Alleviation of Cadmium Adverse Effects by Improving Nutrients Uptake in Bitter Gourd through Cadmium Tolerant Rhizobacteria


 ,
,  , ,
, ,  , , , , and
, , , , and 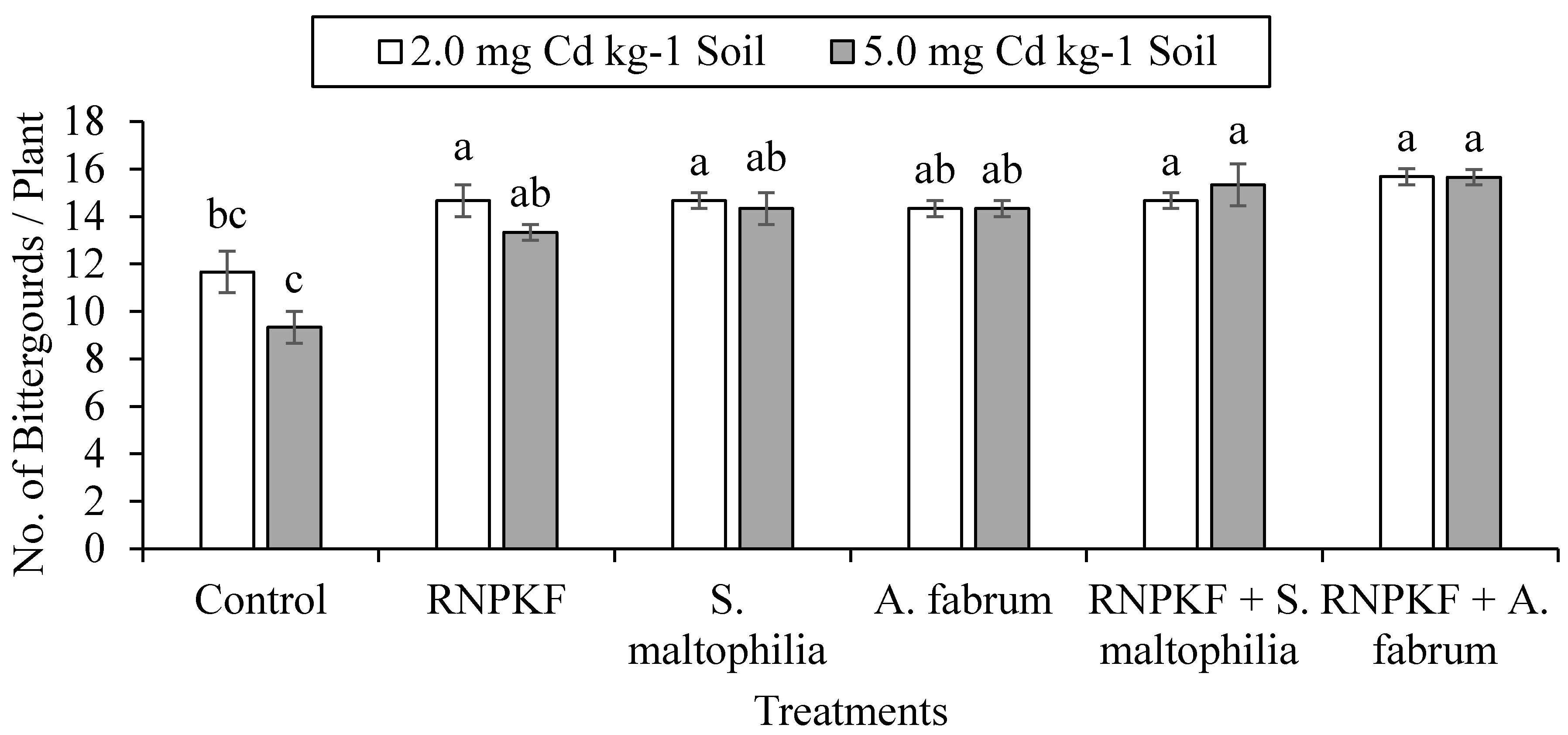{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Treatments
2.2. Collection of Bacterial Strains and Broth
2.3. Seeds Sterilization and Sowing
2.4. Irrigation and Fertilizer Application
2.5. Harvesting and Samples Analyses
2.6. Statistical Analyses
3. Results
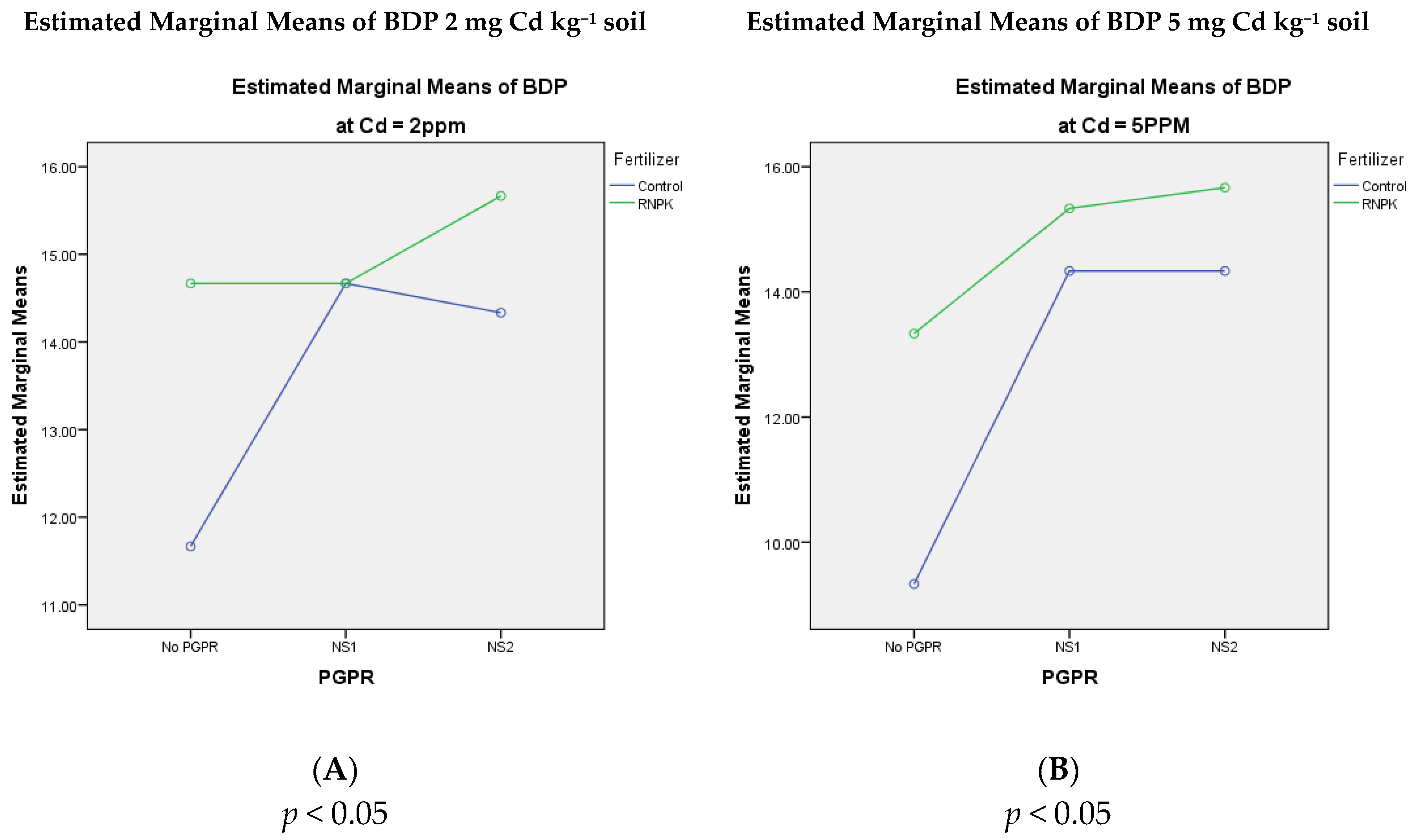
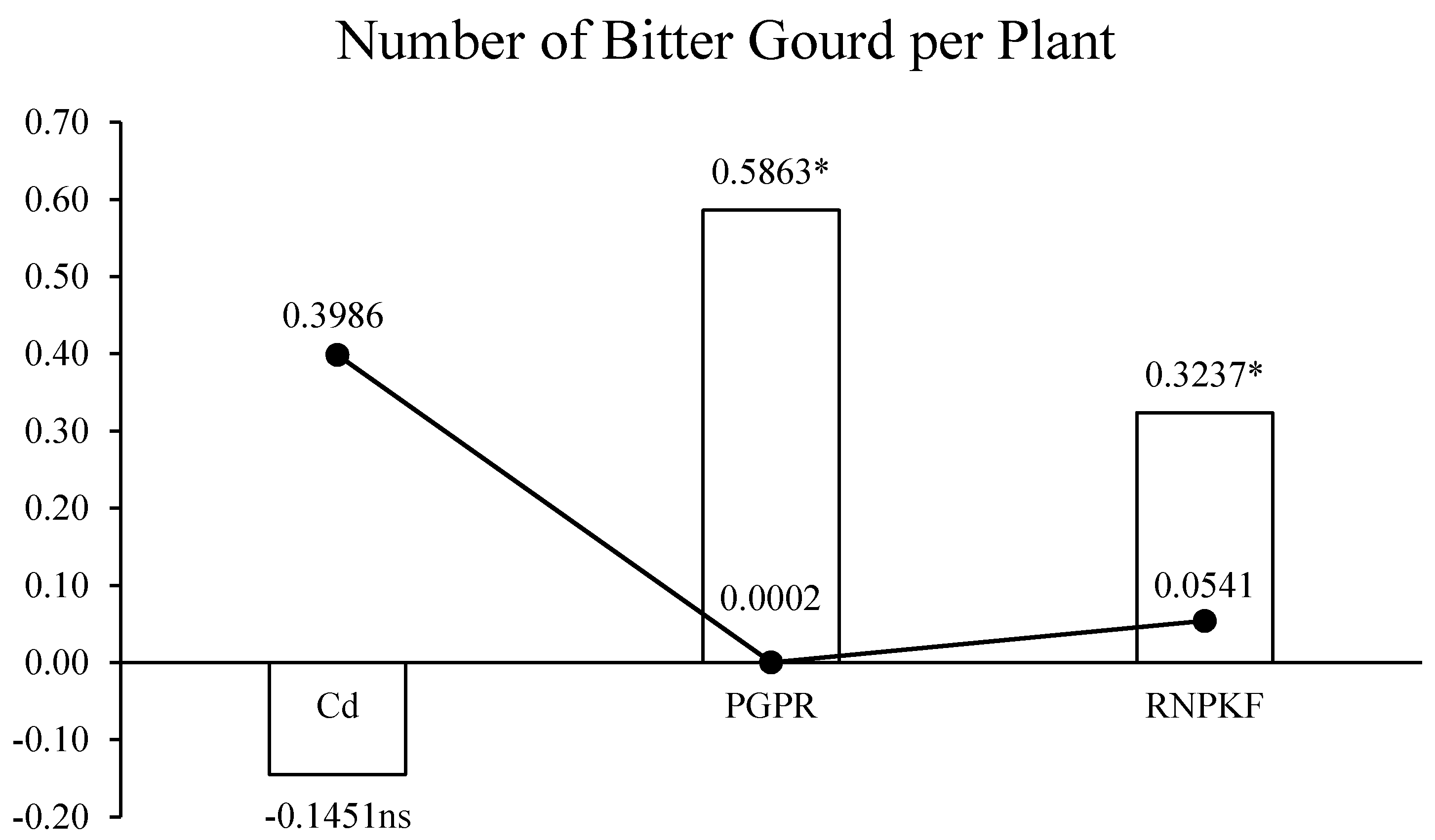
3.1. Number of Bitter Gourds per Plant
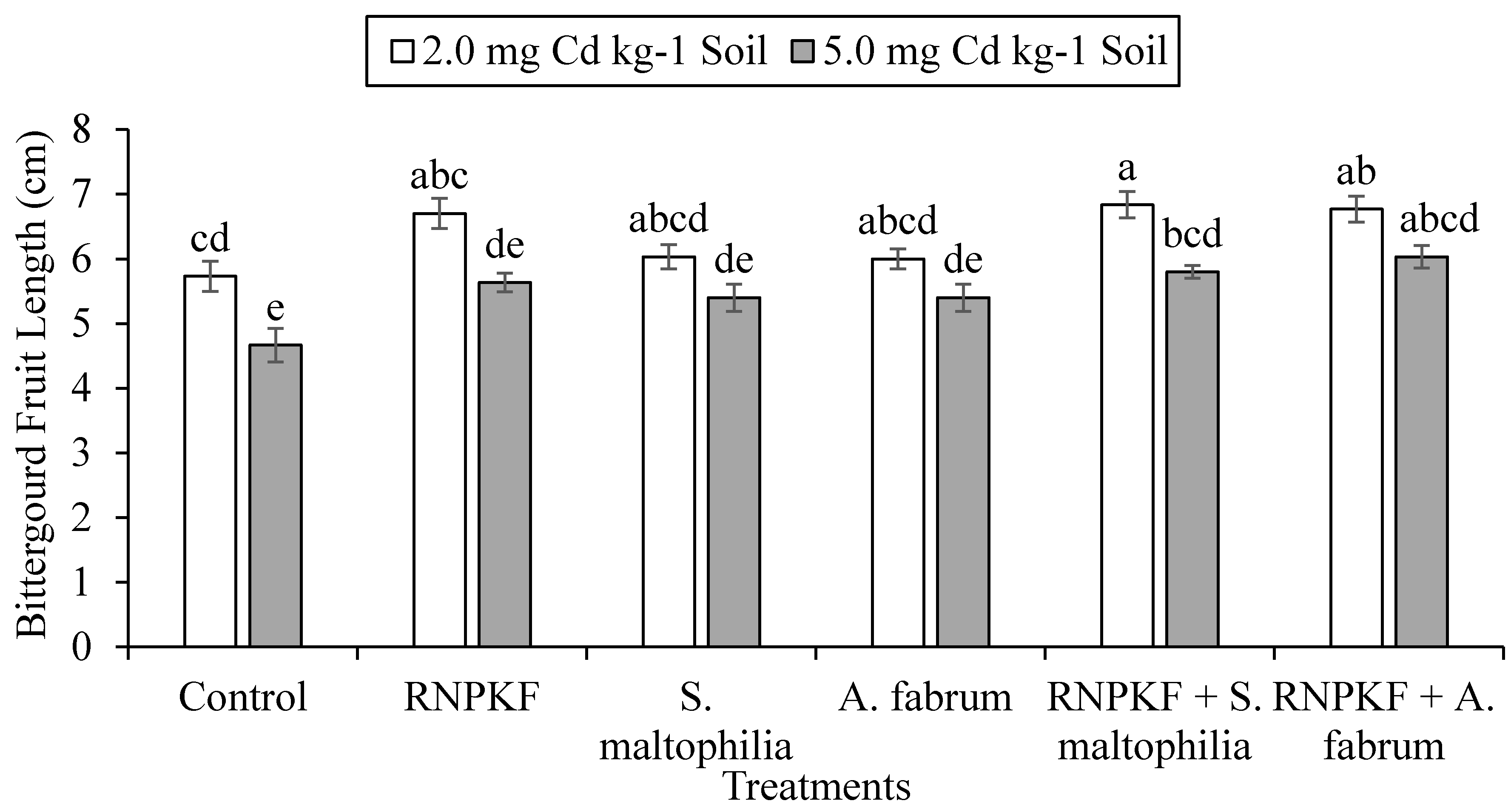
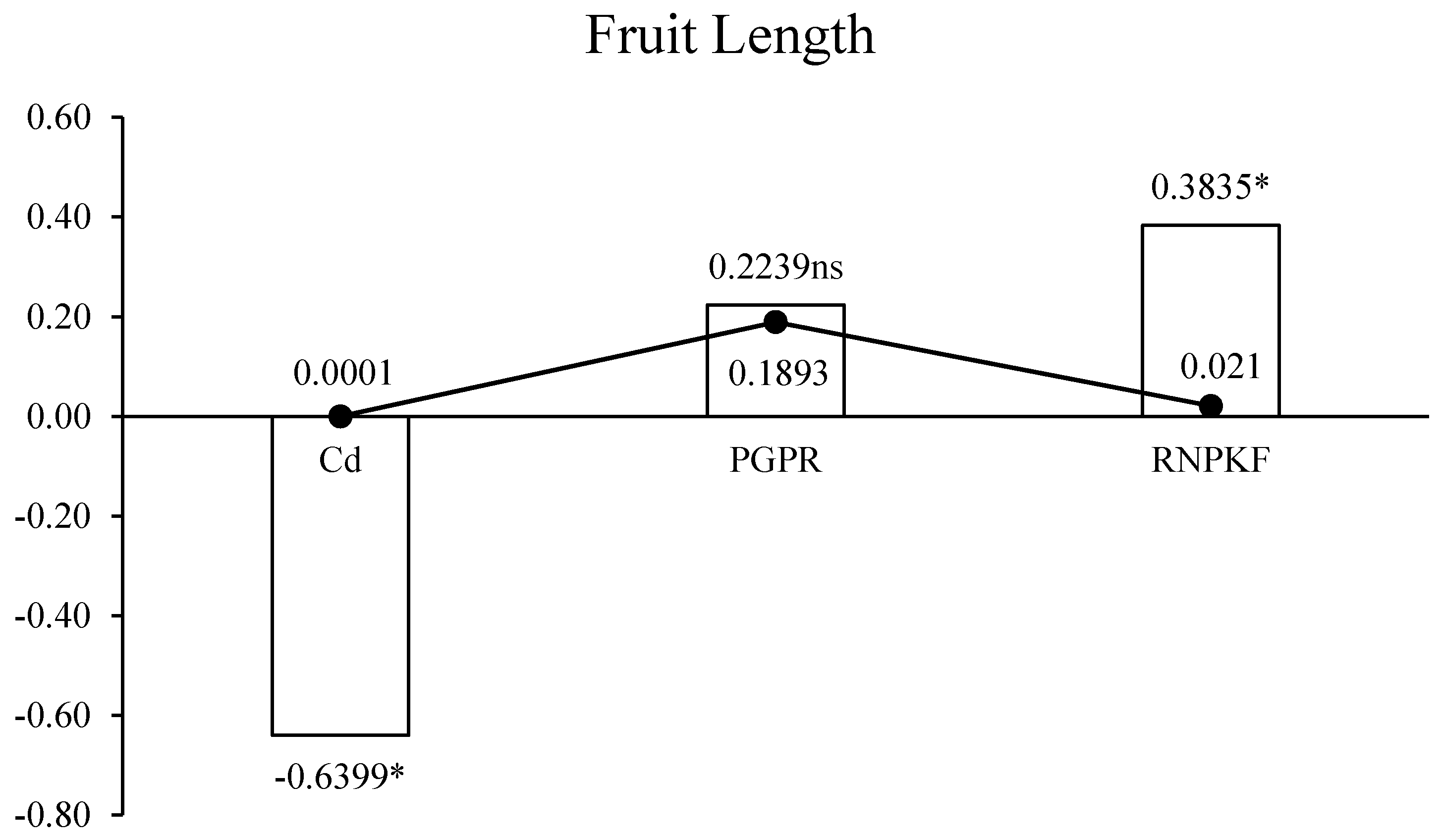
3.2. Bitter Gourd Fruit Length
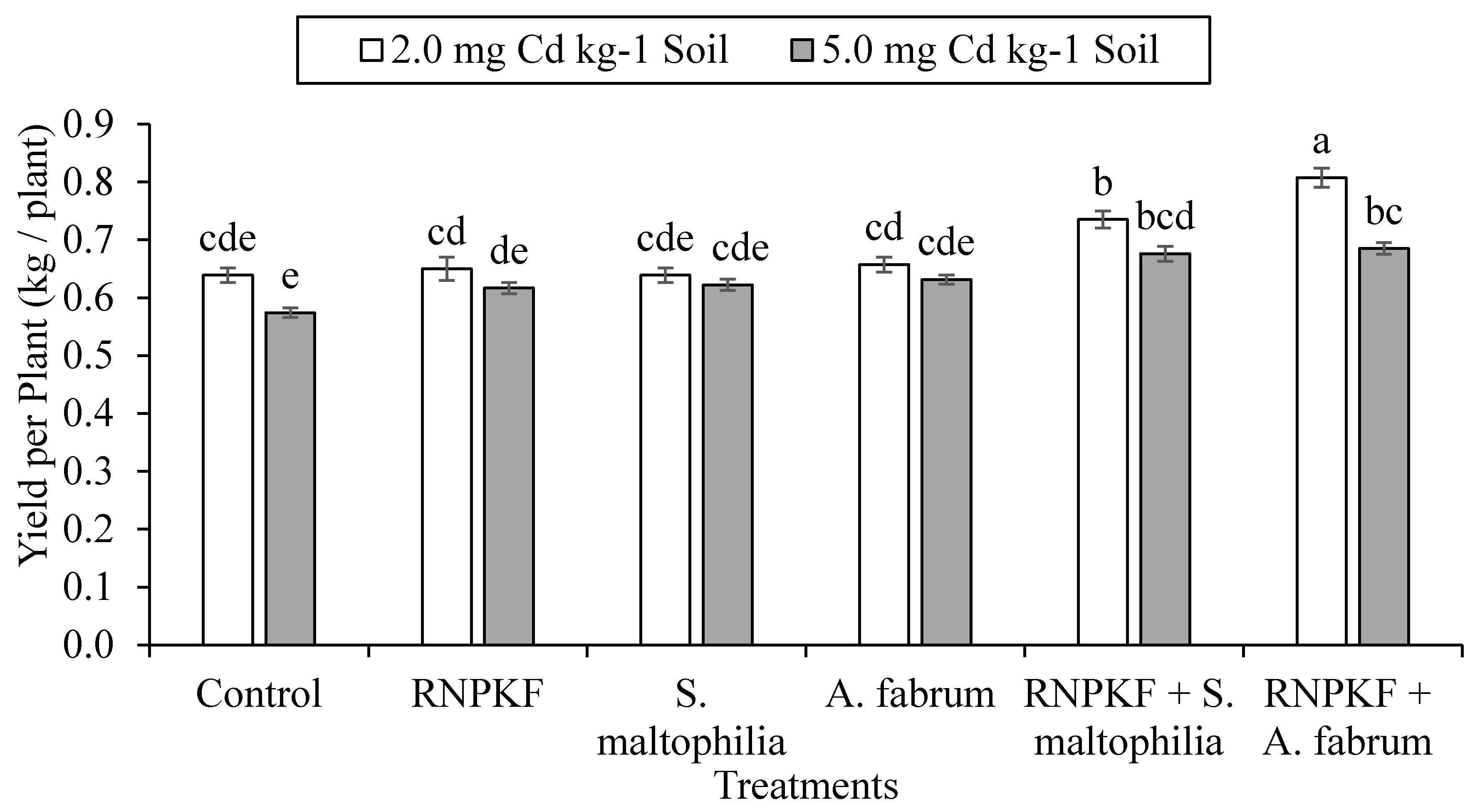
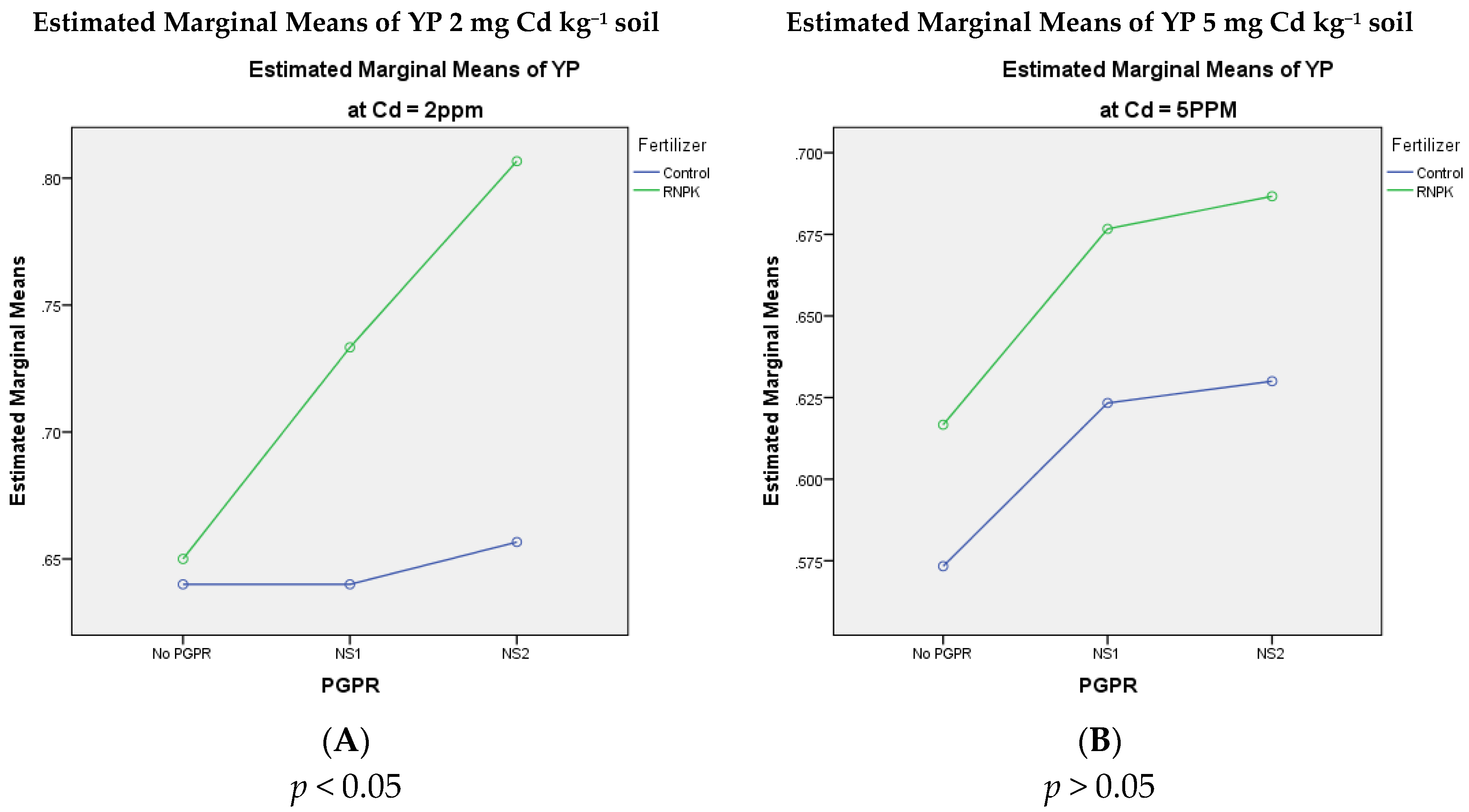
3.3. Bitter Gourd Yield per Plant
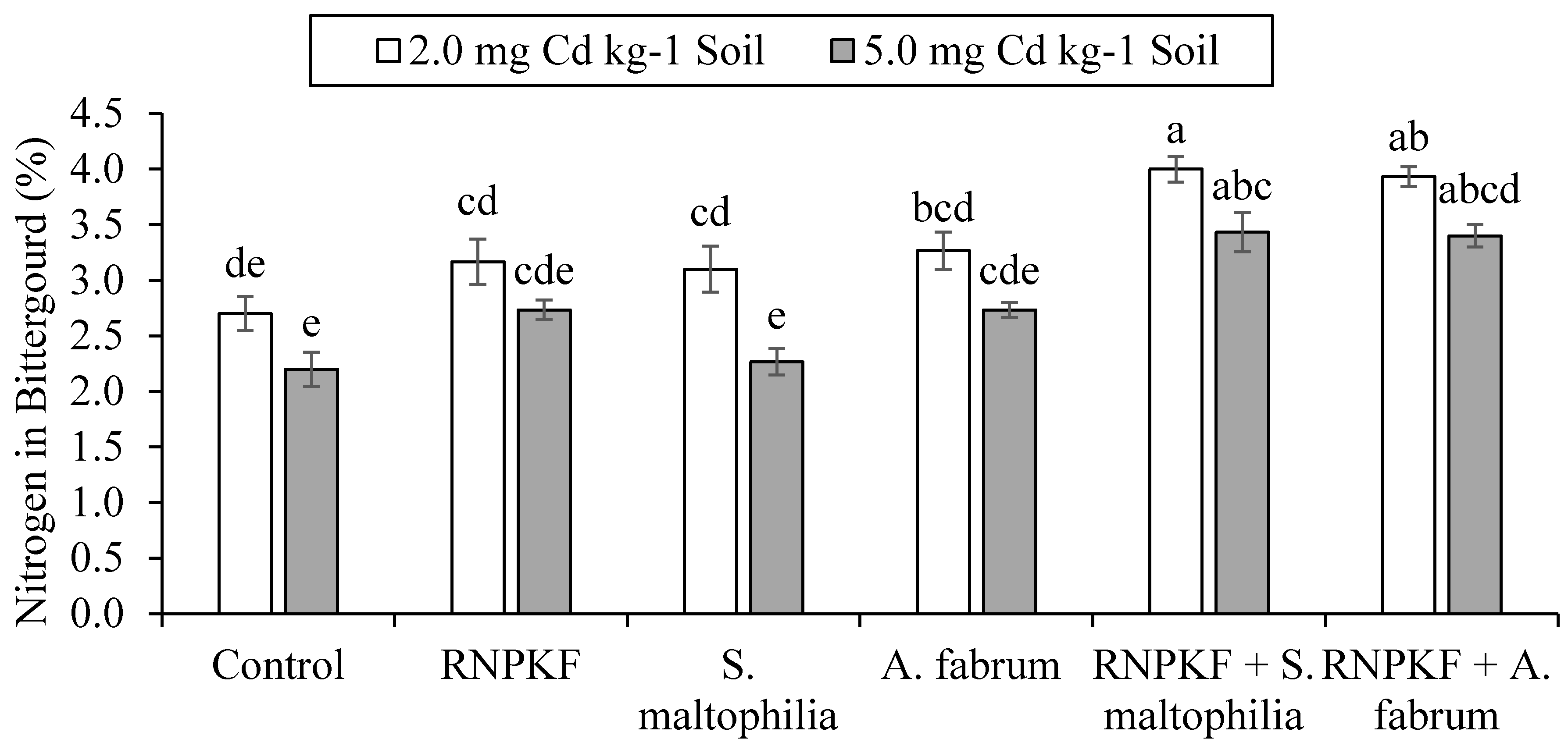
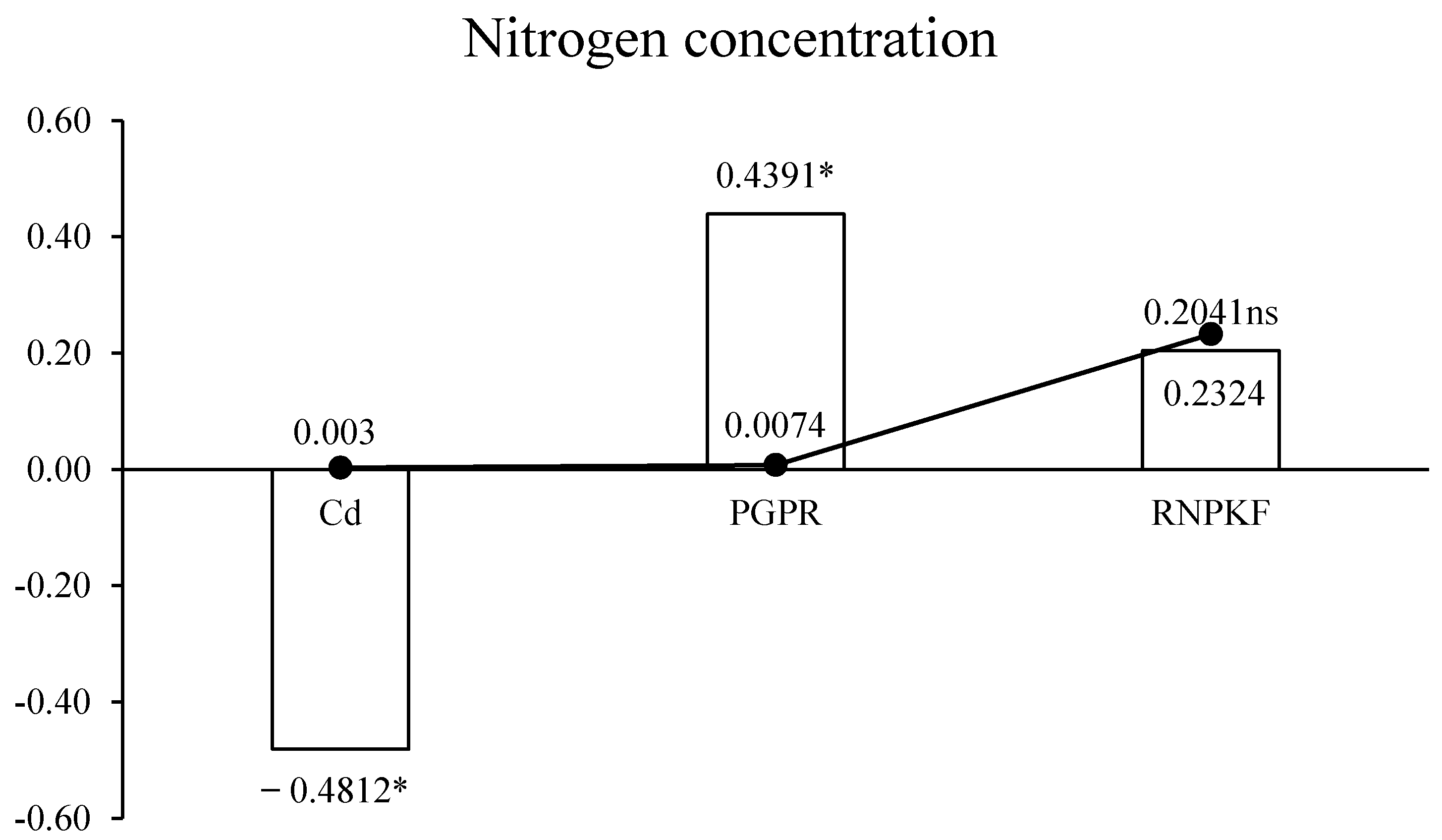
3.4. Nitrogen Concentration in Bitter Gourd
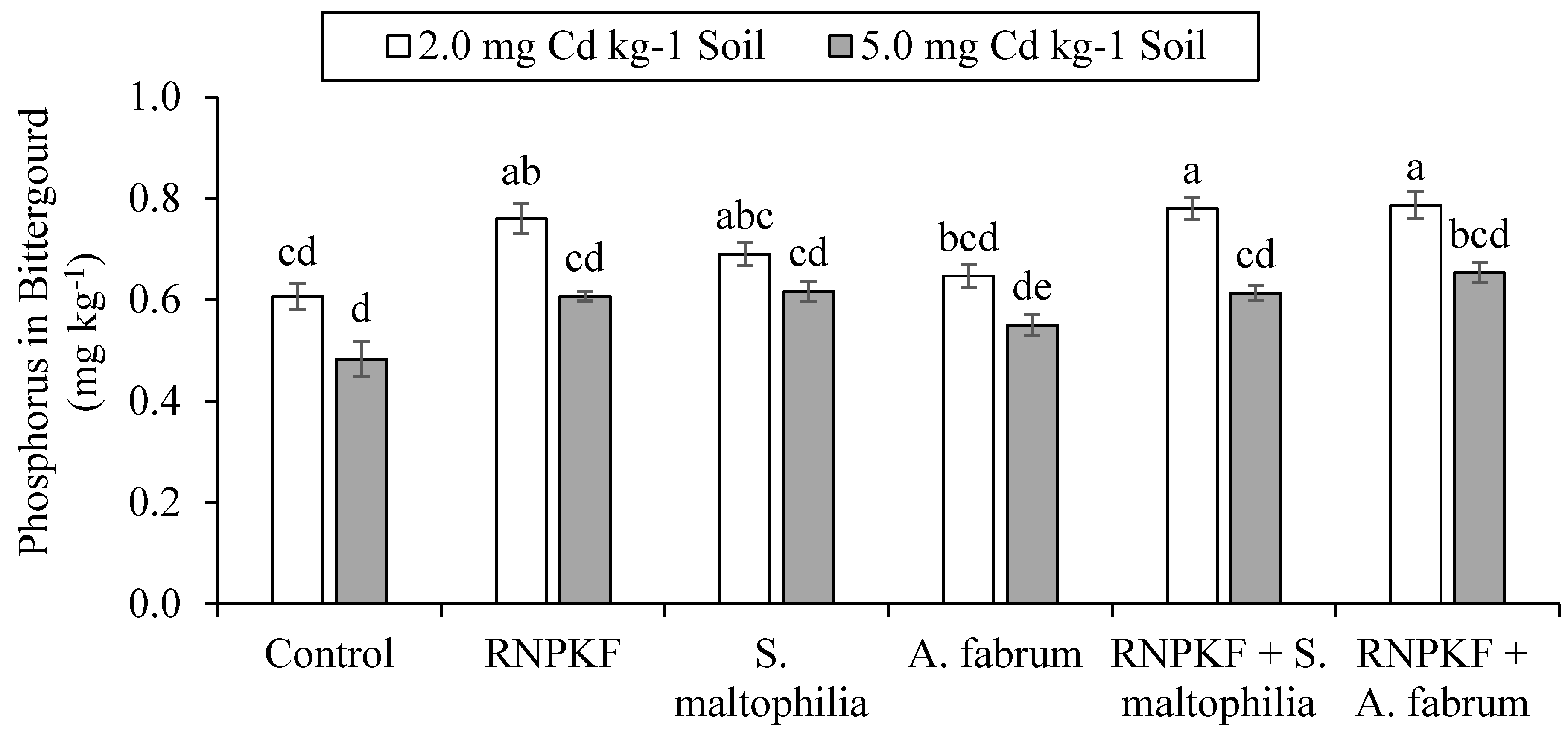
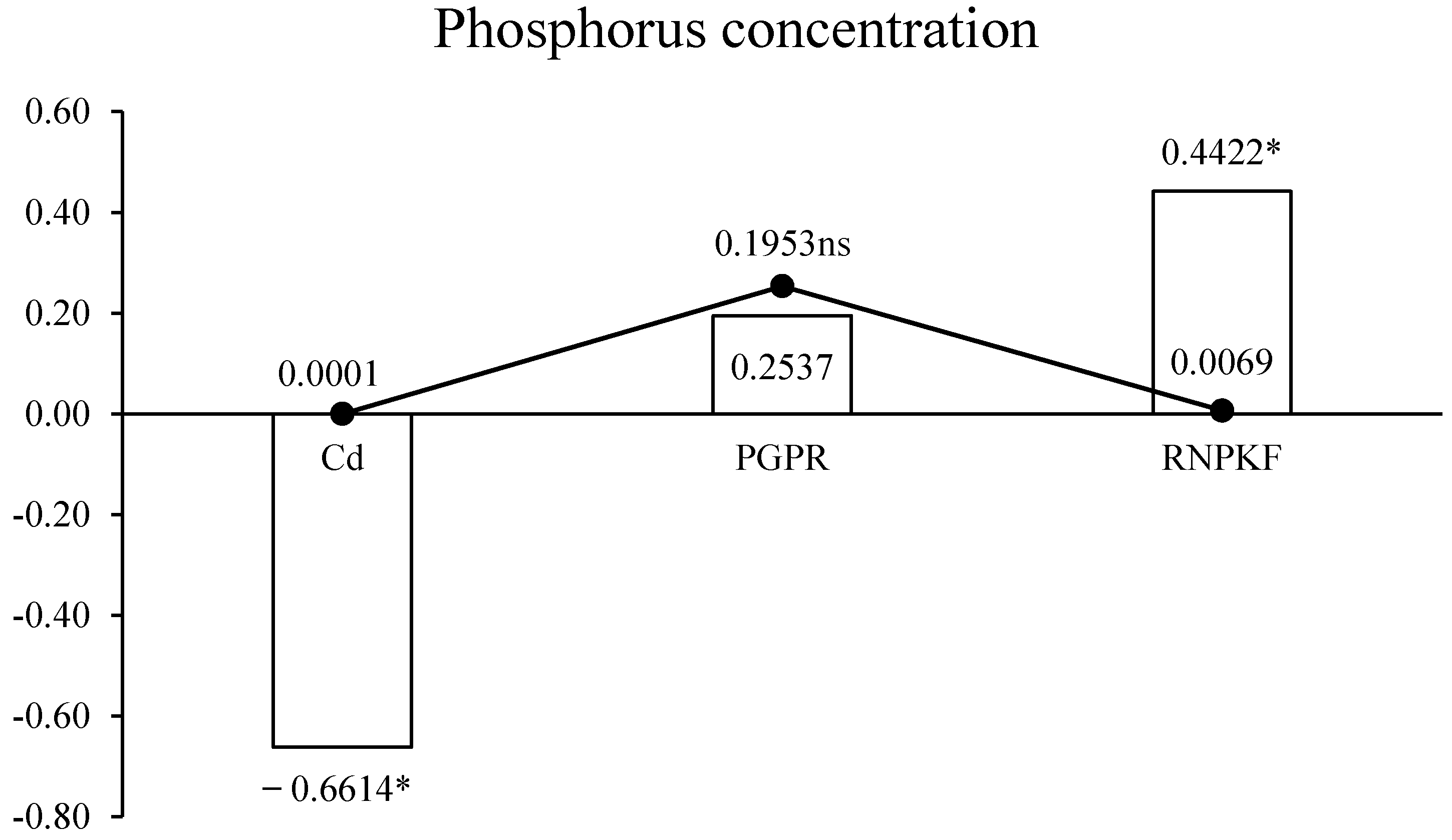
3.5. Phosphorus Concentration in Bitter Gourd
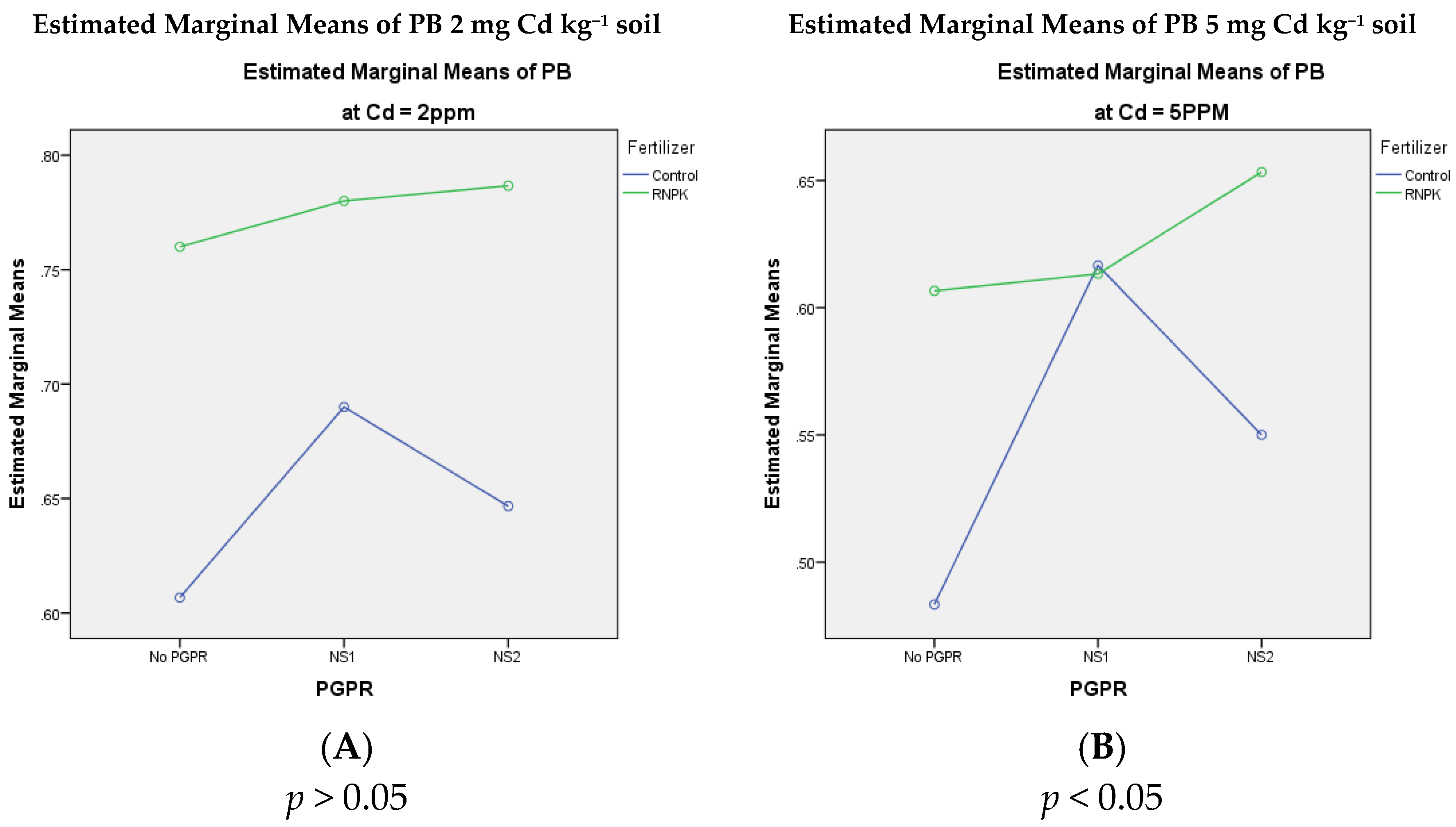
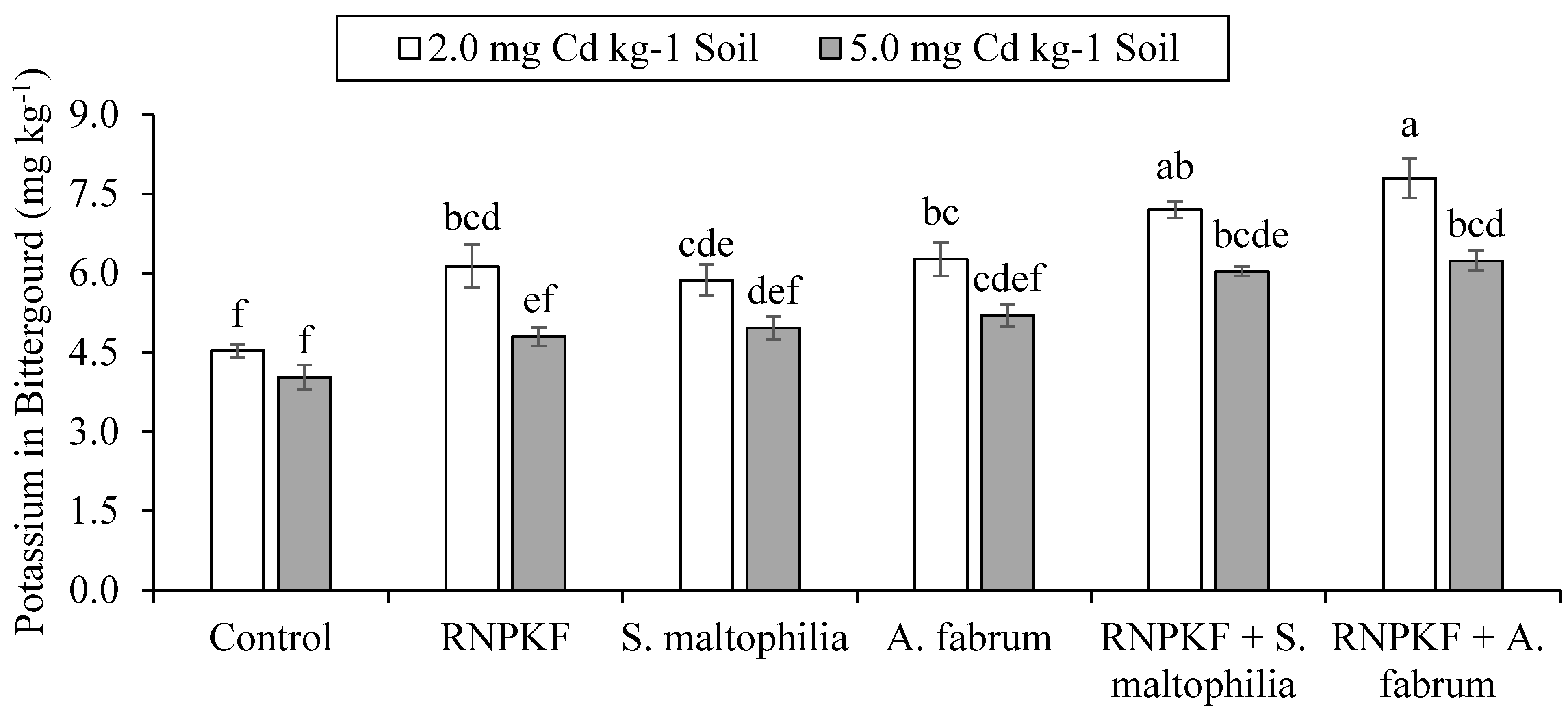
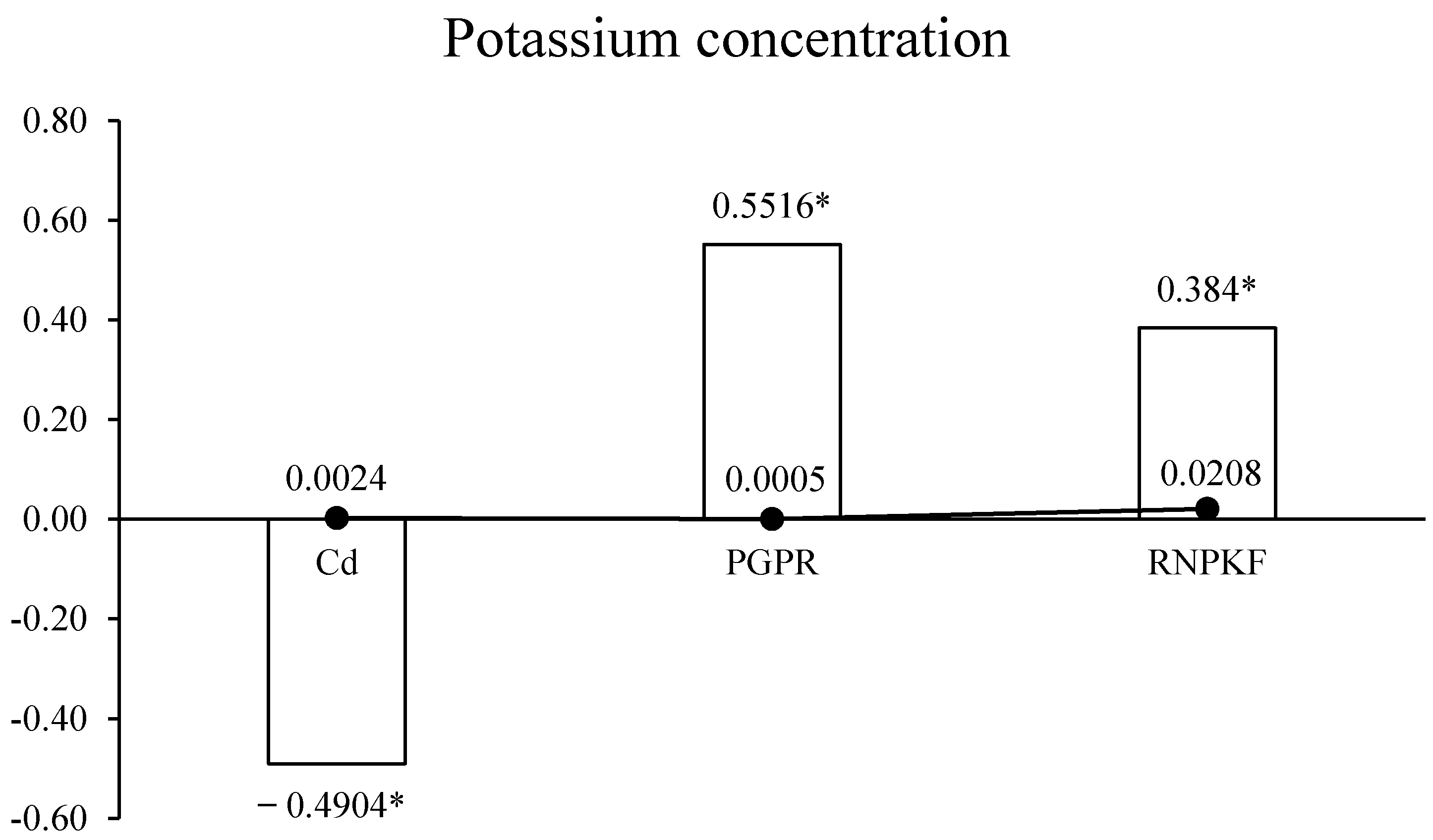
3.6. Potassium Concentration in Bitter Gourd
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fu, F.; Wang, Q. Removal of heavy metal ions from wastewaters: A review. J. Environ. Manag. 2011, 92, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Meena, R.S.; Kumar, S.; Datta, R.; Lal, R.; Vijayakumar, V.; Brtnicky, M.; Sharma, M.P.; Yadav, G.S.; Jhariya, M.K.; Jangir, C.K. Impact of Agrochemicals on Soil Microbiota and Management: A Review. Land 2020, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Nazar, R. Cadmium Toxicity in Plants and Role of Mineral Nutrients in Its Alleviation. Am. J. Plant Sci. 2012, 3, 1476–1489. [Google Scholar] [CrossRef] [Green Version]
