
Immune Modulation Ability of Hepcidin from Teleost Fish

 , , and
, , and 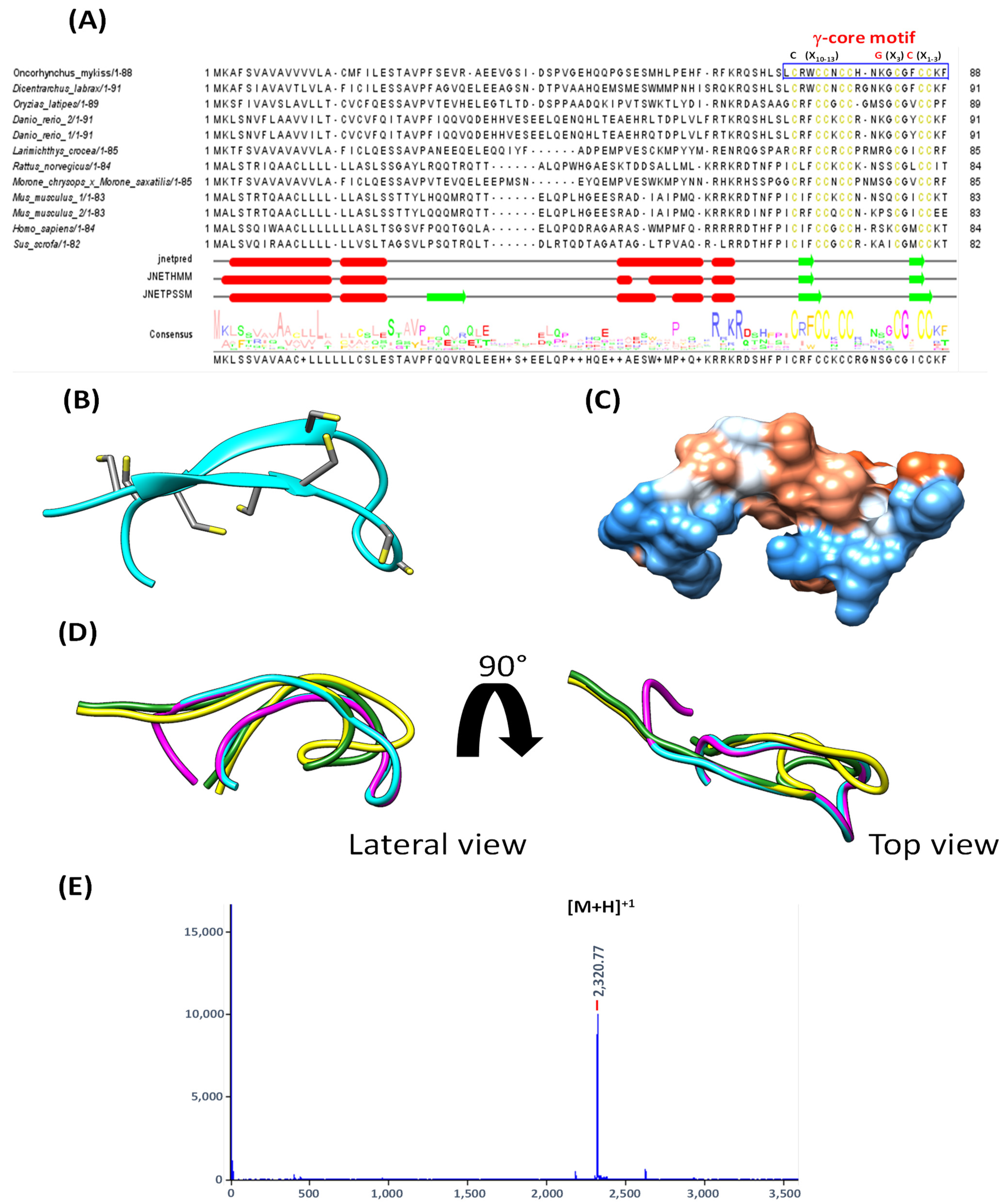{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Peptide Synthesis
2.2. Cell Culture and Incubation with Hep20
2.3. Experiment Fish and Hep20 Inoculation and Sample Collection
2.4. RNA Extraction and RT-qPCR
2.5. In Silico Analysis of the Structure of Hep20
2.6. Statistical Analysis
3. Results
3.1. Sequence and Structure Analysis of Fish Hepcidin
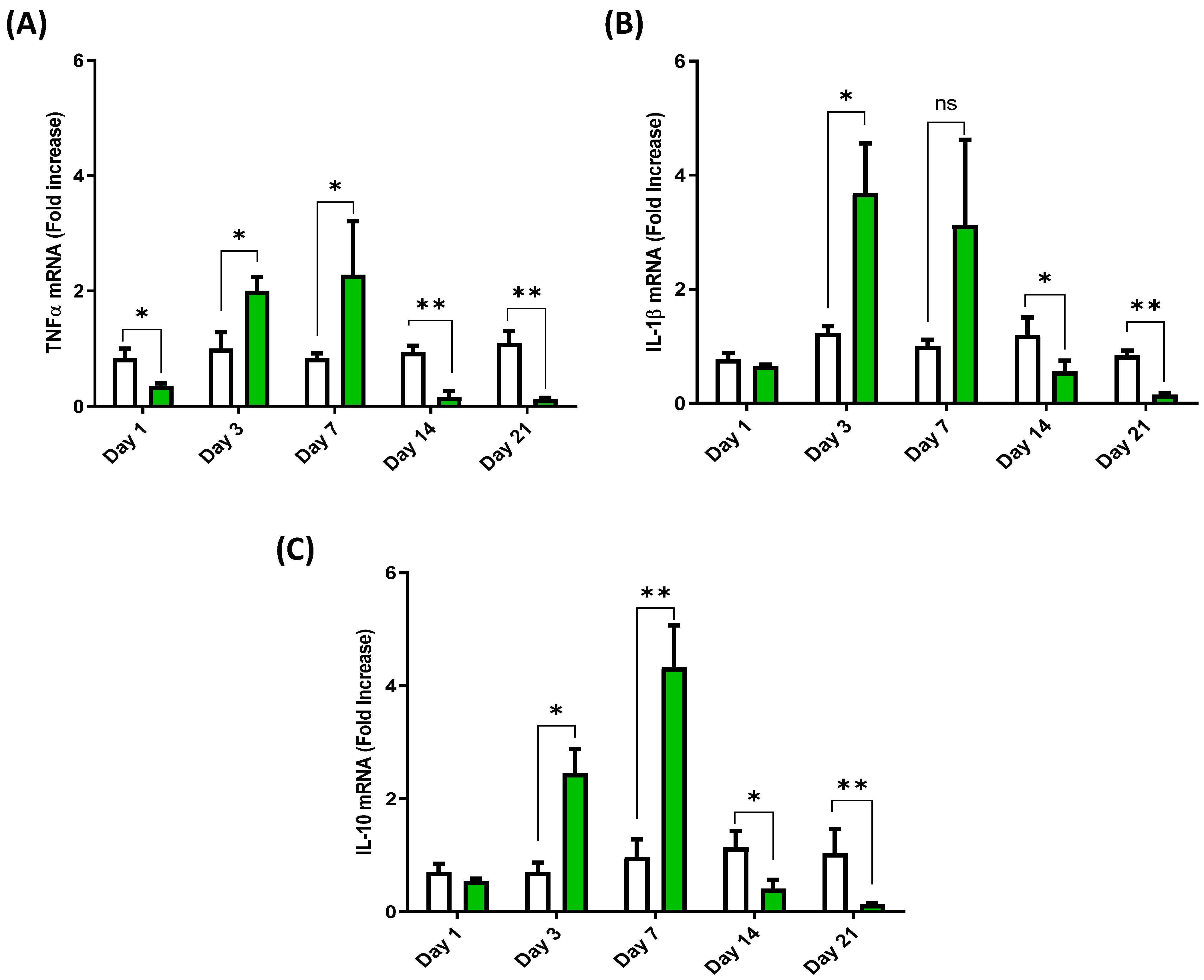
3.2. Cytokine Expression Profile in the Anterior Intestine of European Sea Bass Injected with Hep20
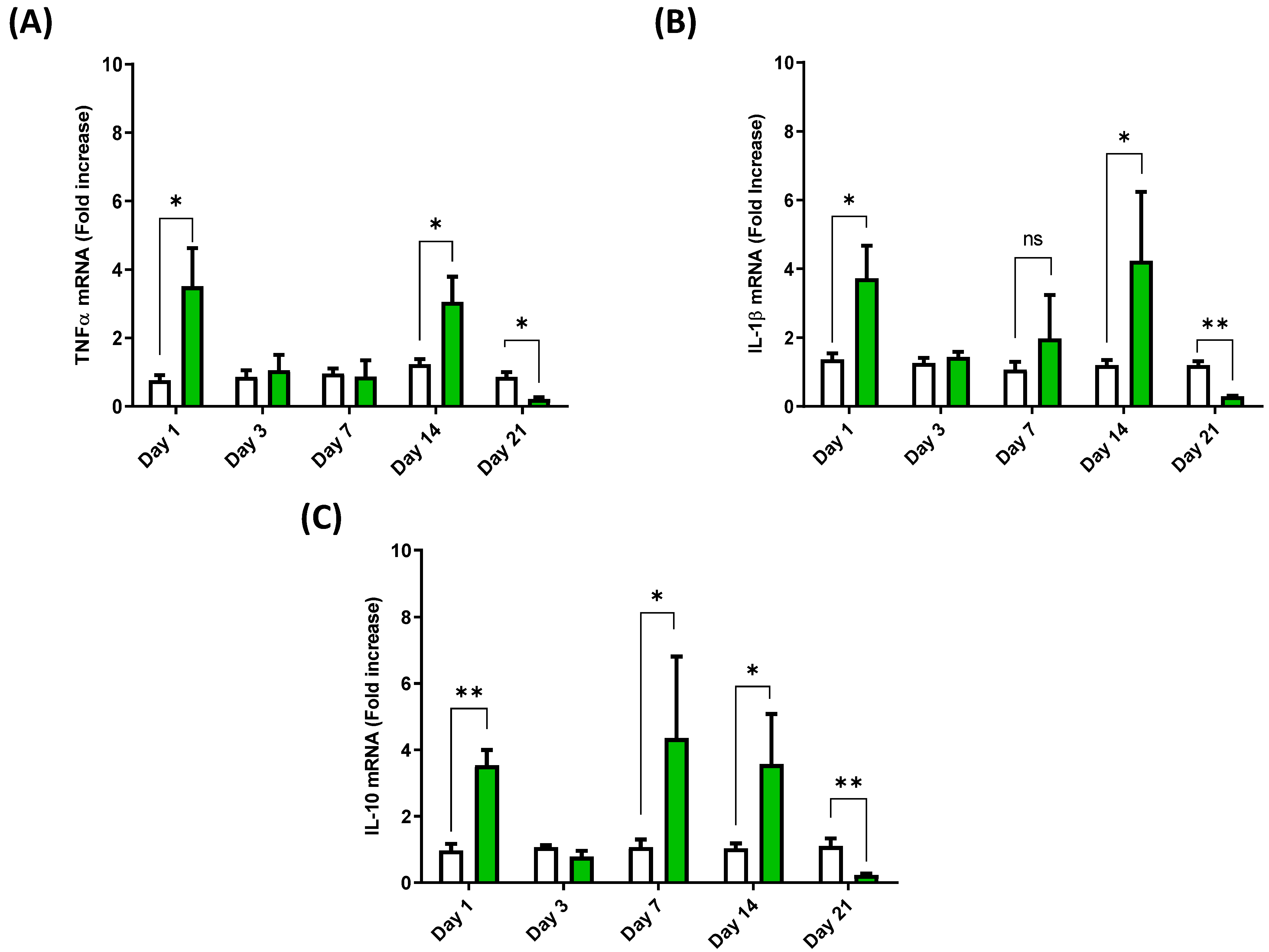
3.3. Cytokine Expression Profile in the Spleen of European Sea Bass Injected with Hep20
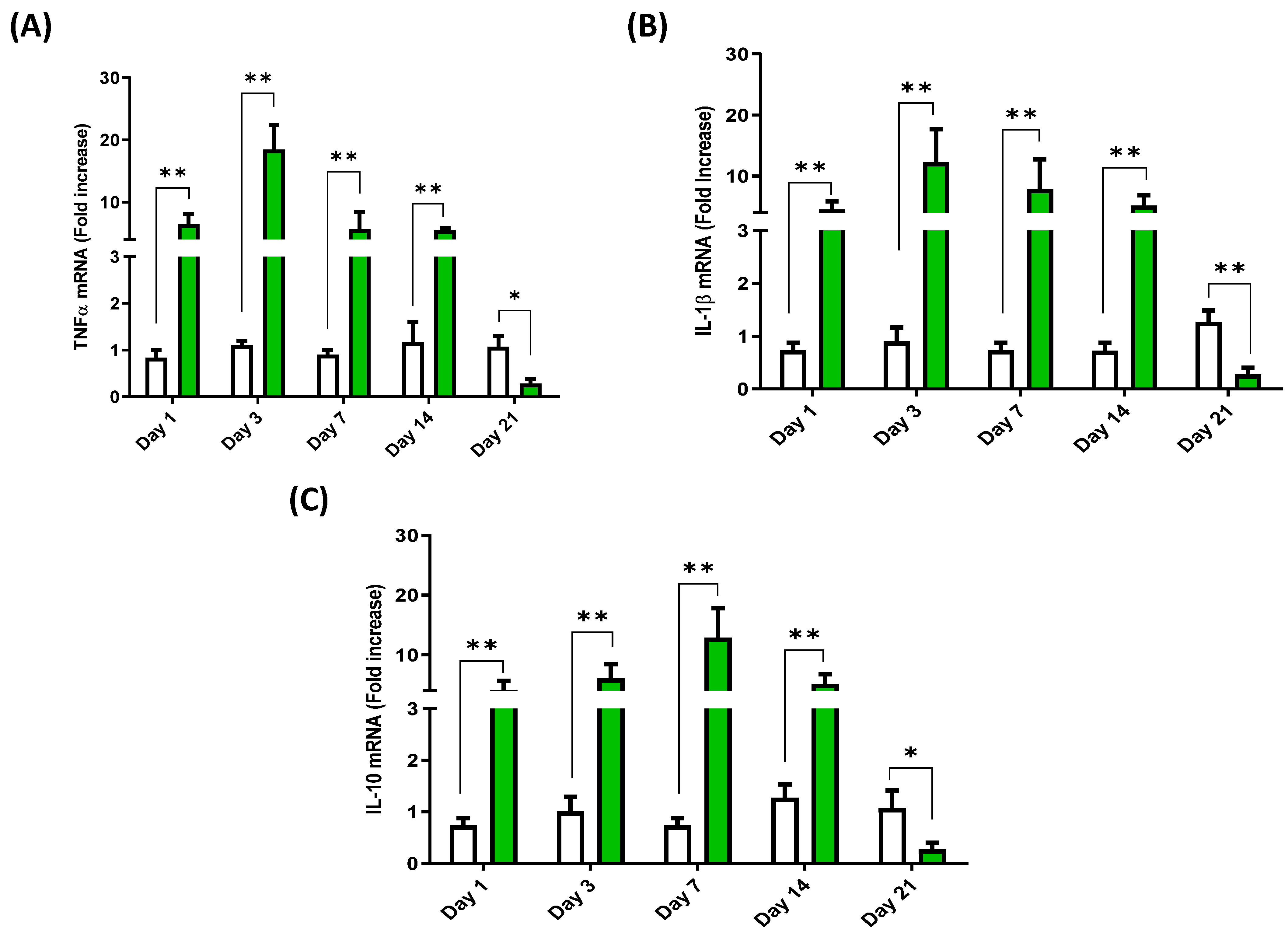
3.4. Cytokine Expression Profile in the Head Kidney of European Sea Bass Injected with Hep20
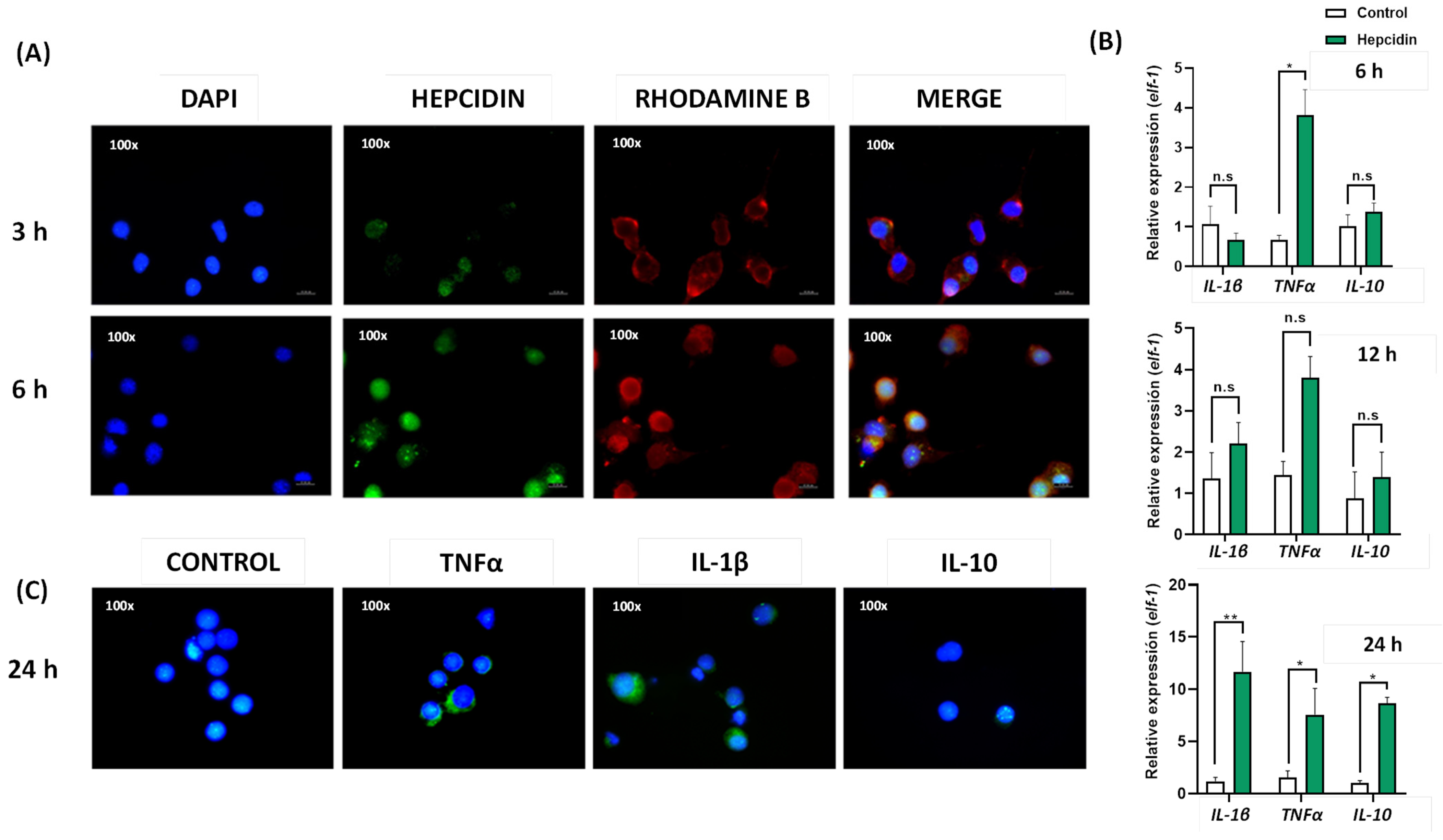
3.5. Cytokine Expression Induced by Hep20 in Cell Culture
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kamysz, W.; Okrój, M.; Łukasiak, J. Novel Properties of Antimicrobial Peptides. Acta Biochim. Pol. 2003, 50, 461–469. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.W.; Haney, E.F.; Gill, E.E. The Immunology of Host Defence Peptides: Beyond Antimicrobial Activity. Nat. Rev. Immunol. 2016, 16, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin Regulates Cellular Iron Efflux by Binding to Ferroportin and Inducing Its Internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armitage, A.E.; Eddowes, L.A.; Gileadi, U.; Cole, S.; Spottiswoode, N.; Selvakumar, T.A.; Ho, L.