
Molecular Characterization of Anaplasma spp. among Dairy, Cashmere, and Meat Goats in Shaanxi Province, Northwestern China

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
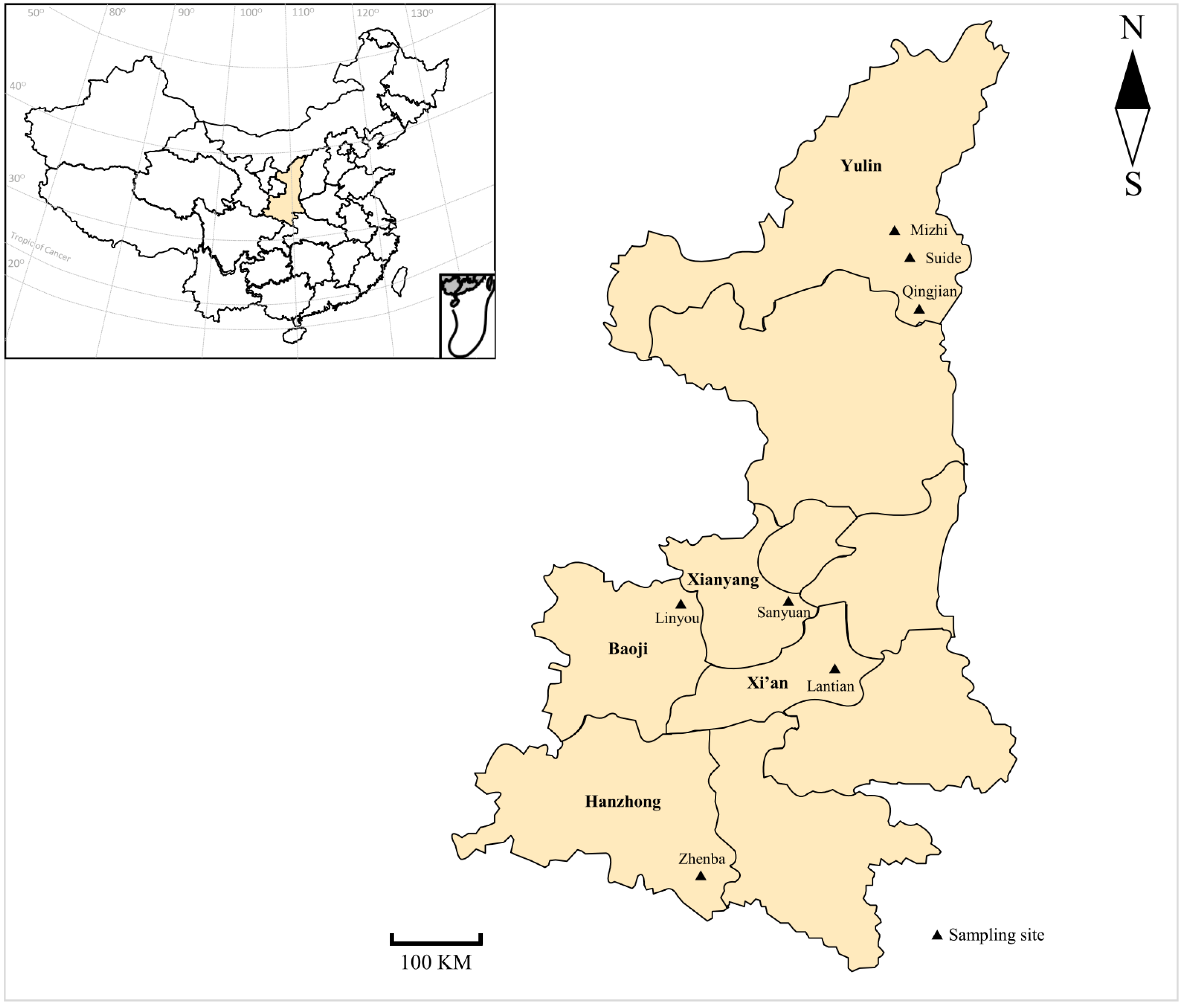
2.1. Sampling
2.2. Genomic DNA Extraction
2.3. PCR Amplification
2.4. Sequencing and Sequence Analysis
2.5. Statistical Analysis
2.6. Nucleotide Sequence Accession Numbers
3. Results
3.1. Prevalence of Anaplasma in Goats in Shaanxi Province
3.2. Anaplasma phagocytophilum in Goats in Shaanxi Province
3.3. Anaplasma bovis in Goats in Shaanxi Province
3.4. Anaplasma ovis in Goats in Shaanxi Province
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dumler, J.S.; Barbet, A.F.; Bekker, C.P.; Dasch, G.A.; Palmer, G.H.; Ray, S.C.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: Unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and ‘HGE agent’ as subjective synonyms of Ehrlichia phagocytophila. Int. J. Syst. Evol. Microbiol. 2001, 51, 2145–2165. [Google Scholar] [PubMed] [Green Version]
- Kocan, K.M.; de la Fuente, J.; Guglielmone, A.A.; Meléndez, R.D. Antigens and alternatives for control of Anaplasma marginale infection in cattle. Clin. Microbiol. Rev. 2003, 16, 698–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madison-Antenucci, S.; Kramer, L.D.; Gebhardt, L.L.; Kauffman, E. Emerging Tick-Borne Diseases. Clin. Microbiol. Rev. 2020, 33, e00083-18. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zheng, Y.C.; Ma, L.; Jia, N.; Jiang, B.G.; Jiang, R.R.; Huo, Q.B.; Wang, Y.W.; Liu, H.B.; Chu, Y.L.; et al. Human infection with a novel tick-borne Anaplasma species in China: A surveillance study. Lancet. Infect. Dis. 2015, 15, 663–670. [Google Scholar] [CrossRef]
- Kocan, K.M.; de la Fuente, J.; Cabezas-Cruz, A. The genus Anaplasma: New challenges after reclassification. Rev. Sci. Tech. 2015, 34, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Matei, I.A.; Estrada-Peña, A.; Cutler, S.J.; Vayssier-Taussat, M.; Varela-Castro, L.; Potkonjak, A.; Zeller, H.; Mihalca, A.D. A review on the eco-epidemiology and clinical management of human granulocytic anaplasmosis and its agent in Europe. Parasit. Vectors 2019, 12, 599. [Google Scholar] [CrossRef]