- Lazar, V.; Cernat, R.; Balotescu, C.; Cotar, A.; Coipan, E.; Cojocaru, C. Correlation between Multiple Antibiotic Resistance and Heavy-Metal Tolerance among some E.coli Strains Isolated from Polluted Waters. Bacteriol. Virusol. Parazitol. Epidemiol. (Buchar. Rom. 1990) 2002, 47, 155–160. [Google Scholar]
- Molaei, A.; Lakzian, A.; Haghnia, G.; Astaraei, A.; Rasouli-Sadaghiani, M.; Ceccherini, M.T.; Datta, R. Assessment of some cultural experimental methods to study the effects of antibiotics on microbial activities in a soil: An incubation study. PLoS ONE 2017, 12, e0180663. [Google Scholar] [CrossRef] [PubMed]
- Molaei, A.; Lakzian, A.; Datta, R.; Haghnia, G.; Astaraei, A.; Rasouli-Sadaghiani, M.; Ceccherini, M.T. Impact of chlortetracycline and sulfapyridine antibiotics on soil enzyme activities. Int. Agrophys. 2017, 31, 499–505. [Google Scholar] [CrossRef] [Green Version]
- Sanita di Toppi, L.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar] [CrossRef]
- Vahter, M.; Berglund, M.; Slorach, S.; Friberg, L.; Sarić, M.; Zheng, X.; Fujita, M. Methods for integrated exposure monitoring of lead and cadmium. Environ. Res. 1991, 56, 78–89. [Google Scholar] [CrossRef]
- Kabata-Pendias, A.; Pendias, H. Trace Elements in Soils and Plants, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2001; p. 331. [Google Scholar]
- Muramoto, S.; Aoyama, I. Effects of fertilizers on the vicissitude of cadmium in rice plant. J. Environ. Sci. Health Part A Environ. Sci. Eng. Toxicol. 1990, 25, 629–636. [Google Scholar] [CrossRef]
