
The Alimentary Tract of African Bony-Tongue, Heterotis niloticus (Cuvier, 1829): Morphology Study

 ,
,  , , ,
, , ,  , and
, and 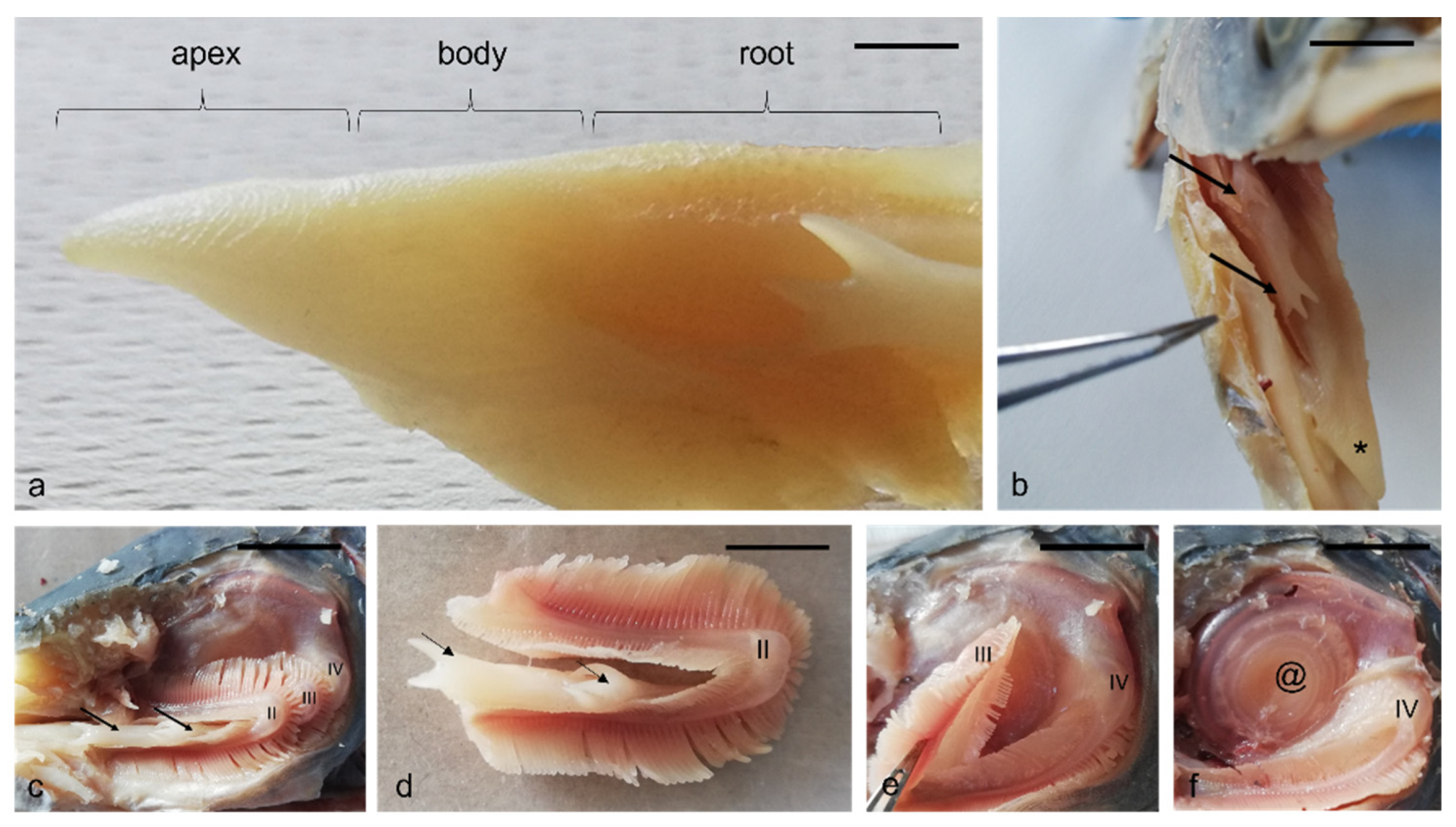{kind=link}
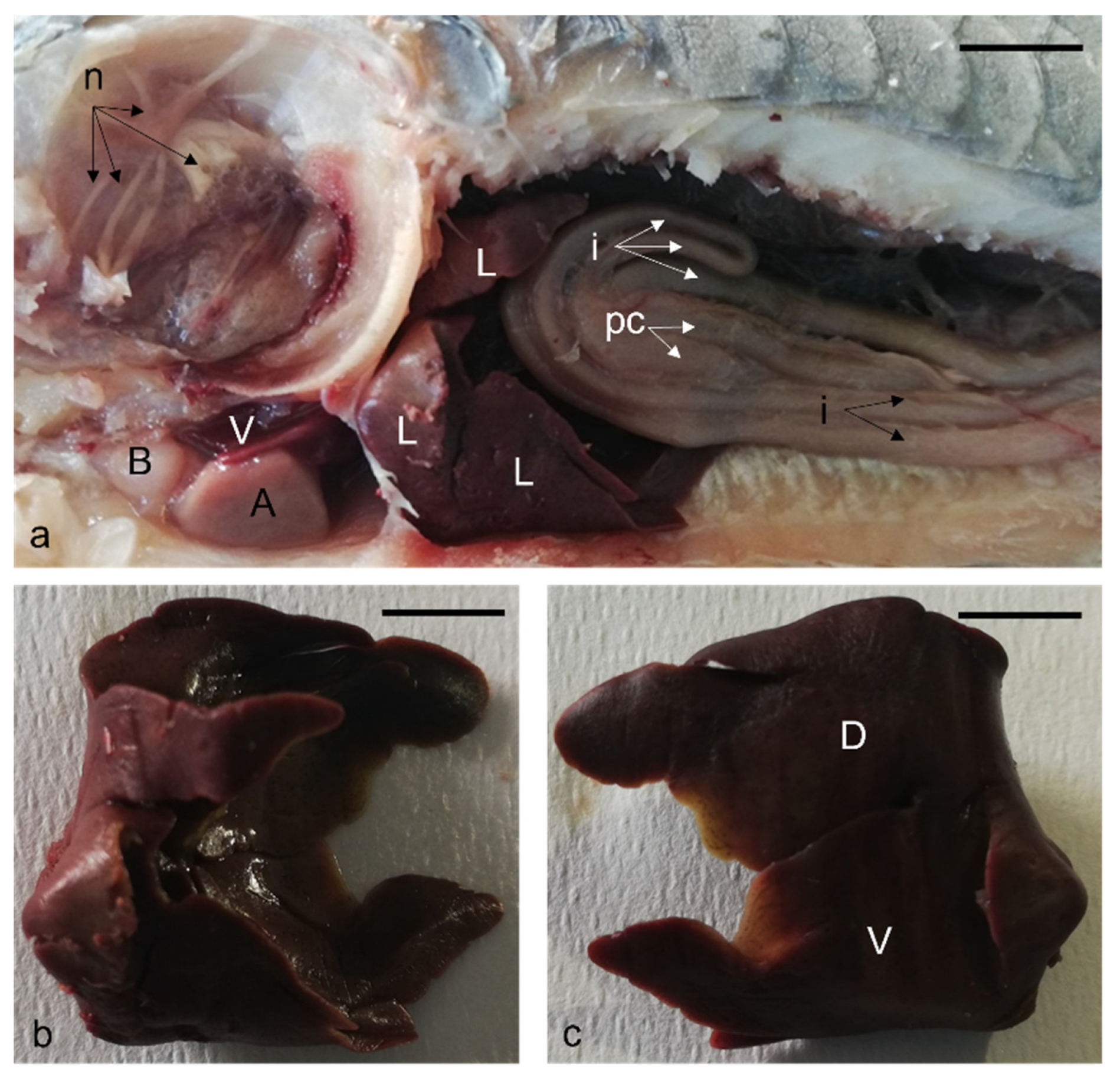{kind=link}
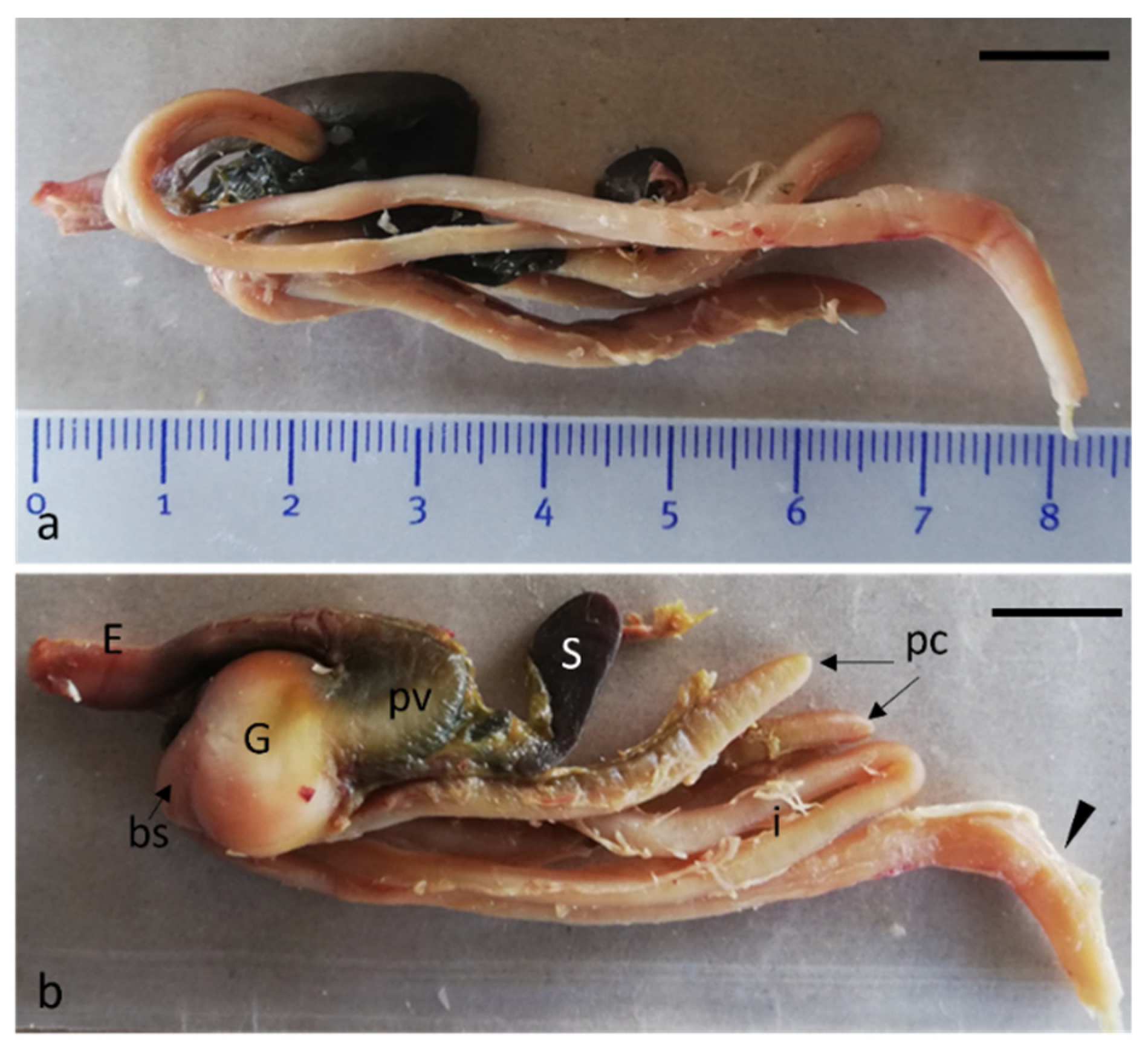{kind=link}
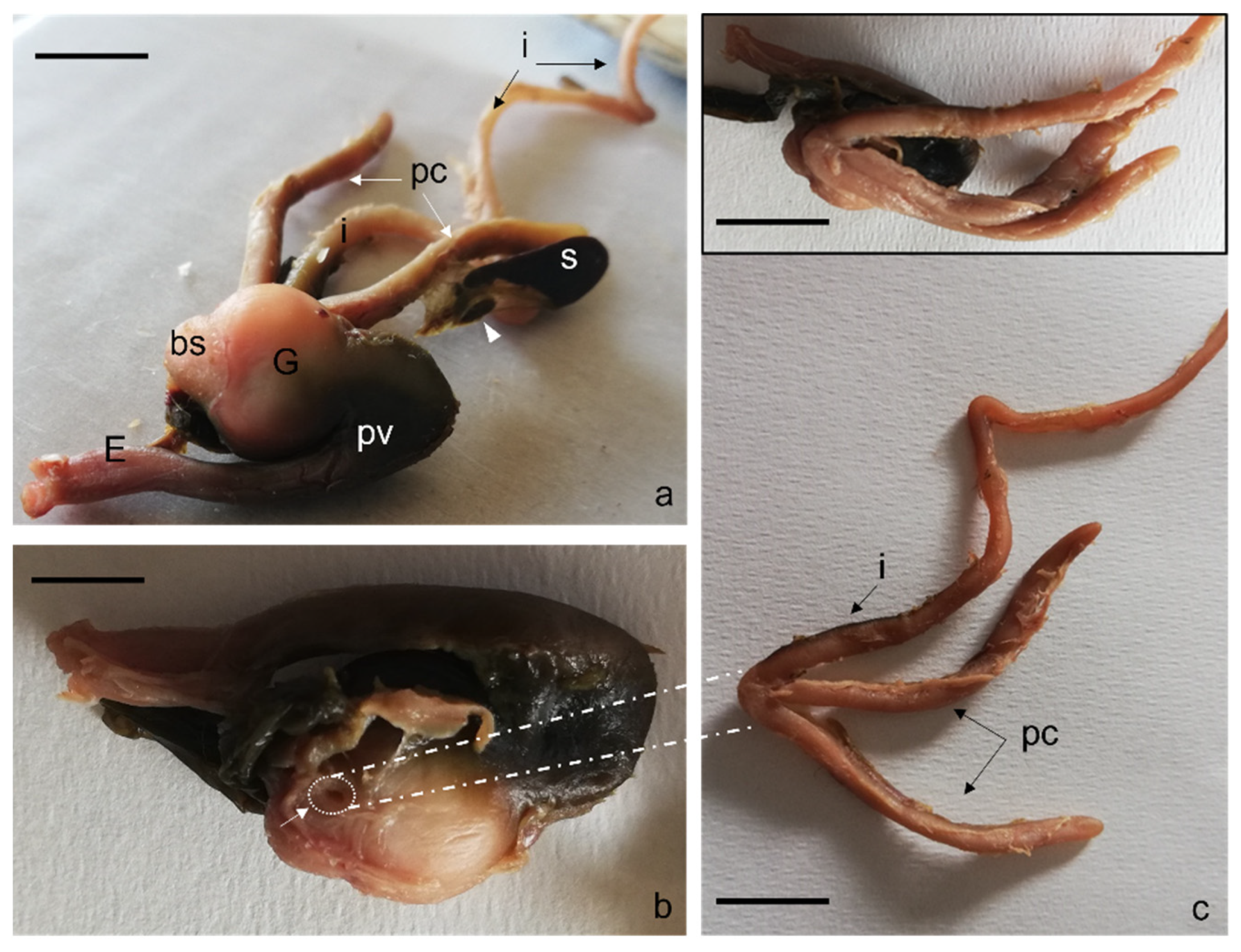{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Histochemical Staining
2.2. Immunocytochemical Staining
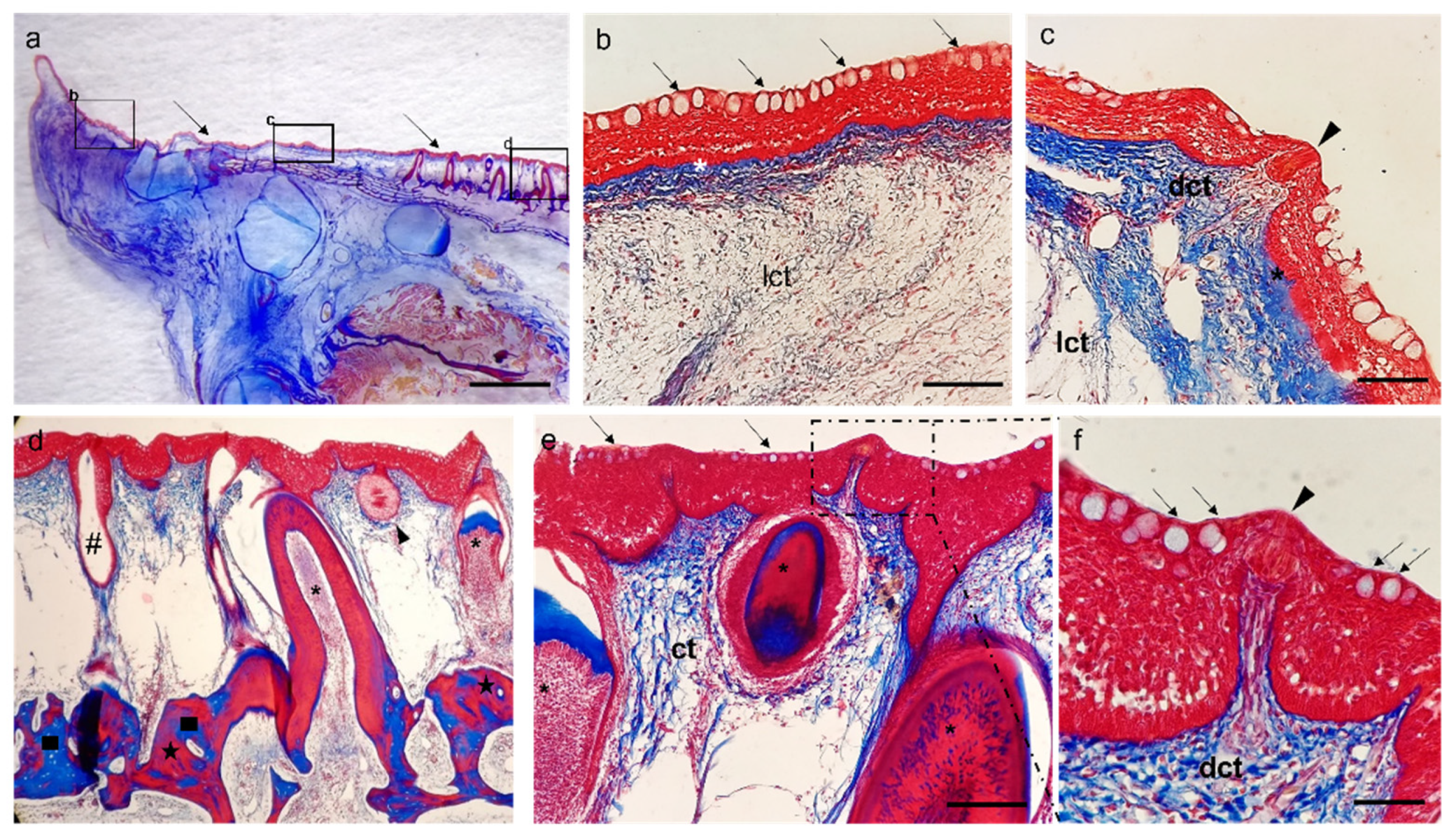
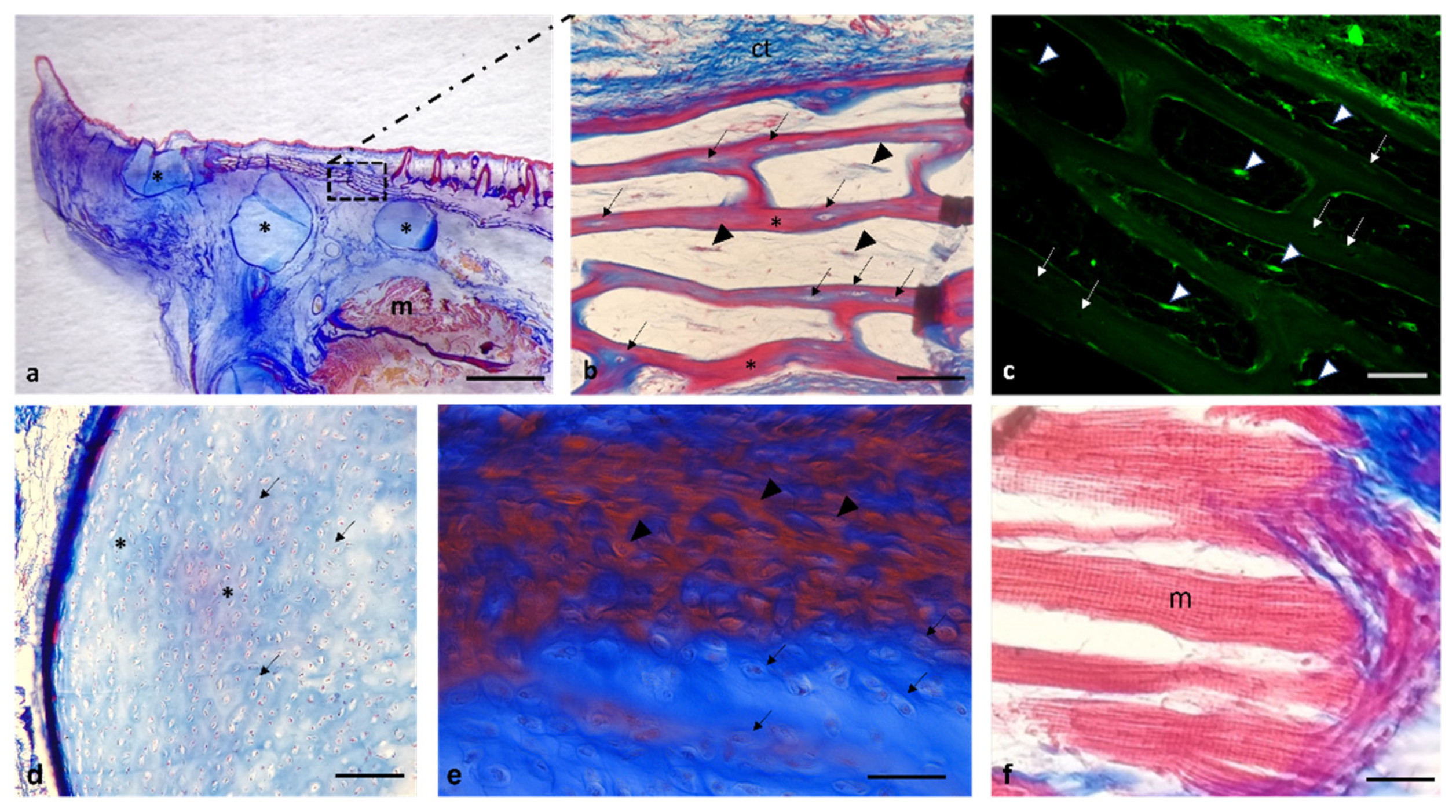
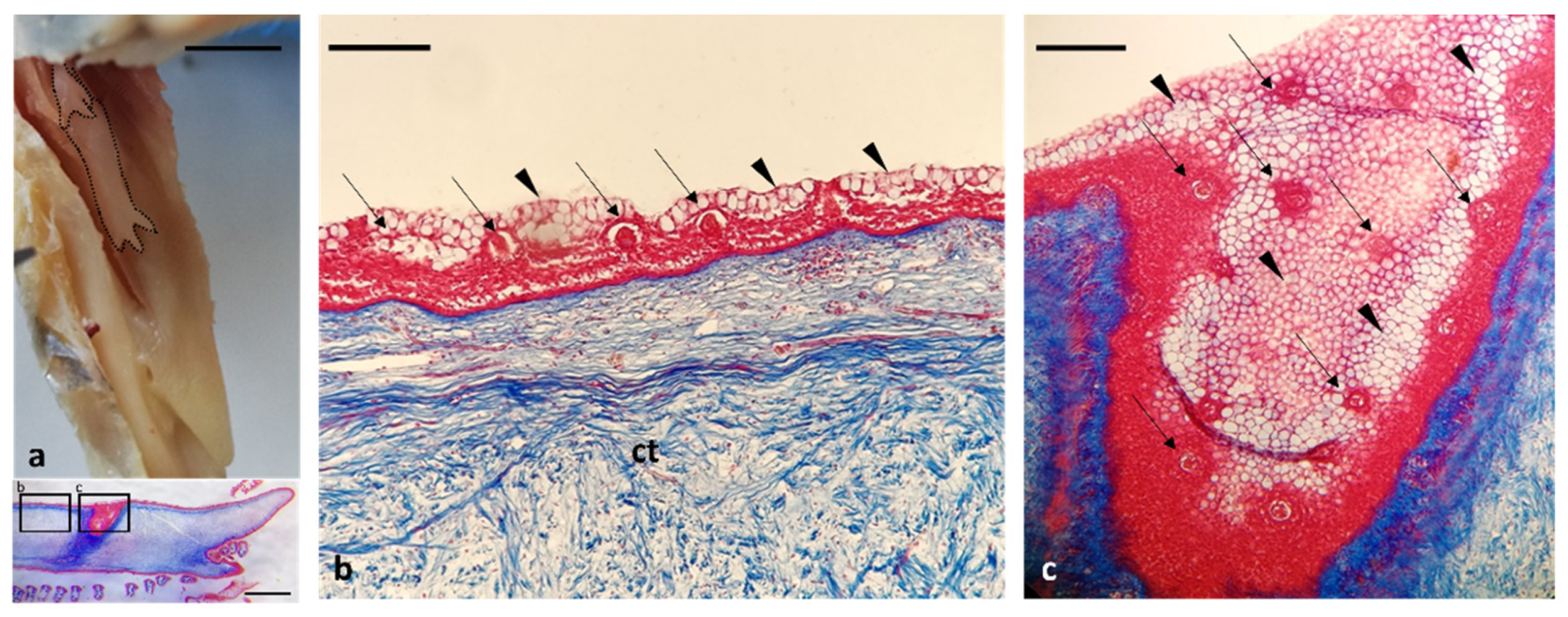
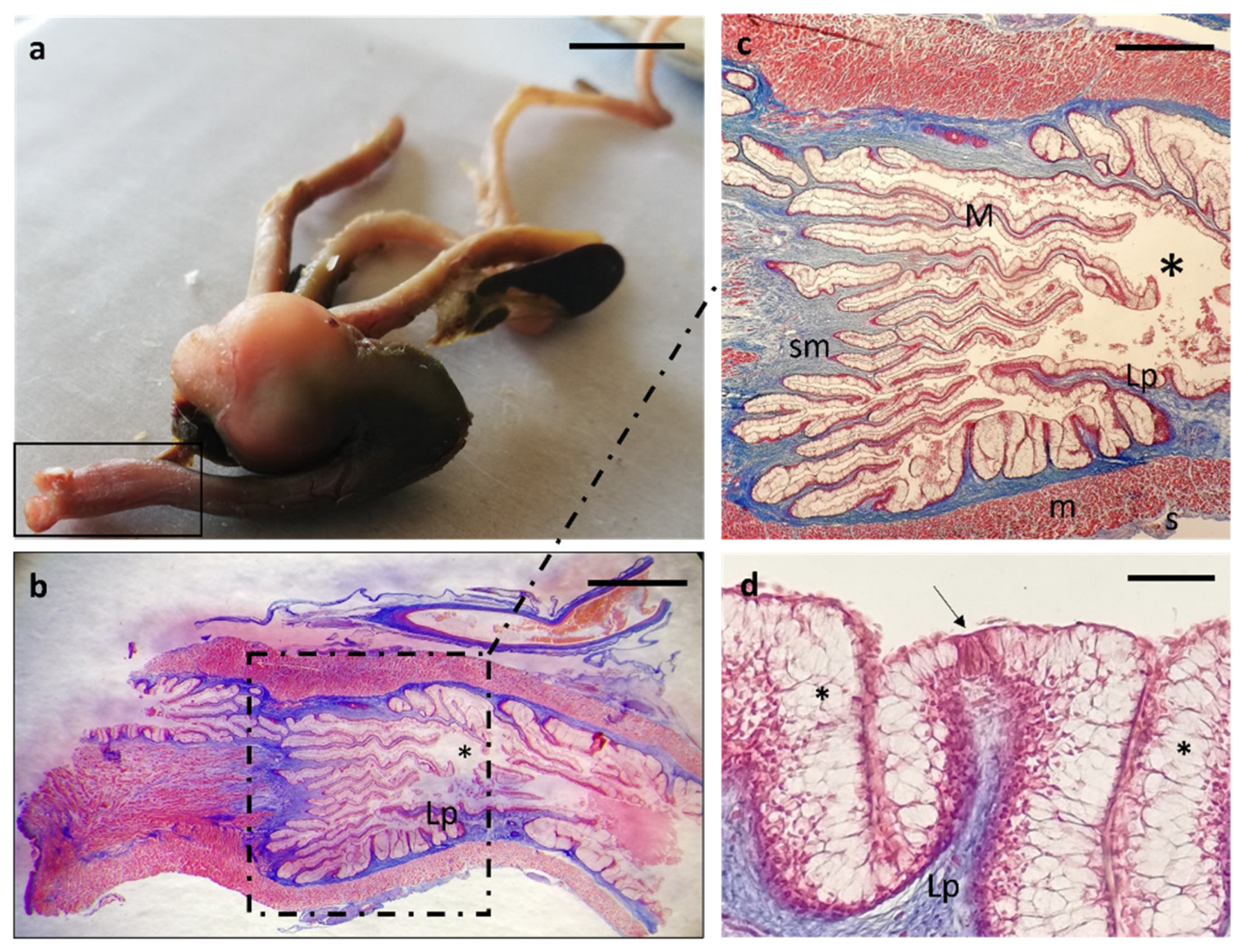
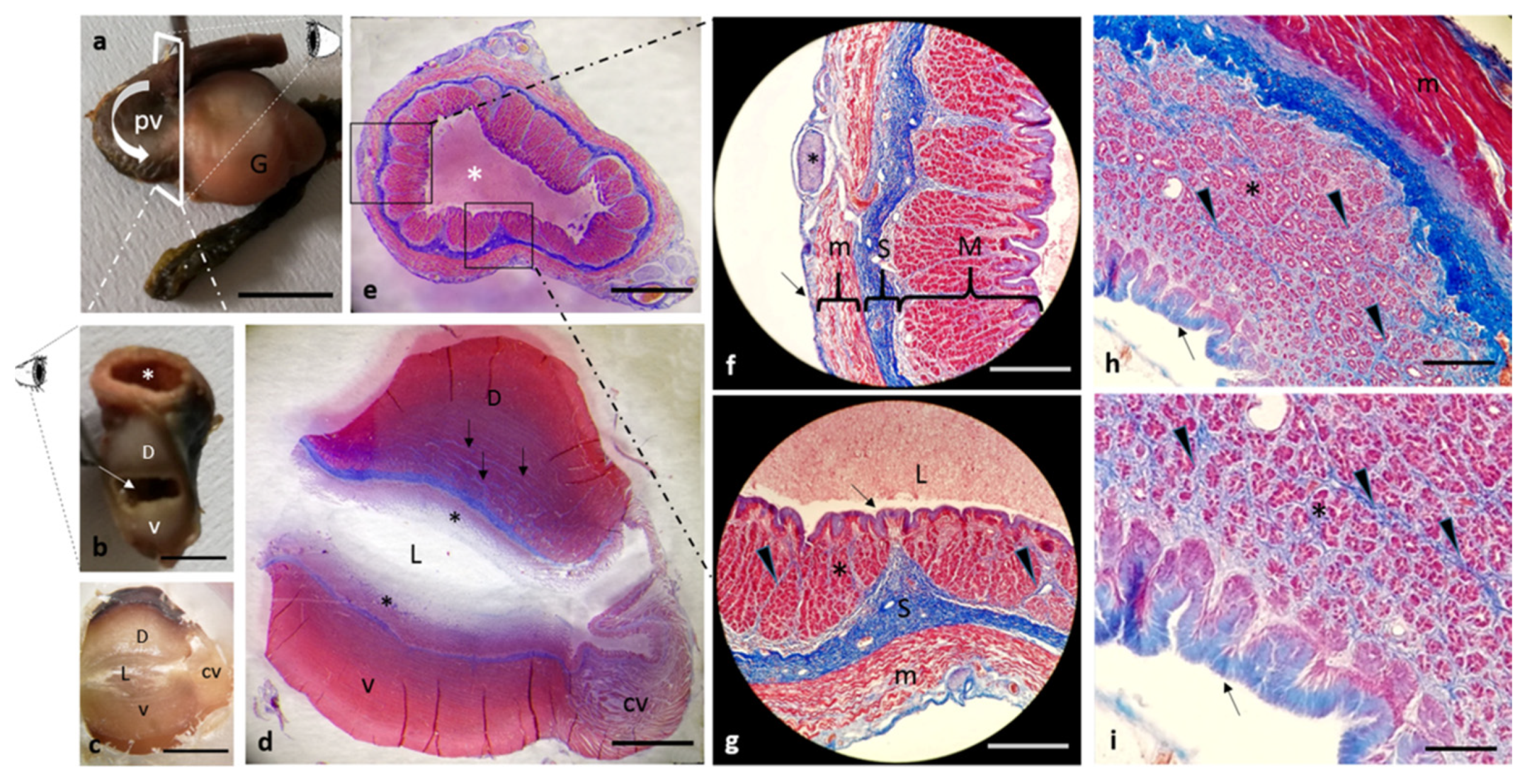
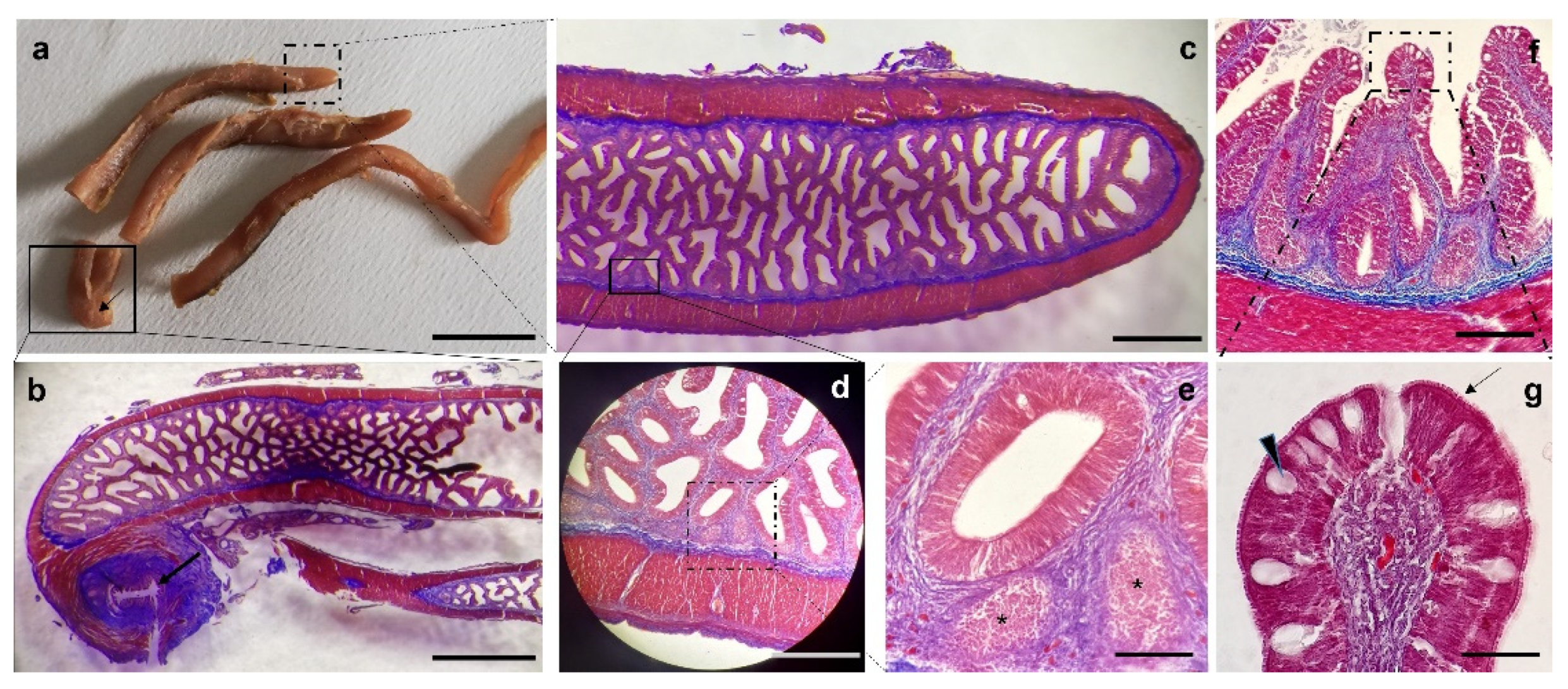
