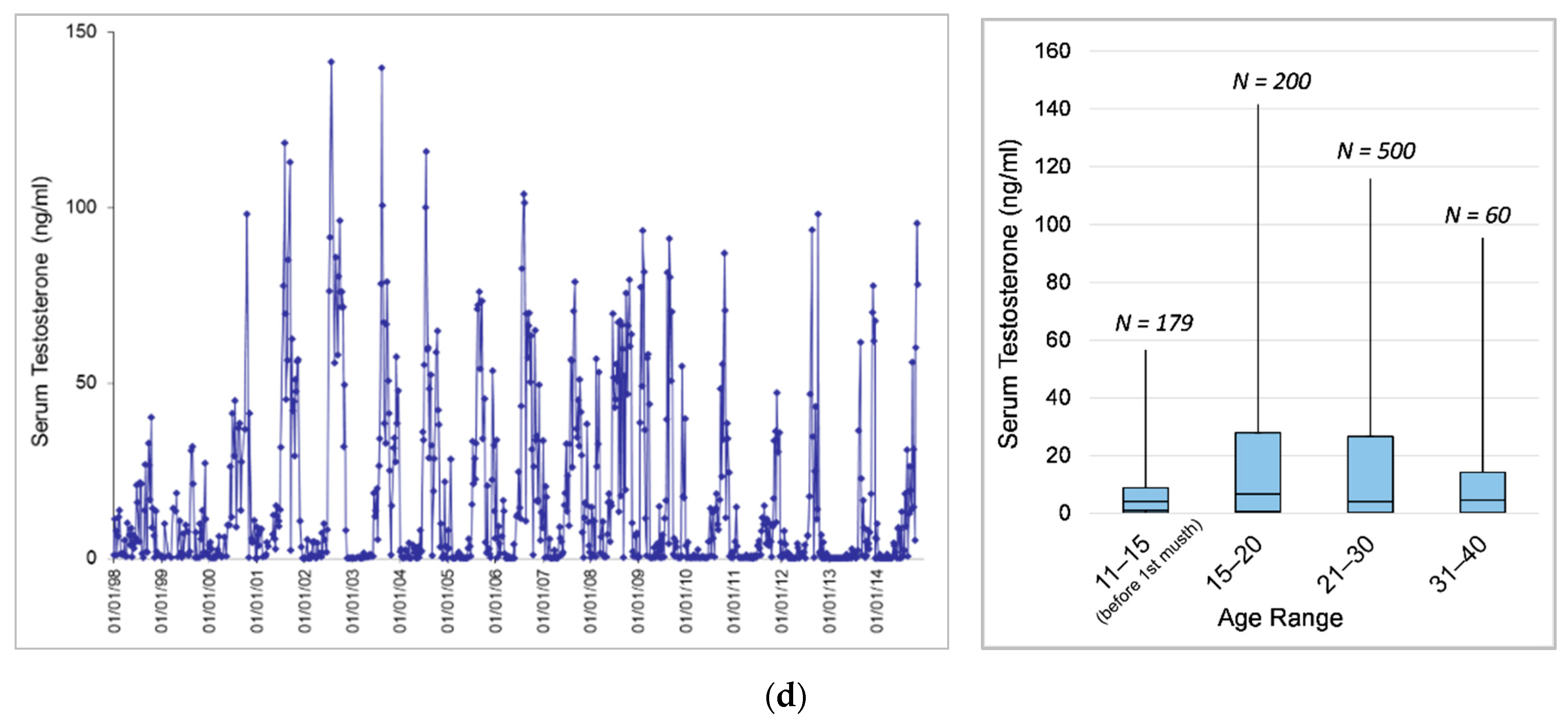
3.1. Longitudinal Serum Testosterone Patterns
3.1.1. Musth Patterns
3.1.2. Musth Threshold Testosterone Concentrations
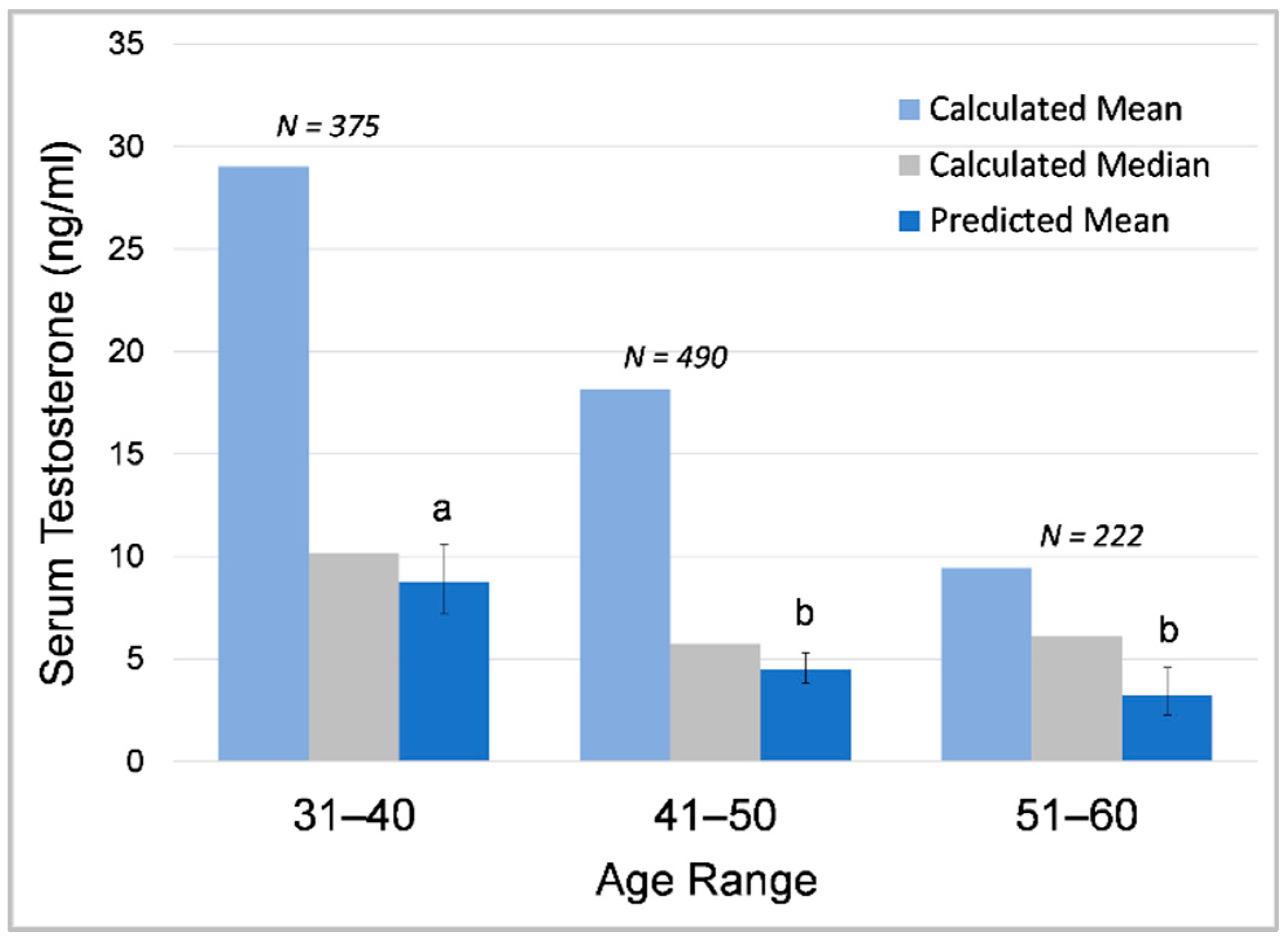
3.1.3. Effects of Age and Dominance Status on Testosterone across Bulls
3.1.4. Effects of Age on Testosterone within Individuals
3.2. Longitudinal Serum Cortisol Patterns
3.2.1. Effects of Testosterone, Musth, and Puberty on Cortisol
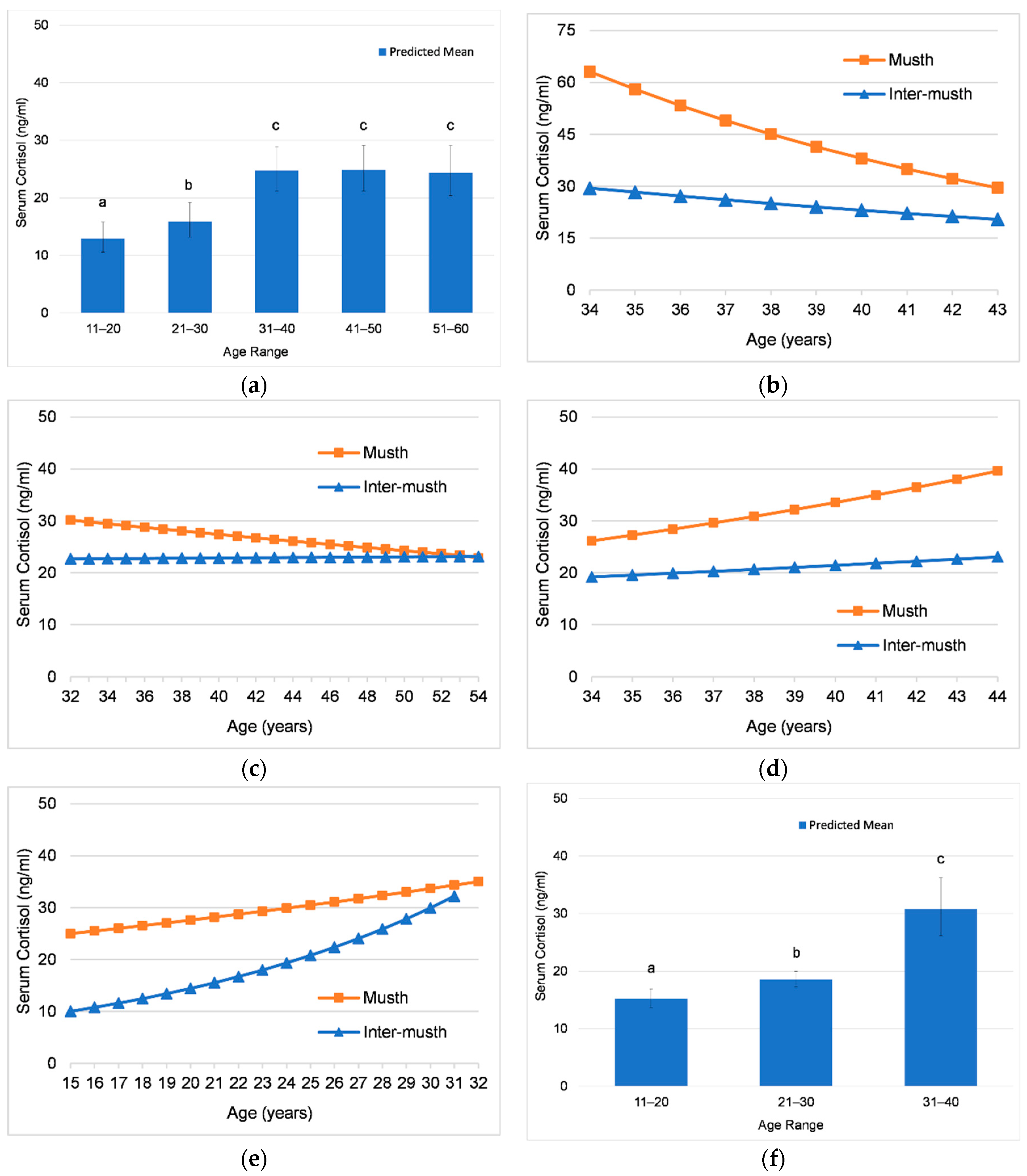
3.2.2. Effects of Age on Cortisol
3.2.3. Effects of Origin of Birth on Cortisol
3.2.4. Effects of Social Life Events on Cortisol
3.2.5. Effects of Mycobacterium Tuberculosis Detection and Treatment on Cortisol
3.2.6. Effects of Health Decline on Cortisol
4. Discussion
4.1. Longitudinal Serum Testosterone Patterns
4.1.1. Musth Patterns
4.1.2. Musth Threshold Testosterone Concentrations
4.1.3. Effects of Age and Dominance Status on Testosterone across Bulls
4.1.4. Effects of Age on Testosterone within Individuals
4.2. Longitudinal Serum Cortisol Patterns
4.2.1. Effects of Puberty, Musth, and Testosterone on Cortisol
4.2.2. Effects of Age on Cortisol across Bulls
4.2.3. Effects of Age on Cortisol within Individuals
4.2.4. Effects of Origin of Birth on Cortisol
4.2.5. Effects of Social Life Events on Cortisol
4.2.6. Effects of Mycobacterium Tuberculosis Detection and Treatment on Cortisol
4.3. Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cooper, K.A.; Harder, J.D.; Clawson, D.H.; Fredrick, D.L.; Lodge, G.A.; Peachey, H.C.; Spellmire, T.J.; Winstel, D.P. Serum testosterone and musth in captive male African and Asian elephants. Zoo Biol. 1990, 9, 297–306. [Google Scholar] [CrossRef]
- Eisenberg, J.F.; McKay, G.M.; Jainudeen, M.R. Reproductive behavior of the Asiatic elephant (Elephas maximus maximus L.). Behaviour 1971, 38, 193–225. [Google Scholar] [CrossRef] [PubMed]
- Jainudeen, M.R.; Katongole, C.B.; Short, R.V. Plasma testosterone levels in relation to musth and sexual activity in the male Asiatic elephant, Elephas maximus. J. Reprod. Fertil. 1972, 29, 99–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lincoln, G.A.; Ratnasooriya, W.D. Testosterone secretion, musth behaviour and social dominance in captive male Asian elephants living near the equator. Reproduction 1996, 108, 107. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.E.; Buss, I.O.; Hess, D.L.; Schmidt, M.J. Testosterone and dihydrotestosterone concentrations in elephant serum and temporal gland secretions. Biol. Reprod. 1984, 30, 352–362. [Google Scholar] [CrossRef] [Green Version]
- Poole, J.H. Rutting behavior in African elephants: The phenomenon of musth. Behaviour 1987, 102, 283–316. [Google Scholar] [CrossRef]
- Poole, J.H. Announcing intent: The aggressive state of musth in African elephants. Anim. Behav. 1989, 37, 140–152. [Google Scholar] [CrossRef]
- Poole, J.H.; Moss, C.J. Musth in the African elephant, Loxodonta africana. Nature 1981, 292, 830–831. [Google Scholar] [CrossRef]
- Hall-Martin, A.J.; Van der Walt, L.A. Plasma testosterone levels in relation to musth in the male African Elephant. Koedoe 1984, 27, 3. [Google Scholar] [CrossRef] [Green Version]
- Poole, J.H. Mate guarding, reproductive success and female choice in African elephants. Anim. Behav. 1989, 37, 842–849. [Google Scholar] [CrossRef]
- Rasmussen, L.E.; Riddle, H.S.; Krishnamurthy, V. Mellifluous matures to malodorous in musth. Nature 2002, 415, 975–976. [Google Scholar] [CrossRef] [PubMed]
- Scott, N.L. Chemical Communication and Musth in Captive Male Elephants. Ph.D. Dissertation, Portland State University, Portland, OR, USA, 2002. [Google Scholar]
- Rasmussen, L.E.L.; Hess, D.L.; Haight, J.D. Chemical analysis of temporal gland secretions collected from an Asian bull elephant during a four-month musth episode. J. Chem. Ecol. 1990, 16, 2167–2181. [Google Scholar] [CrossRef] [PubMed]
- Jainudeen, M.R.; McKay, G.M.; Eisenberg, J.F. Observations on musth in the domesticated Asiatic elephant (Elephas maximus). Mammalia 1972, 36, 247–261. [Google Scholar] [CrossRef]
