
Two Different Copy Number Variations of the SOX5 and SOX8 Genes in Yak and Their Association with Growth Traits

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sample Collection and Growth Trait Measurements
2.3. Genomic DNA Extraction and RNA Isolation
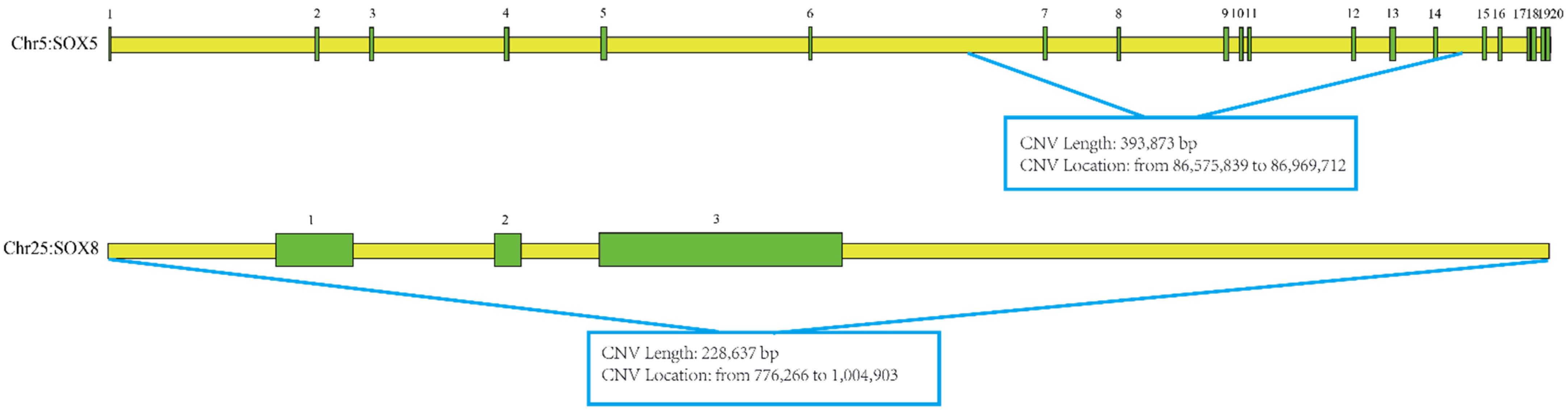
2.4. Candidate Gene information
2.5. Primer Design
2.6. Quantitative PCR Analysis
2.7. Statistical Analysis
3. Results
3.1. Association Analysis between CNV and Growth Traits
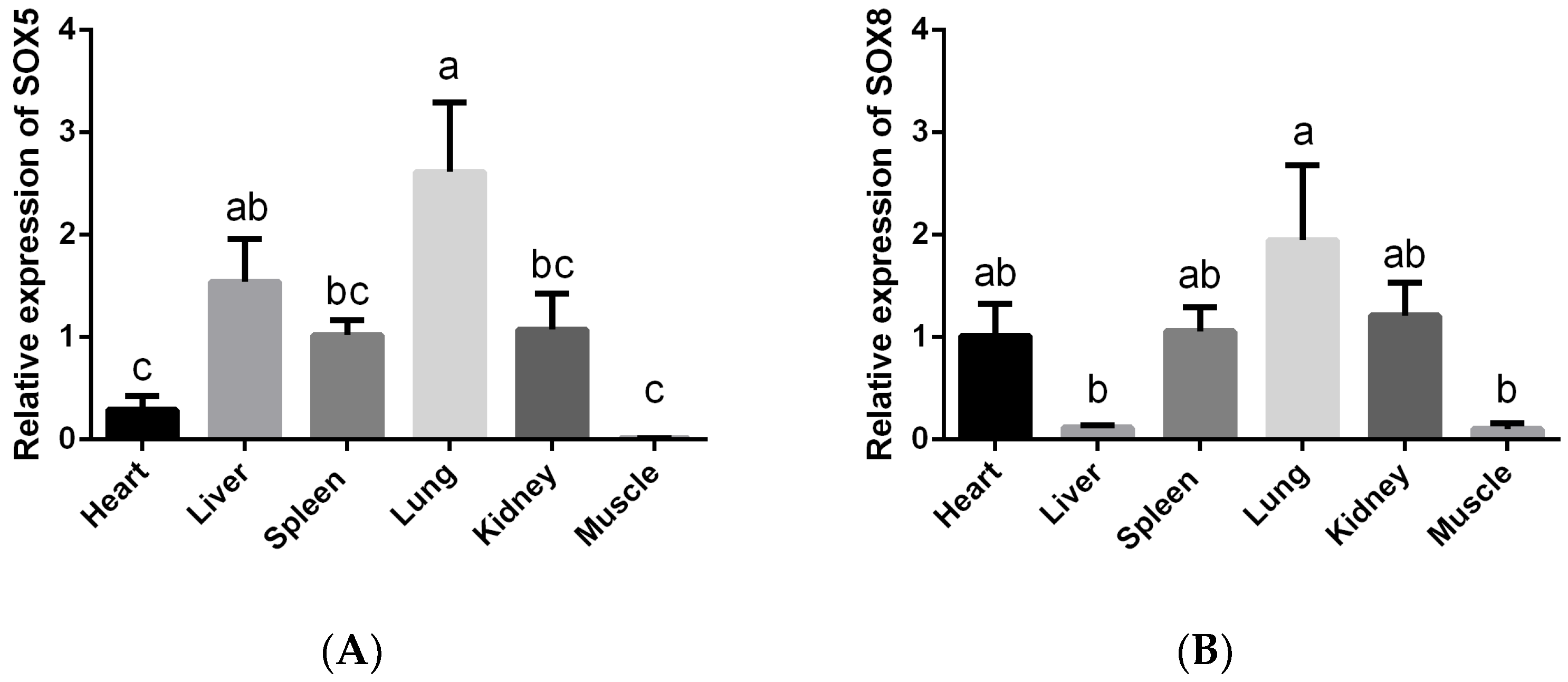
3.2. Gene Expression Profiling of SOX5 and SOX8
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Z.; Jiang, M. Metabolomic profiles in yak mammary gland tissue during the lactation cycle. PLoS ONE 2019, 14, e0219220. [Google Scholar] [CrossRef] [PubMed]
- Long, L.; Zhu, Y.; Li, Z.; Zhang, H.; Liu, L.; Bai, J. Differential expression of skeletal muscle mitochondrial proteins in yak, dzo, and cattle: A proteomics-based study. J. Vet. Med. Sci. 2020, 82, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pei, J.; Bao, P.; Cao, M.; Guo, S.; Song, R.; Song, W.; Liang, C.; Yan, P.; Guo, X. Mitogenomic diversity and phylogeny analysis of yak (Bos grunniens). BMC Genom. 2021, 22, 325. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.W.; Lin, Y.T.; Ding, S.T.; Lo, L.L.; Wang, P.H.; Lin, E.C.; Liu, F.W.; Lu, Y.W. Efficient SNP Discovery by Combining Microarray and Lab-on-a-Chip Data for Animal Breeding and Selection. Microarrays 2015, 4, 570–595. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.; Hou, D.; Feng, Y.; Li, T.; Jing, Z.; Li, W.; Han, R.; Li, G.; Sun, G.; Tian, Y.; et al. Molecular characterization and a duplicated 31-bp indel within the LDB2 gene and its associations with production performance in chickens. Gene 2020, 761, 145046. [Google Scholar] [CrossRef]
- Du, H.; Zheng, X.; Zhao, Q.; Hu, Z.; Wang, H.; Zhou, L.; Liu, J.F. Analysis of Structural Variants Reveal Novel Selective Regions in the Genome of Meishan Pigs by Whole Genome Sequencing. Front. Genet. 2021, 12, 550676. [Google Scholar] [CrossRef]
- Dong, Y.; Shi, S.; Bao, Y.; Yao, H.; Jing, Y.; Lin, Z. Development of 90 EST-SNP markers in blood clam (Tegillarca granosa) using high resolution melting (HRM). Conserv. Genet. Resour. 2014, 7, 309–314. [Google Scholar] [CrossRef]