- Woldehiwet, Z. The natural history of Anaplasma phagocytophilum. Vet. Parasitol. 2010, 167, 108–122. [Google Scholar] [CrossRef]
- Ben Said, M.; Belkahia, H.; Karaoud, M.; Bousrih, M.; Yahiaoui, M.; Daaloul-Jedidi, M.; Messadi, L. First molecular survey of Anaplasma bovis in small ruminants from Tunisia. Vet. Microbiol. 2015, 179, 322–326. [Google Scholar] [CrossRef]
- Battilani, M.; De Arcangeli, S.; Balboni, A.; Dondi, F. Genetic diversity and molecular epidemiology of Anaplasma. Infect. Genet. Evol. 2017, 49, 195–211. [Google Scholar] [CrossRef]
- Hornok, S.; Elek, V.; de la Fuente, J.; Naranjo, V.; Farkas, R.; Majoros, G.; Földvári, G. First serological and molecular evidence on the endemicity of Anaplasma ovis and A. marginale in Hungary. Vet. Microbiol. 2007, 122, 316–322. [Google Scholar] [CrossRef]
- Bakken, J.S.; Dumler, J.S. Human granulocytic anaplasmosis. Infect. Dis. Clin. 2015, 29, 341–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, A.S.; Bakken, J.S.; Folk, S.M.; Paddock, C.D.; Bloch, K.C.; Krusell, A.; Sexton, D.J.; Buckingham, S.C.; Marshall, G.S.; Storch, A.G.; et al. Diagnosis and management of tickborne rickettsial diseases: Rocky Mountain spotted fever, ehrlichioses, and anaplasmosis—United States: A practical guide for physicians and other health-care and public health professionals. MMWR Recomm. Rep. 2006, 55, 1–27. [Google Scholar] [PubMed]
- Wormser, G.P.; Dattwyler, R.J.; Shapiro, E.D.; Halperin, J.J.; Steere, A.C.; Klempner, M.S.; Krause, P.J.; Bakken, J.S.; Strle, F.; Stanek, G.; et al. The clinical assessment, treatment, and prevention of lyme disease, human granulocytic anaplasmosis, and babesiosis: Clinical practice guidelines by the Infectious Diseases Society of America. Clin. Infect. Dis. 2006, 43, 1089–1134. [Google Scholar] [CrossRef] [PubMed]
- WOAH. Manual of Standards for Diagnostic Tests and Vaccines for Terrestrial Animals. 2008. Available online: https://www.oie.int/en/what-we-do/standards/codes-and-manuals/ (accessed on 20 July 2021).
