
Minor Sea Turtle Nesting Areas May Remain Unnoticed without Specific Monitoring: The Case of the Largest Mediterranean Island (Sicily, Italy)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Data Analysis
3. Results
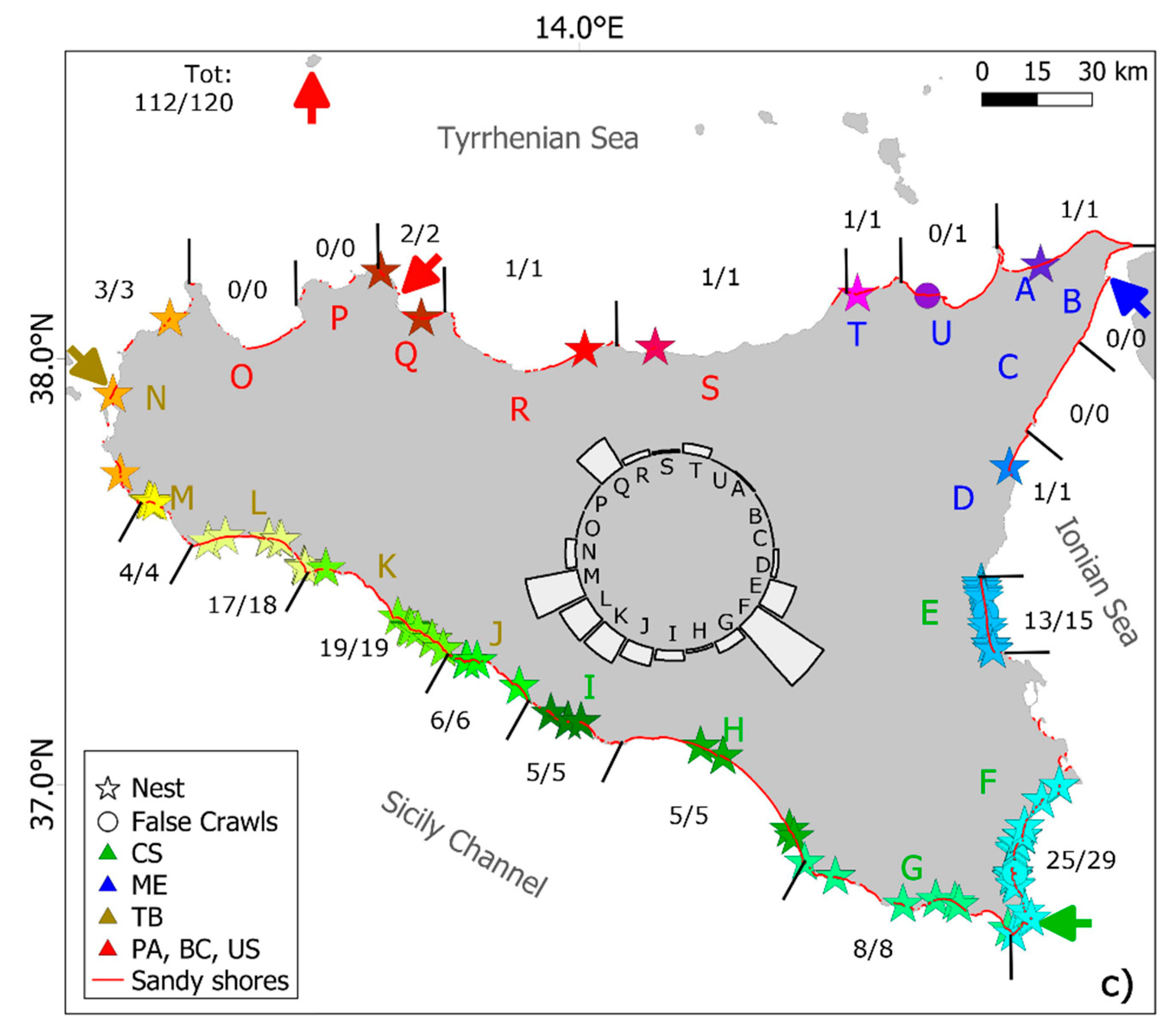
3.1. Spatio-Temporal Distribution of Sea Turtle Nesting Activity
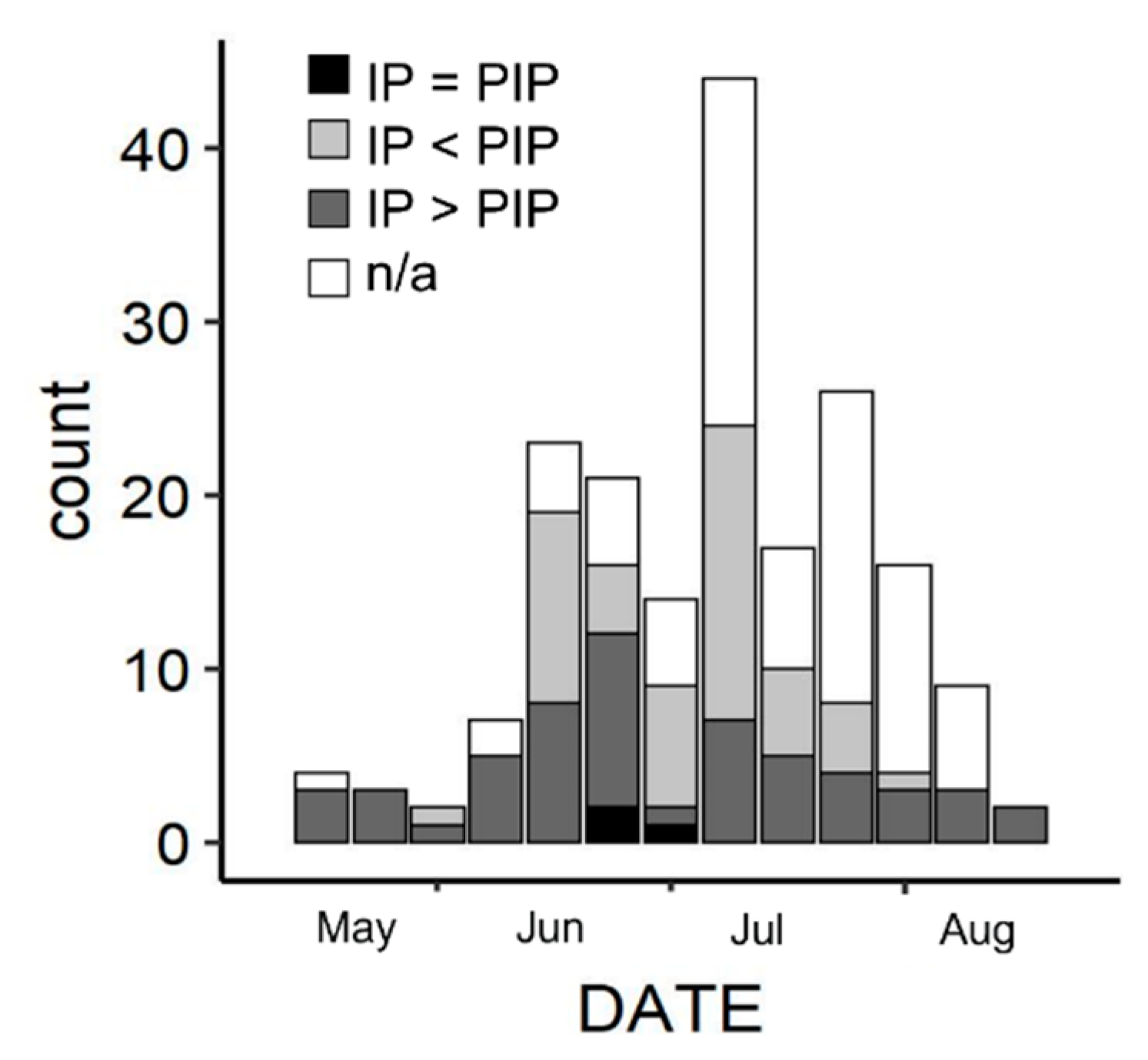
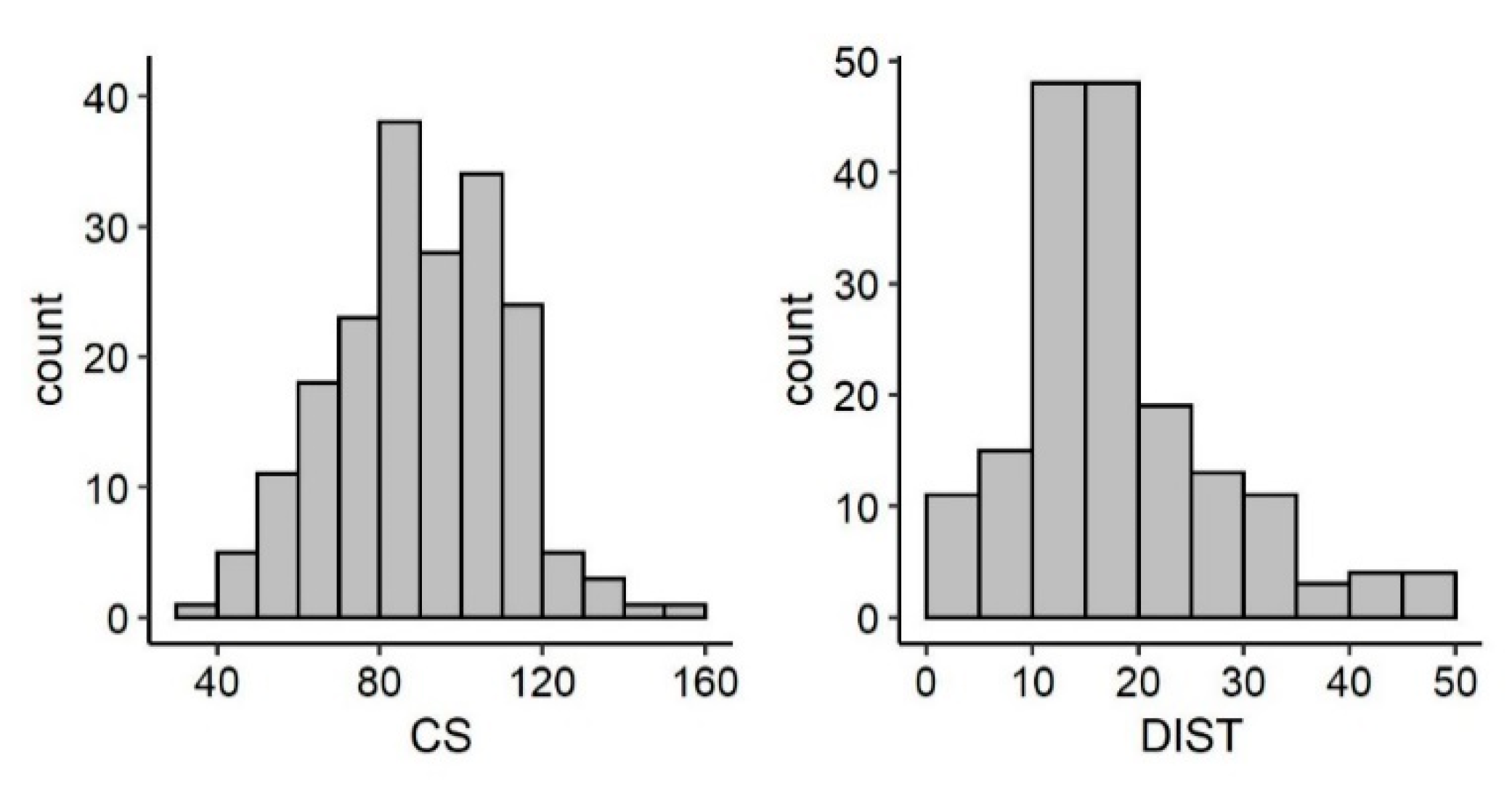
3.2. Reproductive Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, J.D. Reproduction in sea turtles. In The Biology of Sea Turtles; Lutz, P.L., Musick, J.A., Eds.; CRC Marine Science Series; CRC Press, Inc.: Boca Raton, FL, USA, 1997; pp. 51–81. [Google Scholar]