- Somgird, C.; Sripiboon, S.; Mahasawangkul, S.; Boonprasert, K.; Brown, J.L.; Stout, T.A.; Colenbrander, B.; Thitaram, C. Differential testosterone response to GnRH-induced LH release before and after musth in adult Asian elephant (Elephas maximus) bulls. Theriogenology 2016, 85, 1225–1232. [Google Scholar] [CrossRef]
- Brown, J.L.; Somerville, M.; Riddle, H.S.; Keele, M.; Duer, C.K.; Freeman, E.W. Comparative endocrinology of testicular, adrenal and thyroid function in captive Asian and African elephant bulls. Gen. Comp. Endocrinol. 2007, 151, 153–162. [Google Scholar] [CrossRef]
- Möstl, E.; Palme, R. Hormones as indicators of stress. Domest. Anim. Endocrinol. 2002, 23, 67–74. [Google Scholar] [CrossRef]
- Ralph, C.R.; Tilbrook, A.J. Invited Review: The usefulness of measuring glucocorticoids for assessing animal welfare. J. Anim. Sci. 2016, 94, 457–470. [Google Scholar] [CrossRef] [Green Version]
- Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 2019, 199, 229–243. [Google Scholar] [CrossRef]
- Buwalda, B.; Scholte, J.; de Boer, S.F.; Coppens, C.M.; Koolhaas, J.M. The acute glucocorticoid stress response does not differentiate between rewarding and aversive social stimuli in rats. Horm. Behav. 2012, 61, 218–226. [Google Scholar] [CrossRef]
- Busch, D.S.; Hayward, L.S. Stress in a conservation context: A discussion of glucocorticoid actions and how levels change with conservation-relevant variables. Biol. Conserv. 2009, 142, 2844–2853. [Google Scholar] [CrossRef]
- Menargues, A.; Urios, V.; Mauri, M. Welfare assessment of captive Asian elephants (Elephas maximus) and Indian rhinoceros (Rhinoceros unicornis) using salivary cortisol measurement. Anim. Welf. 2008, 17, 305–312. [Google Scholar]
- Boyle, S.A.; Roberts, B.; Pope, B.M.; Blake, M.R.; Leavelle, S.E.; Marshall, J.J.; Smith, A.; Hadicke, A.; Falcone, J.F.; Knott, K.; et al. Assessment of Flooring Renovations on African Elephant (Loxodonta africana) Behavior and Glucocorticoid Response. PLoS ONE 2015, 10, e0141009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dathe, H.H.; Kuckelkorn, B.; Minnemann, D. Salivary cortisol assessment for stress detection in the Asian elephant (Elephas maximus): A pilot study. Zoo Biol. 1992, 11, 285–289. [Google Scholar] [CrossRef]
- Schmid, J.; Heistermann, M.; Gansloßer, U.; Hodges, J. Introduction of foreign female Asian elephants (Elephas maximus) into an existing group: Behavioural reactions and changes in cortisol levels. Anim. Welf. 2001, 10, 357–372. [Google Scholar]
- Burks, K.D.; Mellen, J.D.; Miller, G.W.; Lehnhardt, J.; Weiss, A.; Figueredo, A.J.; Maple, T.L. Comparison of two introduction methods for African elephants (Loxodonta africana). Zoo Biol. 2004, 23, 109–126. [Google Scholar] [CrossRef]
- Chave, E.; Edwards, K.L.; Paris, S.; Prado, N.; Morfeld, K.A.; Brown, J.L. Variation in metabolic factors and gonadal, pituitary, thyroid, and adrenal hormones in association with musth in African and Asian elephant bulls. Gen. Comp. Endocrinol. 2019, 276, 1–13. [Google Scholar] [CrossRef]
- Yon, L.; Kanchanapangka, S.; Chaiyabutr, N.; Meepan, S.; Stanczyk, F.Z.; Dahl, N.; Lasley, B. A longitudinal study of LH, gonadal and adrenal steroids in four intact Asian bull elephants (Elephas maximus) and one castrate African bull (Loxodonta africana) during musth and non-musth periods. Gen. Comp. Endocrinol. 2007, 151, 241–245. [Google Scholar] [CrossRef]
- Glaeser, S.S.; Edwards, K.L.; Wielebnowski, N.; Brown, J.L. Effects of physiological changes and social life events on adrenal glucocorticoid activity in female zoo-housed Asian elephants (Elephas maximus). PLoS ONE 2020, 15, e0241910. [Google Scholar] [CrossRef]
- Fanson, K.V.; Keeley, T.; Fanson, B.G. Cyclic changes in cortisol across the estrous cycle in parous and nulliparous Asian elephants. Endocr. Connect. 2014, 3, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Kajaysri, J.; Nokkaew, W. Assessment of pregnancy status of Asian elephants (Elephas maximus) by measurement of progestagen and glucocorticoid and their metabolite concentrations in serum and feces, using enzyme immunoassay (EIA). J. Vet. Med. Sci. 2014, 76, 363–368. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L.; Lehnhardt, J. Serum and urinary hormones during pregnancy and the peri- and postpartum period in an Asian elephant (Elephas, maximus). Zoo Biol. 1995, 14, 555–564. [Google Scholar] [CrossRef]
