
Comparative Susceptibilities of Selected California Chinook Salmon and Steelhead Populations to Isolates of L Genogroup Infectious Hematopoietic Necrosis Virus (IHNV)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Populations
2.2. Virus
2.3. Experimental Studies of Fish Susceptibility to Mortality
2.4. Temperature
2.5. Viral Persistence Studies
2.6. Statistical Analyses
3. Results
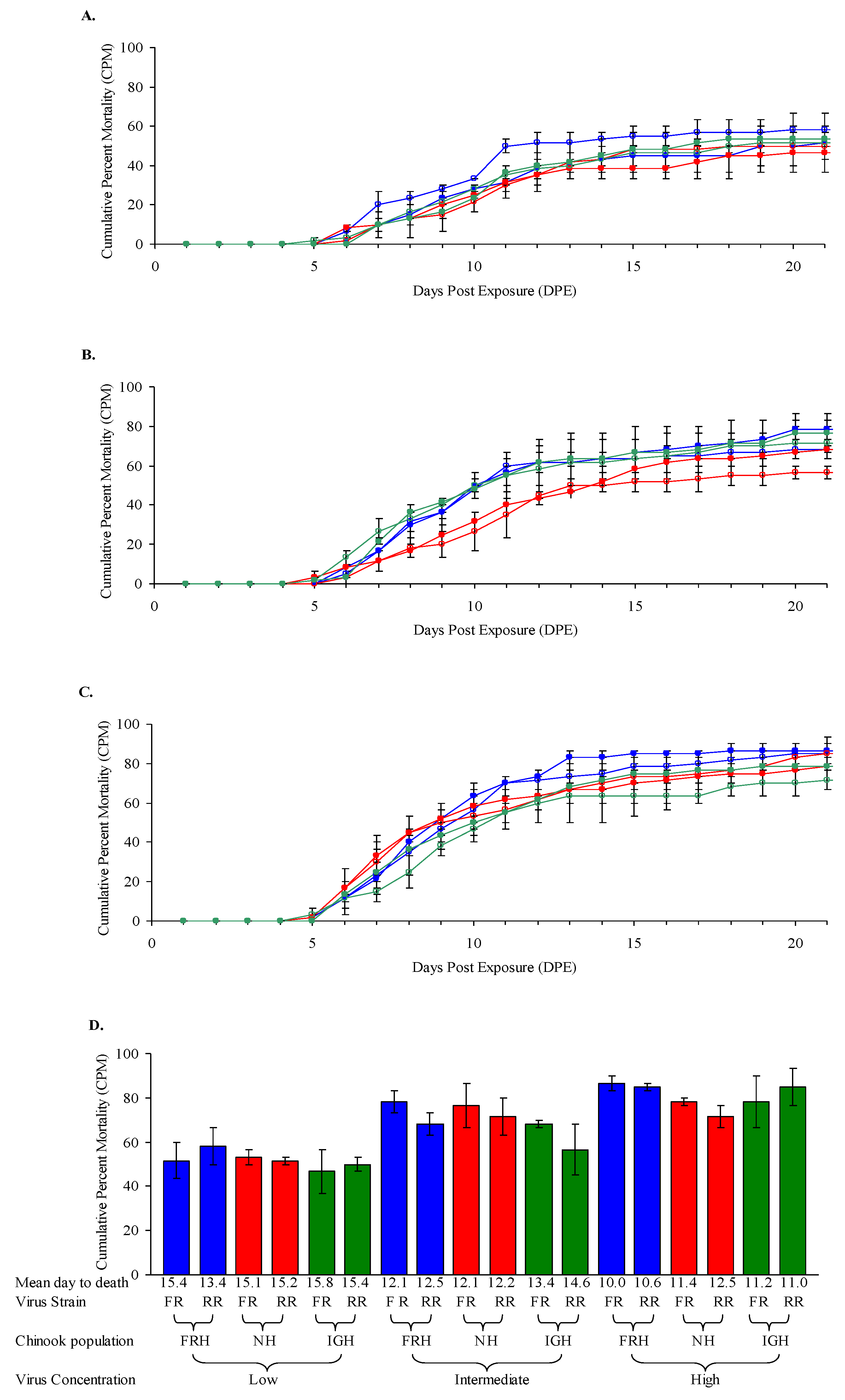
3.1. Chinook Salmon Fry Susceptibility at 12 °C
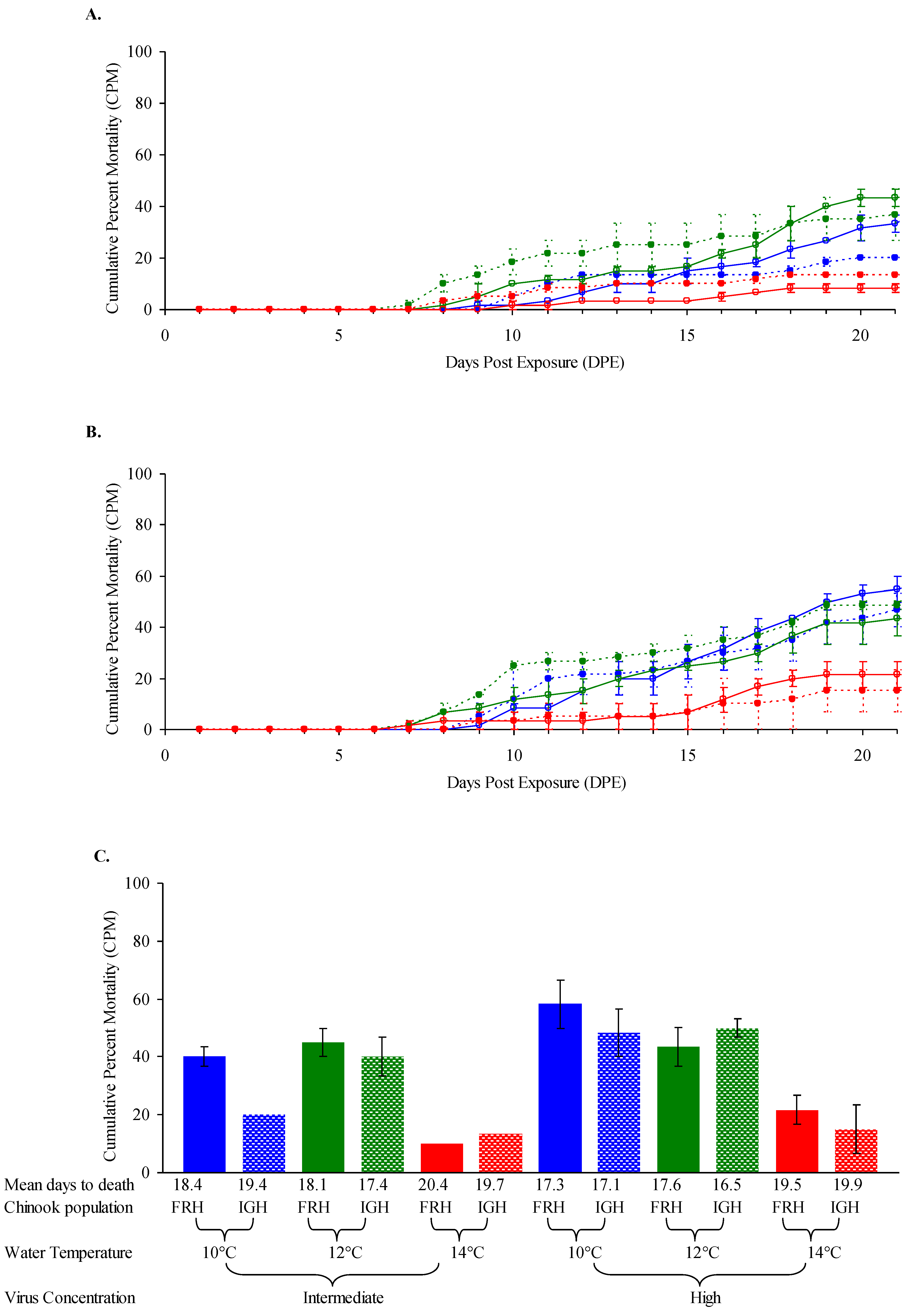
3.2. Chinook Salmon Fingerling Susceptibility at Three Temperatures
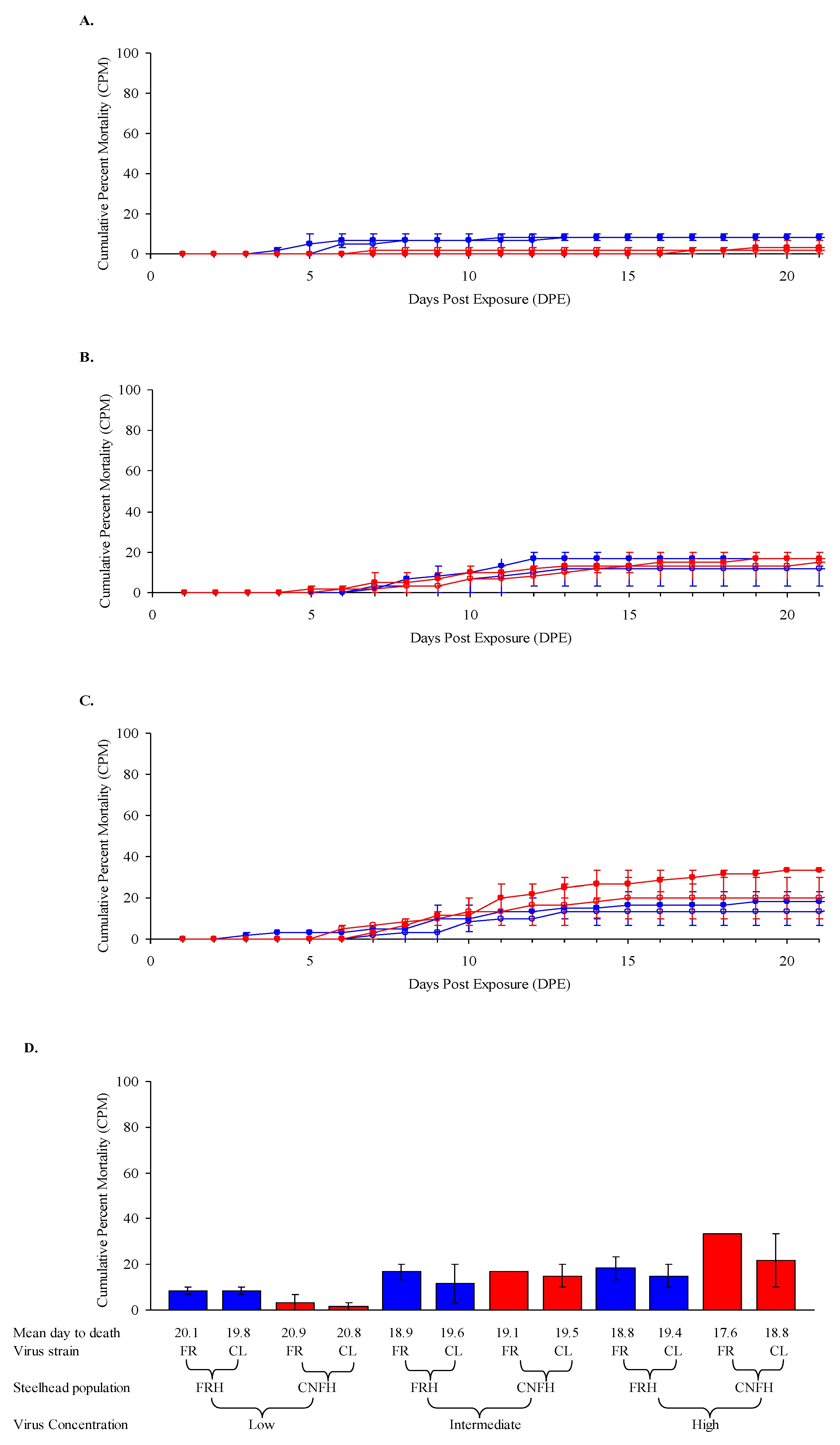
3.3. Steelhead Fry Susceptibility at 12 °C
3.4. Viral Persistence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, I.V.; Amend, D.F. A natural epizootic of infectious hematopoietic necrosis in fry of sockeye salmon (Oncorhynchus nerka) at Chilko Lake, British Columbia. J. Fish. Res. Board Can. 1976, 33, 1564–1567. [Google Scholar] [CrossRef]
- Pilcher, K.S.; Fryer, J.L. The viral diseases of fish, a review through 1978. Part I, Diseases of proven viral etiology. CRC Crit. Rev. Microbiol. 1980, 7, 287–364. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K. Infectious Hematopoietic Necrosis. Fish Viruses and Fish Viral Diseases; Cornell University Press: Ithaca, NY, USA, 1988; pp. 83–114. [Google Scholar]
- Bootland, L.M.; Leong, J.C. Infectious Hematopoietic Necrosis Virus (IHNV). In Fish Diseases and Disorders: Viral, Bacterial, and Fungal Infections; Woo, P.T.K., Bruno, D.W., Eds.; CAB International Publishing: New York, NY, USA, 1999; Volume 3, pp. 57–121. [Google Scholar]
- Winton, J.R. Recent advances in detection and control of infectious hematopoietic necrosis virus in aquaculture. Ann. Rev. Fish Dis. 1991, 1, 83–93. [Google Scholar] [CrossRef]
- LaPatra, S.E.; Groberg, W.J.; Rohovec, J.S.; Fryer, J.L. Size-related susceptibility of salmonids to two strains of infectious hematopoietic necrosis virus. Trans. Am. Fish. Soc. 1990, 119, 25–30. [Google Scholar] [CrossRef]
- LaPatra, S.E.; Groff, J.M.; Fryer, J.L.; Hedrick, R.P. Comparative pathogenesis of three strains of infectious hematopoietic necrosis virus in rainbow trout Oncorhynchus mykiss. Dis. Aquat. Org. 1990, 8, 105–112. [Google Scholar] [CrossRef]