- Fuentes, M.M.P.B.; Gredzens, C.; Bateman, B.L.; Boettcher, R.; Ceriani, S.A.; Godfrey, M.H.; Helmers, D.; Ingram, D.K.; Kamrowski, R.L.; Pate, M.; et al. Conservation hotspots for marine turtle nesting in the United States based on coastal development. Ecol. Appl. 2016, 26, 2706–2717. [Google Scholar] [CrossRef] [PubMed]
- Casale, P.; Broderick, A.C.; Camiñas, J.A.; Cardona, L.; Carreras, C.; Demetropoulos, A.; Fuller, W.J.; Godley, B.J.; Hochscheid, S.; Kaska, Y.; et al. Mediterranean sea turtles: Current knowledge and priorities for conservation and research. Endang. Species Res. 2018, 36, 229–267. [Google Scholar] [CrossRef] [Green Version]
- Durrieu de Madron, X.; Guieu, C.; Sempéré, R.; Conan, P.; Cossa, D.; D’Ortenzio, F.; Estournel, C.; Gazeau, F.; Rabouille, C.; Stemmann, L.; et al. Marine ecosystems’ responses to climatic and anthropogenic forcings in the Mediterranean. Prog. Oceanogr. 2011, 91, 97–166. [Google Scholar] [CrossRef]