- Liptrap, R.; Raeside, J. Effect of corticotrophin and corticosteroids on plasma interstitial cell-stimulating hormone and urinary steroids in the boar. J. Endocrinol. 1968, 42, 33–43. [Google Scholar] [CrossRef]
- Howland, B.E.; Sanford, L.M.; Palmer, W.M. Changes in serum levels of LH, FSH, prolactin, testosterone, and cortisol associated with season and mating in male pygmy goats. J. Androl. 1985, 6, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Borg, K.E.; Esbenshade, K.L.; Johnson, B.H. Cortisol, growth hormone, and testosterone concentrations during mating behavior in the bull and boar. J. Anim. Sci. 1991, 69, 3230–3240. [Google Scholar] [CrossRef] [Green Version]
- Liptrap, R.M.; Raeside, J.I. A relationship between plasma concentrations of testosterone and corticosteroids during sexual and aggressive behaviour in the boar. J. Endocrinol. 1978, 76, 75–85. [Google Scholar] [CrossRef]
- Mooring, M.S.; Patton, M.L.; Lance, V.A.; Hall, B.M.; Schaad, E.W.; Fetter, G.A.; Fortin, S.S.; McPeak, K.M. Glucocorticoids of bison bulls in relation to social status. Horm. Behav. 2006, 49, 369–375. [Google Scholar] [CrossRef]
- Sands, J.; Creel, S. Social dominance, aggression and faecal glucocorticoid levels in a wild population of wolves, Canis lupus. Anim. Behav. 2004, 67, 387–396. [Google Scholar] [CrossRef]
- Evans, K.E.; Harris, S. Adolescence in male African elephants, Loxodonta africana, and the importance of sociality. Anim. Behav. 2008, 76, 779–787. [Google Scholar] [CrossRef]
- Keerthipriya, P.; Nandini, S.; Vidya, T.N.C. Effects of Male Age and Female Presence on Male Associations in a Large, Polygynous Mammal in Southern India: The Asian Elephant. Front. Ecol. Evol. 2021, 9, 348. [Google Scholar] [CrossRef]
- Goldenberg, S.Z.; de Silva, S.; Rasmussen, H.B.; Douglas-Hamilton, I.; Wittemyer, G. Controlling for behavioural state reveals social dynamics among male African elephants, Loxodonta africana. Anim. Behav. 2014, 95, 111–119. [Google Scholar] [CrossRef]
- Hartley, M.; Wood, A.; Yon, L. Facilitating the social behaviour of bull elephants in zoos. Int. Zoo Yearb. 2019, 53, 62–77. [Google Scholar] [CrossRef]
- Ganswindt, A.; Muenscher, S.; Henley, M.; Henley, S.; Heistermann, M.; Palme, R.; Thompson, P.; Bertschinger, H. Endocrine correlates of musth and the impact of ecological and social factors in free-ranging African elephants (Loxodonta africana). Horm. Behav. 2010, 57, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Vidya, T.N.C.; Sukumar, R. Social and reproductive behaviour in elephants. Curr. Sci. 2005, 89, 1200–1207. [Google Scholar]
- Brown, J.L.; Kersey, D.C.; Freeman, E.W.; Wagener, T. Assessment of Diurnal Urinary Cortisol Excretion in Asian and African Elephants Using Different Endocrine Methods. Zoo Biol. 2010, 29, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.E.L.; Perrin, T.E. Physiological Correlates of Musth: Lipid Metabolites and Chemical Composition of Exudates. Physiol. Behav. 1999, 67, 539–549. [Google Scholar] [CrossRef]
- Edwards, K.L.; Bansiddhi, P.; Paris, S.; Galloway, M.; Brown, J.L. The development of an immunoassay to measure immunoglobulin A in Asian elephant feces, saliva, urine and serum as a potential biomarker of well-being. Conserv. Physiol. 2019, 7, coy077. [Google Scholar] [CrossRef]
- Brown, J.L.; Bush, M.; Wildt, D.E.; Raath, J.R.; de Vos, V.; Howard, J.G. Effects of GnRH analogues on pituitary-testicular function in free-ranging African elephants (Loxodonta africana). J. Reprod. Fertil. 1993, 99, 627–634. [Google Scholar] [CrossRef] [Green Version]
