
The Brown Seaweeds of Scotland, Their Importance and Applications
by
 , ,
, ,
Omar Al-Dulaimi
1, 
Mostafa E. Rateb
1,
Andrew S. Hursthouse
1 ,
,
Gary Thomson
2 and
Mohammed Yaseen
1,* 1
School of Computing, Engineering & Physical Sciences, University of the West of Scotland, Paisley PA1 2BE, UK
2
Odour & Dust Solutions, Standalane, Ayrshire KA3 5BG, UK
*
Author to whom correspondence should be addressed.
Environments 2021, 8(6), 59; https://0-doi-org.brum.beds.ac.uk/10.3390/environments8060059
Submission received: 22 April 2021
/
Revised: 16 June 2021
/
Accepted: 17 June 2021
/
Published: 21 June 2021
Abstract
:More than 50% of the UK coastline is situated in Scotland under legislative jurisdiction; therefore, there is a great opportunity for regionally focused economic development by the rational use of sustainable marine bio-sources. We review the importance of seaweeds in general, and more specifically, wrack brown seaweeds which are washed from the sea and accumulated in the wrack zone and their economic impact. Rules and regulations governing the harvesting of seaweed, potential sites for harvesting, along with the status of industrial application are discussed. We describe extraction and separation methods of natural products from these seaweeds along with their phytochemical profiles. Many potential applications for these derivatives exist in agriculture, energy, nutrition, biomaterials, waste treatment (composting), pharmaceuticals, cosmetics and other applications. The chemical diversity of the natural compounds present in these seaweeds is an opportunity to further investigate a range of chemical scaffolds, evaluate their biological activities, and develop them for better pharmaceutical or biotechnological applications. The key message is the significant opportunity for the development of high value products from a seaweed processing industry in Scotland, based on a sustainable resource, and locally regulated.
1. Introduction
Research and development efforts to harvest all forms of Scottish renewable resources are supported by many stakeholders. The potential advantage of seaweed harvesting as a renewable resource is an economic opportunity for Scotland given the importance of these seaweeds as a source of biopolymers, food supplements, minerals and vitamins, and their role as soil conditioners and composting accelerators and the devolved legislative and regulatory responsibility. In this review, we present an overview of an economically important group of the seaweeds: the brown seaweeds that live attached to rocks and are widely distributed in Scotland. Their harvesting, chemical derivatives, and uses will also be discussed. Macroalgae are organisms living in aquatic environments; they use the light to produce energy, and they are macroscopic, multicellular organisms [1] living independently or with other organisms. When they inhabit seas, they are resistant to harsh environmental conditions, and they can adapt themselves to live in different salinity, pH, and water depths [2,3,4,5].
Algae exist in multicellular forms, and where they can be seen by the naked eye, they are described as macroalgae, with less visible forms being known as microalgae. Macroalgae are a taxonomically diverse group that vary in morphology and pigmentation. They can be classified by colour into red (Rhodophyta), brown (Phaeophyta), and green (Chlorophyta or green algae) or by size into macroalgae and microalgae [3,6]. The pigments fucoxanthin, chlorophyll a and b, and phycobilins were isolated from brown, green, and red algae, respectively, and are considered to be responsible for the different algal colours [6,7].
Scotland has over 11,000 km of coastline which makes up more than half of the coastline of the whole of the UK and is one of the largest masses of deep water coast in Europe, with its depth reaching up to 30 meters [8,9,10,11]. This makes the marine industry a matter of high interest in Scotland’s economy, and the development of the aquaculture industry is crucial for creating new jobs, improving the social and environmental conditions in coastal areas, and future economic impact in Scotland [12,13].
Data recorded revealed that more than 90% of the UK aquaculture production is in Scotland, with fish farming supporting around 2300 jobs in Scotland [14]. The marine cultivation industry contributes to the food chain; it is an important pillar for commercial fisheries, conserves endangered species, and maintains the seashore’s community [15,16,17]. In the Scottish territories, the production from aquatic cultures is concentrated along the western shores and islands where favourable environmental conditions are present [18].
Herein, we discuss the distribution of the rock/brown seaweed in Scotland, the biodiversity in their habitat, and their ecological importance. The procedures, rules, and regulations embedded by the Scottish Government to control the irrational harvesting activities of the seaweeds in Scotland are explained. An important perspective on biotechnological applications of seaweed in waste management in the UK is also covered, with added emphasis given to the challenges of biomass treatment for the waste management sector. Additionally, we highlight the use of edible algae as food supplements and as successful alternatives to animal proteins. Finally, the phytochemical composition and some of the important secondary metabolites produced by seaweed are described and discussed.