- Witt, M.J.; Hawkes, L.A.; Godfrey, M.H.; Godley, B.J.; Broderick, A.C. Predicting the impacts of climate change on a globally distributed species: The case of the loggerhead turtle. J. Exp. Biol. 2010, 213, 901–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pike, D.A. Forecasting the viability of sea turtle eggs in a warming world. Glob. Chang. Biol. 2014, 20, 7–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, A.F.; Saad, A.; Jony, M. Discovery of a regionally important green turtle Chelonia mydas rookery in Syria. Oryx 2008, 42, 456–459. [Google Scholar] [CrossRef] [Green Version]
- Mingozzi, T.; Masciari, G.; Paolillo, G.; Pisani, B.; Russo, M.; Massolo, A. Discovery of a regular nesting area of loggerhead turtle Caretta caretta in southern Italy: A new perspective for national conservation. Biodivers. Conserv. 2007, 16, 3519–3541. [Google Scholar] [CrossRef]
- Casale, P.; Palilla, G.; Salemi, A.; Napoli, A.; Prinzi, M.; Genco, L.; Bonaviri, D.; Mastrogiacomo, A.; Oliverio, M.; Lo Valvo, M. Exceptional sea turtle nest records in 2011 suggest an underestimated nesting potential in Sicily (Italy). Acta Herpetol. 2012, 7, 181–188. [Google Scholar]
- Ceriani, S.A.; Brost, B.; Meylan, A.B.; Meylan, P.A.; Casale, P. Bias in sea turtle productivity estimates: Error and factors involved. Mar. Biol. 2021, 168, 41. [Google Scholar] [CrossRef]
- Team, Q.D. QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2022. Available online: http://qgis.org (accessed on 1 March 2022).