- Schulte, B.A.; Rasmussen, L.E.L. Musth, Sexual Selection, Testosterone, and Metabolites. In Advances in Chemical Signals in Vertebrates; Johnston, R.E., Müller-Schwarze, D., Sorensen, P.W., Eds.; Springer: Boston, MA, USA, 1999; pp. 383–397. [Google Scholar]
- Lee, B. AZA Regional Studbook for Asian Elephant (Elephas maximus); Association of Zoos and Aquariums: Portland, OR, USA, 2019. [Google Scholar]
- Miller, M.A.; Finnegan, M.; Storms, T.; Garner, M.; Lyashchenko, K.P. Outbreak of Mycobacterium Tuberculosis in a Herd of Captive Asian Elephants (Elephas maximus): Antemortem Diagnosis, Treatement, and Lessons Learned. J. Zoo Wildl. Med. 2018, 49, 747–754. [Google Scholar] [CrossRef]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.-S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Harrison, X.A.; Donaldson, L.; Correa-Cano, M.E.; Evans, J.; Fisher, D.N.; Goodwin, C.E.D.; Robinson, B.S.; Hodgson, D.J.; Inger, R. A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 2018, 6, e4794. [Google Scholar] [CrossRef] [Green Version]
- Edwards, K.L.; Shultz, S.; Pilgrim, M.; Walker, S.L. Irregular ovarian activity, body condition and behavioural differences are associated with reproductive success in female eastern black rhinoceros (Diceros bicornis michaeli). Gen. Comp. Endocrinol. 2015, 214, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.L.; Trotter, J.; Jones, M.; Brown, J.L.; Steinmetz, H.W.; Walker, S.L. Investigating temporary acyclicity in a captive group of Asian elephants (Elephas maximus): Relationship between management, adrenal activity and social factors. Gen. Comp. Endocrinol. 2016, 225, 104–116. [Google Scholar] [CrossRef]
- LaDue, C.A.; Schulte, B.A.; Kiso, W.K.; Freeman, E.W. Musth and sexual selection in elephants: A review of signalling properties and potential fitness consequences. Behaviour 2021, 1, 1–36. [Google Scholar] [CrossRef]
- Niemuller, C.A.; Liptrap, R.M. Altered androstenedione to testosterone ratios and LH concentrations during musth in the captive male Asian elephant (Elephas maximus). J. Reprod. Fertil. 1991, 91, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Scallan, J.; Glaeser, S.; Backues, K.; Freeman, E. Serum Testosterone Concentrations in an ex situ male Asian Elephant (Elephas maximus) in relation to a temporary breeding loan. J. Elephant Manag. Assoc. (JEMA) 2011, 22, 20–33. [Google Scholar]
- Rasmussen, H.B.; Ganswindt, A.; Douglas-Hamilton, I.; Vollrath, F. Endocrine and behavioral changes in male African elephants: Linking hormone changes to sexual state and reproductive tactics. Horm. Behav. 2008, 54, 539–548. [Google Scholar] [CrossRef]
- Scott, N.L.; Riddle, H. Assessment of Musth in Captivity: A Survey of Factors Affecting the Frequency and Duration of Musth in Captive Male Elephants Elephas maximus—Loxodonta africana. J. Elephant Manag. Assoc. (JEMA) 2003, 14, 11–15. [Google Scholar]
- LaDue, C.; Scott, N.; Margulis, S. A survey of musth among captive male elephants in North America: Updated results and implications for management. J. Elephant Manag. Assoc. (JEMA) 2014, 25, 18–24. [Google Scholar]
- Keerthipriya, P.; Vidya, T.N.C. A Comparison of the Occurrence of Musth in the Kabini Elephant Population with Other Populations. Gajah 2019, 50, 4–16. [Google Scholar]
- Thongtip, N.; Saikhun, J.; Mahasawangkul, S.; Kornkaewrat, K.; Pongsopavijitr, P.; Songsasen, N.; Pinyopummin, A. Potential factors affecting semen quality in the Asian elephant (Elephas maximus). Reprod. Biol. Endocrinol. 2008, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Leung, A.; Sinha-Hikim, A.P. Reproductive aging in the male brown-Norway rat: A model for the human. Endocrinology 1993, 133, 2773–2781. [Google Scholar] [CrossRef] [PubMed]
- Bremner, W.J.; Vitiello, M.V.; Prinz, P.N. Loss of Circadian Rhythmicity in Blood Testosterone Levels with Aging in Normal Men. J. Clin. Endocrinol. Metab. 1983, 56, 1278–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morley, J.E.; Kaiser, F.E.; Perry, H.M.; Patrick, P.; Morley, P.M.K.; Stauber, P.M.; Vellas, B.; Baumgartner, R.N.; Garry, P.J. Longitudinal changes in testosterone, luteinizing hormone, and follicle-stimulating hormone in healthy older men. Metabolism 1997, 46, 410–413. [Google Scholar] [CrossRef]