1.1. The Distribution of Brown Seaweeds in Scotland
One of the important seaweeds in the coastal areas of Scotland are the rockweeds, which are large brown seaweeds of the taxonomic order Fucales. Figure 1 illustrates the taxonomic order of these seaweeds. Fucus vesiculosus, or bladder wrack, is a brown seaweed that is distributed all over the rocky shores of the UK. This type of alga is usually collected from the middle tide zone or mid-littoral zone, where it grows at high density. The leaf or leaf-like part of F. vesiculosus can be found to grow up to 2 meters in length, particularly in the north-eastern rocky coastline of Maine, USA [19]. Its usual habitat is shared with Ascophyllum nodosum between Fucus serratus and Fucus spiralis in the higher shore [20]. The algae have binary branching growth stalks with air-filled pods and smooth fronds. The male and female plants grow to 20 cm length on average with a regeneration rate of 0.5 cm per week; its life span is 2–5 years. However, the growth rate varies from location to location but increases with warmer temperatures and more light, especially between June and August [21]. F. vesiculosus is used as a source of energy, food, and health supplement, and as skin care products. Its decoction produces a vapour with benefits in the treatment of rheumatoid arthritis and hypertension, and it can also be used as a tonic drink [22,23,24].
F. serratus, or toothed wrack, inhabits the lower shore around the rocky coast. It is around 60 cm long and has toothed fronds, with pinpoint white hairs. This brown alga has no air vesicles and it is distributed around most of the UK coastline [20]. The plant grows at a rate of 0.2 cm/week, which is higher in summer compared to other times, and they cover the shore at a higher density reaching 10–20 plants per m2. Its 2–5 years life span is comparable to that of F. vesiculosus. This alga is used as soil conditioner and is industrially processed for the production of alginate [25].
F. spiralis, or the spiral wrack, has twisted fronds and naturally inhabits the intertidal zone usually on the rock of the high tide, which is only flooded during high tide. The spiral rick is adapted to live in low moisture conditions and can grow up to 40 cm long; it does not have air bladders and lives for up to 4 years. It is distributed around all the UK coasts below the zone of Pelvetia canaliculata. The twisted fronds give this seaweed the name spiralis. F. spiralis prefers rocks with cracks that can effectively protect their zygotes [26,27,28].
Ascophyllum nodosum, or knotted wrack, that predominantly distributed in sheltered rocks. This seaweed has long fronds supported by air bulbs to support its floating. The fronds are relatively long and can reach up to 200 cm. An epiphytic algae Polysiphonia lanosa usually lives as clusters adhered to the fronds of A. nodosum. Knotted wrack often shares the habitant with F. vesiculosus [20]. As previously mentioned, the knotted wrack is distributed all over the shores of the UK. The var. mackayi is distributed in the west coast of Scotland; this has binary-branched fronds with few air sacs [29,30,31]. Mycosphaerella ascophylli is a fungus that lives attached to A. nodosum and has an important role not only in protecting the knotted wrack from dryness but also in mitigating the high salinity effect on it [32].
The holdfasts of A. nodosum are thought to persist for several decades from which new fronds regenerate [33]; the external cells’ layer is repeatedly stripped off in a phenomenon not detected in other seaweed [34,35]. A. nodosum prefers sheltered coasts, as high waves decrease its growth. A. nodosum can competitively eliminate F. vesiculosus from the natural habitat. The knotted rack usually lives longer than the genus Fucus with up to 10 years. The industries of alginates extraction, fertilisers from seaweeds, and seaweeds for dietary purposes in Scotland use A. nodosum as a main source. Data show that around 32,000 tonnes of A. nodosum is harvested in Scotland and Ireland, which threatens the rational use of natural resources and requires immediate interventions [36,37,38]. Pelvetia canaliculata, or the channelled wrack, is distributed in most areas of the UK and it can live up to one week out of water in the habitat in the high shore area just above its competitive seaweed, F. spiralis [39].