- Kolo, A.O.; Collins, N.E.; Brayton, K.A.; Chaisi, M.; Blumberg, L.; Frean, J.; Gall, C.A.; Wentzel, M.J.; Wills-Berriman, S.; Boni, L.D.; et al. Anaplasma phagocytophilum and other Anaplasma spp. in various hosts in the Mnisi Community, Mpumalanga Province, South Africa. Microorganisms 2020, 8, 1812. [Google Scholar] [CrossRef]
- Hosseini-Chegeni, A.; Tavakoli, M.; Goudarzi, G.; Telmadarraiy, Z.; Sharifdini, M.; Faghihi, F.; Ghanbari, M.K. Molecular detection of Anaplasma marginale and Anaplasma ovis (Rickettsiales: Anaplasmataceae) in Ixodid Tick Species in Iran. Arch. Razi. Inst. 2020, 75, 39–46. [Google Scholar]
- Sun, X.F.; Zhao, L.; Wen, H.L.; Luo, L.M.; Yu, X.J. Anaplasma species in China. Lancet Infect. Dis. 2015, 15, 1263–1264. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Ma, M.; Wang, Z.; Wang, J.; Peng, Y.; Li, Y.; Guan, G.; Luo, J.; Yin, H. Molecular survey and genetic identification of Anaplasma species in goats from central and southern China. Appl. Environ. Microbiol. 2012, 78, 464–470. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Li, Y.; Liu, Z.; Liu, J.; Niu, Q.; Ren, Q.; Chen, Z.; Guan, G.; Luo, J.; Yin, H. Molecular detection and characterization of Anaplasma spp. in sheep and cattle from Xinjiang, northwest China. Parasit. Vectors 2015, 8, 108. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Liu, Z.; Niu, Q.; Liu, J.; Han, R.; Guan, G.; Li, Y.; Liu, G.; Luo, J.; Yin, H. Anaplasma phagocytophilum in sheep and goats in central and southeastern China. Parasit. Vectors 2016, 9, 593. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Yang, J.; Guan, G.; Liu, Z.; Luo, J.; Song, M. Molecular investigation of Anaplasma species in sheep from Heilongjiang Province, northeast China identified four Anaplasma species and a novel genotype of Anaplasma capra. Parasitol. Int. 2020, 76, 102072. [Google Scholar] [CrossRef]
- Wang, K.; Yan, Y.; Zhou, Y.; Zhao, S.; Jian, F.; Wang, R.; Zhang, L.; Ning, C. Seasonal dynamics of Anaplasma spp. in goats in warm-temperate zone of China. Ticks Tick-Borne Dis. 2021, 12, 101673. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Sun, E.; Wen, Y.; Ye, C.; Liu, F.; Jiang, P.; Tao, X. Molecular evidence of coinfection of Anaplasma species in small ruminants from Anhui Province, China. Parasitol. Int. 2019, 71, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Zhan, L.; Cao, W.C.; Jiang, J.F.; Zhang, X.A.; Wu, X.M.; Zhang, W.Y.; Liu, W.; Zuo, S.Q.; Cao, Z.W.; Yang, H.; et al. Anaplasma phagocytophilum in livestock and small rodents. Vet. Microbiol. 2010, 144, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, M.; Rikihisa, Y.; Lin, Q.; Isogai, E.; Tahara, K.; Itagaki, A.; Hiramitsu, Y.; Tajima, T. Novel genetic variants of Anaplasma phagocytophilum, Anaplasma bovis, Anaplasma centrale, and a novel Ehrlichia sp. in wild deer and ticks on two major islands in Japan. Appl. Environ. Microbiol. 2006, 72, 1102–1109. [Google Scholar] [CrossRef] [Green Version]
- Barlough, J.E.; Madigan, J.E.; DeRock, E.; Bigornia, L. Nested polymerase chain reaction for detection of Ehrlichia equi genomic DNA in horses and ticks (Ixodes pacificus). Vet. Parasitol. 1996, 63, 319–329. [Google Scholar] [CrossRef]
- de la Fuente, J.; Atkinson, M.W.; Naranjo, V.; Fernández de Mera, I.G.; Mangold, A.J.; Keating, K.A.; Kocan, K.M. Sequence analysis of the msp4 gene of Anaplasma ovis strains. Vet. Microbiol. 2007, 119, 375–381. [Google Scholar] [CrossRef]