-P.; Townsend, A.R.M.; Drakesmith, H. Hepcidin Regulation by Innate Immune and Infectious Stimuli. Blood 2011, 118, 4129–4139. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Obiefuna, V.; Hodgkinson, J.W.; McAllister, M.; Belosevic, M. Teleost Antimicrobial Peptide Hepcidin Contributes to Host Defense of Goldfish (Carassius auratus L.) against Trypanosoma carassii. Dev. Comp. Immunol. 2019, 94, 11–15. [Google Scholar] [CrossRef]
- Barroso, C.; Carvalho, P.; Nunes, M.; Gonçalves, J.F.M.; Rodrigues, P.N.S.; Neves, J.V. The Era of Antimicrobial Peptides: Use of Hepcidins to Prevent or Treat Bacterial Infections and Iron Disorders. Front. Immunol. 2021, 12, 754437. [Google Scholar] [CrossRef]
- Alvarez, C.A.; Guzmán, F.; Cárdenas, C.; Marshall, S.H.; Mercado, L. Antimicrobial Activity of Trout Hepcidin. Fish Shellfish Immunol. 2014, 41, 93–101. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, L.-P.; Li, M.-F.; Sun, L. Turbot (Scophthalmus maximus) Hepcidin-1 and Hepcidin-2 Possess Antimicrobial Activity and Promote Resistance against Bacterial and Viral Infection. Fish Shellfish Immunol. 2014, 38, 127–134. [Google Scholar] [CrossRef]
- Gui, L.; Zhang, P.; Zhang, Q.; Zhang, J. Two Hepcidins from Spotted Scat (Scatophagus argus) Possess Antibacterial and Antiviral Functions in Vitro. Fish Shellfish Immunol. 2016, 50, 191–199. [Google Scholar] [CrossRef]
- Álvarez, C.A.; Gomez, F.A.; Mercado, L.; Ramírez, R.; Marshall, S.H. Piscirickettsia Salmonis Imbalances the Innate Immune Response to Succeed in a Productive Infection in a Salmonid Cell Line Model. PLoS ONE 2016, 11, e0163943. [Google Scholar] [CrossRef] [Green Version]