- Radwan, M.A.; Salama, A.K. Market basket survey for some heavy metals in Egyptian fruits and vegetables. Food Chem. Toxicol. 2006, 44, 1273–1278. [Google Scholar] [CrossRef]
- Steenland, K.; Boffetta, P. Lead and cancer in humans: Where are we now? Am. J. Ind. Med. 2000, 38, 295–299. [Google Scholar] [CrossRef]
- Hossain, M.A.; Hasanuzzaman, M.; Fujita, M. Up-regulation of antioxidant and glyoxalase systems by exogenous glycinebetaine and proline in mung bean confer tolerance to cadmium stress. Physiol. Mol. Biol. Plants 2010, 16, 259–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.L.; Lee, I.-J. Endophytic Penicillium funiculosum LHL06 secretes gibberellin that reprograms Glycine max L. growth during copper stress. BMC Plant Biol. 2013, 13, 86–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo, R.A.; Gratão, P.L.; Monteiro, C.C.; Carvalho, R.F. What is new in the research on cadmium-induced stress in plants? Food Energy Secur. 2012, 1, 133–140. [Google Scholar] [CrossRef]
- Llamas, A.; Ullrich, C.I.; Sanz, A. Cd2+ effects on transmembrane electrical potential difference, respiration and membrane permeability of rice (Oryza sativa L) roots. Plant Soil 2000, 219, 21–28. [Google Scholar] [CrossRef]
- Larbi, A.; Morales, F.; Abadia, A.; Gogorcena, Y.; Lucena, J.J.; Abadia, J. Effects of Cd and Pb in sugar beet plants grown in nutrient solution: Induced Fe deficiency and growth inhibition. Funct. Plant Biol. 2002, 29, 1453–1464. [Google Scholar] [CrossRef]