- Akyar, I. GLP: Good Laboratory Practice. In Modern Approaches to Quality Control; Eldin, A.B., Ed.; IntechOpen Limited: London, UK, 2011; pp. 35–56. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Shi, K.; Li, J.; Yan, Y.; Chen, Q.; Wang, K.; Zhou, Y.; Li, D.; Chen, Y.; Yu, F.; Peng, Y.; et al. Dogs as New Hosts for the emerging zoonotic pathogen Anaplasma capra in China. Front. Cell. Infect. Microbiol. 2019, 9, 394. [Google Scholar] [CrossRef]
- Guimarães, A.; Raimundo, J.M.; Peixoto, M.P.; da Silva, C.B.; Pires, M.S.; Santos, H.A.; Baldani, C.D. Molecular detection, characterization of Anaplasma spp. in domestic cats from Rio de Janeiro state. Acta Trop. 2019, 191, 239–242. [Google Scholar] [CrossRef]
- Félix, M.L.; Armúa-Fernández, M.T.; Parodi, P.; Bazzano, V.; Mangold, A.J.; Venzal, J.M. Detection of a putative novel genotype of Anaplasma in gray-brocket deer (Mazama gouazoubira) from Uruguay. Exp. Appl. Acarol. 2020, 81, 575–583. [Google Scholar] [CrossRef]
- Seo, M.G.; Kwon, O.D.; Kwak, D. Anaplasma bovis infection in a horse: First clinical report and molecular analysis. Vet. Microbiol. 2019, 233, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.M.; Dumler, J.S.; Bakken, J.S.; Walker, D.H. Identification of a granulocytotropic Ehrlichia species as the etiologic agent of human disease. J. Clin. Microbiol. 1994, 32, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poitout, F.M.; Shinozaki, J.K.; Stockwell, P.J.; Holland, C.J.; Shukla, S.K. Genetic variants of Anaplasma phagocytophilum infecting dogs in Western Washington State. J. Clin. Microbiol. 2005, 43, 796–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zobba, R.; Ben Said, M.; Belkahia, H.; Pittau, M.; Cacciotto, C.; Pinna Parpaglia, M.L.; Messadi, L.; Alberti, A. Molecular epidemiology of Anaplasma spp. related to A. phagocytophilum in Mediterranean small ruminants. Acta Trop. 2020, 202, 105286. [Google Scholar] [CrossRef]
- Ebani, V.; Cerri, D.; Fratini, F.; Ampola, M.; Andreani, E. Seroprevalence of Anaplasma phagocytophilum in domestic and wild animals from central Italy. New Microbiol. 2008, 31, 371–375. [Google Scholar]
- Cicculli, V.; Capai, L.; Quilichini, Y.; Masse, S.; Fernández-Alvarez, A.; Minodier, L.; Bompard, P.; Charrel, R.; Falchi, A. Molecular investigation of tick-borne pathogens in ixodid ticks infesting domestic animals (cattle and sheep) and small rodents (black rats) of Corsica, France. Ticks Tick-Borne Dis. 2019, 10, 606–613. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Z.; Guan, G.; Liu, Q.; Li, Y.; Chen, Z.; Ma, M.; Liu, A.; Ren, Q.; Luo, J.; et al. Prevalence of Anaplasma phagocytophilum in ruminants, rodents and ticks in Gansu, north-western China. J. Med. Microbiol. 2013, 62, 254–258. [Google Scholar] [CrossRef] [Green Version]
- Belkahia, H.; Said, M.B.; El Hamdi, S.; Yahiaoui, M.; Gharbi, M.; Daaloul-Jedidi, M.; Mhadhbi, M.; Jedidi, M.; Darghouth, M.A.; Klabi, I.; et al. First molecular identification and genetic characterization of Anaplasma ovis in sheep from Tunisia. Small Rumin. Res. 2014, 121, 404–410. [Google Scholar] [CrossRef]
- Torina, A.; Alongi, A.; Naranjo, V.; Estrada-Peña, A.; Vicente, J.; Scimeca, S.; Marino, A.M.; Salina, F.; Caracappa, S.; de la Fuente, J. Prevalence and genotypes of Anaplasma species and habitat suitability for ticks in a Mediterranean ecosystem. Appl. Environ. Microbiol. 2008, 74, 7578–7584. [Google Scholar] [CrossRef] [Green Version]
- Jahfari, S.; Coipan, E.C.; Fonville, M.; van Leeuwen, A.D.; Hengeveld, P.; Heylen, D.; Heyman, P.; van Maanen, C.; Butler, C.M.; Földvári, G.; et al. Circulation of four Anaplasma phagocytophilum ecotypes in Europe. Parasit. Vectors 2014, 7, 365. [Google Scholar] [CrossRef] [Green Version]