Himanthalia elongata, or thongweed, has strap-like long fronds, which may grow up to 200 cm. This seaweed can live up to two years and survives in most areas of the UK except the south east of England. It lives in the low shore area just below F. serratus and above Laminaria. Thongweed starts its life cycle as small button fronds then produces from 2–4 long strips reach up to 2 m in length. H. elongata is edible seaweed that is eaten in Ireland as fresh salad, dried herb, or pickled herb [24,40].
1.2. The Biodiversity of the Brown Seaweed Habitat and Their Ecological Importance
The importance of seaweeds is prominent, and they have a great impact on inshore and offshore areas, and coastlines of Scotland. They modify the marine climate and preserve the biodiversity. Laminaria hyperborea and other macroalgae play an essential role in the protection of life. These seaweeds can store more carbon and alleviate its free levels, and also diminish global warming’s negative effects such as higher acidity and lower oxygen levels. It is frequently observed that the global warming is one of the greatest threats to life on earth [41,42,43].
Seaweeds are an important link in the food chain as they serve as food and residence for a variety of organisms and many marine species [44,45]. The living organisms which grow on others such as plants and animals are detected on the surfaces of the algae fronds or blades and take their essential nutrients from the environment around them [46].
The structure which holds or anchors the algae as a sessile structure to the seabed is known as the holdfast, which is a habitat for a diversity of species of marine organisms such as invertebrates [47]. Different marine organisms have been detected living around the holdfast and other parts of algae, providing diversity, adding to the biomass of the ecosystem, and participating in the food cycle in aquatic media [48]. A single kelp can host about 8000 individual macroinvertebrates [47], and the diversity of the hosted organisms increases with the increase in volume of the algae as a larger area of habitat is offered [49]. Starfish, sea cucumbers, crabs, shrimp, and snails are examples of these invertebrates [47].
The species of algae can affect the number and diversity of organisms which are hosted near to their holdfast; if we compare L. hyperborea with Laminaria digitata, another kelp which is widely distributed in the UK [50], the former has longer and harder strips which protect their inhabitants more and are less affected by their blades, a property makes them more favourable than L. digitata [51,52]. To summarise, these simple differences in the structures of the kelp markedly affect the types and numbers of organisms hosted and the solidity of the seabed which holds the kelp, affecting both their distribution and their associated understorey gathering [53].
Algae provide an excellent protection strategy for several marine species, defending them against their natural predators [54]. The interaction between organisms (micro and macro) themselves and with their surroundings creates a dynamic ecosystem. Some studies revealed that the biochemical compounds produced by the seaweeds have antimicrobial effects in addition to their innate effect against herbivorous organisms [55,56,57]. Other studies documented that the microorganisms associated with some algae produce natural compounds that have antimicrobial effects [58,59].
The Water Frame Directive (WFD), which is an EU directive, commits its members to a highly standard code of practice during their activities in aquatic areas [60,61]. The WFD uses the status of seaweeds, their abundance, and the diversity in their habitat as an indicator of the marine ecological condition [62,63]. As the UK (and Scotland) withdraws from the EU, the consequence for detailed environmental considerations is unclear at the time of writing, but complex interdependencies on the regulation of marine resources exist (REF SG report in comment link). Large quantities of carbon are part of the dissolved CO2 content of surface aquatic systems and algae absorb a good proportion of that, making them play an important role in carbon sequestration; in addition, 50% of the photosynthesis processes are carried out by algae, and a lab experiment showed that algae showed a daily CO2 uptake equal to 159 mg per litre [64,65,66,67,68]. Other services provided by the seaweed to the ecosystem are protection from strong waves and stabilising the coastal sediments [62]. Taking the role and importance of seaweeds in controlling the marine ecosystem into account, the Scottish government and their civil service directorates performed a comprehensive study of the current environmental conditions and what the effect may be in disturbing the biomass of natural seaweeds in the Scottish coastal zone. Marine Scotland suggests that the uncontrolled harvesting of the seaweeds could lead to severe environmental problems and should be accomplished in a sustainable way [62].
Macroalgae have been harvested for many years for use as primary materials or as a source for bioactive materials by many pharmaceutical and food industries. Currently, in Scotland, this marine industry is small; however, expansion in the future is anticipated as industrial interest rises and a proposal to harvest large quantities of Laminaria hyperborea has been submitted to the Scottish Government. In [62], however opposition to this plan initiated by change.org is presented [69]. Therefore, what are the effects of seaweed harvesting on the natural seabed and shores of Scotland? To answer this question, we need to consider the following factors: the type of the seaweeds identified for harvesting, the method used for collection (manual or mechanical), the quantity taken, the timing of extraction, the geographical location, and the period of time left between harvests for regrowth [62,70]. The Scottish authorities considered all the points of discussion surrounding the subject of seaweeds harvesting and decided to implement a policy to relieve the negative impacts of seaweed harvesting on Scotland’s marine ecosystem [71].