- Li, J.; Zhu, X.; Ma, L.; Xu, H.; Cao, X.; Luo, R.; Chen, H.; Sun, X.; Cai, Y.; Lan, X. Detection of a new 20-bp insertion/deletion (indel) within sheep PRND gene using mathematical expectation (ME) method. Prion 2017, 11, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Ju, X.; Huang, X.; Zhang, M.; Lan, X.; Wang, D.; Wei, C.; Jiang, H. Effects of eight InDel variants in FHIT on milk traits in Xinjiang brown cattle. Anim. Biotechnol. 2021, 32, 486–494. [Google Scholar] [CrossRef]
- Guo, X.; Pei, J.; Wu, X.; Bao, P.; Ding, X.; Xiong, L.; Chu, M.; Lan, X.; Yan, P. Detection of InDel and CNV of SPAG17 gene and their associations with bovine growth traits. Anim. Biotechnol. 2020, 33, 440–447. [Google Scholar] [CrossRef]
- Ren, H.; Tang, Q.; Xue, T.; Wang, Q.; Xu, H.; Zhang, Q.; Pan, C.A. 24-bp indel within the sheep AHR gene is associated with litter size. Anim. Biotechnol. 2021, 4, 1–6. [Google Scholar] [CrossRef]
- Lin, W.; Ren, T.; Li, W.; Liu, M.; He, D.; Liang, S.; Luo, W.; Zhang, X. Novel 61-bp Indel of RIN2 Is Associated With Fat and Hatching Weight Traits in Chickens. Front. Genet. 2021, 12, 672888. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.J.; Cao, X.K.; Dong, D.; Liu, M.; Hao, D.; Shen, X.M.; Huang, Y.Z.; Lei, C.Z.; Ma, Y.; Bai, Y.Y.; et al. Integrative analysis of APOL3 gene CNV for adult cattle stature. Anim. Biotechnol. 2020, 31, 440–446. [Google Scholar] [CrossRef]
- Ma, Y.L.; Wen, Y.F.; Cao, X.K.; Cheng, J.; Huang, Y.Z.; Ma, Y.; Hu, L.Y.; Lei, C.Z.; Qi, X.L.; Cao, H.; et al. Copy number variation (CNV) in the IGF1R gene across four cattle breeds and its association with economic traits. Arch. Anim. Breed. 2019, 62, 171–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, R.; Cheng, J.; Cao, X.K.; Ma, Y.L.; Chaogetu, B.; Huang, Y.Z.; Lan, X.Y.; Lei, C.Z.; Hu, L.Y.; Chen, H. Copy Number Variation of the SHE Gene in Sheep and Its Association with Economic Traits. Animals 2019, 9, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bickhart, D.M.; Liu, G.E. The challenges and importance of structural variation detection in livestock. Front. Genet. 2014, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Spielmann, M.; Klopocki, E. CNVs of noncoding cis-regulatory elements in human disease. Curr. Opin. Genet. Dev. 2013, 23, 249–256. [Google Scholar] [CrossRef]