- Foley, J.E.; Nieto, N.C.; Massung, R.; Barbet, A.; Madigan, J.; Brown, R.N. Distinct ecologically relevant strains of Anaplasma phagocytophilum. Emerg. Infect. Dis. 2009, 15, 842–843. [Google Scholar] [CrossRef] [PubMed]
- Massung, R.F.; Mather, T.N.; Levin, M.L. Reservoir competency of goats for the Ap-variant 1 strain of Anaplasma phagocytophilum. Infect. Immun. 2006, 74, 1373–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugat, T.; Lagrée, A.C.; Maillard, R.; Boulouis, H.J.; Haddad, N. Opening the black box of Anaplasma phagocytophilum diversity: Current situation and future perspectives. Front. Cell. Infect. Microbiol. 2015, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Langenwalder, D.B.; Schmidt, S.; Silaghi, C.; Skuballa, J.; Pantchev, N.; Matei, I.A.; Mihalca, A.D.; Gilli, U.; Zajkowska, J.; Ganter, M.; et al. The absence of the drhm gene is not a marker for human-pathogenicity in European Anaplasma phagocytophilum strains. Parasit. Vectors 2020, 13, 238. [Google Scholar] [CrossRef]
- Rar, V.; Tkachev, S.; Tikunova, N. Genetic diversity of Anaplasma bacteria: Twenty years later. Infect. Genet. Evol. 2021, 91, 104833. [Google Scholar] [CrossRef]
- Ooshiro, M.; Zakimi, S.; Matsukawa, Y.; Katagiri, Y.; Inokuma, H. Detection of Anaplasma bovis and Anaplasma phagocytophilum from cattle on Yonaguni Island, Okinawa, Japan. Vet. Parasitol. 2008, 154, 360–364. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Cui, Y.; Yan, Y.; Wang, X.; Wang, R.; Jian, F.; Zhang, L.; Ning, C. A rapid, simple and sensitive loop-mediated isothermal amplification method to detect Anaplasma bovis in sheep and goats samples. Parasitol. Int. 2018, 67, 70–73. [Google Scholar] [CrossRef]
- Sakamoto, L.; Ichikawa, Y.; Sakata, Y.; Matsumoto, K.; Inokuma, H. Detection of Anaplasma bovis DNA in the peripheral blood of domestic dogs in Japan. Jpn. J. Infect. Dis. 2010, 63, 349–352. [Google Scholar] [CrossRef]
- Tateno, M.; Nishio, T.; Sakuma, M.; Nakanishi, N.; Izawa, M.; Asari, Y.; Okamura, M.; Maruyama, S.; Miyama, T.S.; Setoguchi, A.; et al. Molecular epidemiologic survey of Bartonella, Ehrlichia, and Anaplasma infections in Japanese Iriomote and Tsushima leopard cats. J. Wildl. Dis. 2013, 49, 646–652. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.P.; Tie, W.F.; Meng, S.; Li, D.; Wang, J.L.; Du, L.Y.; Xie, G.C. Extensive genetic diversity of Anaplasma bovis in ruminants in Xi’an, China. Ticks Tick-Borne Dis. 2020, 11, 101477. [Google Scholar] [CrossRef]
- Guo, W.P.; Wang, X.; Li, Y.N.; Xu, G.; Wang, Y.H.; Zhou, E.M. GroEL gene typing and genetic diversity of Anaplasma bovis in ticks in Shaanxi, China. Infect. Genet. Evol. 2019, 74, 103927. [Google Scholar] [CrossRef] [PubMed]
- Cabezas-Cruz, A.; Gallois, M.; Fontugne, M.; Allain, E.; Denoual, M.; Moutailler, S.; Devillers, E.; Zientara, S.; Memmi, M.; Chauvin, A.; et al. Epidemiology and genetic diversity of Anaplasma ovis in goats in Corsica, France. Parasit. Vectors 2019, 12, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enkhtaivan, B.; Narantsatsral, S.; Davaasuren, B.; Otgonsuren, D.; Amgalanbaatar, T.; Uuganbayar, E.; Zoljargal, M.; Myagmarsuren, P.; Suganuma, K.; Molefe, N.I.; et al. Molecular detection of Anaplasma ovis in small ruminants and ixodid ticks from Mongolia. Parasitol. Int. 2019, 69, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.P.; Huang, B.; Zhao, Q.; Xu, G.; Liu, B.; Wang, Y.H.; Zhou, E.M. Human-pathogenic Anaplasma spp., and Rickettsia spp. in animals in Xi’an, China. PLoS Negl. Trop. Dis. 2018, 12, e0006916. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Categories | Sampling Sites | Farms | Management | No. Total | No. Tested |
