
Prevalence and Genotyping of Anaplasma phagocytophilum Strains from Wild Animals, European Bison (Bison bonasus) and Eurasian Moose (Alces alces) in Poland

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
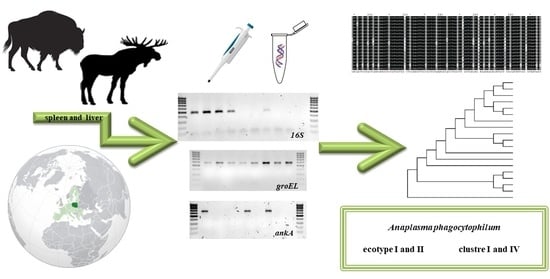
1. Introduction
2. Materials and Methods
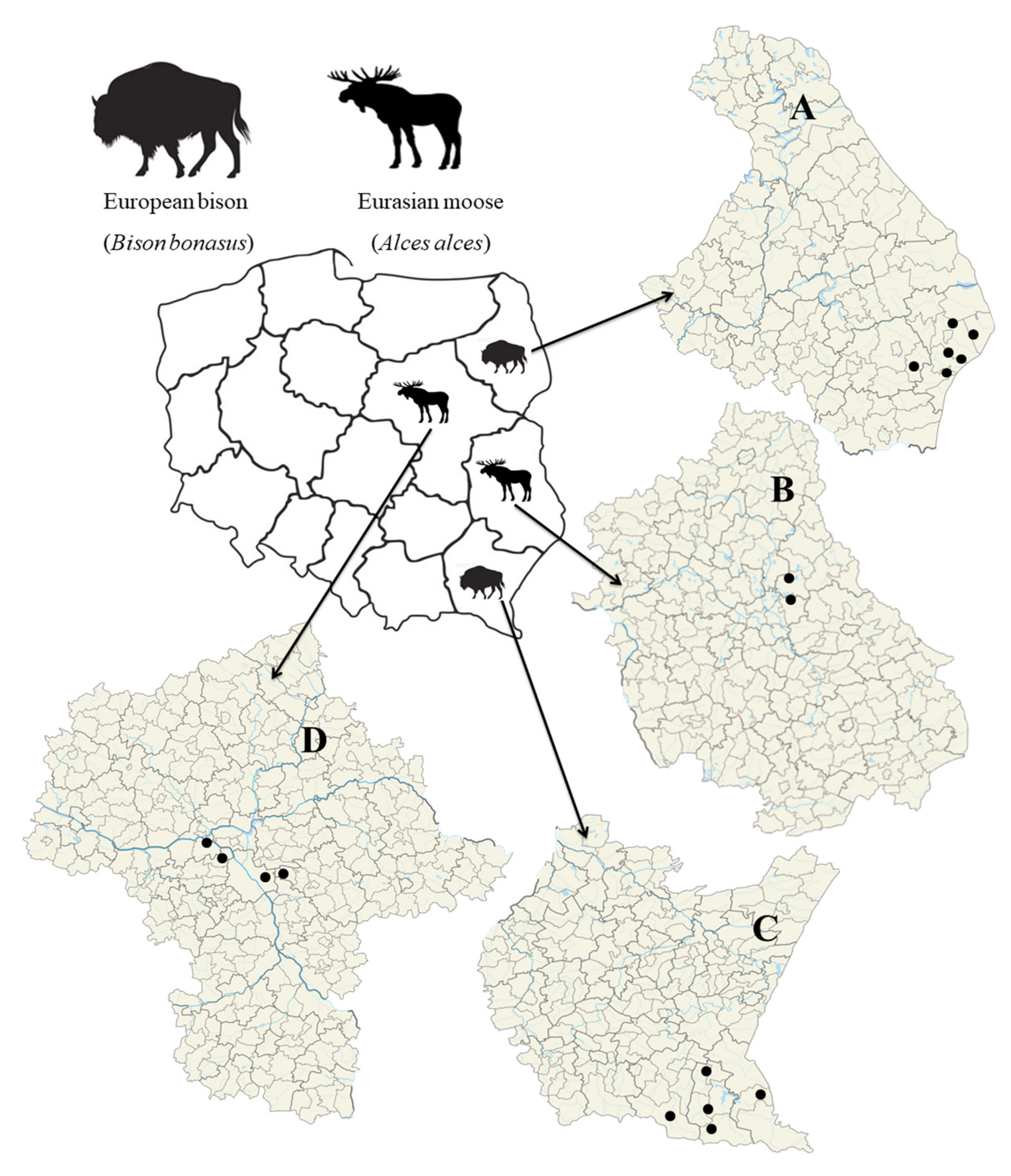
2.1. Materials
2.2. Molecular Methods

2.3. Statistical Methods
3. Results
3.1. Prevalence
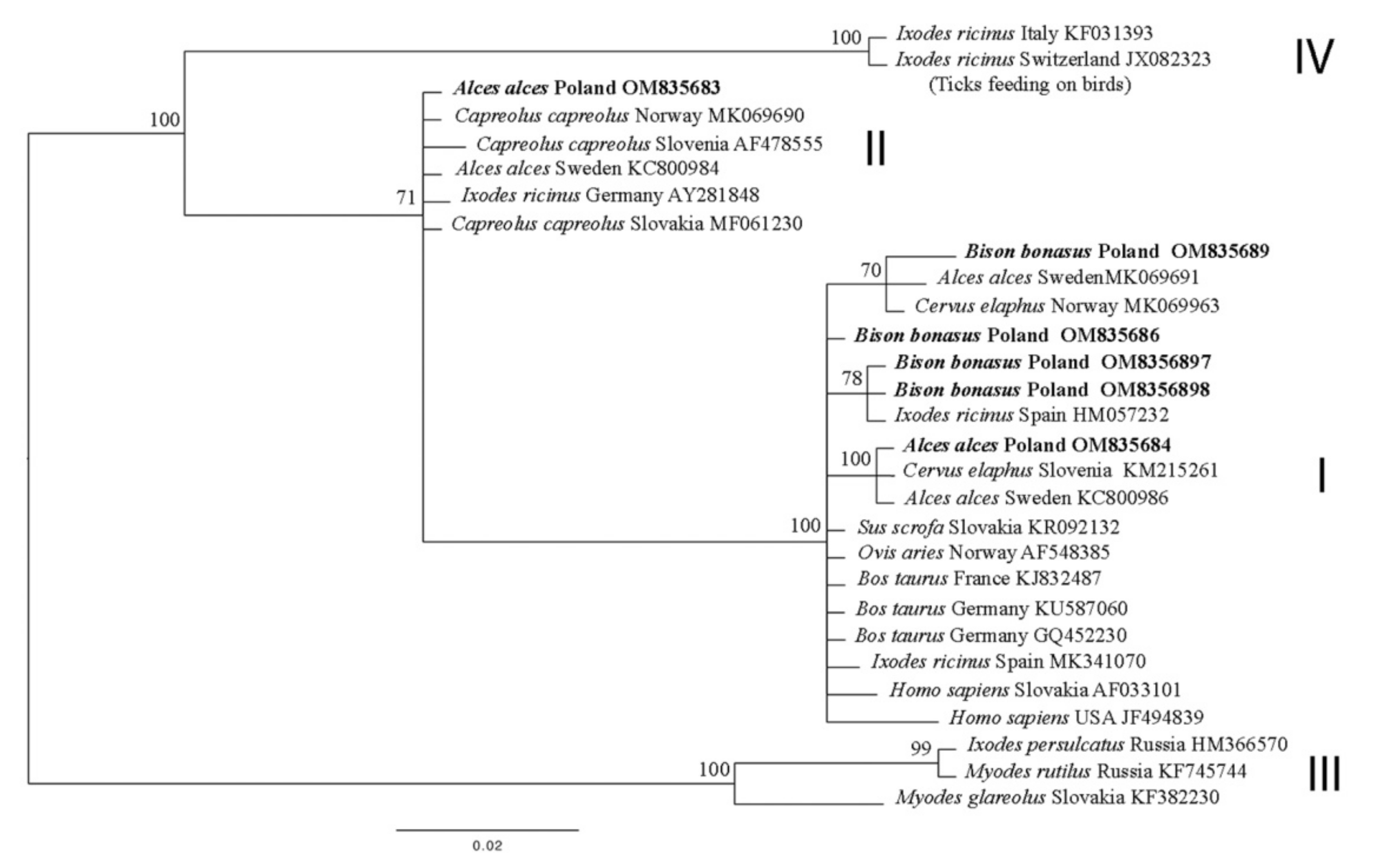
3.2. 16S rDNA
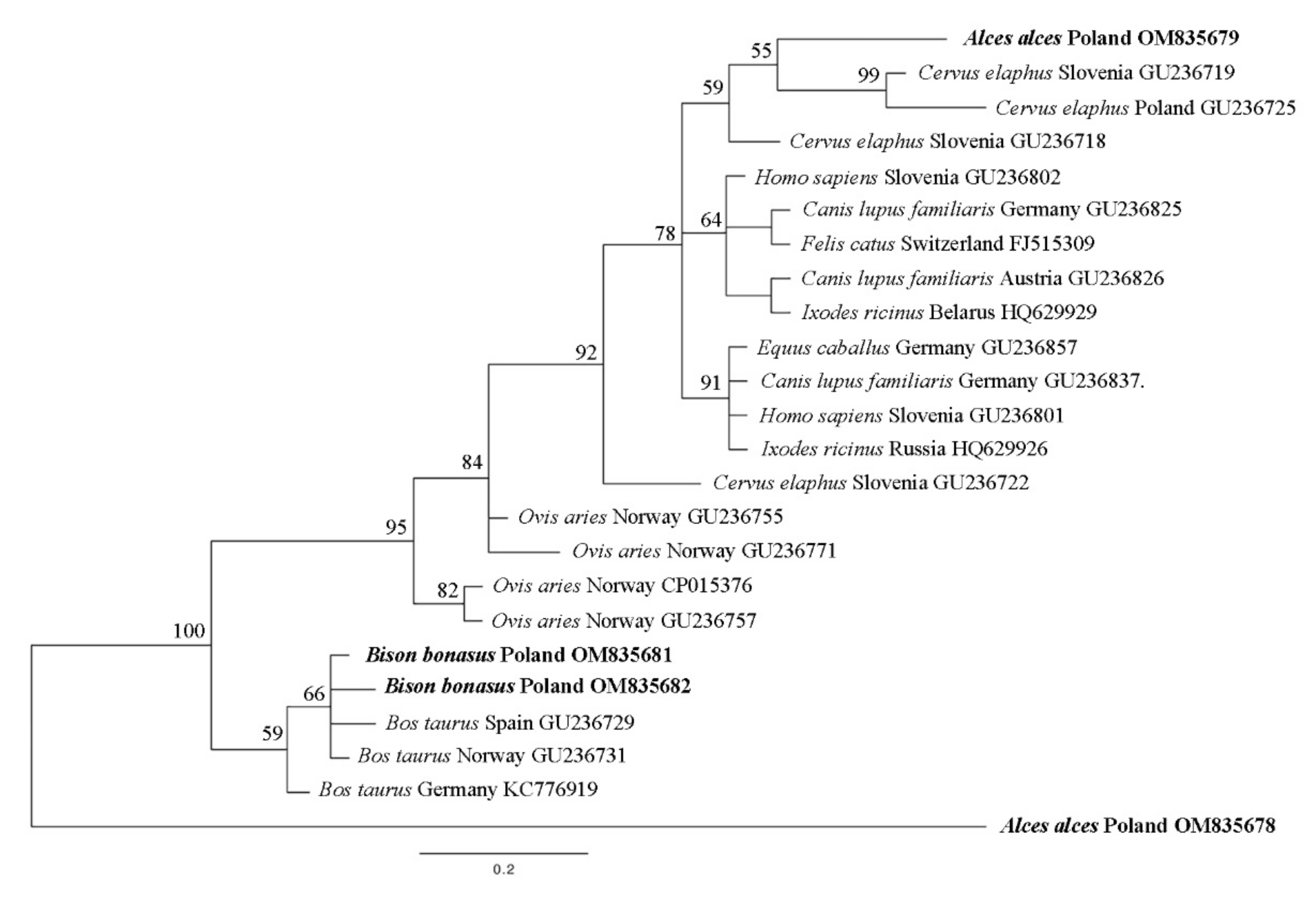
3.3. groEL
3.4. ankA
3.5. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scharf, W.; Schauer, S.; Freyburger, F.; Petrovec, M.; Schaarschmidt-Kiener, D.; Liebisch, G.; Runge, M.; Ganter, M.; Kehl, A.; Dumler, J.S.; et al. Distinct host species correlate with Anaplasma phagocytophilum ankA gene clusters. J. Clin. Microbiol. 2011, 49, 790–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Severo, M.S.; Stephens, K.D.; Kotsyfakis, M.; Pedra, J.H. Anaplasma phagocytophilum: Deceptively simple or simply deceptive? Future Microbiol. 2012, 7, 719–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, W.S.; Brownlee, A.; Wilson, D.R.; MacLeod, J. Tick-borne fever (a hitherto undescribed disease of sheep). J. Comp. Pathol. 1932, 45, 301–307. [Google Scholar] [CrossRef]
- Dumler, J.S.; Barbet, A.F.; Bekker, C.P.; Dasch, G.A.; Palmer, G.H.; Ray, S.C.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: Unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and ‘HGE agent’ as subjective synonyms of Ehrlichia phagocytophila. Int. J. Syst. Evol. Microbiol. 2001, 51 Pt 6, 2145–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzięgiel, B.; Adaszek, Ł.; Krzysiak, M.; Skrzypczak, M.; Adaszek, M.; Furmaga, B.; Winiarczyk, S. The occurrence of Anaplasma phagocytophilum in wild bison from the Bialowieza Primeval Forest in Eastern Poland. Berl. Munch. Tierarztl. Wochenschr. 2015, 128, 310–314. [Google Scholar] [PubMed]
- Jaarsma, R.I.; Sprong, H.; Takumi, K.; Kazimirova, M.; Silaghi, C.; Mysterud, A.; Rudolf, I.; Beck, R.; Földvári, G.; Tomassone, L.; et al. Anaplasma phagocytophilum evolves in geographical and biotic niches of vertebrates and ticks. Parasit. Vectors 2019, 12, 328. [Google Scholar] [CrossRef]