To alleviate the negative outcomes of seaweed harvesting, several measures have been adapted to protect some areas from industrial harvesting and to ensure the sustainability of the resource. One of these measures is the restriction of harvesting in the following areas: wave-sensitive areas, areas of specific biological communities, hauling-out areas, wrecks areas, historical areas, and areas saved for crofters [62,70]. To ensure that industrial harvesting is performed appropriately to let the seaweed regrow and alleviate and negative ecological impacts, a concept of licencing is confirmed to protect Scotland’s marine environment, and to protect the rights of the harvesters, and other stakeholders [62,70].
2. Some of the Important Uses of the Seaweeds
2.1. Seaweed as Food Supplements
Seaweed are used as food for human beings, feed for animals, or as additives after collection, cleaning, and packing. Different processing stages of seaweed leads to the production of raw extracts or primary materials which enter the supply chains of pharmaceutical and cosmetics industries [3,72].
Algae have nutritional benefits and have been used for long time [72]. In western countries, the use of algae as food is limited, but it is in continuous development [73], and its use is focusing on the isolation of polymers for biotechnological and pharmaceutical applications [74]. Meanwhile, in East Asia, seaweed was used for a long time as a food, due to the high levels of proteins, vitamins, and minerals [75]. They are good candidates for animal feeding, and as food additives and supplements. The brown seaweed, A. nodosum, was incorporated in animal feed by a Canadian company to increase their immunity and combat microbial invasion [76,77,78].
Alginates and other polymers derived from algae find a place in the food, pharmaceuticals, and cosmetics industries. Usually, alginates and other polysaccharides polymers are used as emulsifying agents, thickening agents, gelling agents and food stabilisers in food industries [79,80,81]. Hydrocolloids such as alginates have the ability to stabilise oil in water emulsion by adsorption on the oil surfaces to prevent flocculation [82]. Propylene glycol alginate is an emulsifier of choice in the food industry and sometimes, it is preferred over methylcellulose [82]. The use of alginates and alginate derivatives is well represented in biotechnological applications and their use in drug delivery and the microencapsulation of pancreatic islets’ beta cells (β cells) is widely researched [83,84,85,86]. Alginate has interesting properties, as it is flexible, absorbs tensile stress, and it is edible. These properties have identified it as a green alternative to non-biodegradable food packaging [87]; unfortunately its porous texture makes it permeable to humidity and oxygen, which cause food deterioration [87]. These shortages in alginate’s role as food packaging is monitored by the incorporation of safe and food friendly antimicrobials to decrease the effects of microbes, minimise food deterioration, and prolong food life [88,89,90,91]. Additionally, the use of algae as a biofuel cannot be ignored [62]. Laminaria sp., and A. nodosum contain massive quantities of alginate, a property which makes them the first choice of alginate extraction for industrial purposes [92,93,94].
East Asian countries were recorded as the largest consumers of seaweed worldwide [95]. For example, the average consumption in Japan is around one and half kilograms per adult of dry seaweed annually [74,96]. The Japanese use more than one hundred species of seaweed [97]. A. nodosum is incorporated into a diet of overweight people to reduce the energy intake after consumption [98]; alginate has a unique characteristic of gel formation on contact with acids and calcium ions at low temperature. This property is utilised to induce glut after the ingestion of alginate [99,100]. Algae have high fibre contents, which make the digestion process in humans a complex process, while the ruminant animals’ digestive system has the required enzymatic machinery and it can digest the high fibre contents in the algal meal and increase the availability of nutrients [101]. The monogastric animals need preliminary digestive processing to obtain benefits from the proteins of seaweed [102]. In general, the proteins of red seaweed are digested better than brown seaweed and are comparable in levels to fruits and vegetables [103].