|---|---|---|---|---|---|
| Guanzhong dairy goat | Sanyuan | Farm 1 | Good | 200 | 26 |
| Lantian | Farm 2 | Medium | 280 | 39 | |
| Farm 3 | Good | 150 | 17 | ||
| Farm 4 | Good | 146 | 15 | ||
| Farm 5 | Poor | 240 | 27 | ||
| Linyou | Farm 6 | Poor | 500 | 91 | |
| Farm 7 | Good | 100 | 12 | ||
| Saanen dairy goat | Sanyuan | Farm 8 | Good | 120 | 22 |
| Shaanbei white cashmere goat | Mizhi | Farm 9 | Medium | 200 | 37 |
| Suide | Farm 10 | Poor | 220 | 39 | |
| Farm 11 | Good | 200 | 24 | ||
| Qingjian | Farm 12 | Medium | 200 | 39 | |
| Shaannan white goat | Zhenba | Farm 13 | Poor | 230 | 40 |
| Farm 14 | Poor | 210 | 39 | ||
| Farm 15 | Poor | 210 | 42 |
| Pathogen | Target Gene | Primer | Sequence (5′–3′) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|---|
| A. phagocytophilum | 16S rRNA | EE1 | TCCTGGCTCAGAACGAACGCTGGCGGC | 1430 | [25,26] |
| EE2 | AGTCACTGACCCAACCTTAAATGGCTG | ||||
| SSAP2f | GCTGAATGTGGGGATAATTTAT | 641 | |||
| SSAP2r | ATGGCTGCTTCCTTTCGGTTA | ||||
| A. bovis | 16S rRNA | EE1 | TCCTGGCTCAGAACGAACGCTGGCGGC | 1430 | [25,26] |
| EE2 | AGTCACTGACCCAACCTTAAATGGCTG | ||||
| AB1f | CTCGTAGCTTGCTATGAGAAC | 551 | |||
| AB1r | TCTCCCGGACTCCAGTCTG | ||||
| A. ovis | msp4 | MSP4f | CCGGATCCTTAGCTGAACAGGAATCTTGC | 867 | [27] |
| MSP4r | GGGAGCTCCTATGAATTACAGAGAATTGTTTAC |
| Factor | Production Categories | No. Tested | No. Positive (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | AP a | AB b | AO c | AP + AB d | AB + AO e | AP + AO f | AP + AB + AO g | ||||
| Production | |||||||||||
| Guanzhong dairy goat | Farm 1 | 26 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| Farm 2 | 39 | 23 (59.0) | 13 (33.3) | 18 (46.2) | 0 (0) | 8 (20.5) | 0 (0) | 0 (0) | 0 (0) | ||
| Farm 3 | 17 | 1 (5.9) | 1 (5.9) | 1 (5.9) | 0 (0) | 1 (5.9) | 0 (0) | 0 (0) | 0 (0) | ||
| Farm 4 | 15 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | ||
| Farm 5 | 27 | 22 (81.5) | 21 (77.8) | 15 (55.6) | 0 (0) | 14 (51.9) | 0 (0) | 0 (0) | 0 (0) | ||
| Farm 6 | 91 | 65 (71.4) | 38 (41.8) | 45 (49.5) | 4 (4.4) | 19 (20.9) | 0 (0) | 1 (1.1) | 1 (1.1) | ||
| Farm 7 | 12 | 3 (25.0) | 0 (0) | 3 (25.0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | ||
| Saanen dairy goat | Farm 8 | 22 | 1 (4.5) | 0 (0) | 1 (4.5) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| Subtotal | 249 | 115 (46.2) | 73 (29.3) | 83 (33.3) | 4 (1.6) | 42 (16.9) | 0 (0) | 1 (0.4) | 1 (0.4) | ||
| Shaanbei white cashmere goat | Farm 9 | 37 | 20 (54.1) | 0 (0) | 2 (5.4) | 20 (54.1) | 0 (0) | 2 (5.4) | 0 (0) | 0 (0) | |
| Farm 10 | 39 | 31 (79.5) | 8 (20.5) | 23 (59.0) | 18 (46.2) | 3 (7.7) | 6 (15.4) | 1 (2.6) | 4 (10.3) | ||
| Farm 11 | 24 | 6 (25.0) | 2 (8.3) | 3 (12.5) | 4 (16.7) | 1 (4.2) | 0 (0) | 0 (0) | 1 (4.2) | ||
| Farm 12 | 39 | 24 (61.5) | 14 (35.9) | 10 (25.6) | 10 (25.6) | 5 (12.8) | 2 (5.1) | 3 (7.7) | 0 (0) | ||
| Subtotal | 139 | 81 (58.3) | 24 (17.3) | 38 (27.3) | 52 (37.4) | 9 (6.5) | 10 (7.2) | 4 (2.9) | 5 (3.6) | ||
| Shaannan white goat | Farm 13 | 40 | 36 (90.0) | 32 (80.0) | 33 (82.5) | 0 (0) | 29 (72.5) | 0 (0) | 0 (0) | 0 (0) | |
| Farm 14 | 39 | 34 (87.2) | 33 (84.6) | 10 (25.6) | 0 (0) | 9 (23.1) | 0 (0) | 0 (0) | 0 (0) | ||
| Farm 15 | 42 | 32 (76.2) | 26 (61.9) | 20 (47.6) | 0 (0) | 14 (33.3) | 0 (0) | 0 (0) | 0 (0) | ||
| Subtotal | 121 | 102 (84.3) | 91 (75.2) | 63 (52.1) | 0 (0) | 52 (43.0) | 0 (0) | 0 (0) | 0 (0) | ||
| Ages | |||||||||||
| <2 months | 15 | 12 (80.0) | 4 (26.7) | 12 (80.0) | 4 (26.7) | 2 (13.3) | 2 (13.3) | 0 (0) | 2 (13.3) | ||