- Muller, M.N.; Wrangham, R.W. Dominance, cortisol and stress in wild chimpanzees (Pan troglodytes schweinfurthii). Behav. Ecol. Sociobiol. 2004, 55, 332–340. [Google Scholar] [CrossRef] [Green Version]
- Ganswindt, A.; Palme, R.; Heistermann, M.; Borragan, S.; Hodges, J. Non-invasive assessment of adrenocortical function in the male African elephant (Loxodonta africana) and its relation to musth. Gen. Comp. Endocrinol. 2003, 134, 156–166. [Google Scholar] [CrossRef]
- Ganswindt, A.; Rasmussen, H.B.; Heistermann, M.; Hodges, J.K. The sexually active states of free-ranging male African elephants (Loxodonta africana): Defining musth and non-musth using endocrinology, physical signals, and behavior. Horm. Behav. 2005, 47, 83–91. [Google Scholar] [CrossRef]
- Ganswindt, A.; Heistermann, M.; Hodges, K. Physical, Physiological, and Behavioral Correlates of Musth in Captive African Elephants (Loxodonta africana). Physiol. Biochem. Zool. 2005, 78, 505–514. [Google Scholar] [CrossRef]
- Ghosal, R.; Ganswindt, A.; Seshagiri, P.B.; Sukumar, R. Endocrine Correlates of Musth in Free-Ranging Asian Elephants (Elephas maximus) Determined by Non-Invasive Faecal Steroid Hormone Metabolite Measurements. PLoS ONE 2013, 8, e84787. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Reddy, V.P.; Kokkiligadda, A.; Shivaji, S.; Umapathy, G. Non-invasive assessment of reproductive status and stress in captive Asian elephants in three south Indian zoos. Gen. Comp. Endocrinol. 2014, 201, 37–44. [Google Scholar] [CrossRef]
- Goymann, W. On the use of non-invasive hormone research in uncontrolled, natural environments: The problem with sex, diet, metabolic rate and the individual. Methods Ecol. Evol. 2012, 3, 757–765. [Google Scholar] [CrossRef]
- Barboza, P.S.; Hartbauer, D.W.; Hauer, W.E.; Blake, J.E. Polygynous mating impairs body condition and homeostasis in male reindeer (Rangifer tarandus tarandus). J. Comp. Physiol. B 2004, 174, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Khonmee, J.; Brown, J.L.; Rojanasthien, S.; Thumasanukul, D.; Kongphoemphun, A.; Siriaroonrat, B.; Tipkantha, W.; Punyapornwithaya, V.; Thitaram, C. Seasonality of fecal androgen and glucocorticoid metabolite excretion in male goral (Naemorhedus griseus) in Thailand. Anim. Reprod. Sci. 2014, 146, 70–78. [Google Scholar] [CrossRef]
- Metrione, L.C.; Hunter, D.; Penfold, L.M. Seasonal Changes in Fecal Glucocorticoid Metabolite Concentrations in Bison (Bison bison) Living with or without Wolves (Canis lupus). J. Wildl. Dis. 2020, 56, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.L.; Miller, M.A.; Carlstead, K.; Brown, J.L. Relationships between housing and management factors and clinical health events in elephants in North American zoos. PLoS ONE 2019, 14, e0217774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.L.; Carlstead, K.; Bray, J.D.; Dickey, D.; Farin, C.; Ange-van Heugten, K. Individual and environmental risk factors associated with fecal glucocorticoid metabolite concentrations in zoo-housed Asian and African elephants. PLoS ONE 2019, 14, e0217326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norkaew, T.; Brown, J.L.; Bansiddhi, P.; Somgird, C.; Thitaram, C.; Punyapornwithaya, V.; Punturee, K.; Vongchan, P.; Somboon, N.; Khonmee, J. Influence of season, tourist activities and camp management on body condition, testicular and adrenal steroids, lipid profiles, and metabolic status in captive Asian elephant bulls in Thailand. PLoS ONE 2019, 14, e0210537. [Google Scholar] [CrossRef] [Green Version]
- McEwen, B.S.; Wingfield, J.C. The concept of allostasis in biology and biomedicine. Horm. Behav. 2003, 43, 2–15. [Google Scholar] [CrossRef]
- Creel, S.; Dantzer, B.; Goymann, W.; Rubenstein, D.R. The ecology of stress: Effects of the social environment. Funct. Ecol. 2013, 27, 66–80. [Google Scholar] [CrossRef] [Green Version]