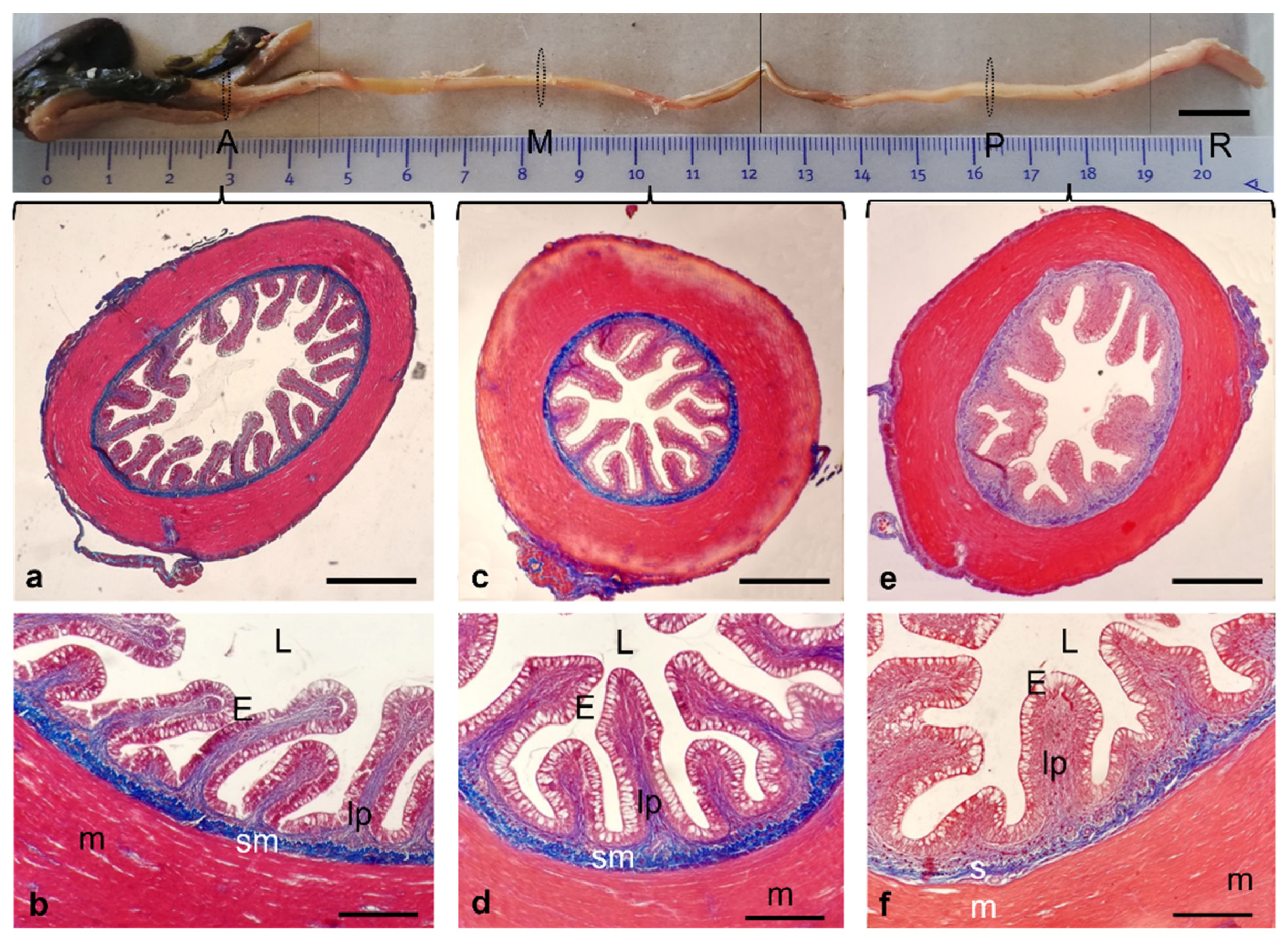
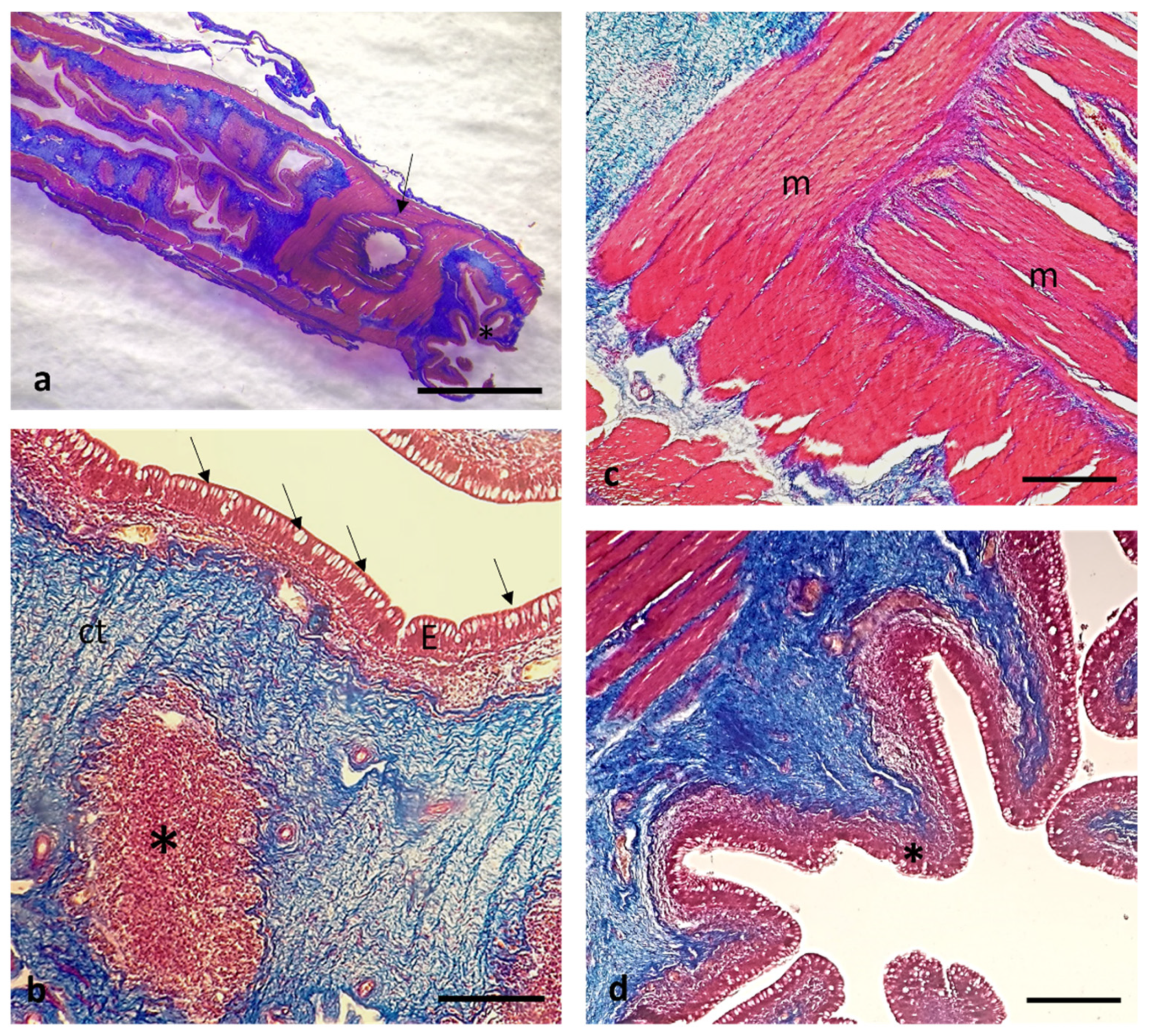
3. Results
Histological Features
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Mbega, J. Biodiversité des poissons du bassin inférieur de l’Ogooué (Gabon). Tropicultura 2004, 22. Available online: https://agris.fao.org/agris-search/search.do?recordID=DJ2012048960 (accessed on 25 March 2022).
- Adite, A.; Winemiller, K.O.; Fiogbe, E.D. Ontogenetic, seasonal, and spatial variation in the diet of Heterotis niloticus (Osteoglossiformes: Osteoglossidae) in the Sô River and Lake Hlan, Benin, West Africa. Environ. Biol. Fishes 2005, 73, 367–378. [Google Scholar] [CrossRef]