| 3–6 months | 99 | 44 (44.4) | 27 (27.3) | 25 (25.3) | 14 (14.1) | 15 (15.2) | 2 (2.0) | 3 (3.0) | 1 (1.0) | ||
| 7–12 months | 60 | 45 (75.0) | 26 (43.3) | 27 (45.0) | 14 (23.3) | 13 (21.7) | 4 (6.7) | 1 (1.7) | 2 (3.3) | ||
| >12 months | 335 | 197 (58.8) | 131 (39.1) | 120 (35.8) | 24 (7.2) | 73 (21.8) | 2 (0.6) | 1 (0.3) | 1 (0.3) | ||
| Total | 509 | 298 (58.5) | 188 (36.9) | 184 (36.1) | 56 (11.0) | 103 (20.2) | 10 (2.0) | 5 (1.0) | 6 (1.2) | ||
| Pathogen | Meat Goat | Cashmere Goat | Dairy Goat | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| n | χ2 | n | χ2 | OR (95% CI) | p | n | χ2 | OR (95% CI) | p | |
| Any Anaplasma | 102 | REF | 81 | 21.018 | 3.84 (2.12–6.97) | <0.001 | 115 | 48.773 | 6.26 (3.61–10.84) | <0.001 |
| A. phagocytophilum | 91 | REF | 24 | 88.038 | 14.54 (7.95–26.57) | <0.001 | 73 | 69.486 | 7.31 (4.46–11.99) | <0.001 |
| A. bovis | 63 | REF | 38 | 16.651 | 2.89 (1.72–4.84) | <0.001 | 83 | 11.962 | 2.17 (1.39–3.39) | 0.001 |
| A. ovis | 0 | REF | 52 | 56.583 | NA | <0.001 | 4 | 1.965 | NA | 0.161 |
| Pathogen | Production Categories | Ages | Sampling Sites (All) | Sampling Sites (Positive) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| χ2 | df | p | χ2 | df | p | χ2 | df | p | χ2 | df | p | |
| Any Anaplasma | 48.743 | 2 | <0.001 | 17.659 | 3 | 0.001 | 174.903 | 14 | <0.001 | 117.44 | 12 | <0.001 |
| A. phagocytophilum | 105.376 | 2 | <0.001 | 6.378 | 3 | 0.095 | 187.211 | 14 | <0.001 | 96.34 | 9 | <0.001 |
| A. bovis | 18.812 | 2 | <0.001 | 19.641 | 3 | <0.001 | 126.079 | 14 | <0.001 | 98.871 | 12 | <0.001 |
| A. ovis | 136.408 | 2 | <0.001 | 19.113 | 3 | <0.001 | 167.124 | 14 | <0.001 | 48.265 | 4 | <0.001 |
| Pathgen | Factor | Specimens Size of Factor (n) | Univariate Model | Multivariate Model | ||||
|---|---|---|---|---|---|---|---|---|
| OR | 95% CI | p | OR | 95% CI | p | |||
| Any Anaplasma (n = 298) | Production categories | |||||||
| meat goat | 102 | 0.36 | 0.25–0.52 | <0.001 | 0.97 | 0.35–4.79 | 0.982 | |
| cashmere goat | 81 | 0.98 | 0.66–1.47 | 0.939 | NA | NA | NA | |
| dairy goat | 115 | 5.26 | 3.17–9.16 | <0.001 | 0.79 | 0.48–1.27 | 0.532 | |
| Ages | ||||||||
| <2 months | 12 | 2.91 | 0.91–12.89 | 0.101 | 0.52 | 0.07–4.95 | 0.004 | |
| 3–6 months | 44 | 0.49 | 0.31–0.76 | 0.002 | 0.26 | 0.10–0.63 | 0.017 | |
| 7–12 months | 45 | 2.32 | 1.29–4.42 | 0.007 | 0.27 | 0.09–0.79 | <0.001 | |
| >12 months | 197 | 1.03 | 0.71–1.49 | 0.869 | NA | NA | NA | |
| Sampling sites | ||||||||
| Lantian | 46 | 0.35 | 0.23–0.54 | <0.001 | 0.98 | 0.36–4.82 | 0.982 | |
| Linyou | 68 | 1.49 | 0.95–2.36 | 0.086 | 0.97 | 0.32–4.73 | 0.981 | |
| Mizhi | 20 | 0.82 | 0.42–1.62 | 0.565 | NA | NA | NA | |
| Qingjian | 24 | 2.95 | 1.39–7.01 | 0.008 | 9.8 | 2.14–47.59 | 0.004 | |
| Suide | 37 | 0.6 | 0.35–1.03 | 0.062 | 2.97 | 0.89–10.33 | 0.081 | |
| Sanyuan | 1 | 0.98 | 0.82–1.60 | 0.971 | NA | NA | NA | |
| Zhenba | 102 | 5.26 | 3.17–9.16 | <0.001 | 8.94 | 3.45–24.83 | <0.001 | |
| A. phagocytophilum (n = 188) | Production categories | |||||||
| meat goat | 91 | 9.1 | 5.74–14.79 | <0.001 | 0.98 | 0.86–1.72 | 0.98 | |
| cashmere goat | 24 | 0.26 | 0.16–0.42 | <0.001 | 0.89 | 0.37–2.31 | 0.837 | |
| dairy goat | 73 | 0.52 | 0.36–0.75 | 0.001 | 0.41 | 0.25–0.69 | 0.01 | |
| Ages | ||||||||
| <2 months | 4 | 0.61 | 0.17–1.82 | 0.407 | NA | NA | NA | |
| 3–6 months | 27 | 0.58 | 0.35–0.93 | 0.028 | 0.89 | 0.29–2.68 | 0.836 | |
| 7–12 months | 26 | 1.35 | 0.78–2.33 | 0.276 | NA | NA | NA | |
| >12 months | 131 | 1.32 | 0.9–1.94 | 0.16 | 3.23 | 1.32–8.04 | <0.001 | |