- LaPatra, S.E.; Fryer, J.L.; Rohovec, J.S. Virulence comparison of different electropherotypes of infectious hematopoietic necrosis virus. Dis. Aquat. Org. 1993, 16, 115–120. [Google Scholar] [CrossRef]
- Garver, K.A.; Batts, W.N.; Kurath, G. Virulence comparisons of infectious hematopoietic necrosis virus U and M genogroups in sockeye salmon and rainbow trout. J. Aquat. Anim. Health 2006, 18, 232–243. [Google Scholar] [CrossRef]
- Breyta, R.; Jones, A.; Kurath, G. Differential susceptibility in steelhead trout populations to an emergent MD strain of infectious hematopoietic necrosis virus. Dis. Aquat. Org. 2014, 112, 17–28. [Google Scholar] [CrossRef] [Green Version]
- LaPatra, S.E. Factors affecting pathogenicity of infectious hematopoietic necrosis virus (IHNV) for salmonid fish. J. Aquat. Anim. Health 1998, 10, 121–131. [Google Scholar] [CrossRef]
- Bootland, L.M.; Leong, J.C. Infectious Haematopoietic Necrosis Virus (IHNV). In Fish Diseases and Disorders: Viral, Bacterial, and Fungal Infections; Woo, P.T.K., Leatherland, J.F., Bruno, D.W., Eds.; CAB International Publishing: Poole, UK, 2011; Volume 3, pp. 66–109. [Google Scholar]
- Amend, D.F. Control of infectious hematopoietic necrosis virus disease by elevating the water temperature. J. Fish. Res. Board Can. 1970, 27, 265–270. [Google Scholar] [CrossRef]
- Kurath, G.; Garver, K.A.; Troyer, R.M.; Emmenegger, E.J.; Einer-Jensen, K.; Anderson, E.D. Phylogeography of infectious hematopoietic necrosis virus in North America. J. Gen. Virol. 2003, 84, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Black, A.; Breyta, R.; Bedford, T.; Kurath, G. Geography and host species shape the evolutionary dynamics of U genogroup infectious hematopoietic necrosis virus. Virus Evol. 2016, 2, vew034. [Google Scholar] [CrossRef] [Green Version]
- Breyta, R.; Black, A.; Kaufman, J.; Kurath, G. Spatial and temporal heterogeneity of infectious hematopoietic necrosis virus in Pacific Northwest salmonids. Infect. Genet. Evol. 2016, 45, 347–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garver, K.; Troyer, R.; Kurath, G. Two distinct phylogenetic clades of infectious hematopoietic necrosis virus overlap within the Columbia River basin. Dis. Aquat. Org. 2003, 55, 