- Boone, P.M.; Campbell, I.M.; Baggett, B.C.; Soens, Z.T.; Rao, M.M.; Hixson, P.M.; Patel, A.; Bi, W.; Cheung, S.W.; Lalani, S.R.; et al. Deletions of recessive disease genes: CNV contribution to carrier states and disease-causing alleles. Genom. Res. 2013, 23, 1383–1394. [Google Scholar] [CrossRef] [Green Version]
- E, G.X.; Zhu, Y.B.; Basang, W.D.; Na, R.S.; Han, Y.G.; Zeng, Y. Comparative and selection sweep analysis of CNV was associated to litter size in Dazu black goats. Anim. Biotechnol. 2021, 32, 792–797. [Google Scholar] [CrossRef]
- Mei, C.; Junjvlieke, Z.; Raza, S.H.; Wang, H.; Cheng, G.; Zhao, C.; Zhu, W.; Zan, L. Copy number variation detection in Chinese indigenous cattle by whole genome sequencing. Genomics 2020, 112, 831–836. [Google Scholar] [CrossRef]
- Zhang, L.; Jia, S.; Yang, M.; Xu, Y.; Li, C.; Sun, J.; Huang, Y.; Lan, X.; Lei, C.; Zhou, Y.; et al. Detection of copy number variations and their effects in Chinese bulls. BMC Genom. 2014, 15, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Utsunomiya, Y.T.; Xu, L.; Hay, H.A.; Bickhart, D.M.; Alexandre, P.A.; Rosen, B.D.; Schroeder, S.G.; Carvalheiro, R.; de Neves, H.H.; et al. Genome-wide CNV analysis reveals variants associated with growth traits in Bos indicus. BMC Genom. 2016, 17, 419. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Shen, X.; Yang, Y.; Yang, H.; Qi, A.; Yang, S.; Qu, K.; Lan, X.; Huang, B.; Chen, H. Two Different Copy Number Variations of the CLCN2 Gene in Chinese Cattle and Their Association with Growth Traits. Animals 2021, 12, 41. [Google Scholar] [CrossRef] [PubMed]
- Kamachi, Y.; Kondoh, H. Sox proteins: Regulators of cell fate specification and differentiation. Development 2013, 140, 4129–4144. [Google Scholar] [CrossRef] [Green Version]
- Collignon, J.; Sockanathan, S.; Hacker, A.; Cohen, T.M.; Norris, D.; Rastan, S.; Stevanovic, M.; Goodfellow, P.N.; Lovell, B.R. A comparison of the properties of Sox-3 with Sry and two related genes, Sox-1 and Sox-2. Development 1996, 122, 509–520. [Google Scholar] [CrossRef]
- Uwanogho, D.; Rex, M.; Cartwright, E.J.; Pearl, G.; Healy, C.; Scotting, P.J.; Sharpe, P.T. Embryonic expression of the chicken Sox2, Sox3 and Sox11 genes suggests an interactive role in neuronal development. Mech. Dev. 1995, 49, 23–36. [Google Scholar] [CrossRef]