- Remesar, S.; Díaz, P.; Prieto, A.; García-Dios, D.; Fernández, G.; López, C.M.; Panadero, R.; Díez-Baños, P.; Morrondo, P. Prevalence and molecular characterization of Anaplasma phagocytophilum in roe deer (Capreolus capreolus) from Spain. Ticks Tick Borne Dis. 2020, 11, 101351. [Google Scholar] [CrossRef]
- Cafiso, A.; Bazzocchi, C.; Cavagna, M.; di Lorenzo, E.; Serra, V.; Rossi, R.; Comazzi, S. Molecular Survey of Babesia spp. and Anaplasma phagocytophilum in Roe Deer from a Wildlife Rescue Center in Italy. Animals 2021, 11, 3335. [Google Scholar] [CrossRef]
- Myczka, A.W.; Steiner-Bogdaszewska, Ż.; Filip-Hutsch, K.; Oloś, G.; Czopowicz, M.; Laskowski, Z. Detection of Anaplasma phagocytophilum in Wild and Farmed Cervids in Poland. Pathogens 2021, 10, 1190. [Google Scholar] [CrossRef]
- Alberti, A.; Zobba, R.; Chessa, B.; Addis, M.F.; Sparagano, O.; Pinna Parpaglia, M.L.; Cubeddu, T.; Pintori, G.; Pittau, M. Equine and canine Anaplasma phagocytophilum strains isolated on the island of Sardinia (Italy) are phylogenetically related to pathogenic strains from the United States. Appl. Environ. Microbiol. 2005, 71, 6418–6422. [Google Scholar] [CrossRef] [Green Version]
- Jahfari, S.; Coipan, E.C.; Fonville, M.; van Leeuwen, A.D.; Hengeveld, P.; Heylen, D.; Heyman, P.; van Maanen, C.; Butler, C.M.; Földvári, G.; et al. Circulation of four Anaplasma phagocytophilum ecotypes in Europe. Parasit. Vectors 2014, 7, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stigum, V.M.; Jaarsma, R.I.; Sprong, H.; Rolandsen, C.M.; Mysterud, A. Infection prevalence and ecotypes of Anaplasma phagocytophilum in moose Alces alces, red deer Cervus elaphus, roe deer Capreolus capreolus and Ixodes ricinus ticks from Norway. Parasit. Vectors 2019, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Adamska, M. The role of different species of wild ungulates and Ixodes ricinus ticks in the circulation of genetic variants of Anaplasma phagocytophilum in a forest biotope in north-western Poland. Ticks Tick Borne Dis. 2020, 11, 101465. [Google Scholar] [CrossRef] [PubMed]