187–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troyer, R.M.; Kurath, G. Molecular epidemiology of infectious hematopoietic necrosis virus reveals complex virus traffic and evolution within southern Idaho aquaculture. Dis. Aquat. Org. 2003, 55, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Kelley, G.O.; Bendorf, C.M.; Yun, S.C.; Kurath, G.; Hedrick, R.P. Genotypes and phylogeographical relationships of infectious hematopoietic necrosis virus in California, USA. Dis. Aquat. Org. 2007, 77, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Parisot, T.J.; Yasutake, W.T.; Klontz, G. Virus diseases of the Salmonidae in western United States I. Etiology and epizootiology. Ann. N. Y. Acad. Sci. 1965, 24, 52–55. [Google Scholar] [CrossRef]
- Wingfield, W.H.; Chan, L.D. Studies on the Sacramento River chinook disease and its causative agent, a Symposium on Diseases of Fish and Shellfish. In American Fisheries Society, Special Publication 5; Snieszko, S.F., Ed.; American Fisheries Society: Washington, DC, USA, 1970; pp. 307–318. [Google Scholar]
- Troyer, R.M.; LaPatra, S.E.; Kurath, G. Genetic analyses reveal unusually high diversity of infectious haematopoietic necrosis virus in rainbow trout aquaculture. J. Gen. Virol. 2000, 81, 2823–2832. [Google Scholar] [CrossRef]
- Bendorf, C.M.; Kelley, G.O.; Yun, S.C.; Kurath, G.; Andree, K.B.; Hedrick, R.P. Genetic diversity of infectious hematopoietic necrosis virus from Feather River and Lake Oroville, California, and virulence of selected isolates for Chinook salmon and rainbow trout. J. Aquat. Anim. Health 2007, 19, 254–269. [Google Scholar] [CrossRef]
- Peñaranda, M.M.; Purcell, M.K.; Kurath, G. Differential virulence mechanisms of infectious hematopoietic necrosis virus in rainbow trout (Oncorhynchus mykiss) include host entry and virus replication kinetics. J. Gen. Virol. 2009, 90, 2172–2182. [Google Scholar] [CrossRef]
- Hernandez, D.G.; Purcell, M.K.; Friedman, C.S.; Kurath, G. Susceptibility of ocean-and stream-type Chinook salmon to isolates of the L, U, and M genogroups of infectious hematopoietic necrosis virus (IHNV). Dis. Aquat. Org. 2016, 121, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, D.G.; Brown, W.; Naish, K.A.; Kurath, G. Virulence and infectivity of UC, MD, and L strains of infectious hematopoietic necrosis virus (IHNV) in four populations of Columbia River Basin Chinook salmon. Viruses 2021, 13, 701. [Google Scholar] [CrossRef] [PubMed]