It is widely known that animal proteins make up the main contribution to human food protein intake. However, it is increasingly debated that the production of protein from animals could be reduced because of the ethical and environmental impacts of this part of the food chain [104]. However, the safety of using seaweed as food and animal feeding is under debate [105]. To mitigate the effects of the production of animal protein and prepare resilient strategies for future food supply shortages [106], we need to identify alternatives to the modern technologies introduced to intensify food production from plant and animal sources, thus helping to reduce output of greenhouse gases and detrimental impacts on the natural habitats and diversity of other species [107]. The identification of algae as a safe protein source can mitigate the negative impacts of protein production from animals and terrestrial plants [105] as the high levels of proteins, abundant in algae, are comparable to that in animal products such as meat and eggs [108].
Seaweed do not need fresh water, land for cultivation, or even special agricultural techniques, which make their cultivation, and harvesting more feasible [109,110,111]. This promotes the rational consumption of fresh water which is typically used to irrigate the crops with high protein contents [112]. Green microalgae (Chlorella spp.) have been cultivated for commercial use since 2004 [113,114]. Other studies were also interested in the cultivation of seaweeds for alginate production [115,116]. The main purpose of these cultivation activities is to find a suitable protein source which is safe and sustainable, and protein quantities in algae vary significantly depending on the species, geographical distribution, temperature, and time of the collection [117]. The collection of Ulva lactuca in August is preferred to collection in April as the proteins level is higher [118]. The highest quantities of proteins were recorded from the red macroalgae and the lowest levels from brown seaweeds [74].
2.2. Seaweeds Used in Green Waste Management
Compost, meeting the regulatory quality standard (e.g., PAS100) is an end of waste product produced from waste organic matter, and is formed by the decomposition of the biomass due to metabolic processes of the microbiota in situ [119]. The composting process for seaweed has been investigated as a way to limit the negative impact of excessive growth or accumulation of seaweed on tourist beaches and on trade at seaports where excessive growth may limit the movement of ships, etc. [120,121]. Strong performance over conventional compost media was observed, highlighting the speed to reach and duration of the thermophilic phase, stabilising waste within 4 months.
The use of seaweed-derived extract in the composting of solid bio-waste materials appears to enhance the production of useful microbiota (bacterial and fungal colonies). These microorganisms are the primary players in the composting process as they break down the organic biomass to a rich nutrient-enhanced compost. The composting process aids carbon sequestration and reduces methane release from landfills, reducing its carbon footprint. The potential use of seaweed in composting processes for green waste management would improve the sustainability of algae in Scotland and provide the beneficial application of material causing nuisance (on beaches and affecting water transport). However, we need to remember that the harvesting process must be undertaken in a controllable and well-designed manner as seaweed provides a significant ecosystem service. In addition, the use of algae in the composting of the green waste could reduce the time of composting, improving the productivity of the process. Other researchers revealed that the effect of the phosphorus and potassium provided by seaweed, in addition to the slightly alkaline media produced by algae, and better aeration of the soil improved the production of Licopersicon esculentum (Tomato) [121].
Algae have a relatively low C/N contents so the ideal composting process can be achieved by mixing a material with high carbon content such as trees, shrubs, etc. to reach the required 30 C/N mixture and kick off the composting process. Aerobic metabolism consumes the carbon of the biomass, the stabilisation stage of the compost is achieved at 15 C/N mixture [122]. Higher carbon contents in the waste biomass extends the time of composting; hence, the stabilisation process and achievement of higher fungal contents in the compost takes a longer time [123,124]. This means that higher fungal content of the compost is one of the signs that indicates compost stabilisation. On the other hand, the lower C/N contents metabolised faster and produced more ammonia as an undesirable product of the composting process [122].
The composting of marine-derived bio-waste may yield a compost with a high salt concentration that negatively affects the yield of salt-sensitive plants, so washing the seaweed could potentially reduce salt content, although it has been observed that the washing step is not really crucial as it does not decrease the intracellular salt content of the algal cells [120,125].
Seaweed-based treatments offer an economic and effective biological solution which activates and stabilises the decomposition of organic matter, reducing both the cost and time associated with widely used composting approaches to waste treatment. These treatments are also expected to have more widespread application to industrial “nuisance” (dusts and odours) and remediation options. BCx®, a compost accelerator from EKOGEA, contains 100% A. nodosum used to enhance the composting process and achieve a mature compost within 6 weeks. This compost accelerator provides nutrients to the microbiota and eliminates NH3 and H2S via ion exchange. Additionally, it is claimed that it interacts with potentially toxic elements and optimises moisture content during the composting process [126]. The use of seaweeds and their extracts in the acceleration of composting processes could present a great opportunity for the UK to decrease the resources used in green waste management.