- Jouglin, M.; Chagneau, S.; Faille, F.; Verheyden, H.; Bastian, S.; Malandrin, L. Detecting and characterizing mixed infections with genetic variants of Anaplasma phagocytophilum in roe deer (Capreolus capreolus) by developing an ankA cluster-specific nested PCR. Parasit. Vectors 2017, 10, 377. [Google Scholar] [CrossRef] [PubMed]
- Bown, K.J.; Lambin, X.; Ogden, N.H.; Begon, M.; Telford, G.; Woldehiwet, Z.; Birtles, R.J. Delineating Anaplasma phagocytophilum ecotypes in coexisting, discrete enzootic cycles. Emerg. Infect. Dis. 2009, 15, 1948–1954. [Google Scholar] [CrossRef] [PubMed]
- Massung, R.F.; Owens, J.H.; Ross, D.; Reed, K.D.; Petrovec, M.; Bjoersdorff, A.; Coughlin, R.T.; Beltz, G.A.; Murphy, C.I. Sequence analysis of the ank gene of granulocytic ehrlichiae. J. Clin. Microbiol. 2000, 38, 2917–2922. [Google Scholar] [CrossRef] [Green Version]
- Von Loewenich, F.D.; Baumgarten, B.U.; Schröppel, K.; Geissdörfer, W.; Röllinghoff, M.; Bogdan, C. High diversity of ankA sequences of Anaplasma phagocytophilum among Ixodes ricinus ticks in Germany. J. Clin. Microbiol. 2003, 41, 5033–5040. [Google Scholar] [CrossRef] [Green Version]
- Majazki, J.; Wüppenhorst, N.; Hartelt, K.; Birtles, R.; von Loewenich, F.D. Anaplasma phagocytophilum strains from voles and shrews exhibit specific ankA gene sequences. BMC Vet. Res. 2013, 9, 235. [Google Scholar] [CrossRef] [Green Version]
- Krasiński, Z.A. Żubr Puszcz Imperator; Wydawnictwo BPN: Białowieża, Poland, 2005; pp. 1–24. (In Polish) [Google Scholar]
- Wawrzyniak, P. Population dynamics, its impact upon the habitat and necessity of managament the moose (Alces alces) population in Poland. In Proceedings of the Conference “Zarządzanie Populacjami Zwierząt”, Warsaw, Poland, 5 August 2016; Łowiec Polski: Warsaw, Poland, 2016; pp. 17–27. (In Polish). [Google Scholar]