- Utter, F.; Miller, G.; Stahl, G.; Teel, D. Genetic population structure of Chinook salmon, Oncorhynchus tshawytscha, in the Pacific northwest. Fish. Bull. 1989, 87, 239–264. [Google Scholar]
- Bartley, D.M.; Gall, G.A.E. Genetic structure and gene flow in Chinook salmon populations of California. Trans. Am. Fish. Soc. 1990, 119, 55–71. [Google Scholar] [CrossRef]
- Gall, G.A.E.; Bartley, D.M.; Brodziak, J.; Gomulkiewicz, R.; Mangel, M. Geographic variation in population genetic structure of Chinook salmon from California and Oregon. Fish. Bull. 1992, 90, 77–100. [Google Scholar]
- Banks, M.A.; Baldwin, B.A.; Hedgecock, D. Research on Chinook salmon stock structure using microsatellite DNA. Bull. Natl. Res. Inst. Aquac. Suppl. 1996, 2, 5–9. [Google Scholar]
- Kim, T.J.; Parker, K.M.; Hedrick, P.W. Major histocompatibility complex differentiation in Sacramento River Chinook salmon. Genetics 1999, 151, 1115–1122. [Google Scholar] [CrossRef]
- Nielsen, J.L. The evolutionary history of steelhead (Oncorhynchus mykiss) along the US Pacific Coast: Developing a conservation strategy using genetic diversity. ICES J. Mar. Sci. 1999, 56, 449–458. [Google Scholar] [CrossRef]
- Banks, M.A.; Rashbrook, V.K.; Calavetta, M.J.; Dean, C.A.; Hedgecock, D. Analysis of microsatellite DNA resolves genetic structure and diversity of Chinook salmon (Oncorhynchus tshawytscha) in California’s Central Valley. Can. J. Fish. Aquat. Sci. 2000, 57, 915–927. [Google Scholar] [CrossRef] [Green Version]
- Hedgecock, D.; Banks, M.A.; Rashbrook, V.K.; Dean, C.A.; Blankenship, S.M. Application of population genetics to conservation of Chinook salmon diversity in the Central Valley. In Contributions to the Biology of Central Valley Salmonids; Brown, R., Ed.; Department of Water Resources: Sacramento, CA, USA, 2001; Volume 1, pp. 45–70. [Google Scholar]
- Williamson, K.S.; May, B. Homogenization of fall-run Chinook salmon gene pools in the Central Valley of California, USA. N. Am. J. Fish. Manag. 2005, 25, 993–1009. [Google Scholar] [CrossRef]