- Hays, G.C.; Ashworth, J.S.; Barnsley, M.J.; Broderick, A.C.; Emery, D.R.; Godley, B.J.; Henwood, A.; Jones, E.L. The importance of sand albedo for the thermal conditions on sea turtle nesting beaches. Oikos 2001, 93, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. GLM and GAM for absence–presence and proportional data. In Mixed Effects Models and Extensions in Ecology with R.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 245–259. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Fuller, W.J.; Godley, B.J.; Hodgson, D.J.; Reece, S.E.; Witt, M.J.; Broderick, A.C. Importance of spatio-temporal data for predicting the effects of climate change on marine turtle sex ratios. Mar. Ecol. Prog. Ser. 2013, 488, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Maffucci, F.; Corrado, R.; Palatella, L.; Borra, M.; Marullo, S.; Hochscheid, S.; Lacorata, G.; Iudicone, D. Seasonal heterogeneity of ocean warming: A mortality sink for ectotherm colonizers. Sci. Rep. 2016, 6, 23983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carreras, C.; Pascual, M.; Tomás, J.; Marco, A.; Hochsheid, S.; Castillo, J.J.; Gozalbes, P.; Parga, M.; Piovano, S.; Cardona, L. Sporadic nesting reveals long distance colonisation in the philopatric loggerhead sea turtle (Caretta caretta). Sci. Rep. 2018, 8, 1435. [Google Scholar] [CrossRef] [Green Version]
- Sönmez, B.; Elginöz, E.; Ilgaz, M.; Altınkaya, H. Nesting activity of loggerhead turtles (2013–2020) and 20 years abundance trend (2001–2020) on Çıralı Beach, Turkey. Reg. Stud. Mar. Sci. 2021, 44, 101758. [Google Scholar] [CrossRef]
- Bohm, E.; Magazzu, G.; Wald, L.; Zoccolotti, M. Coastal currents on the Sicilian shelf south of Messina. Oceanol. Acta 1987, 10, 137–142. [Google Scholar]
- Yakimov, M.M.; Denaro, R.; Genovese, M.; Cappello, S.; D’Auria, G.; Chernikova, T.N.; Timmis, K.N.; Golyshin, P.N.; Giluliano, L. Natural microbial diversity in superficial sediments of Milazzo Harbor (Sicily) and community successions during microcosm enrichment with various hydrocarbons. Environ. Microbiol. 2005, 7, 1426–1441. [Google Scholar] [CrossRef]
- Dimitriadis, C.; Fournari-Konstantinidou, I.; Sourbès, L.; Koutsoubas, D.; Mazaris, A.D. Reduction of sea turtle population recruitment caused by nightlight: Evidence from the Mediterranean region. Ocean. Coast. Manag. 2018, 153, 108–115. [Google Scholar] [CrossRef]
- Lorne, J.K.; Salmon, M. Effects of exposure to artificial lighting on orientation of hatchling sea turtles on the beach and in the ocean. Endang. Species Res. 2007, 3, 23–30. [Google Scholar] [CrossRef]
- Girondot, M.; Monsinjon, J.; Guillon, J.-M. Delimitation of the embryonic thermosensitive period for sex determination using an embryo growth model reveals a potential bias for sex ratio prediction in turtles. J. Therm. Biol. 2018, 73, 32–40. [Google Scholar] [CrossRef]
- Wibbels, T. Critical approaches to sex determination in sea turtles. In The Biology of Sea Turtles. Volume II.; Lutz, P.L., Musick, J.A., Wyneken, J., Eds.; CRC Marine Biology Series; CRC Press, Inc.: Boca Raton, FL, USA, 2003; pp. 103–134. [Google Scholar]
- Katselidis, K.A.; Schofield, G.; Stamou, G.; Dimopoulos, P.; Pantis, J.D. Females first? Past, present and future variability in offspring sex ratio at a temperate sea turtle breeding area. Anim. Conserv. 2012, 15, 508–518. [Google Scholar] [CrossRef]
- Yntema, C.L.; Mrosovsky, N. Critical periods and pivotal temperatures for sexual differentiation in loggerhead sea turtles. Can. J. Zool. 1982, 60, 1012–1016. [Google Scholar] [CrossRef]
- Lolavar, A.; Wyneken, J. Effect of rainfall on loggerhead turtle nest temperatures, sand temperatures and hatchling sex. Endang. Species Res. 2015, 28, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Godley, B.J.; Broderick, A.C.; Downie, J.R.; Glen, F.; Houghton, J.D.R.; Kirkwood, I.; Reece, S.; Hays, G.C. Thermal conditions in nests of loggerhead turtles: Further evidence suggesting female skewed sex ratios of hatchling production in the Mediterranean. J. Exp. Mar. Biol. Ecol. 2001, 263, 45–63. [Google Scholar] [CrossRef]