- Matei, I.A.; Estrada-Peña, A.; Cutler, S.J.; Vayssier-Taussat, M.; Varela-Castro, L.; Potkonjak, A.; Zeller, H.; Mihalca, A.D. A review on the eco-epidemiology and clinical management of human granulocytic anaplasmosis and its agent in Europe. Parasit. Vectors 2019, 12, 599. [Google Scholar] [CrossRef]
- Szewczyk, T.; Werszko, J.; Myczka, A.W.; Laskowski, Z.; Karbowiak, G. Molecular detection of Anaplasma phagocytophilum in wild carnivores in north-eastern Poland. Parasit. Vectors 2019, 12, 465. [Google Scholar] [CrossRef] [Green Version]
- Massung, R.F.; Levin, M.L.; Munderloh, U.G.; Silverman, D.J.; Lynch, M.J.; Gaywee, J.K.; Kurtti, T.J. Isolation and propagation of the Ap-Variant 1 strain of Anaplasma phagocytophilum in a tick cell line. J. Clin. Microbiol. 2007, 45, 2138–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myczka, A.W.; Szewczyk, T.; Laskowski, Z. The Occurrence of Zoonotic Anaplasma phagocytophilum Strains, in the Spleen and Liver of Wild Boars from North-West and Central Parts of Poland. Acta Parasitol. 2021, 66, 1082–1085. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Altman, D.G.; Machin, D.; Bryant, T.N.; Gardner, M.J. Statistics with Confidence, 2nd ed.; BMJ Books: London, UK, 2000. [Google Scholar]
- Matsumoto, K.; Grzeszczuk, A.; Brouqui, P.; Raoult, D. Rickettsia raoultii and Anaplasma phagocytophilum in Dermacentor reticulatus ticks collected from Bialowieza Primeval Forest European bison (Bison bonasus bonasus), Poland. Clin. Microbiol. Infect. 2009, 15, 286–287. [Google Scholar] [CrossRef] [Green Version]
- Karbowiak, G.; Víchová, B.; Werszko, J.; Demiaszkiewicz, A.W.; Pyziel, A.M.; Sytykiewicz, H.; Szewczyk, T.; Peťko, B. The infection of reintroduced ruminants—Bison bonasus and Alces alces—with Anaplasma phagocytophilum in northern Poland. Acta Parasitol. 2015, 60, 645–648. [Google Scholar] [CrossRef]
- De la Fuente, J.; Golsteyn Thomas, E.J.; van den Bussche, R.A.; Hamilton, R.G.; Tanaka, E.E.; Druhan, S.E.; Kocan, K.M. Characterization of Anaplasma marginale isolated from North American bison. Appl. Environ. Microbiol. 2003, 69, 5001–5005. [Google Scholar] [CrossRef] [Green Version]
- Raczyński, J. European Bison Pedigree Book; Wydawnictwo BPN: Białowieża, Poland, 2020. [Google Scholar]