- Neves, J.V.; Ramos, M.F.; Moreira, A.C.; Silva, T.; Gomes, M.S.; Rodrigues, P.N.S. Hamp1 but Not Hamp2 Regulates Ferroportin in Fish with Two Functionally Distinct Hepcidin Types. Sci. Rep. 2017, 7, 14793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melino, S.; Garlando, L.; Patamia, M.; Paci, M.; Petruzzelli, R. A Metal-Binding Site Is Present in the Amino Terminal Region of the Bioactive Iron Regulator Hepcidin-25. J. Pept. Res. 2005, 66 (Suppl. 1), 65–71. [Google Scholar] [CrossRef]
- Nemeth, E.; Preza, G.C.; Jung, C.-L.; Kaplan, J.; Waring, A.J.; Ganz, T. The N-Terminus of Hepcidin Is Essential for Its Interaction with Ferroportin: Structure-Function Study. Blood 2006, 107, 328–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.-G.; Liu, S.-S.; Sun, B.; Wang, X.-L.; Wang, N.; Chen, S.-L. Iron-Metabolic Function and Potential Antibacterial Role of Hepcidin and Its Correlated Genes (Ferroportin 1 and Transferrin Receptor) in Turbot (Scophthalmus maximus). Fish Shellfish Immunol. 2013, 34, 744–755. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.A.; Santana, P.A.; Guzmán, F.; Marshall, S.; Mercado, L.A. Detection of the Hepcidin Prepropeptide and Mature Peptide in Liver of Rainbow Trout. Dev. Comp. Immunol. 2013, 41, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.A.; Acosta, F.; Montero, D.; Guzmán, F.; Torres, E.; Vega, B.; Mercado, L. Synthetic Hepcidin from Fish: Uptake and Protection against Vibrio anguillarum in Sea Bass (Dicentrarchus labrax). Fish Shellfish Immunol. 2016, 55, 662–670. [Google Scholar] [CrossRef]