- Nielsen, J.L.; Pavey, S.A.; Wiacek, T.; Williams, I. Genetics of Central Valley O. mykiss populations: Drainage and watershed scale analyses. San Fr. Estuary Watershed Sci. 2005, 3, 3–31. [Google Scholar]
- Foott, S. IHNV Resistance in Steelhead Fry. STT-BCW-04-COL; U.S. Fish and Wildlife Service, California-Nevada Fish Health Center: Anderson, CA, USA, 2004. [Google Scholar]
- Lannan, C.N.; Winton, J.R.; Fryer, J.L. Fish cell lines: Establishment and characterization of nine cell lines from salmonids. In Vitro 1984, 20, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Fijan, N.; Sulimanovic, D.; Bearzotti, M.; Muzinic, D.; Zwillenberg, L.O.; Chilmonczyk, S.; Vautherot, J.F.; de Kinkelin, P. Some properties of the Epithelioma papulosum cyprini (EPC) cell line from carp Cyprinus carpio. Ann. L’Institut Pasteur Virol. 1983, 134E, 207–220. [Google Scholar] [CrossRef]
- Winton, J.R.; Batts, W.; deKinkelin, P.; LeBerre, M.; Bremont, M.; Fijan, N. Current lineages of the epithelioma papulosum cyprini (EPC) cell line are contaminated with fathead minnow, Pimephales promelas, cells. J. Fish Dis. 2010, 33, 701–704. [Google Scholar] [CrossRef]
- LaPatra, S.E. General procedures for virology AFS-FHS (American Fisheries Society-Fish Health Section). In FHS BLUE book: Suggested Procedures for the Detection and Identification of Certain Finfish and Shellfish Pathogens; AFS-FHS: Bethesda, MD, USA, 2004. [Google Scholar]
- Arkush, K.D.; Bovo, G.; de Kinkelin, P.; Winton, J.R.; Wingfield, W.H.; Hedrick, R.P. Biochemical and antigenic properties of the first isolates of infectious hematopoietic necrosis virus from salmonid fish in Europe. J. Aquat. Anim. Health 1989, 1, 148–153. [Google Scholar] [CrossRef]
- Wertheimer, A.C.; Winton, J.R. Differences in Susceptibility among Three Stocks of Chinook Salmon, Oncorhynchus Tshawytscha to Two Isolates of Infectious Hematopoietic Necrosis Virus, Auke Bay, AK (U.S.A.); NOAA/National Marine Fisheries Service: Silver Spring, MD, USA, 1982; pp. 1–15. [Google Scholar]
- Bergmann, S.M.; Fichtner, D.; Skall, H.F.; Schlotfeldt, H.J.; Olesen, N.J. Age- and weight-dependent susceptibility of rainbow trout Oncorhynchus mykiss to isolates of infectious haematopoietic necrosis virus (IHNV) of varying virulence. Dis. Aquat. Org. 2003, 55, 205–210. [Google Scholar] [CrossRef]