- Klich, D.; Łopucki, R.; Perlińska-Teresiak, M.; Lenkiewicz-Bardzińska, A.; Olech, W. Human-Wildlife Conflict: The Human Dimension of European Bison Conservation in the Bieszczady Mountains (Poland). Animals 2021, 11, 503. [Google Scholar] [CrossRef]
- Pūraitė, I.; Rosef, O.; Paulauskas, A.; Radzijevskaja, J. Anaplasma phagocytophilum infection in moose (Alces alces) in Norway. Microbes Infect. 2015, 17, 823–828. [Google Scholar] [CrossRef]
- Razanske, I.; Rosef, O.; Radzijevskaja, J.; Krikstolaitis, R.; Paulauskas, A. Impact of tick-borne Anaplasma phagocytophilum infections in calves of moose (Alces alces) in southern Norway. Folia Parasitol. 2021, 68, 023. [Google Scholar] [CrossRef] [PubMed]
- Malmsten, J.; Dalin, A.M.; Moutailler, S.; Devillers, E.; Gondard, M.; Felton, A. Vector-Borne Zoonotic Pathogens in Eurasian Moose (Alces alces alces). Vector Borne Zoonotic Dis. 2019, 19, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Elliott, J.A.; Dickson, C.C.; Kantar, L.; O’Neal, M.R.; Lichtenwalner, A.; Bryant, A.; Jakubas, W.J.; Pekins, P.J.; de Urioste-Stone, S.M.; Kamath, P.L. Prevalence and risk factors of Anaplasma infections in Eastern moose (Alces alces americana) and winter ticks (Dermacentor albipictus) in Maine USA. J. Wildl. Dis. 2021, 57, 844–855. [Google Scholar] [CrossRef] [PubMed]
- Víchová, B.; Majláthová, V.; Nováková, M.; Stanko, M.; Hviščová, I.; Pangrácová, L.; Chrudimský, T.; Čurlík, J.; Petko, B. Anaplasma infections in ticks and reservoir host from Slovakia. Infect. Genet. Evol. 2014, 22, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Chastagner, A.; Dugat, T.; Vourc’h, G.; Verheyden, H.; Legrand, L.; Bachy, V.; Chabanne, L.; Joncour, G.; Maillard, R.; Boulouis, H.J.; et al. Multilocus sequence analysis of Anaplasma phagocytophilum reveals three distinct lineages with different host ranges in clinically ill French cattle. Vet. Res. 2014, 45, 114. [Google Scholar] [CrossRef]