| Sampling sites | ||||||||
| Lantian | 35 | 0.64 | 0.4–0.98 | 0.045 | 1.17 | 0.67–2.05 | 0.575 | |
| Linyou | 38 | 0.99 | 0.92–1.85 | 0.992 | NA | NA | NA | |
| Mizhi | 0 | 0.97 | 0.85–1.72 | 0.979 | NA | NA | NA | |
| Qingjian | 14 | 0.42 | 0.17–0.88 | 0.031 | 0.96 | 0.67–1.82 | 0.979 | |
| Suide | 10 | 0.54 | 0.29–0.97 | 0.045 | 0.96 | 0.57–1.79 | 0.978 | |
| Sanyuan | 0 | 0.97 | 0.77–1.59 | 0.973 | NA | NA | NA | |
| Zhenba | 91 | 9.1 | 5.74–14.79 | <0.001 | 12.84 | 6.53–26.8 | <0.001 | |
| A. bovis (n = 184) | Production categories | |||||||
| meat goat | 63 | 2.4 | 1.58–3.64 | <0.001 | 1.97 | 0.96–4.82 | 0.089 | |
| cashmere goat | 38 | 0.58 | 0.37–0.88 | 0.012 | 1.28 | 0.47–2.77 | 0.423 | |
| dairy goat | 83 | 0.79 | 0.55–1.13 | 0.196 | 0.45 | 0.21–0.86 | 0.314 | |
| Ages | ||||||||
| <2 months | 12 | 7.49 | 2.34–33.19 | 0.002 | 5.06 | 0.88–42.36 | <0.001 | |
| 3–6 months | 25 | 0.53 | 0.32–0.86 | 0.013 | 0.73 | 0.32–1.56 | 0.645 | |
| 7–12 months | 27 | 1.52 | 0.88–2.62 | 0.131 | 0.66 | 0.3–1.47 | 0.578 | |
| >12 months | 120 | 0.96 | 0.66–1.41 | 0.831 | NA | NA | NA | |
| Sampling sites | ||||||||
| Lantian | 34 | 0.66 | 0.42–1.03 | 0.07 | 1.03 | 0.51–2.41 | 0.982 | |
| Linyou | 48 | 1.73 | 1.12–2.69 | 0.014 | 1.05 | 0.63–2.67 | 0.982 | |
| Mizhi | 2 | 0.09 | 0.01–0.3 | 0.001 | 1.05 | 0.66–2.83 | 0.985 | |
| Qingjian | 10 | 2.76 | 1.43–5.46 | 0.003 | 0.97 | 0.65–1.43 | 0.981 | |
| Suide | 26 | 0.42 | 0.21–0.77 | 0.008 | 0.98 | 0.77–1.69 | 0.983 | |
| Sanyuan | 1 | 0.97 | 0.83–1.69 | 0.973 | NA | NA | NA | |
| Zhenba | 63 | 2.4 | 1.58–3.64 | <0.001 | 0.97 | 0.89–1.87 | 0.981 | |
| A. ovis (n = 56) | Production categories | |||||||
| meat goat | 0 | 0.98 | 0.86–1.65 | 0.985 | NA | NA | NA | |
| cashmere goat | 52 | 9.86 | 3.69–24.18 | <0.001 | 0.98 | 0.37–4.01 | 0.937 | |
| dairy goat | 4 | 0.07 | 0.02–0.16 | <0.001 | 1.02 | 0.57–2.36 | 0.981 | |
| Ages | ||||||||
| <2 months | 4 | 3.09 | 0.83–9.41 | 0.061 | 1.03 | 0.49–2.45 | 0.986 | |
| 3–6 months | 14 | 1.44 | 0.73–2.7 | 0.268 | NA | NA | NA | |
| 7–12 months | 14 | 2.95 | 1.46–5.7 | 0.002 | 1.04 | 0.63–2.71 | 0.986 | |
| >12 months | 24 | 0.34 | 0.19–0.6 | <0.001 | 0.96 | 0.32–3.97 | 0.932 | |
| Sampling sites | ||||||||
| Lantian | 0 | 0.98 | 0.89–1.77 | 0.986 | NA | NA | NA | |
| Linyou | 4 | 0.28 | 0.08–0.69 | 0.015 | 1.03 | 0.41–2.09 | 0.991 | |
| Mizhi | 20 | 14.25 | 6.89–29.94 | <0.001 | 0.97 | 0.41–4.07 | 0.992 | |
| Qingjian | 10 | 9.74 | 4.76–19.91 | <0.001 | 0.98 | 0.37–3.89 | 0.994 | |
| Suide | 22 | 2.75 | 1.36–5.29 | 0.003 | 0.98 | 0.42–3.88 | 0.995 | |
| Sanyuan | 0 | 0.96 | 0.66–1.71 | 0.984 | NA | NA | NA | |
| Zhenba | 0 | 0.96 | 0.71–1.69 | 0.985 | NA | NA | NA | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Fu, M.; Yu, Z.; Wang, J.; Song, J.; Zhao, G. Molecular Characterization of Anaplasma spp. among Dairy, Cashmere, and Meat Goats in Shaanxi Province, Northwestern China. Animals 2022, 12, 1566. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121566
Yang X, Fu M, Yu Z, Wang J, Song J, Zhao G. Molecular Characterization of Anaplasma spp. among Dairy, Cashmere, and Meat Goats in Shaanxi Province, Northwestern China. Animals. 2022; 12(12):1566. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121566
Chicago/Turabian StyleYang, Xin, Mingzhe Fu, Zhengqing Yu, Junwei Wang, Junke Song, and Guanghui Zhao. 2022. "Molecular Characterization of Anaplasma spp. among Dairy, Cashmere, and Meat Goats in Shaanxi Province, Northwestern China" Animals 12, no. 12: 1566. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121566