- Houghten, R.A. General Method for the Rapid Solid-Phase Synthesis of Large Numbers of Peptides: Specificity of Antigen--Antibody Interaction at the Level of Individual Amino Acids. Proc. Natl. Acad. Sci. USA 1985, 82, 5131–5135. [Google Scholar] [CrossRef] [Green Version]
- Ganassin, R.C.; Bols, N.C. Development of a Monocyte/Macrophage-like Cell Line, RTS11, from Rainbow Trout Spleen. Fish Shellfish Immunol. 1998, 8, 457–476. [Google Scholar] [CrossRef]
- Gómez, F.; Henríquez, V.; Marshall, S. Additional Evidence of the Facultative Intracellular Nature of the Fish Bacterial Pathogen Piscirickettsia Salmonis. Arch. Med. Vet. 2009, 41, 261–267. [Google Scholar] [CrossRef] [Green Version]
- Rojas, V.; Galanti, N.; Bols, N.C.; Marshall, S.H. Productive Infection of Piscirickettsia Salmonis in Macrophages and Monocyte-like Cells from Rainbow Trout, a Possible Survival Strategy. J. Cell. Biochem. 2009, 108, 631–637. [Google Scholar] [CrossRef]
- Schmitt, P.; Wacyk, J.; Morales-Lange, B.; Rojas, V.; Guzmán, F.; Dixon, B.; Mercado, L. Immunomodulatory Effect of Cathelicidins in Response to a β-Glucan in Intestinal Epithelial Cells from Rainbow Trout. Dev. Comp. Immunol. 2015, 51, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Morales-Lange, B.; Djordjevic, B.; Gaudhaman, A.; Press, C.M.; Olson, J.; Mydland, L.T.; Mercado, L.; Imarai, M.; Castex, M.; Øverland, M. Dietary Inclusion of Hydrolyzed Debaryomyces Hansenii Yeasts Modulates Physiological Responses in Plasma and Immune Organs of Atlantic Salmon (Salmo salar) Parr Exposed to Acute Hypoxia Stress. Front. Physiol. 2022, 13, 429. [Google Scholar] [CrossRef] [PubMed]