- Silaghi, C.; Nieder, M.; Sauter-Louis, C.; Knubben-Schweizer, G.; Pfister, K.; Pfeffer, M. Epidemiology, genetic variants and clinical course of natural infections with Anaplasma phagocytophilum in a dairy cattle herd. Parasit. Vectors 2018, 11, 20. [Google Scholar] [CrossRef] [Green Version]
- Malmsten, J.; Widén, D.G.; Rydevik, G.; Yon, L.; Hutchings, M.R.; Thulin, C.G.; Söderquist, L.; Aspan, A.; Stuen, S.; Dalin, A.M. Temporal and spatial variation in Anaplasma phagocytophilum infection in Swedish moose (Alces alces). Epidemiol. Infect. 2014, 142, 1205–1213. [Google Scholar] [CrossRef] [Green Version]
- Masuzawa, T.; Uchishima, Y.; Fukui, T.; Okamoto, Y.; Muto, M.; Koizumi, N.; Yamada, A. Detection of Anaplasma phagocytophilum from Wild Boars and Deer in Japan. Jpn. J. Infect. Dis. 2011, 64, 333–336. [Google Scholar] [CrossRef]
- Kazimírová, M.; Hamšíková, Z.; Špitalská, E.; Minichová, L.; Mahríková, L.; Caban, R.; Sprong, H.; Fonville, M.; Schnittger, L.; Kocianová, E. Diverse tick-borne microorganisms identified in free-living ungulates in Slovakia. Parasit. Vectors 2018, 11, 495. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hosts Group | Host | Country | Gen Bank No. |
|---|---|---|---|
| Carnivores | Red fox (Vulpes vulpes) | Switzerland | KX180948.1 |
| Poland | MH328211.1 | ||
| Dog (Canis lupus familiaris) | Croatia | KY114936.1 | |
| Germany | JX173651.1 | ||
| Republic of South Africa (RSA) | MK814406.1 | ||
| Iraq | MN453475.1 | ||
| Japan | LC334014.1 | ||
| European badger (Meles meles) | Poland | MH328211 | |
| Coyote (Canis latrans) | United States of America (USA) | AF170728.1 | |
| Raccoon dog (Nyctereutes procyonoides) | South Korea | KY458557.1 | |
| Cat (Felis catus) | South Korea | KR021165.1 | |
| European polecat (Mustela putorius) | Poland | MH328208.1 | |
| Humans | Homo sapiens | Austria | KT454992.1 |
| Belgium | KM259921.1 | ||
| USA | AF093788.1 | ||
| South Korea | KP306520.1 | ||
| Small mammals | Bank vole (Clethrionomys glareolus) | United Kingdom | AY082656.1 |
| Natal multimammate mouse (Mastomys natalensis) | RSA | MK814411.1 | |
| Northern red-backed vole (Myodes rutilus) | Russia (Sverdlovsk region) | HQ630622.1 | |