- Lefebvre, V.; Behringer, R.R.; Crombrugghe, B. L-Sox5, Sox6 and Sox9 control essential steps of the chondrocyte differentiation pathway. Osteoarthr. Cartil. 2001, 9, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, H.; Kim, J.E.; Nakashima, K.; Balmes, G.; Iwai, N.; Deng, J.M.; Zhang, Z.; Martin, J.F.; Behringer, R.R.; Nakamura, T.; et al. Osteo-chondroprogenitor cells are derived from Sox9 expressing precursors. Proc. Natl. Acad. Sci. USA 2005, 102, 14665–14670. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, K.; Schinke, T.; Haberland, M.; Priemel, M.; Schilling, A.F.; Mueldner, C.; Rueger, J.M.; Sock, E.; Wegner, M.; Amling, M. The high mobility group transcription factor Sox8 is a negative regulator of osteoblast differentiation. J. Cell Biol. 2005, 168, 899–910. [Google Scholar] [CrossRef] [Green Version]
- Jia, C.; Wang, H.; Li, C.; Wu, X.; Zan, L.; Ding, X.; Guo, X.; Bao, P.; Pei, J.; Chu, M.; et al. Genome-wide detection of copy number variations in polled yak using the Illumina BovineHD BeadChip. BMC Genom. 2019, 20, 376. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, R.P.; Bailey, D.R.; Shannon, N.H. Linear body measurements of cattle before and after 20 years of selection for postweaning gain when fed two different diets. J. Anim. Sci. 1993, 71, 1712–1720. [Google Scholar] [CrossRef] [PubMed]
- Bickhart, D.M.; Hou, Y.; Schroeder, S.G.; Alkan, C.; Cardone, M.F.; Matukumalli, L.K.; Song, J.; Schnabel, R.D.; Ventura, M.; Taylor, J.F.; et al. Copy number variation of individual cattle genomes using next-generation sequencing. Genom. Res. 2012, 22, 778–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, J.S.; Cheong, H.S.; Kim, L.H.; Nam Gung, S.; Park, T.J.; Chun, J.Y.; Kim, J.Y.; Pasaje, C.F.; Lee, J.S.; Shin, H.D. Identification of copy number variations and common deletion polymorphisms in cattle. BMC Genom. 2010, 11, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, P.; Harley, V.R. Acquisition of SOX transcription factor specificity through protein-protein interaction, modulation of Wnt signalling and post-translational modification. Int. J. Biochem. Cell Biol. 2010, 42, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, V. Roles and regulation of SOX transcription factors in skeletogenesis. Curr. Top. Dev. Biol. 2019, 133, 171–193. [Google Scholar] [PubMed]