- Broderick, A.C.; Godley, B.J. Population and nesting ecology of the green turtle, Chelonia mydas, and the loggerhead turtle, Caretta caretta, in northern Cyprus. Zool. Middle East 1996, 13, 27–46. [Google Scholar] [CrossRef]
- Margaritoulis, D.; Rees, A.F.; Dean, C.J.; Riggall, T. Reproductive data of loggerhead turtles in Laganas Bay, Zakynthos Island, Greece, 2003–2009. Mar. Turtle Newsl. 2011, 131, 2–6. [Google Scholar]
- Margaritoulis, D. Nesting activity and reproductive output of loggerhead sea turtles, Caretta caretta, over 19 seasons (1984–2002) at Laganas Bay, Zakynthos, Greece: The largest rookery in the Mediterranean. Chelonian Conserv. Biol. 2005, 4, 916–929. [Google Scholar]
- Foley, A.M.; Peck, S.A.; Harman, G.R. Effects of sand characteristics and inundation on the hatching success of loggerhead sea turtle (Caretta caretta) clutches on low-relief mangrove islands in southwest Florida. Chelonian Conserv. Biol. 2006, 5, 32–41. [Google Scholar] [CrossRef]
- Candan, O. Impact of nest relocation on the reproductive success of Loggerhead Turtles, Caretta caretta, in the Göksu Delta, Turkey (Reptilia: Cheloniidae). Zool. Middle East 2018, 64, 38–46. [Google Scholar] [CrossRef]
- Mrosovsky, N. Against Oversimplifying the Issues on Relocating Turtle Eggs. Environ. Manag. 2008, 41, 465–467. [Google Scholar] [CrossRef]
- Foley, A.M.; Peck, S.A.; Harman, G.R.; Richardson, L.W. Loggerhead turtle (Caretta caretta) nesting habitat on low-relief mangrove islands in southwest Florida and consequences to hatchling sex ratios. Herpetologica 2000, 56, 433–445. [Google Scholar]
- Howard, R.; Bell, I.; Pike, D.A. Thermal tolerances of sea turtle embryos: Current understanding and future directions. Endang. Species Res. 2014, 26, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Ceriani, S.A.; Roth, J.D.; Tucker, A.D.; Evans, D.R.; Addison, D.S.; Sasso, C.R.; Ehrhart, L.M.; Weishampel, J.F. Carry-over effects and foraging ground dynamics of a major loggerhead breeding aggregation. Mar. Biol. 2015, 162, 1955–1968. [Google Scholar] [CrossRef]
- Belfiore, V.; (Legambiente val di noto association, Noto, Siracusa, Italy). Personal communications, 2013.
- Betulla, A.; (Beach Owner, Pachino, Siracusa, Italy). Personal communication, 2016.
- Cavarra, D.; (Beach Owner, Siracusa, Italy). Personal communications, 2016.
- Cilea, F.; (Lipu Association Saline di Priolo, Priolo Gargallo, Siracusa, Italy). Personal communications, 2020.
- Di Rosa, A.; (Wwf Italy, Modica, Ragusa, Italy). Personal Communications, 2017.
- Fabrizi, D.; (Local Citizen, Sampieri, Scicli, Ragusa, Italy). Personal communications, 1944.
- Falesi, L.; (Archeo Club Association Val di noto, Noto, Siracusa, Italy). Personal communications, 2016.
- Galia, F.; (WWF Italy, Licata, Agrigento, Italy). Personal communications, 1994.
- Galia, F.; Freggi, D.; d’Angelo, S.; Lo Valvo, M. An unusual nest activity along southern Sicilian coasts: An hope for sea turtle survival? In Proceedings of the 26th Annual Symposium on Sea Turtle Biology and Conservation, Island of Crete, Greece, 3–8 April 2006; pp. 295–296. [Google Scholar]
- Genco, L.; Culmone, G.; Cortellaro, G.; Federico, R.; Piacentino, A.; Prinzi, M. Prima segnalazione di nidificazione di tartaruga comune Caretta caretta in Sicilia nord-occidentale (Italia meridionale). Atti Soc. It. Sci. Nat. Mus. Civ. Stor. Nat. Milano 2008, 149, 325–330. [Google Scholar]
- Insacco, G.; Inclimona, V.; Barlotta, A.; Ricotta, V.; Denise, G.; Spadola, F.; Blasi, M.F.; Scaravelli, D. The activity of the Sea Turtle rehabilitation in the Regional Rescue Center of Comiso (Sicily, Ragusa, Italy) in 10 years. In Proceedings of the 4th Mediterranean Conference on Marine Turtles, Napoli, Italy, 7–10 November 2011. [Google Scholar]
- Isgrò, C.; (MuMa Museum, Milazzo, Messina, Italy). Personal communications, 2013.