- Morales-Lange, B.; Agboola, J.O.; Hansen, J.Ø.; Lagos, L.; Øyås, O.; Mercado, L.; Mydland, L.T.; Øverland, M. The Spleen as a Target to Characterize Immunomodulatory Effects of Down-Stream Processed Cyberlindnera Jadinii Yeasts in Atlantic Salmon Exposed to a Dietary Soybean Meal Challenge. Front. Immunol. 2021, 12, 3345. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A Multiple Sequence Alignment Editor and Analysis Workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Bowdish, D.M.E.; Davidson, D.J.; Scott, M.G.; Hancock, R.E.W. Immunomodulatory Activities of Small Host Defense Peptides. Antimicrob. Agents Chemother. 2005, 49, 1727–1732. [Google Scholar] [CrossRef] [Green Version]
- Bowdish, D.M.E.; Davidson, D.J.; Hancock, R.E.W. A Re-Evaluation of the Role of Host Defence Peptides in Mammalian Immunity. Curr. Protein Pept. Sci. 2005, 6, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.W.D.D. (Ed.) Mammalian Host Defence Peptides; Cambridge University Press: New York, NY, USA, 2004. [Google Scholar]
- Galindo-Villegas, J.; Mulero, I.; García-Alcazar, A.; Muñoz, I.; Peñalver-Mellado, M.; Streitenberger, S.; Scapigliati, G.; Meseguer, J.; Mulero, V. Recombinant TNFα as Oral Vaccine Adjuvant Protects European Sea Bass against Vibriosis: Insights into the Role of the CCL25/CCR9 Axis. Fish Shellfish Immunol. 2013, 35, 1260–1271. [Google Scholar] [CrossRef]