- Lamb, A.N.; Rosenfeld, J.A.; Neill, N.J.; Talkowski, M.E.; Blumenthal, I.; Girirajan, S.; Keelean-Fuller, D.; Fan, Z.; Pouncey, J.; Stevens, C.; et al. Haploinsufficiency of SOX5 at 12p12.1 is associated with developmental delays with prominent language delay, behavior problems, and mild dysmorphic features. Hum. Mutat. 2012, 33, 728–740. [Google Scholar] [CrossRef] [Green Version]
- Nesbitt, A.; Bhoj, E.J.; McDonald, G.K.; Yu, Z.; Denenberg, E.; Sarmady, M.; Tischler, T.; Cao, K.; Dubbs, H.; Zackai, E.H.; et al. Exome sequencing expands the mechanism of SOX5-associated intellectual disability: A case presentation with review of sox-related disorders. Am. J. Med. Genet. A 2015, 167, 2548–2554. [Google Scholar] [CrossRef]
- Bell, K.M.; Western, P.S.; Sinclair, A.H. SOX8 expression during chick embryogenesis. Mech. Dev. 2000, 94, 257–260. [Google Scholar] [CrossRef]
- Chimal, M.J.; Rodriguez, L.J.; Montero, J.A.; Gañan, Y.; Macias, D.; Merino, R.; Hurle, J.M. Analysis of the molecular cascade responsible for mesodermal limb chondrogenesis: Sox genes and BMP signaling. Dev. Biol. 2003, 257, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Ge, Q.; Guo, Y.; Zheng, W.; Zhao, S.; Cai, Y.; Qi, X. Molecular mechanisms detected in yak lung tissue via transcriptome-wide analysis provide insights into adaptation to high altitudes. Sci. Rep. 2021, 11, 7786. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Level | Gene | Primer Pair Sequences 1 (5′-3′) | Amplicon Length (bp) | Tm (°C) |
|---|---|---|---|---|
| DNA | BTF3 | F: AACCAGGAGAAACTCGCCAA | 166 | 63 |
| R: TTCGGTGAAATGCCCTCTCG | ||||
| SOX5 | F: GCTTCCCAGTTCGCTTAG | 104 | 55.6 | |
| R: TTTCTGCCTTGGATGCTC | ||||
| SOX8 | F: CCTTGGGTCACTCGGGTTG | 141 | 63 | |
| R: GCGGCTCGGATTCTTTCG | ||||
| mRNA | GAPDH | F: CCACGAGAAGTATAACAACACC | 120 | 56.1 |
| R: GTCATAAGTCCCTCCACGAT | ||||
| SOX5 | F: AAGAAACTGGCTGCGTCTCA | 168 | 56.1 | |
| R: TAATGGCGGCAGTTGACCTT | ||||
| SOX8 | F: CCGCACATCAAGACGGAGCA | 213 | 64 | |
| R: TGACGGGTAGCCAGGGAACG |
| Age | Growth Trait 1 | CNV Type 2 (Mean ± SE) | p-Value 3 | ||
|---|---|---|---|---|---|
| Deletion (56) | Normal (123) | Duplication (147) | |||
| 6 months | BW (kg) | 82.89 ± 1.39 | 84.96 ± 0.94 | 84.33 ± 0.86 | 0.469 |