- Quillet, E.; Dorson, M.; Le Guillou, S.; Benmansour, A.; Boudinot, P. Wide range of susceptibility to rhabdoviruses in homozygous clones of rainbow trout. Fish Shellfish Immunol. 2007, 22, 510–519. [Google Scholar] [CrossRef]
- Parisot, T.J.; Pelnar, J. An interim report on Sacramento River Chinook disease: A virus-like disease of Chinook salmon. Progress. Fish-Cult. 1962, 24, 51–55. [Google Scholar] [CrossRef]
- Foott, J.S.; Free, D.; McDowell, T.; Arkush, K.D.; Hedrick, R.P. Infectious hematopoietic necrosis virus transmission and disease among juvenile Chinook salmon exposed in culture compared to environmentally relevant conditions. San Fr. Estuary Watershed Sci. 2006, 4, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.F.; Aikens, C.M.; Fryer, J.L.; Rohovec, J.S. Virulence of four isolates of infectious hematopoietic necrosis virus in salmonid fishes and comparative replication in salmonid fish cell lines. Calif. Fish Game 1990, 76, 137–145. [Google Scholar]
- Ma, J.; Bruce, T.J.; Oliver, L.P.; Cain, K.D. Co-infection of rainbow trout (Oncorhynchus mykiss) with infectious hematopoietic necrosis virus and Flavobacterium psychrophilum. J. Fish Dis. 2019, 42, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Watson, S.W.; Guenther, W.; Rucker, R.R. A Virus Disease of Sockeye Salmon: Interim Report; U.S. Fish and Wildlife Service Special Scientific Report; U.S. Department of Interior, Fish and Wildlife Service: Bailey’s Crossroads, VA, USA, 1954; Volume 138, p. 70160618. [Google Scholar]
- Amend, D.F.; Smith, L. Pathophysiology of infectious hematopoietic necrosis virus disease in rainbow trout: Hematological and blood chemical changes in moribund fish. Infect. Immun. 1975, 11, 171–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulcahy, D.; Pascho, R.J.; Jenes, C.K. Titre distribution patterns of infectious haematopoietic necrosis virus in ovarian fluids of hatchery and feral salmon populations. J. Fish Dis. 1983, 6, 183–188. [Google Scholar] [CrossRef]
- Meyers, T.R.; Thomas, J.B.; Follett, J.E.; Saft, R.R. Infectious hematopoietic necrosis virus: Trends in prevalence and the risk management approach in Alaskan sockeye salmon culture. J. Aquat. Anim. Health 1990, 2, 85–98. [Google Scholar] [CrossRef]