- Raida, M.K.; Buchmann, K. Development of Adaptive Immunity in Rainbow Trout, Oncorhynchus Mykiss (Walbaum) Surviving an Infection with Yersinia Ruckeri. Fish Shellfish Immunol. 2008, 25, 533–541. [Google Scholar] [CrossRef]
- Román, L.; Acosta, F.; Padilla, D.; El Aamri, F.; Bravo, J.; Vega, B.; Rodriguez, E.; Vega, J.; Déniz, S.; Real, F. The in Vitro Immunomodulatory Effect of Extracellular Products (ECPs) of Vagococcus fluvialis L21 on European Sea Bass (Dicentrarchus labrax) Leucocytes. Fish Shellfish Immunol. 2015, 42, 517–521. [Google Scholar] [CrossRef]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of Antimicrobial Peptide Action and Resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mookherjee, N.; Hancock, R.E.W. Cationic Host Defence Peptides: Innate Immune Regulatory Peptides as a Novel Approach for Treating Infections. Cell. Mol. Life Sci. 2007, 64, 922–933. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Hancock, R.E.W. Peptide Design for Antimicrobial and Immunomodulatory Applications. Biopolymers 2013, 100, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Hilchie, A.L.; Wuerth, K.; Hancock, R.E.W. Immune Modulation by Multifaceted Cationic Host Defense (Antimicrobial) Peptides. Nat. Chem. Biol. 2013, 9, 761–768. [Google Scholar] [CrossRef]
- Lee, S.-C.; Pan, C.-Y.; Chen, J.-Y. The Antimicrobial Peptide, Epinecidin-1, Mediates Secretion of Cytokines in the Immune Response to Bacterial Infection in Mice. Peptides 2012, 36, 100–108. [Google Scholar] [CrossRef]