- Khanmirzaei., A.; Bazargan, K.; Moezzi, A.A.; Richards, B.K.; Shahbazi, K. Single and Sequential Extraction of Cadmium in Some Highly Calcareous Soils of Southwestern Iran. J. Soil Sci. Plant Nutr. 2013, 13, 153–164. [Google Scholar] [CrossRef]
- Greger, M.; Brammer, E.; Lindberg, S.; Larsson, G.; Idestam-almquist, J. Uptake and physiological effects of cadmium in sugar beet (Beta vulgaris) related to mineral provision. J. Exp. Bot. 1991, 42, 729–737. [Google Scholar] [CrossRef]
- Danso Marfo, T.; Datta, R.; Vranová, V.; Ekielski, A. Ecotone Dynamics and Stability from Soil Perspective: Forest-Agriculture Land Transition. Agriculture 2019, 9, 228. [Google Scholar] [CrossRef] [Green Version]
- Marfo, T.D.; Datta, R.; Pathan, S.I.; Vranová, V. Ecotone Dynamics and Stability from Soil Scientific Point of View. Diversity 2019, 11, 53. [Google Scholar] [CrossRef] [Green Version]
- Yadav, G.S.; Datta, R.; Imran Pathan, S.; Lal, R.; Meena, R.S.; Babu, S.; Das, A.; Bhowmik, S.; Datta, M.; Saha, P. Effects of conservation tillage and nutrient management practices on soil fertility and productivity of rice (Oryza sativa L.)–rice system in north eastern region of India. Sustainability 2017, 9, 1816. [Google Scholar] [CrossRef] [Green Version]
- Lukin, S.V.; Selyukova, S.V. Ecological Assessment of the Content of Cadmium in Soils and Crops in Southwestern Regions of the Central Chernozemic Zone, Russia. Eurasian Soil Sci. 2018, 51, 1547–1553. [Google Scholar] [CrossRef] [Green Version]
- Burd, G.I.; Dixon, D.G.; Glick, B.R. A plant growth-promoting bacterium that decreases nickel toxicity in seedlings. Appl. Environ. Microbiol. 1998, 64, 3663–3668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafar-ul-Hye, M.; Shahjahan, A.; Danish, S.; Abid, M.; Qayyum, M.F. Mitigation of cadmium toxicity induced stress in wheat by ACC-deaminase containing PGPR isolated from cadmium polluted wheat rhizosphere. Pak. J. Bot. 2018, 50, 1727–1734. [Google Scholar]