| WH (cm) | 94.75 ± 0.71 | 94.72 ± 0.48 | 94.00 ± 0.44 | 0.468 | |
| BL (cm) | 92.57 ± 0.98 | 92.30 ± 0.66 | 91.33 ± 0.60 | 0.418 | |
| CG (cm) | 123.13 ± 1.05 | 124.02 ± 0.71 | 124.47 ± 0.65 | 0.550 | |
| 12 months | BW (kg) | 81.24 ± 1.45 | 83.58 ± 0.99 | 82.72 ± 0.89 | 0.410 |
| WH (cm) | 89.87 ± 0.57 | 90.62 ± 0.39 | 90.58 ± 0.35 | 0.518 | |
| BL (cm) | 95.07 ± 0.68 | 95.82 ± 0.46 | 96.30 ± 0.42 | 0.297 | |
| CG (cm) | 116.42 ± 0.68 | 117.34 ± 0.46 | 117.38 ±.042 | 0.449 | |
| 18 months | BW (kg) | 119.60 ± 1.95 | 122.70 ± 1.33 | 123.36 ± 1.29 | 0.264 |
| WH (cm) | 103.77 ± 0.79 a | 103.44 ± 0.55 a | 100.03 ± 0.50 b | <0.01 ** | |
| BL (cm) | 102.32 ± 0.78 | 101.84 ± 0.55 | 101.55 ± 0.49 | 0.699 | |
| CG (cm) | 139.62 ± 1.38 | 139.28 ± 0.96 | 137.24 ± 0.87 | 0.183 | |
| 30 months | BW (kg) | 152.95 ± 2.43 | 155.60 ± 1.64 | 156.41 ± 1.49 | 0.479 |
| WH (cm) | 100.33 ± 0.79 | 99.77 ± 0.51 | 99.37 ± 0.47 | 0.558 | |
| BL (cm) | 112.31 ± 0.91 | 112.81 ± 0.59 | 113.42 ± 0.54 | 0.525 | |
| CG (cm) | 147.03 ± 1.34 | 146.32 ± 0.87 | 148.03 ± 0.81 | 0.352 | |
| Age | Growth Trait | CNV Type 1 (Mean ± SE) | p-Value 2 | ||
|---|---|---|---|---|---|
| Deletion (109) | Normal (100) | Duplication (117) | |||
| 6 months | BW (kg) | 83.24 ± 0.99 | 84.83 ± 1.04 | 84.90 ± 0.97 | 0.411 |
| WH (cm) | 94.44 ± 0.51 | 94.09 ± 0.53 | 94.62 ± 0.49 | 0.757 | |
| BL (cm) | 91.46 ± 0.70 | 92.98 ± 0.73 | 91.41 ± 0.67 | 0.211 | |
| CG (cm) | 123.73 ± 0.75 | 124.25 ± 0.79 | 124.22 ± 0.73 | 0.863 | |
| 12 months | BW (kg) | 82.27 ± 1.04 | 83.62 ± 1.09 | 82.56 ± 1.00 | 0.646 |
| WH (cm) | 90.24 ± 0.41 | 90.73 ± 0.43 | 90.46 ± 0.40 | 0.717 | |
| BL (cm) | 95.40 ± 0.49 | 96.23 ± 0.51 | 96.10 ± 0.47 | 0.437 | |
| CG (cm) | 117.02 ± 0.49 | 117.35 ± 0.51 | 117.24 ± 0.47 | 0.888 | |
| 18 months | BW (kg) | 122.94 ± 1.43 | 122.27 ± 1.52 | 122.01 ± 1.42 | 0.893 |
| WH (cm) | 102.76 ± 0.61 | 102.05 ± 0.62 | 101.13 ± 0.58 | 0.154 | |
| BL (cm) | 102.21 ± 0.58 | 101.76 ± 0.59 | 101.43 ± 0.55 | 0.626 | |
| CG (cm) | 138.83 ± 1.03 | 138.62 ± 1.05 | 137.87 ± 0.98 | 0.775 | |
| 30 months | BW (kg) | 153.95 ± 1.76 | 157.33 ± 1.75 | 155.27 ± 1.70 | 0.391 |
| WH (cm) | 100.47 ± 0.56 | 99.25 ± 0.55 | 99.35 ± 0.53 | 0.221 | |
| BL (cm) | 113.36 ± 0.64 | 112.20 ± 0.63 | 113.45 ± 0.62 | 0.290 | |