- Pan, C.-Y.; Wu, J.-L.; Hui, C.-F.; Lin, C.-H.; Chen, J.-Y. Insights into the Antibacterial and Immunomodulatory Functions of the Antimicrobial Peptide, Epinecidin-1, against Vibrio Vulnificus Infection in Zebrafish. Fish Shellfish Immunol. 2011, 31, 1019–1025. [Google Scholar] [CrossRef]
- Brogden, K.A.; Johnson, G.K.; Vincent, S.D.; Abbasi, T.; Vali, S. Oral Inflammation, a Role for Antimicrobial Peptide Modulation of Cytokine and Chemokine Responses. Expert Rev. Anti. Infect. Ther. 2013, 11, 1097–1113. [Google Scholar] [CrossRef]
- Rehaume, L.M.; Hancock, R.E.W. Neutrophil-Derived Defensins as Modulators of Innate Immune Function. Crit. Rev. Immunol. 2008, 28, 185–200. [Google Scholar] [CrossRef]
- Presicce, P.; Giannelli, S.; Taddeo, A.; Villa, M.L.; Della Bella, S. Human Defensins Activate Monocyte-Derived Dendritic Cells, Promote the Production of Proinflammatory Cytokines, and up-Regulate the Surface Expression of CD91. J. Leukoc. Biol. 2009, 86, 941–948. [Google Scholar] [CrossRef]
- Rajanbabu, V.; Pan, C.-Y.; Lee, S.-C.; Lin, W.-J.; Lin, C.-C.; Li, C.-L.; Chen, J.-Y. Tilapia Hepcidin 2-3 Peptide Modulates Lipopolysaccharide-Induced Cytokines and Inhibits Tumor Necrosis Factor-Alpha through Cyclooxygenase-2 and Phosphodiesterase 4D. J. Biol. Chem. 2010, 285, 30577–30586. [Google Scholar] [CrossRef] [Green Version]
- Prasad, S.V.; Fiedoruk, K.; Daniluk, T.; Piktel, E.; Bucki, R. Expression and Function of Host Defense Peptides at Inflammation Sites. Int. J. Mol. Sci. 2020, 21, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigro, E.; Colavita, I.; Sarnataro, D.; Scudiero, O.; Daniele, A.; Salvatore, F.; Pessi, A. Host