- Danish, S.; Kiran, S.; Fahad, S.; Ahmad, N.; Ali, M.A.; Tahir, F.A.; Rasheed, M.K.; Shahzad, K.; Li, X.; Wang, D.; et al. Alleviation of chromium toxicity in maize by Fe fortification and chromium tolerant ACC deaminase producing plant growth promoting rhizobacteria. Ecotoxicol. Environ. Saf. 2019, 185, 109706. [Google Scholar] [CrossRef]
- Arshad, M.; Frankenberger, W.T.J. Ethylene: Agricultural Sources and Applications; Kluwer Academic Publishers: New York, NY, USA, 2002. [Google Scholar]
- Penrose, D.M.; Glick, B.R. Enzymes that regulate ethylene levels—1-Aminocyclopropane-1-carboxylic acid (ACC) deaminase, ACC synthase and ACC oxidase. Indian J. Exp. Biol. 1997, 35, 1–17. [Google Scholar]
- Yang, J.; Kloepper, J.W.; Ryu, C.M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef]
- Zafar-ul-Hye, M.; Danish, S.; Abbas, M.; Ahmad, M.; Munir, T.M. ACC deaminase producing PGPR Bacillus amyloliquefaciens and agrobacterium fabrum along with biochar improve wheat productivity under drought stress. Agronomy 2019, 9, 343. [Google Scholar] [CrossRef] [Green Version]
- Danish, S.; Zafar-ul-Hye, M. Co-application of ACC-deaminase producing PGPR and timber-waste biochar improves pigments formation, growth and yield of wheat under drought stress. Sci. Rep. 2019, 9, 5999. [Google Scholar] [CrossRef] [Green Version]