| CG (cm) | 147.99 ± 0.96 a | 145.18 ± 0.92 b | 148.45 ± 0.91 a | 0.027 * | |
| Age | CNV Combination Type | Growth Trait 1 (Mean ± SE) | |||
|---|---|---|---|---|---|
| BW (kg) | WH (cm) | BL (cm) | CG (cm) | ||
| 6 months | Deletion/Deletion (26) | 85.42 ± 2.21 | 95.38 ± 0.94 | 93.92 ± 1.23 | 124.31 ± 1.35 |
| Deletion/Normal (24) | 79.54 ± 2.09 | 93.71 ± 1.08 | 92.42 ± 1.80 | 121.13 ± 1.50 | |
| Deletion/Duplication (6) | 85.33 ± 5.12 | 96.17 ± 2.61 | 87.33 ± 1.36 | 126.00 ± 2.63 | |
| Normal/Deletion (41) | 82.54 ± 1.47 | 94.44 ± 0.81 | 90.68 ± 1.18 | 122.49 ± 1.35 | |
| Normal/Normal (41) | 87.85 ± 1.42 | 95.02 ± 0.96 | 93.98 ± 1.20 | 125.39 ± 1.27 | |
| Normal/Duplication (41) | 84.55 ± 1.64 | 94.68 ± 0.82 | 92.24 ± 1.37 | 124.17 ± 1.25 | |
| Duplication/Deletion (42) | 82.57 ± 1.61 | 93.86 ± 0.72 | 90.69 ± 1.00 | 124.60 ± 1.15 | |
| Duplication/Normal (35) | 85.00 ± 1.85 | 93.26 ± 0.84 | 92.20 ± 1.32 | 125.06 ± 1.39 | |
| Duplication/Duplication (70) | 85.07 ± 1.27 | 94.46 ± 0.66 | 91.27 ± 0.71 | 124.10 ± 0.92 | |
| p value2 | 0.109 | 0.787 | 0.216 | 0.501 | |
| 12 months | Deletion/Deletion (26) | 81.68 ± 2.58 | 90.08 ± 0.78 | 94.44 ± 1.08 | 117.08 ± 0.79 |
| Deletion/Normal (24) | 79.17 ± 1.95 | 89.46 ± 0.81 | 95.33 ± 0.67 | 115.08 ± 0.96 | |
| Deletion/Duplication (6) | 87.33 ± 4.04 | 90.67 ± 1.76 | 96.67 ± 2.39 | 119.00 ± 1.84 | |
| Normal/Deletion (41) | 82.67 ± 1.45 | 89.73 ± 0.65 | 95.00 ± 0.68 | 116.33 ± 0.97 | |
| Normal/Normal (41) | 85.13 ± 1.59 | 91.63 ± 0.67 | 96.65 ± 1.10 | 118.65 ± 0.81 | |
| Normal/Duplication (41) | 82.95 ± 1.87 | 90.50 ± 0.70 | 95.80 ± 0.92 | 117.05 ± 0.65 | |
| Duplication/Deletion (42) | 82.24 ± 1.60 | 90.83 ± 0.68 | 96.36 ± 0.78 | 117.64 ± 0.75 | |
| Duplication/Normal (35) | 84.88 ± 1.87 | 90.57 ± 0.71 | 96.37 ± 0.77 | 117.43 ± 0.90 | |
| Duplication/Duplication (70) | 81.93 ± 1.32 | 90.42 ± 0.53 | 96.23 ± 0.50 | 117.20 ± 0.62 | |
| p value | 0.460 | 0.638 | 0.690 | 0.269 | |
| 18 months | Deletion/Deletion (26) | 122.86 ± 2.82 | 104.46 ± 1.42 a | 104.25 ± 1.32 | 141.00 ± 1.68 |
| Deletion/Normal (24) | 116.20 ± 2.04 | 103.13 ± 1.17 a,b,c | 100.09 ± 1.11 | 137.91 ± 1.72 | |
| Deletion/Duplication (6) | 119.50 ± 7.54 | 103.50 ± 0.99 a,b | 103.17 ± 1.87 | 140.67 ± 4.06 | |
| Normal/Deletion (41) | 121.23 ± 2.88 | 103.56 ± 1.08 a,b | 101.12 ± 1.15 | 138.94 ± 2.38 | |