- Hawkley, L.C.; Cole, S.W.; Capitanio, J.P.; Norman, G.J.; Cacioppo, J.T. Effects of social isolation on glucocorticoid regulation in social mammals. Horm. Behav. 2012, 62, 314–323. [Google Scholar] [CrossRef] [Green Version]
- Prado-Oviedo, N.A.; Bonaparte-Saller, M.K.; Malloy, E.J.; Meehan, C.L.; Mench, J.A.; Carlstead, K.; Brown, J.L. Evaluation of Demographics and Social Life Events of Asian (Elephas maximus) and African Elephants (Loxodonta africana) in North American Zoos. PLoS ONE 2016, 11, e0154750. [Google Scholar] [CrossRef]
- Greco, B.J.; Meehan, C.L.; Hogan, J.N.; Leighty, K.A.; Mellen, J.; Mason, G.J.; Mench, J.A. The Days and Nights of Zoo Elephants: Using Epidemiology to Better Understand Stereotypic Behavior of African Elephants (Loxodonta africana) and Asian Elephants (Elephas maximus) in North American Zoos. PLoS ONE 2016, 11, e0144276. [Google Scholar] [CrossRef] [PubMed]
- Bottasso, O.; Bay, M.L.; Besedovsky, H.; Del Rey, A. The Immuno-endocrine Component in the Pathogenesis of Tuberculosis. Scand. J. Immunol. 2007, 66, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Rosen, L.E.; Olea-Popelka, F.; Deem, S.L.; Isaza, R.; Schmitt, D.; Miller, M. Survey of Antituberculosis Drug Administration and Adverse Effects in Elephants in North America. J. Zoo Wildl. Med. 2019, 50, 23–32. [Google Scholar] [CrossRef]
- Samaras, N.; Samaras, D.; Frangos, E.; Forster, A.; Philippe, J. A review of age-related dehydroepiandrosterone decline and its association with well-known geriatric syndromes: Is treatment beneficial? Rejuvenation Res. 2013, 16, 285–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Rodríguez, L.; Blas, J.; Viñuela, J.; Marchant, T.A.; Bortolotti, G.R. Condition and androgen levels: Are condition-dependent and testosterone-mediated traits two sides of the same coin? Anim. Behav. 2006, 72, 97–103. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elephant | Origin | Birth Year | Age Range (Years) | Offspring Sired | Effects Analyzed |
|---|---|---|---|---|---|
| Male1 | Wild | 1960 | ~34–43 | 4 | M, Age, DS |
| Male2 | Zoo-born Oregon Zoo | 1962 | 32–54 | 7 | M, Age, TM, DM, Tbactive, TBtr, DS |
| Male3 | Wild | 1971 | ~34–44 | 6 | M, Age, DM, Tbactive, TBtr, DS |
| Male4 | Zoo-born Oregon Zoo | 1983 | 12–32 | 0 | R, M, Age, TM, DM, Tbactive, TBtr, DS |
| Scale | Definition | Pictoral Definition | |
|---|---|---|---|
| 0 | No visible swelling or secretion. |  |  |
| 1 | Swollen temporal gland area. Opening may be enlarged. May not be symmetrical. |  |  |
| 2 | Temporal gland area wet. TGS less than 1/4 way to jawline. |  | |
| 3 | TGS between 1/4 and 3/4 way to jawline. |  |  |
| 4 | TGS from 3/4 to all the way to jawline. |  | |
| 5 | TGS staining is dried (lighter color than wet). |  | |
| Scale | Definition | Pictoral Definition | |
|---|---|---|---|
| 0 | No visible UD. Urination occurs normally (with penis fully extended). |  |  |
| 1 | Occasional drops (without penis extended). Urination occurs with penis partially extended. |  |  |
| 2 | Regular drops and/or some steady streams (without penis extended). Urination occurs with penis less extended less than #1. Legs (upper or lower) a little wet or stained with urine. |  |  |
| 3 | Steady streams from the opening and some dribbling from the skin of the sheath (without penis extended). Penis does not drop to urinate. Legs (upper or lower) half wet or stained with urine. Skin around sheath wet with urine. |  |  |
| 4 | Heavy steady streams falling from the opening and skin of the sheath so the stream looks wider than #3 (without penis extended). Penis does not drop to urinate. Legs entirely wet with urine. Skin around sheath wet with urine. |  | |
| 5 | UD staining is dried (lighter color than wet). |  | |
| Scale | Definition |
|---|---|
| 0 | No aggression, moves readily |
| 1 | Somewhat “spacey” or uncooperative |
| 2 | Reluctant to move, bangs on doors occasionally |
| 3 | Considerable hesitation moving, bangs on doors often |
| 4 | Very aggressive, refuses to move |
| Musth Status | Serum Testosterone | TGS a | UD b | Behavior c | |
|---|---|---|---|---|---|
| Concentration | Duration | ||||