- Jalali, J.; Gaudin, P.; Capiaux, H.; Ammar, E.; Lebeau, T. Isolation and screening of indigenous bacteria from phosphogypsum-contaminated soils for their potential in promoting plant growth and trace elements mobilization. J. Environ. Manag. 2020, 260, 110063. [Google Scholar] [CrossRef]
- Brtnicky, M.; Dokulilova, T.; Holatko, J.; Pecina, V.; Kintl, A.; Latal, O.; Vyhnanek, T.; Prichystalova, J.; Datta, R. Long-Term Effects of Biochar-Based Organic Amendments on Soil Microbial Parameters. Agronomy 2019, 9, 747. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.A.; Hussain, I.; Rasheed, R.; Iqbal, M.; Riaz, M.; Arif, M.S. Advances in microbe-assisted reclamation of heavy metal contaminated soils over the last decade: A review. J. Environ. Manag. 2017, 198, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Danish, S.; Zafar-ul-hye, M.; Mohsin, F.; Hussan, M. ACC-deaminase producing plant growth promoting rhizobacteria and biochar mitigate adverse effects of drought stress on maize growth. PLoS ONE 2020, 15, e0230615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danish, S.; Zafar-Ul-Hye, M. Combined role of ACC deaminase producing bacteria and biochar on cereals productivity under drought. Phyton 2020, 89, 217–227. [Google Scholar] [CrossRef]
- Parewa, H.P.; Meena, V.S.; Jain, L.K.; Choudhary, A. Sustainable crop production and soil health management through plant growth-promoting rhizobacteria. In Role of Rhizospheric Microbes in Soil: Stress Management and Agricultural Sustainability; Springer: Singapore, 2018; Volume 1, pp. 299–329. [Google Scholar]