Information revealed from the compost certification scheme “report 2018” from REAL (Renewable Energy Assurance Limited) indicated that projects using sustainable seaweed resources in the management of green waste are important for the benefits of many stakeholders: the company, the customers, farmers, compost makers, community councils, community, and the sustainability of marine resources. The REAL scheme ensures that the end users of compost and bio-fertilisers are utilising trusted products produced under a quality control system governed by assured controlled practice [127].
In 2018, around 179 certified producers of compost that are distributed throughout the UK dealt with around 3.5–5 million tonnes of waste materials and produced 1.86 million tonnes of compost. Approximately 76% of those processes were undertaken in England, 13% in Scotland, 7% in Wales and 4% in Northern Ireland. About 68% of all these processes were carried out in the open air, turned windrows, and more than 75% of them processed green waste [127] (summarised in Figure 2). All information about the certified compost producers, addresses, and contact details can be retrieved from the Renewable Energy Assurance Ltd. (REAL) website http://www.qualitycompost.org.uk/ (accessed on 22 April 2021) under the REAL’s Certification Scheme for quality composts section.
Compost feedstock varies between sites but is generally composed of green waste (grass, flowers, pruning, clippings, and leaves). Permitted industrial and animal by-product (ABP) wastes such as food waste are typically processed at in-vessel composting facilities. In Scotland, 70% of the certified composting operations only process green waste, as shown in Figure 3.
2.3. Role of Seaweeds in Agriculture
A. nodosum extract is added to the soil to serve as a bio-stimulant and increases ion exchange, the transportation of micronutrients, provides natural polysaccharides that may increase the vitality of the soil microbiota, and holds water in the soil. Natural compounds in the extract may act as plant growth promoters and increase the plants growth rate [128,129,130,131,132]. The addition of extracted seaweed is much better than addition of the dry one, because the transportation and exchange of materials is faster in the former. The process of preparing an extract from the seaweed to use as plant growth accelerator processes includes the use of a cold press where no heat or chemical agents are used. Others used chemical extraction active constituents for that purpose, and alkaline solutions and heat were used to extract enough natural materials for application in agriculture [133].
Seaweed extracts from A. nodosum have important effects on the growth rate and quantity of phytochemicals of two varieties of the broccoli cultivars. The quantities of phenolics, flavonoids, and isothiocyanates were increased in all tests compared to control, which led to the improvement of the quality of the plant under study [129]. A. nodosum is a seaweed extensively studied for its benefits for the soil and for the growth of cultivated plants, thus it is effectively used as fertiliser and soil conditioner in addition to its value as a nutritional additive, as well as its role in food supplementation for humans and as animal feed [134]. The extracts of the seaweeds are applied by various techniques: dipping of the roots, spray, or soil immersion to distribute the nutrients, minerals, and growth promoters [135]. After the addition of these extracts via different techniques, the plants become more resistant to biotic and abiotic stress [134,136]. It was also recognised that the production of specific compounds in plants under study increased, such as the antioxidants produced by spinach after treatment with seaweed extract [137].
3. Current and Future Seaweed Harvesting Activity in Scotland
As highlighted in the introduction, in Scotland, harvesting activities are limited to small batches contributing to food, pharmaceutical, and biotechnological industries. Although these activities are involved in the collection of brown, red, and green algae, A. nodosum and L. hyperborea are the main targets for harvesters as they contain the highest quantities of alginates and proteins [138]. There are seven official sites in Scotland that are licenced to harvest seaweeds for industrial purposes [62]. These places are shown in Figure 5 using a map downloaded from visit Scotland.com [139]. Some of the seaweeds such as Laminaria spp. are washed out of the marine environment to the shoreline. The seaweeds drift to the beach because of wind and tidal action, and have some economic value. They are gathered by local communities in islands and used to improve the soil conditions and as fertilisers in the Machair (From Scots Gaelic, meaning Lowlands), and the fertile lands on the northwest coastlines of Ireland and Scotland, particularly the Outer Hebrides [140,141].