- Adite, A.; Winemiller, K.; Fiogbe, E. Population structure and reproduction of the African bonytongue Heterotis niloticus in the Sô River-floodplain system (West Africa): Implications for management. Ecol. Freshw. Fish 2006, 15, 30–39. [Google Scholar] [CrossRef]
- Froese, R.; Zeller, D.; Kleisner, K.; Pauly, D. What catch data can tell us about the status of global fisheries. Mar. Biol. 2012, 159, 1283–1292. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic PublicationFishBase. 2019. Available online: http://www.fishbase.org (accessed on 25 March 2022).
- Moreau, S.; Perly, B.; Biguet, J. Interactions de la chloroquine avec la ferriprotoporphyrine IX: Étude par résonance magnétique nucléaire. Biochimie 1982, 64, 1015–1025. [Google Scholar] [CrossRef]
- Odo, G.E.; Nwamba, H.O.; Eyo, J.E. Aspects of the biology of Heterotis niloticus Cuvier 1829 (osteoglossiformes: Osteoglossidae) in the anambra flood river system, Nigeria. Anim. Res. Int. 2009, 6, 2. [Google Scholar] [CrossRef]
- Koenig, L.A.; Gallant, J.R. Sperm competition, sexual selection and the diverse reproductive biology of Osteoglossiformes. J. Fish Biol. 2021, 99, 740–754. [Google Scholar] [CrossRef]
- Obermiller, L.E.; Pfeiler, E. Phylogenetic relationships of elopomorph fishes inferred from mitochondrial ribosomal DNA sequences. Mol. Phylogenetics Evol. 2003, 26, 202–214. [Google Scholar] [CrossRef]