| Inter-musth and Non-musth | <10 ng/mL | >2 weeks | 0 d | 0 | 0 d |
| Pre-musth | >10 ng/mL | no criteria | 1 | 0 | 1–4 |
| Musth | >10 ng/mL | >4 weeks (including pre/post musth) | 2–4 | 1–4 | 1–4 |
| Post-musth | >10 ng/mL | no criteria | 5 e | 5 e | 1–4 |
| Overall | Inter-Musth Period a | Musth Period | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Individual | Origin of Birth | Age Range (Years) | N | Median (Range) | Mean (SD) | CV% | Median (Range) | Mean (SD) | CV% | Median (Range) | Mean (SD) | CV% |
| Male1 | Wild | ~34–43 | 350 | 5.07 (0.10–258.88) | 29.49 (44.45) | 150.74 | 1.98 (0.10–103.42) | 5.31 (11.44) | 215.99 | 82.10 (0.55–258.88) | 79.53 (45.65) | 57.39 |
| Male2 | Zoo-born | 32–54 | 1087 | 7.25 (0.10–300.26) | 21.19 (33.66) | 158.83 | 2.89 (0.10–120.18) | 6.35 (10.46) | 164.68 | 38.47 (0.10–300.26) | 49.64 (44.16) | 88.95 |
| 34–44 a | 485 b | 8.70 (0.10–300.26) | 28.79 (41.73) | 144.94 | 3.68 (0.10–120.18) | 7.68 (12.08) | 157.28 | 48.53 (0.12–300.26) | 59.39 (49.76) | 83.79 | ||
| Male3 | Wild | ~34–44 | 468 | 2.40 (0.10–99.10) | 9.76 (14.14) | 145.23 | 0.51 (0.10–43.17) | 1.92 (4.25) | 221.58 | 17.23 (0.10–99.10) | 21.31 (16.20) | 76.04 |
| Male4 b | Zoo-born | 12–32 | 939 | 4.60 (0.10–141.65) | 15.24 (23.51) | 154.32 | 0.69 (0.10–76.17) | 3.41 (6.65) | 194.96 | 36.72 (0.10–141.65) | 41.58 (28.57) | 68.72 |
| 11–15 c | 179 | 4.13 (0.10–56.57) | 5.97 (6.76) | 113.12 | - | - | - | - | - | - | ||
| 15–32 d | 760 | 4.67 (0.10–141.65) | 17.24 (26.29) | 146.09 | 0.69 (0.10–76.17) | 3.41 (6.65) | 194.96 | 36.72 (0.10–141.65) | 41.58 (28.57) | 68.72 | ||
| All Elephants | ~12–54 | 2734 | 5.12 (0.10–300.26) | 18.25 (30.26) | 165.78 | 1.34 (0.10–120.18) | 4.56 (8.97) | 196.70 | 34.96 (0.10–300.26) | 46.02 (39.68) | 86.23 | |
| Individual | Threshold Testosterone Concentration (ng/mL) | Testosterone Concentration for First Musth Point Using Methods of Chave et al., 2019 [27] | ||
|---|---|---|---|---|
| Minimum | Median | Mean | ||
| Male1 | 16 | 69 | 72 | 67 |
| Male2 | 11 | 43 | 49 | 51 |
| Male3 | 13 | 16 | 18 | 21 |
| Male4 | 10 | 40 | 39 | 35 |
| Individual | Age Range of Analysis | Age Variable | Effect Size (SE) | Wald | p | Age Effect |
|---|---|---|---|---|---|---|
| Male1 | ~34–43 | Age (years) | −0.066 (0.021) | 9.681 | 0.002 | Decreasing |
| 31–40 (reference) a | ||||||
| 41–50 | −0.097 (0.125) | 0.606 | 0.436 | - | ||
| Male2 | 32–53 | Age (years) | −0.027 (0.004) | 37.043 | <0.001 | Decreasing |
| 31–40 (reference) a | ||||||
| 41–50 | −0.289 (0.056) | 26.262 | <0.001 | Lower | ||
| 51–60 | −0.432 (0.088) | 23.858 | <0.001 | Lower | ||
| Male3 | ~34–44 | Age (years) | −0.090 (0.015) | 38.079 | <0.001 | Decreasing |
| 31–40 (reference) a | ||||||
| 41–50 | −0.495 (0.082) | 36.783 | <0.001 | Lower | ||
| Male4 b | 15–32 | Age (years) | −0.017 (0.005) | 10.509 | 0.001 | Decreasing |
| 11–20 (reference) a | ||||||
| 21–30 | −0.081 (0.063) | 1.655 | 0.198 | - | ||
| 31–40 | 0.167 (0.128) | 1.697 | 0.193 | - |
| All Reproductive States | Inter-Musth Period | Musth Period | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Individual | Origin of Birth | Age Range (Years) | N | Median (Range) | Mean (SD) | CV% | Median (Range) | Mean (SD) | CV% | Median (Range) | Mean (SD) | CV% |
| Male1 | Wild | ~34–43 | 380 | 27.70 (5.40–261.60) | 36.68 (26.51) | 72.29 | 23.30 (5.40–261.60) | 30.28 (25.28) | 83.48 | 44.40 (15.20–125.80) | 49.90 (23.77) | 47.63 |
| Male2 | Zoo-born | 32–54 | 1099 | 23.50 (4.40–141.30) | 28.19 (17.21) | 61.06 | 21.55 (4.40–141.30) | 27.10 (17.78) | 65.60 | 26.95 (5.00–96.00) | 30.18 (16.16) | 53.53 |
| Male3 | Wild | ~34–44 | 525 | 24.80 (2.50–109.70) | 29.92 (19.87) | 66.39 | 19.90 (2.50–106.10) | 24.82 (15.91) | 64.12 | 32.50 (6.50–109.70) | 38.04 (22.24) | 58.48 |
| Male4 a | Zoo-born | 12–32 | 934 | 22.30 (2.50–176.30) | 27.23 (20.08) | 73.75 | 18.50 (2.50–176.30) | 24.72 (20.66) | 83.59 | 33.80 (4.80–128.30) | 34.78 (18.64) | 53.59 |
| All Elephants | ~12–54 | 2938 | 24.00 (2.50–261.60) | 29.29 (20.22) | 69.03 | 24.00 (2.50–261.60) | 26.54 (19.74) | 74.40 | 30.75 (4.80–128.30) | 35.65 (20.26) | 56.85 | |