- Drolet, B.S.; Chiou, P.P.; Heidel, J.; Leong, J.A. Detection of truncated virus particles in a persistent RNA virus infection in vivo. J. Virol. 1995, 69, 2140–2147. [Google Scholar] [CrossRef] [Green Version]
- Hattenberger-Baudouy, A.M.; Danton, M.; Merle, G.; Torchy, C.; de Kinkelin, P. Serological evidence of infectious hematopoietic necrosis in rainbow trout from a French outbreak of disease. J. Aquat. Anim. Health 1989, 1, 126–134. [Google Scholar] [CrossRef]
- LaPatra, S.E.; Turner, T.; Lauda, K.A.; Jones, G.R.; Walker, S. Characterization of the humoral response of rainbow trout to infectious hematopoietic necrosis virus. J. Aquat. Anim. Health 1993, 5, 165–171. [Google Scholar] [CrossRef]
- St Hilaire, S.; Ribble, C.; Traxler, G.; Davies, T.; Kent, M.L. Evidence for a carrier state of infectious hematopoietic necrosis virus in chinook salmon Oncorhynchus tshawytscha. Dis. Aquat. Org. 2001, 46, 173–179. [Google Scholar] [CrossRef]
- Amend, D.F. Detection and transmission of infectious hematopoietic necrosis virus in rainbow trout. J. Wildl. Dis. 1975, 11, 471–478. [Google Scholar] [CrossRef]
- Harmache, A.; LeBerre, M.; Droineau, S.; Giovannini, M.; Bremont, M. Bioluminescence imaging of live infected salmonids reveals that the fin bases are the major portal of entry for Novirhabdovirus. J. Virol. 2006, 80, 3655–3659. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Fish Species a | Origin of Population | Geographic Region (Sub-Region) | L Subgroup Range b | History of IHNV Epidemics |
|---|---|---|---|---|
| Chinook salmon | Feather River Hatchery (FHR) | Sacramento River basin (Feather River) | LII | Yes |
| Chinook salmon | Nimbus Hatchery (NH) | Sacramento River Basin (American River) | LII | Yes |
| Chinook salmon | Irongate Hatchery (IGH) | North Coast California (Klamath River) | LI | No |
| Steelhead trout | Feather River Hatchery (FHR) | Sacramento River Basin (Feather River) | LII | Yes |
| Steelhead trout | Coleman National Fish Hatchery (CNFH) | Sacramento River Basin (Battle Creek) | LII | No c |
| IHNV Isolate | IHNV Isolate Full Name | Origin of Isolate a | Year of Isolation | L Subgroup b | Sequence Type b | Sero-Group |
|---|---|---|---|---|---|---|
| FR04 | FR04CA73-13 | FRH Chinook salmon Adult | 2004 | LII | mG011L | 3 |