- Isgrò, C.; (MuMa Museum, Milazzo, Messina, Italy). Personal communications, 2019.
- Isgrò, C.; (MuMa Museum, Milazzo, Messina, Italy). Personal communications, 2020.
- Lino, L.; (Pronatura Association, Catania, Italy). Personal communications, 1960.
- Pachino; (Local Citizen, Siracusa, Italy). Personal communications, 2002.
- Scala, S.; (Beach Owner, Portopalo di Capopassero, Siracusa, Italy). Personal communications, 2015.
- Torrisi, A.; (Local Citizen, Catania, Italy). Personal communications, 1987.
- Torrisi, A.; (Local Citizen, Catania, Italy). Personal communications, 2007.
- Zaffarana, R.; (Local Citizen, Avola, Siracusa, Italy). Personal communications, 2013.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Natural | Relocated | ||
|---|---|---|---|---|
| Mean min IP (n; Range; SD) | 57 d (83; 36–80 d; 8.8 d) | 58 d (19; 35–78 d; 10.1 d) | ||
| Mean CS (n; Range; SD) | 92.7 (155; 48–151; 20.4) | 88.0 (23; 50–128; 20.1) | ||
| Mean HS (n; Range; SD) | 63% (155; 0–100%; 36.4%) | 53% (23; 0–96.1%; 32.2%) | ||
| Mean ES (n; Range; SD) | 58.% (155; 0–100%; 35.6%) | 47.9% (23; 0–95.5%; 32.6%) | ||
| Mean DATE (n; Range; SD) | 28.6 weeks (136; 22–33; 2.6) s | 29 weeks (23; 26–34; 2.5) | ||
| Mean IPMT (n; Range; SD) | 28.2 °C (88;24.1–30.3 °C; 1.4 °C) | 28.3 °C (20; 23.8–30.0 °C; 1.8 °C) | ||
| Mean DIST (n; Range; SD) | 20.3 m (140; 0–50 m; 10.2 m) | 13.8 m (22; 2–30 m; 10 m) | ||
| SC | GR | 10 | GR | 1 |
| OC | 130 | OC | 118 | |
| WH | 5 | WH | 1 | |
| YL | 72 | YL | 3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prato, O.O.; Paduano, V.; Baldi, G.; Bonsignore, S.; Callea, G.; Camera, C.; Culmone, G.; D’angelo, S.; Fiorentino, D.; Galia, G.; et al. Minor Sea Turtle Nesting Areas May Remain Unnoticed without Specific Monitoring: The Case of the Largest Mediterranean Island (Sicily, Italy). Animals 2022, 12, 1221. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12091221
Prato OO, Paduano V, Baldi G, Bonsignore S, Callea G, Camera C, Culmone G, D’angelo S, Fiorentino D, Galia G, et al. Minor Sea Turtle Nesting Areas May Remain Unnoticed without Specific Monitoring: The Case of the Largest Mediterranean Island (Sicily, Italy). Animals. 2022; 12(9):1221. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12091221
Chicago/Turabian StylePrato, Oleana Olga, Valentina Paduano, Giulia Baldi, Salvatore Bonsignore, Gerlando Callea, Carlo Camera, Girolamo Culmone, Stefania D’angelo, Diego Fiorentino, Gino Galia, and et al. 2022. "Minor Sea Turtle Nesting Areas May Remain Unnoticed without Specific Monitoring: The Case of the Largest Mediterranean Island (Sicily, Italy)" Animals 12, no. 9: 1221. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12091221