| Wild-born | ~34–44 | 905 | 26.20 (2.50–261.60) | 32.76 (23.12) | 70.58 | 21.85 (2.50–261.60) | 27.39 (21.01) | 76.71 | 37.15 (6.50–125.80) | 43.08 (23.67) | 59.54 | |
| Zoo-born | 12–54 | 2033 | 23.10 (2.50–176.30) | 27.75 (18.59) | 66.98 | 20.90 (2.50–176.30) | 26.10 (19.06) | 73.04 | 28.90 (4.80–128.30) | 32.14 (17.39) | 54.11 | |
| Variable | Individual(s) | Effect Size (SE) | Wald | p | Effect on Mean Cortisol |
|---|---|---|---|---|---|
| Testosterone | Male1 | 0.129 (0.014) | 85.048 | <0.001 | Positive |
| Male2 | 0.028 (0.009) | 8.504 | 0.004 | Positive | |
| Male3 | 0.128 (0.013) | 61.284 | <0.001 | Positive | |
| Male4 | 0.070 (0.011) | 38.353 | <0.001 | Positive | |
| All Elephants | 0.075 (0.006) | 161.041 | <0.001 | Positive | |
| Inter-musth (reference) a | |||||
| Musth | Male1 | 0.259 (0.027) | 93.600 | <0.001 | Higher |
| Male2 | 0.060 (0.016) | 14.523 | <0.001 | Higher | |
| Male3 | 0.187 (0.024) | 61.284 | <0.001 | Higher | |
| Male4 | 0.204 (0.024) | 74.687 | <0.001 | Higher | |
| All Elephants | 0.152 (0.011) | 194.919 | <0.001 | Higher |
| Individual | Age Range of Analysis | Age Variable | Effect Size (SE) | Wald | p | Age Effect |
|---|---|---|---|---|---|---|
| Male1 | ~34–43 | Age (years) | −0.026 (0.005) | 32.714 | <0.001 | Decreasing |
| 31–40 (reference) b | ||||||
| 41–50 | −0.110 (0.029) | 14.189 | <0.001 | Lower | ||
| Interaction of Age and Musth | −0.019 (0.009) | 4.346 | 0.037 | Significant | ||
| Male2 | 32–54 | Age (years) | −0.002 (0.001) | 2.079 | 0.149 | -- |
| 31–40 (reference) b | ||||||
| 41–50 | −0.028 (0.017) | 2.714 | 0.099 | -- | ||
| 51–60 | −0.022 (0.021) | 1.092 | 0.296 | -- | ||
| Interaction of Age and Musth | −0.006 (0.002) | 5.605 | 0.018 | Significant | ||
| Male3 | ~34–44 | Age (years) | 0.011 (0.004) | 8.054 | 0.021 | Increasing |
| 31–40 (reference) b | ||||||
| 41–50 | 0.113 (0.023) | 24.856 | <0.001 | Higher | ||
| Interaction of Age and Musth | 0.010 (0.008) | 1.451 | 0.228 | -- | ||
| Male4 a | 15–32 | Age (years) | 0.025 (0.002) | 118.624 | <0.001 | Increasing |
| 11–20 (reference) b | ||||||
| 21–30 | 0.087 (0.026) | 11.171 | 0.001 | Higher | ||
| 31–40 | 0.306 (0.042) | 53.358 | <0.001 | Higher | ||
| Interaction of Age and Musth | −0.023 (0.005) | 22.151 | <0.001 | Significant | ||
| All Elephants | 15–54 | Age (years) | 0.004 (0.011) | 19.502 | <0.001 | Increasing |
| 11–20 (reference) b | ||||||
| 21–30 | 0.090 (0.023) | 15.450 | <0.001 | Higher | ||
| 31–40 | 0.283 (0.035) | 65.123 | <0.001 | Higher | ||
| 41–50 | 0.284 (0.037) | 58.378 | <0.001 | Higher | ||
| 51–60 | 0.276 (0.041) | 44.934 | <0.001 | Higher | ||
| Interaction of Age and Musth | −0.007 (0.001) | 42.213 | <0.001 | Significant |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glaeser, S.S.; Edwards, K.L.; Paris, S.; Scarlata, C.; Lee, B.; Wielebnowski, N.; Finnell, S.; Somgird, C.; Brown, J.L. Characterization of Longitudinal Testosterone, Cortisol, and Musth in Male Asian Elephants (Elephas maximus), Effects of Aging, and Adrenal Responses to Social Changes and Health Events. Animals 2022, 12, 1332. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101332
Glaeser SS, Edwards KL, Paris S, Scarlata C, Lee B, Wielebnowski N, Finnell S, Somgird C, Brown JL. Characterization of Longitudinal Testosterone, Cortisol, and Musth in Male Asian Elephants (Elephas maximus), Effects of Aging, and Adrenal Responses to Social Changes and Health Events. Animals. 2022; 12(10):1332. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101332
Chicago/Turabian StyleGlaeser, Sharon S., Katie L. Edwards, Stephen Paris, Candace Scarlata, Bob Lee, Nadja Wielebnowski, Shawn Finnell, Chaleamchat Somgird, and Janine L. Brown. 2022. "Characterization of Longitudinal Testosterone, Cortisol, and Musth in Male Asian Elephants (Elephas maximus), Effects of Aging, and Adrenal Responses to Social Changes and Health Events" Animals 12, no. 10: 1332. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101332