| RR98 | RR98CA | RR wild Chinook salmon Adult | 1998 | LI | mG013L | 1 |
| CL02 | CL02CA144K1 | CNFH Chinook salmon Adult | 2002 | LII | mG012L | 3 |
| FR92 | FR92CA61-16 | FRH Chinook salmon Adult | 1992 | LII | mG014L | 2 |
| FR69 | FR69Clot119 | FRH Chinook salmon | 1969 | LI | mG2107L | 1 |
| Exp’t | Fish Species (Populations) a | Life Stage, Ave. Weight | Virus Isolate(s) (L Subgroup) b | Challenge Dose(s) c | Temp(s) |
|---|---|---|---|---|---|
| 1 | Chinook salmon (FRH, NH, IGH) | Fry, 0.54–0.57 g | FR04 (LII), RR98 (LI) | low, intermed., high | 12 °C |
| 2 | Chinook salmon (FRH, IGH) | Fingerlings, 2.2 g | FR04 (LII) | intermed., high | 10, 12, 14 °C |
| 3 | Steelhead trout (FRH, CNFH) | Fry, 0.67, 0.68 g | FR04 (LII), CL02 (LII) | low, intermed., high | 12 °C |
| Fish Species (Populations) a | Life Stage, Ave. Weight at Challenge | Virus Exposure a | 120 dpe GKS b | 215–216 dpe GKS + Skin | 227–248 dpe GKS + Skin |
|---|---|---|---|---|---|
| Chinook salmon (NH, IGH) | Fingerlings, 5.1, 4.2 g | FR04 (LII), Low dose | 0/60 | 2/32 | Ns c |
| Mock | 0/60 | 0/30 | ns | ||
| Steelhead trout (FRH) | Fry, fingerlings 0.67 g, 1.1 g | FR04, CL02, FR92, FR69 Low/High dose | ns | ns | 0/163 |
| Mock | ns | ns | 0/45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bendorf, C.M.; Yun, S.C.; Kurath, G.; Hedrick, R.P. Comparative Susceptibilities of Selected California Chinook Salmon and Steelhead Populations to Isolates of L Genogroup Infectious Hematopoietic Necrosis Virus (IHNV). Animals 2022, 12, 1733. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12131733
Bendorf CM, Yun SC, Kurath G, Hedrick RP. Comparative Susceptibilities of Selected California Chinook Salmon and Steelhead Populations to Isolates of L Genogroup Infectious Hematopoietic Necrosis Virus (IHNV). Animals. 2022; 12(13):1733. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12131733
Chicago/Turabian StyleBendorf, Christin M., Susan C. Yun, Gael Kurath, and Ronald P. Hedrick. 2022. "Comparative Susceptibilities of Selected California Chinook Salmon and Steelhead Populations to Isolates of L Genogroup Infectious Hematopoietic Necrosis Virus (IHNV)" Animals 12, no. 13: 1733. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12131733