| Black-striped field mouse (Apodemus agrarius) | South Korea | KR611719.1 | |
| China | GQ412337 | ||
| DQ342324 | |||
| European hedgehog (Erinaceus europaeus) | Germany | FN390878.1 | |
| Insect | Ixodes ricinus | Estonia | MW922755.1 |
| Belarus | HQ629915.1 | ||
| Austria | JX173652.1 | ||
| Turkey | FJ172530.1 | ||
| Ixodes pacificus | USA | KP276588.1 | |
| Ixodes persulcatus | Russia (Irkutsk region) | HM366584.1 | |
| Ixodes tapirus | Panama | MW677508.1 | |
| Haemaphysalis longicornis | China | KF569908 | |
| South Korea | GU064898 | ||
| Haematopota pluvialis | Poland | MH844585.1 | |
| Ungulates | Sheep (Ovis aries) | Norway | CP015376.1 |
| Red deer (Cervus elaphus) | Slovenia | KM215243.1 | |
| Roe deer (Capreolus capreolus) | Spain | MN170723.1 | |
| Wild boar (Sus scrofa) | Poland | MT510541.1 | |
| Llama (Lama glama) | USA | AF241532.1 | |
| Horse (Equus ferus caballus) | USA | AF172166.1 | |
| Cow (Bos taurus taurus) | Turkey | KP745629.1 | |
| Goat (Capra hircus) | China | KF569909.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Myczka, A.W.; Kaczor, S.; Filip-Hutsch, K.; Czopowicz, M.; Plis-Kuprianowicz, E.; Laskowski, Z. Prevalence and Genotyping of Anaplasma phagocytophilum Strains from Wild Animals, European Bison (Bison bonasus) and Eurasian Moose (Alces alces) in Poland. Animals 2022, 12, 1222. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12091222
Myczka AW, Kaczor S, Filip-Hutsch K, Czopowicz M, Plis-Kuprianowicz E, Laskowski Z. Prevalence and Genotyping of Anaplasma phagocytophilum Strains from Wild Animals, European Bison (Bison bonasus) and Eurasian Moose (Alces alces) in Poland. Animals. 2022; 12(9):1222. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12091222
Chicago/Turabian StyleMyczka, Anna W., Stanisław Kaczor, Katarzyna Filip-Hutsch, Michał Czopowicz, Elwira Plis-Kuprianowicz, and Zdzisław Laskowski. 2022. "Prevalence and Genotyping of Anaplasma phagocytophilum Strains from Wild Animals, European Bison (Bison bonasus) and Eurasian Moose (Alces alces) in Poland" Animals 12, no. 9: 1222. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12091222