- Dudgeon, D. The contribution of scientific information to the conservation and management of freshwater biodiversity in tropical Asia. In Aquatic Biodiversity; Springer: Berlin/Heidelberg, Germany, 2003; Volume 171, pp. 295–314. [Google Scholar]
- Mustapha, M. Heterotis niloticus (Cuvier, 1829) a threatened fish species in Oyun reservoir, Offa, Nigeria; the need for its conservation. Asian J. Exp. Biol. Sci. 2010, 1, 1–7. [Google Scholar]
- Bake, G.G.; Sadiku, S. Relationship between the basic morphometric measurements and growth pattern of Heterotis niloticus from River Kaduna floodplain. In Proceedings of the 19th Annual Conference of the Fisheries Society of Nigeria (FISON), Ilorin, Nigeria, 29 November–3 December 2004. [Google Scholar]
- Fagbenro, O.A.; Akinbulumo, M.O.; Adeparusi, O.E.; Raji, A.A. Flesh Yield, Waste Yield, Proximate and Mineral Composition of Four Commercial West African Freshwater Food Fishes. J. Anim. Vet. Adv. 2005, 4, 848–851. [Google Scholar]
- Goulding, M. The Fishes and the Forest; University of California Press: Berkeley, CA, USA, 1980. [Google Scholar]
- Rainboth, W.J.; Food and Agriculture Organization; Danish International Development Agency. Fishes of the Cambodian Me-kong; Food and Agriculture Organization, Ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 1996. [Google Scholar]
- Allen, G.R.; Midgley, S.H.; Allen, M. Field Guide to the Freshwater Fishes of Australia; Western Australian Museum: Perth, Australia, 2002.