| Normal/Normal (41) | 125.09 ± 2.36 | 103.05 ± 0.90 a,b,c | 102.76 ± 0.91 | 139.66 ± 1.58 | |
| Normal/Duplication (41) | 121.68 ± 2.19 | 103.73 ± 0.66 a,b | 101.54 ± 0.90 | 139.22 ± 1.33 | |
| Duplication/Deletion (42) | 124.55 ± 2.58 | 101.03 ± 0.97 a,b,c | 101.90 ± 0.89 | 137.38 ± 1.57 | |
| Duplication/Normal (35) | 123.55 ± 2.37 | 100.09 ± 1.15 b,c | 101.78 ± 0.98 | 137.91 ± 1.49 | |
| Duplication/Duplication (70) | 122.46 ± 1.73 | 99.38 ± 0.74 c | 101.21 ± 0.64 | 136.81 ± 1.42 | |
| p value | 0.486 | <0.01 ** | 0.320 | 0.770 | |
| 30 months | Deletion/Deletion (26) | 154.71 ± 3.67 | 101.20 ± 1.44 | 114.07 ± 1.31 | 149.60 ± 2.67 |
| Deletion/Normal (24) | 149.68 ± 2.81 | 100.21 ± 1.19 | 110.53 ± 1.41 | 145.16 ± 1.33 | |
| Deletion/Duplication (6) | 159.40 ± 5.33 | 98.20 ± 0.73 | 113.80 ± 3.62 | 146.40 ± 1.94 | |
| Normal/Deletion (41) | 150.71 ± 2.98 | 99.79 ± 1.08 | 112.90 ± 1.05 | 146.90 ± 1.52 | |
| Normal/Normal (41) | 160.72 ± 2.85 | 99.15 ± 0.73 | 112.45 ± 1.07 | 144.50 ± 1.54 | |
| Normal/Duplication (41) | 154.70 ± 2.93 | 100.42 ± 0.80 | 113.10 ± 0.88 | 147.65 ± 1.63 | |
| Duplication/Deletion (42) | 156.30 ± 2.93 | 100.74 ± 1.09 | 113.45 ± 0.89 | 148.23 ± 1.53 | |
| Duplication/Normal (35) | 158.64 ± 2.79 | 98.72 ± 0.72 | 113.00 ± 1.21 | 145.93 ± 1.67 | |
| Duplication/Duplication (70) | 155.20 ± 2.20 | 98.80 ± 0.63 | 113.65 ± 0.78 | 149.19 ± 1.09 | |
| p value | 0.225 | 0.497 | 0.716 | 0.289 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Chu, M.; Bao, Q.; Bao, P.; Guo, X.; Liang, C.; Yan, P. Two Different Copy Number Variations of the SOX5 and SOX8 Genes in Yak and Their Association with Growth Traits. Animals 2022, 12, 1587. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121587
Zhang Z, Chu M, Bao Q, Bao P, Guo X, Liang C, Yan P. Two Different Copy Number Variations of the SOX5 and SOX8 Genes in Yak and Their Association with Growth Traits. Animals. 2022; 12(12):1587. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121587
Chicago/Turabian StyleZhang, Zhilong, Min Chu, Qi Bao, Pengjia Bao, Xian Guo, Chunnian Liang, and Ping Yan. 2022. "Two Different Copy Number Variations of the SOX5 and SOX8 Genes in Yak and Their Association with Growth Traits" Animals 12, no. 12: 1587. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121587