Defense Peptide-Derived Privileged Scaffolds for Anti-Infective Drug Discovery. J. Pept. Sci. 2017, 23, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Yount, N.Y.; Waring, A.J.; Gank, K.D.; Welch, W.H.; Kupferwasser, D.; Yeaman, M.R. Structural Correlates of Antimicrobial Efficacy in IL-8 and Related Human Kinocidins. Biochim. Biophys. Acta—Biomembr. 2007, 1768, 598–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigro, E.; Colavita, I.; Sarnataro, D.; Scudiero, O.; Zambrano, G.; Granata, V.; Daniele, A.; Carotenuto, A.; Galdiero, S.; Folliero, V.; et al. An Ancestral Host Defence Peptide within Human β-Defensin 3 Recapitulates the Antibacterial and Antiviral Activity of the Full-Length Molecule. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Álvarez, C.A.; Santana, P.A.; Salinas-Parra, N.; Beltrán, D.; Guzmán, F.; Vega, B.; Acosta, F.; Mercado, L. Immune Modulation Ability of Hepcidin from Teleost Fish. Animals 2022, 12, 1586. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121586
Álvarez CA, Santana PA, Salinas-Parra N, Beltrán D, Guzmán F, Vega B, Acosta F, Mercado L. Immune Modulation Ability of Hepcidin from Teleost Fish. Animals. 2022; 12(12):1586. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121586
Chicago/Turabian StyleÁlvarez, Claudio Andrés, Paula A. Santana, Nicolás Salinas-Parra, Dina Beltrán, Fanny Guzmán, Belinda Vega, Félix Acosta, and Luis Mercado. 2022. "Immune Modulation Ability of Hepcidin from Teleost Fish" Animals 12, no. 12: 1586. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121586