- Pathan, S.I.; Větrovský, T.; Giagnoni, L.; Datta, R.; Baldrian, P.; Nannipieri, P.; Renella, G. Microbial expression profiles in the rhizosphere of two maize lines differing in N use efficiency. Plant Soil 2018, 433, 401–413. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-Ul-Hye, M.; Hussain, S.; Riaz, M.; Qayyum, M.F. Mitigation of drought stress in maize through inoculation with drought tolerant ACC deaminase containing PGPR under axenic conditions. Pak. J. Bot. 2020, 52, 49–60. [Google Scholar] [CrossRef]
- Zafar-Ul-Hye, M.; Zahra, M.B.; Danish, S.; Abbas, M.; Rehim, A.; Akbar, M.N.; Iftikhar, A.; Gul, M.; Nazir, I.; Abid, M.; et al. Multi-strain inoculation with pgpr producing acc deaminase is more effective than single-strain inoculation to improve wheat (Triticum aestivum) growth and yield. Phyton 2020, 89, 405–413. [Google Scholar] [CrossRef]
- Glick, B.; Penrose, D.; Li, J. A Model for the Lowering of Plant Ethylene Concentrations by Plant Growth-promoting Bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [CrossRef]
- Ahmed, N.; Ahsen, S.; Ali, M.A.; Hussain, M.B.; Hussain, S.B.; Rasheed, M.K.; Butt, B.; Irshad, I.; Danish, S. Rhizobacteria and silicon synergy modulates the growth, nutrition and yield of mungbean under saline soil. Pak. J. Bot. 2020, 52, 9–15. [Google Scholar] [CrossRef]
- Miniraj, N.; Prasanna, K.P.; Peter, K.V. Bitter gourd Momordica spp. Genet. Improv. Veg. Plants 1993, 239–246. [Google Scholar] [CrossRef]
- Lea Lojkova, V.V. Pavel Formánek, Ida Drápelová, Martin Brtnicky, Rahul Datta Enantiomers of Carbohydrates and Their Role in Ecosystem Interactions: A Review. Symmetry 2020, 12, 470. [Google Scholar] [CrossRef] [Green Version]
- 6 Benefits of Bitter Melon (Bitter Gourd) and Its Extract. Available online: https://www.healthline.com/nutrition/bitter-melon#section8 (accessed on 27 May 2020).