- Hellberg, H.; Bjerkås, I. The Anatomy of the Oesophagus, Stomach and Intestine in Common Wolffish (Anarhichas lupus L.): A Basis for Diagnostic Work and Research. Acta Vet. Scand. 2000, 41, 283–297. [Google Scholar] [CrossRef] [PubMed]
- El-Bakary, N.; El-Gammal, H.L. Comparative histological, histochemical and ultrastructural studies on the liver of flathead grey mullet (Mugil cephalus) and sea bream (Sparus aurata). Glob. Vet. 2010, 4, 548–553. [Google Scholar]
- Abbate, F.; Latella, G.; Montalbano, G.; Guerrera, M.C.; Levanti, M.B.; Ciriaco, E. Scanning Electron Microscopical Study of the Lingual Epithelium of Green Iguana (Iguana iguana). Anat. Histol. Embryol. 2008, 37, 314–316. [Google Scholar] [CrossRef] [PubMed]
- Abbate, F.; Latella, G.; Montalbano, G.; Guerrera, M.C.; Germanà, G.P.; Levanti, M.B. The Lingual Dorsal Surface of the Blue-Tongue Skink (Tiliqua scincoides). Anat. Histol. Embryol. 2009, 38, 348–350. [Google Scholar] [CrossRef] [PubMed]
- Yashpal, M.; Kumari, U.; Mittal, S.; Mittal, A.K. Morphological specializations of the buccal cavity in relation to the food and feeding habit of a carp Cirrhinus mrigala: A scanning electron microscopic investigation. J. Morphol. 2009, 270, 714–728. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Levanti, M.; Laurà, R.; Germanà, G.P.; Montalbano, G.; Cavallaro, M.; Germanà, A. Morphology of the Atlantic salmon (Salmo salar) tongue. Anat. Histol. Embryol. 2020, 49, 686–694. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Levanti, M.; Laurà, R.; Aragona, M.; Mhalhel, K.; Montalbano, G.; Germanà, A. Anatomical, histological and immunohistochemical study of the tongue in the rainbow trout (Oncorhynchus mykiss). Anat. Histol. Embryol. 2020, 49, 848–858. [Google Scholar] [CrossRef]
- Nandi, S.; Saikia, S.K. Scanning electron microscopic and histological studies of the buccal cavity of a phytoplanktivorous small freshwater fish, Amblypharyngodon mola. Microsc. Res. Tech. 2021, 84, 119–124. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Levanti, M.; Laurà, R.; Aragona, M.; Mhalhel, K.; Montalbano, G.; Germanà, A. Morphological characteristics of the blackspot seabream (Pagellu.us bogaraveo) tongue: A structural and immunohistochemical study. Anat. Histol. Embryol. 2022, 51, 103–111. [Google Scholar] [CrossRef]
- Abbate, F.; Germana, G.; De Carlos, F.; Montalbano, G.; Laura, R.; Levanti, M.; Germana, A. The oral cavity of the adult zebrafish (Danio rerio). Anat. Histol. Embryol. 2006, 35, 299–304. [Google Scholar] [CrossRef]
- El Bakary, N.E.S.R. Morphological study of the asymmetrical buccal cavity of the flatfish common solea (Solea solea) and its relation to the type of feeding. Asian Pac. J. Trop. Biomed. 2014, 4, 13–17. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Montalbano, G.; De Carlos, F.; Suárez, A.Á.; Ciriaco, E.; Germanà, A. Morphology of the european sea bass (Dicentrarchus labrax) tongue. Microsc. Res. Tech. 2012, 75, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Abbate, F.; Guerrera, M.C.; Montalbano, G.; Ciriaco, E.; Germanà, A. Morphology of the tongue dorsal surface of gilthead seabream (Sparus aurata). Microsc. Res. Tech. 2012, 75, 1666–1671. [Google Scholar] [CrossRef] [PubMed]
- Gamal, A.M.; Elsheikh, E.H.; Nasr, E.S. Morphological adaptation of the buccal cavity in relation to feeding habits of the omnivorous fish Clarias gariepinus: A scanning electron microscopic study. J. Basic Appl. Zool. 2012, 65, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Kasumyan, A.O. The taste system in fishes and the effects of environmental variables. J. Fish Biol. 2019, 95, 155–178. [Google Scholar] [CrossRef] [Green Version]
- Abbate, F.; Guerrera, M.C.; Levanti, M.; Laurà, R.; Montalbano, G.; Cavallaro, M.; Germanà, A. The tongue of Leopard Gecko (Eublepharis macularius): LM, SEM and confocal laser study. Anat. Histol. Embryol. 2020, 49, 51–59. [Google Scholar] [CrossRef]
- Abuzinadah, O.A. Studies on Red Sea Fish; Swansea University (United Kingdom): Swansea, UK, 1990. [Google Scholar]
- Nasruddin, N.S.; Azmai, M.N.A.; Ismail, A.; Saad, M.Z.; Daud, H.M.; Zulkifli, S.Z. Histological Features of the Gastrointestinal Tract of Wild Indonesian Shortfin Eel, Anguilla bicolor bicolor (McClelland, 1844), Captured in Peninsular Malaysia. Sci. World J. 2014, 2014, 312670. [Google Scholar] [CrossRef] [Green Version]
- Naguib, S.; Ei-Shabaka, H.; Ashour, F. Comparative histological and ultrastructural studies on the stomach of Schilbe mystus and the intestinal swelling of Labeo niloticus. J. Am. Sci. 2011, 7, 251–263. [Google Scholar]
- Day, R.D.; German, D.P.; Manjakasy, J.M.; Farr, I.; Hansen, M.J.; Tibbetts, I.R. Enzymatic digestion in stomachless fishes: How a simple gut accommodates both herbivory and carnivory. J. Comp. Physiol. B 2011, 181, 603–613. [Google Scholar] [CrossRef]
- Agbugui, M.O.; Egbo, H.O.; Abhulimen, F.E. The Biology of the African Bonytongue Heterotis niloticus (Cuvier, 1829) from the Lower Niger River at Agenebode in Edo State, Nigeria. Int. J. Zool. 2021, 2021, 1748736. [Google Scholar] [CrossRef]
- Buddington, R.K.; Chen, J.W.; Diamond, J. Genetic and phenotypic adaptation of intestinal nutrient transport to diet in fish. J. Physiol. 1987, 393, 261–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbate, F.; Guerrera, M.C.; Montalbano, G.; Zichichi, R.; Germanà, A.; Ciriaco, E. Morphology of the Lingual Dorsal Surface and Oral Taste Buds in Italian Lizard (Podarcis sicula). Anat. Histol. Embryol. 2010, 39, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Erdoğan, S.; Iwasaki, S.-i. Function-related morphological characteristics and specialized structures of the avian tongue. Ann. Anat.–Anat. Anz. 2014, 196, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Erdogan, S.; Alan, A. Gross anatomical and scanning electron microscopic studies of the oropharyngeal cavity in the European magpie (Pica pica) and the common raven (Corvus corax). Microsc. Res. Tech. 2012, 75, 379–387. [Google Scholar] [CrossRef]
- Herrel, A.; van der Meijden, A. An analysis of the live reptile and amphibian trade in the USA compared to the global trade in endangered species. Herpetol. J. 2014, 24, 103–110. [Google Scholar]
- Cizek, P.; Hamouzova, P.; Kvapil, P.; Kyllar, M. Light and scanning electron microscopy of the tongue of the sand lizard (Lacerta agilis). Folia Morphol. 2019, 78, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Bels, V.L.; Jamniczky, H.A.; Montuelle, S.; Pallandre, J.P.; Kardong, K.V.; Russell, A.P. Mechanics and kinematics of fluid uptake and intraoral transport in the leopard gecko. J. Zool. 2020, 311, 33–44. [Google Scholar] [CrossRef]
- Monentcham, S.-E.; Kouam, J.; Pouomogne, V.; Kestemont, P. Biology and prospect for aquaculture of African bonytongue, Heterotis niloticus (Cuvier, 1829): A review. Aquaculture 2009, 289, 191–198. [Google Scholar] [CrossRef]
- Agbugui, M.O.; Abhulimen, F.E.; Egbo, H.O. Gross Anatomy and Histological Features of Gymnarchus niloticus (Cuvier, 1829) from the River Niger at Agenebode in Edo State, Nigeria. Int. J. Zool. 2021, 2021, 3151609. [Google Scholar] [CrossRef]
- Madkour, F.; Mohamed, A. Macro-microscopical anatomy of the gizzard of egyptian laughing dove and rock pigeon. Assiut Vet. Med. J. 2019, 65, 278–285. [Google Scholar]
- Domeneghini, C.; Arrighi, S.; Radaelli, G.; Bosi, G.; Mascarello, F. Morphological and histochemical peculiarities of the gut in the white sturgeon, Acipenser transmontanus. Eur. J. Histochem. 1999, 43, 135–145. [Google Scholar] [PubMed]
- Ballesteros, N.A.; Castro, R.; Abos, B.; Rodríguez Saint-Jean, S.S.; Pérez-Prieto, S.I.; Tafalla, C. The pyloric caeca area is a major site for IgM+ and IgT+ B cell recruitment in response to oral vaccination in rainbow trout. PLoS ONE 2013, 8, e66118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casteleyn, C.; Doom, M.; Lambrechts, E.; Van Den Broeck, W.; Simoens, P.; Cornillie, P. Locations of gut-associated lymphoid tissue in the 3-month-old chicken: A review. Avian Pathol. 2010, 39, 143–150. [Google Scholar] [CrossRef]
- Su, J.; Gong, Y.; Mei, L.; Xi, L.; Chi, S.; Yang, Y.; Jin, J.; Liu, H.; Zhu, X.; Xie, S.; et al. The characteristics of glucose homoeostasis in grass carp and Chinese longsnout catfish after oral starch administration: A comparative study between herbivorous and carnivorous species of fish. Br. J. Nutr. 2020, 123, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Levina, A.D.; Mikhailova, E.S.; Kasumyan, A.O. Taste preferences and feeding behaviour in the facultative herbivorous fish, Nile tilapia Oreochromis niloticus. J. Fish Biol. 2021, 98, 1385–1400. [Google Scholar] [CrossRef]
- Salamat, N.; Darbeh, R.; Majd, N.E.; Safahieh, A.R. Histological aspects of gut associated lymphoid tissue in Acanthopagrus latus. Glob. Vet. 2011, 7, 381–385. [Google Scholar]
- Scadeng, M.; McKenzie, C.; He, W.; Bartsch, H.; Dubowitz, D.J.; Stec, D.; St. Leger, J. Morphology of the Amazonian Teleost Genus Arapaima Using Advanced 3D Imaging. Front. Physiol. 2020, 11, 260. [Google Scholar] [CrossRef]
- Hopperdietzel, C.; Hirschberg, R.M.; Hünigen, H.; Wolter, J.; Richardson, K.; Plendl, J. Gross morphology and histology of the alimentary tract of the convict cichlid Amatitlania nigrofasciata. J. Fish Biol. 2014, 85, 1707–1725. [Google Scholar] [CrossRef]
- Zizza, S.; Desantis, S. Morphology and lectin-binding sites of pyloric caeca epithelium in normal and GnRH-treated Atlantic bluefin tuna (Thunnus thynnus) Linnaeus 1758. Microsc. Res. Tech. 2011, 74, 863–873. [Google Scholar] [CrossRef]
- Urbas, R.; Klieser, E.; Neureiter, D.; Brenner, E. Anatomy and Embryology of the Pancreatic Gland. In Textbook of Pancreatic Cancer: Principles and Practice of Surgical Oncology; Søreide, K., Stättner, S., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2021; pp. 145–160. [Google Scholar] [CrossRef]
- Youson, J.H.; Al-Mahrouki, A.A.; Amemiya, Y.; Graham, L.C.; Montpetit, C.J.; Irwin, D.M. The fish endocrine pancreas: Review, new data, and future research directions in ontogeny and phylogeny. Gen. Comp. Endocrinol. 2006, 148, 105–115. [Google Scholar] [CrossRef]
- Caruso, M.; Sheridan, M. Gut anatomy and morphology|Pancreas. In Encyclopedia of Fish Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2011; Volume 2, pp. 1276–1283. [Google Scholar]
- Caruso, M.A.; Sheridan, M.A. New insights into the signaling system and function of insulin in fish. Gen. Comp. Endocrinol. 2011, 173, 227–247. [Google Scholar] [CrossRef] [PubMed]
- Herrera, M.J.; Heras, J.; German, D.P. Comparative transcriptomics reveal tissue level specialization towards diet in prickleback fishes. J. Comp. Physiol. B 2022, 192, 275–295. [Google Scholar] [CrossRef] [PubMed]
- Farag, F.; Wally, Y.; Daghash, S.; Ibrahim, A.J.J.o.v.a. Some gross morphological studies on the internal anatomy of the scaled common carp fish (Cyprinus carpio) in Egypt. J. Vet. Anat. 2014, 7, 15–29. [Google Scholar] [CrossRef]
- Alonso, F.; Mirande, J.M.; Pandolfi, M. Gross anatomy and histology of the alimentary system of Characidae (Teleostei: Ostariophysi: Characiformes) and potential phylogenetic information. Neotrop. Ichthyol. 2015, 13, 273–286. [Google Scholar] [CrossRef]
- Danowitz, M.; Solounias, N. Embryology, comparative anatomy, and congenital malformations of the gastrointestinal tract. Edorium J. Anat. Embryol. 2016, 3, 39–50. [Google Scholar] [CrossRef]
- Youson, J.H.; Al-Mahrouki, A.A. Ontogenetic and Phylogenetic Development of the Endocrine Pancreas (Islet Organ) in Fishes. Gen. Comp. Endocrinol. 1999, 116, 303–335. [Google Scholar] [CrossRef] [Green Version]
- Dutta, H.M.; Datta-Mushi, J.S. Fish Morphology; Taylor & Francis: Abingdon-on-Thames, UK, 1996. [Google Scholar]
- Faccioli, C.K.; Chedid, R.A.; Bombonato, M.T.S.; Vicentini, C.A.; Vicentini, I.B.F. Morphology and histochemistry of the liver of carnivorous fish Hemisorubim platyrhynchos. Int. J. Morphol. 2014, 32, 715–720. [Google Scholar] [CrossRef] [Green Version]
- Agius, C.; Roberts, R.J. Melano-macrophage centres and their role in fish pathology. J. Fish Dis. 2003, 26, 499–509. [Google Scholar] [CrossRef]
- Steinel, N.C.; Bolnick, D.I. Melanomacrophage Centers As a Histological Indicator of Immune Function in Fish and Other Poikilotherms. Front. Immunol. 2017, 8, 827. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, H.J.; Procópio, M.S.; Gomes, J.M.M.; Vieira, F.O.; Russo, R.C.; Balzuweit, K.; Chiarini-Garcia, H.; Santana Castro, A.C.; Rizzo, E.; Corrêa, J.D. Functional dissimilarity of melanomacrophage centres in the liver and spleen from females of the teleost fish Prochilodus argenteus. Cell Tissue Res. 2011, 346, 417–425. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerrera, M.C.; Aragona, M.; Briglia, M.; Porcino, C.; Mhalhel, K.; Cometa, M.; Abbate, F.; Montalbano, G.; Laurà, R.; Levanti, M.; et al. The Alimentary Tract of African Bony-Tongue, Heterotis niloticus (Cuvier, 1829): Morphology Study. Animals 2022, 12, 1565. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121565
Guerrera MC, Aragona M, Briglia M, Porcino C, Mhalhel K, Cometa M, Abbate F, Montalbano G, Laurà R, Levanti M, et al. The Alimentary Tract of African Bony-Tongue, Heterotis niloticus (Cuvier, 1829): Morphology Study. Animals. 2022; 12(12):1565. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121565
Chicago/Turabian StyleGuerrera, Maria Cristina, Marialuisa Aragona, Marilena Briglia, Caterina Porcino, Kamel Mhalhel, Marzio Cometa, Francesco Abbate, Giuseppe Montalbano, Rosaria Laurà, Maria Levanti, and et al. 2022. "The Alimentary Tract of African Bony-Tongue, Heterotis niloticus (Cuvier, 1829): Morphology Study" Animals 12, no. 12: 1565. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121565