- GOP. Fruits, Vegetables and Condiments: Statistics of Pakistan; Ministry of National Food Security and Research (Economic Wing): Islamabad, Pakistan, 2014.
- Dworkin, M.; Foster, J.W. Experiments with some microorganisms which utilize ethane and hydrogen. J. Bacteriol. 1958, 75, 592–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadiq, A.; Ali, B. Growth and yield enhancement of Triticum aestivum L. by rhizobacteria isolated from agronomic plants. Aust. J. Crop Sci. 2013, 7, 1544–1550. [Google Scholar]
- Chapman, H.D.; Pratt, P.F. Methods of Analysis for Soils, Plants and Water; University of California, Division of Agricultural Sciences: Berkeley, CA, USA, 1961. [Google Scholar]
- Jones, J.B.; WolfH, B.; Mills, H.A. Plant Analysis Handbook: A Practical Sampling, Preparation, Analysis, and Interpretation Guide; Micro-Macro Publishing Inc.: Athens, GA, USA, 1991. [Google Scholar]
- Nadeem, F.; Ahmad, R.; Rehmani, M.I.A.; Ali, A.; Ahmad, M.; Iqbal, J. Qualitative and Chemical Analysis of Rice Kernel to Time of Application of Phosphorus in Combination with Zinc Under Anaerobic Conditions. Asian J. Agric. Biol. 2013, 1, 67–75. [Google Scholar]
- Bremner, M. Chapter 37: Nitrogen-Total. In Methods of Soil Analysis: Part 3 Chemical Methods; American Society of Agronomy: Madison, WI, USA, 1996; pp. 1085–1122. [Google Scholar]
- Roth, E.; Mancier, V.; Fabre, B. Adsorption of cadmium on different granulometric soil fractions: Influence of organic matter and temperature. Geoderma 2012, 189–190, 133–143. [Google Scholar] [CrossRef]
- Papoyan, A.; Kochian, L.V. Identification of Thlaspi caerulescens genes that may be involved in heavy metal hyperaccumulation and tolerance. Characterization of a novel heavy metal transporting ATPase. Plant Physiol. 2004, 136, 3814–3823. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R.; Patten, C.L.; Holguin, G.; Penrose, D.M. Biochemical and Genetic Mechanisms Used by Plant Growth Promoting Bacteria; Imperial College Press: London, UK, 1999. [Google Scholar]
- Ouzounidou, G.; Ciamporova, M.; Moustakas, M.; Karataglis, S. Responses of Maize (Zea-mays L) Plants to Copper Stress.1. Growth, Mineral-Content and Ultrastructure of Roots. Environ. Exp. Bot. 1995, 35, 167–176. [Google Scholar] [CrossRef]
- De Filippis, L.F.; Hampp, R.; Ziegler, H. The effects of sublethal concentrations of zinc, cadmium and mercury on Euglena. Arch. Microbiol. 1981, 128, 407–411. [Google Scholar] [CrossRef]
- Matile, P.; Schellenberg, M.; Vicentini, F. Planta Localization of chlorophyllase in the chloroplast envelope. Planta 1997, 201, 96–99. [Google Scholar] [CrossRef] [Green Version]
- Panković, D.; Plesničar, M.; Arsenijević-Maksimović, I.; Petrović, N.; Sakač, Z.; Kastori, R. Effects of nitrogen nutrition on photosynthesis in Cd-treated sunflower plants. Ann. Bot. 2000, 86, 841–847. [Google Scholar] [CrossRef] [Green Version]
- Jalloh, M.A.; Chen, J.; Zhen, F.; Zhang, G. Effect of different N fertilizer forms on antioxidant capacity and grain yield of rice growing under Cd stress. J. Hazard. Mater. 2009, 162, 1081–1085. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.S.; Dietz, K.J. The Significance of Amino Acids and Amino Acid derived Molecules in Plant Responses and Adaptation to Heavy Metal Stress. J. Exp. Bot. 2006, 57, 711–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarwar, N.; Saifullah, S.M.; Malhi, S.S.; Zia, M.H.; Naeem, A.; Bibia, S.; Farida, G. Role of mineral nutrition in minimizing cadmium accumulation by plants. J. Sci. Food Agric. 2010, 90, 925–937. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhao, S.C.; Liu, R.L.; Zhou, W.; Jin, J.Y. Changes of photosynthetic activities of maize (Zea mays L.) seedlings in response to cadmium stress. Photosynthetica 2009, 47, 277–283. [Google Scholar] [CrossRef]
- Shen, W.; Nada, K.; Tachibana, S. Involvement of polyamines in the chilling tolerance of cucumber cultivars. Plant Physiol. 2000, 124, 431–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, R.; Kelkar, A.; Baraniya, D.; Molaei, A.; Moulick, A.; Meena, R.; Formanek, P. Enzymatic degradation of lignin in soil: A review. Sustainability 2017, 9, 1163. [Google Scholar] [CrossRef] [Green Version]
- Datta, R.; Anand, S.; Moulick, A.; Baraniya, D.; Pathan, S.I.; Rejsek, K.; Vranova, V.; Sharma, M.; Sharma, D.; Kelkar, A.; et al. How enzymes are adsorbed on soil solid phase and factors limiting its activity: A Review. Int. Agrophys. 2017, 31, 287–302. [Google Scholar] [CrossRef]
- Tripathi, M.; Munot, H.P.; Shouche, Y.; Meyer, J.M.; Goel, R. Isolation and functional characterization of siderophore-producing lead- and cadmium-resistant Pseudomonas putida KNP9. Curr. Microbiol. 2005, 50, 233–237. [Google Scholar] [CrossRef]















© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zafar-ul-Hye, M.; Naeem, M.; Danish, S.; Fahad, S.; Datta, R.; Abbas, M.; Rahi, A.A.; Brtnicky, M.; Holátko, J.; Tarar, Z.H.; et al. Alleviation of Cadmium Adverse Effects by Improving Nutrients Uptake in Bitter Gourd through Cadmium Tolerant Rhizobacteria. Environments 2020, 7, 54. https://0-doi-org.brum.beds.ac.uk/10.3390/environments7080054
Zafar-ul-Hye M, Naeem M, Danish S, Fahad S, Datta R, Abbas M, Rahi AA, Brtnicky M, Holátko J, Tarar ZH, et al. Alleviation of Cadmium Adverse Effects by Improving Nutrients Uptake in Bitter Gourd through Cadmium Tolerant Rhizobacteria. Environments. 2020; 7(8):54. https://0-doi-org.brum.beds.ac.uk/10.3390/environments7080054
Chicago/Turabian StyleZafar-ul-Hye, Muhammad, Muhammad Naeem, Subhan Danish, Shah Fahad, Rahul Datta, Mazhar Abbas, Ashfaq Ahmad Rahi, Martin Brtnicky, Jiří Holátko, Zahid Hassan Tarar, and et al. 2020. "Alleviation of Cadmium Adverse Effects by Improving Nutrients Uptake in Bitter Gourd through Cadmium Tolerant Rhizobacteria" Environments 7, no. 8: 54. https://0-doi-org.brum.beds.ac.uk/10.3390/environments7080054