In 2014, global seaweed production was around 28 million tonnes, the vast majority of which (96%) was attributable to aquaculture [142]. Nevertheless, in some regions, including the UK, the harvesting of ‘wild’ seaweeds is still an economically important practice. The diversity of harvesting practices and technologies reflects the habitat variability in which seaweeds are found. The harvesting of cultivated seaweed varies from manual collection to mechanical collection from intertidal zones and the use of cranes mounted on ships in deep water [143]. The harvest method collects total biomass using nets at the end of the season or to collect specific materials such as carrageenan, which is accumulated in older seaweeds, or to partially harvest only the new growth and leave other parts for regeneration and further re-harvesting [144,145]. Globally, five methods of seaweed harvesting are practiced. The first method uses scythes as a tool to assist in manual collection. The second uses the Norwegian dredger which has a head comprising a large toothed dredge which is actively dragged through kelp (L. hyperborea) beds. This harvests the grown, or in other words, the large L. hyperborea and leaves the smaller kelp to mature. The third tool uses a cutting technique instead of towing to harvest seaweed near the shore; this tool is named the Norwegian cutter and vessels are used to harvest A. nodosum. The fourth method is the collection of the shore-cast seaweeds by hand, and the mechanical gathering of the seaweeds using a caterpillar tractor [62]. In Scotland, the manual harvesting technique is mostly used for seaweed collection [146]. The application of mechanical devices to harvest is possible in some areas where it is feasible to use mechanical harvesting methods [53].
In the Outer Hebrides and Caithness, harvesters usually collect the brown seaweed A. nodosum from the intertidal zone by hand or machines. To enable sustainable harvesting and let the seaweed regenerate again, harvesters need to cut the stalk of A. nodosum quarter of a meter from the rock contact point. Other seaweeds are harvested by hand from Fife, Caithness, and Bute, leaving 15 cm of stalk attached to the rock for regeneration [62]. Around 30% of brown kelp is washed away from the seabed naturally and storms occasionally detach up to 90% of a local seaweed population and wash it ashore [147,148]. Recently, some marine companies in Scotland increased their activities in the market and proposed the potential harvesting of a large quantity of L. hyperborea.
Marine Biopolymers Limited is one of those companies which asked for a sustainable resource of L. hyperborea, a seaweed abundant in Scotland, and its quantities are estimated to hit the 20 million tonnes of wet weight [149]. This recent report by Burrows (2018) also documented the distribution of L. hyperborea in Scotland [149]. Although the industrial harvesting of L. hyperborea is relatively new in Scotland, it is a well-established industry in other European countries such as France, Iceland, and Norway [70,150]. This new industry needs a comprehensive overview of the measures to control the harvesting activities to alleviate the negative impact of seaweed harvesting on the environment. Of course, a benefit of this development would be to support coastal communities socially and economically but a monitoring plan in addition to the adaptation of this plan according to the essential needs and environmental necessities is an essential prerequisite to develop the seaweed harvesting activities for industrial purposes [70]. All reports concerned about the effects of harvesting of the seaweeds recommend avoiding the harvesting in the following areas: special areas of conservation, nature conservation marine protected areas containing kelp forest needs protection, and protected priority marine features [70,151,152].
The Scottish Government announced more than eighty species and habitats as priority marine features with protection imposed [153]. Those features are mostly of national and international priority. The GEN 9 Natural Heritage policy as a part of the Scottish marine plan clarified that the priority marine features must not be affected by the application of the development marine environmental plan [154]. Comprehensive information about the legislative framework and requirement for an environmental report can be extracted from a Wild Seaweed Harvesting report prepared by ABP mer, a company that provides consultation services [70]. These limitations mean that the process of seaweed harvesting in Scotland is controlled by many statutory rules, and harvesters need to take several approvals and design evaluation plans for the environmental effects of seaweed harvesting before starting their business [70,155,156].
The Crown Estate Scotland owns the territorial waters of Scotland which covers 13.8 miles from the coastlines. The Crown Estate Scotland leases or gives licence to beneficiaries [154]. All harvesters need prior permission to collect seaweed or shore-cast seaweed, and in addition to that, the Scottish Natural Heritage must approve the collection process of the seaweed from the site of special scientific interest or European designated sites as special areas of protection. The marine Scotland licencing operations team takes into consideration all the regulations and legal acts governing seaweeds harvesting, ensuring a sustainable policy is followed, a low carbon environment to control the environmental changes and a better economy for the societies living in the coastal areas of Scotland [154]. The authorities have the right to know from the harvesters the biomass available, the harvesters’ strategy for sustainable harvesting, and the measures adopted by the harvesters to implement and adapt harvesting processes [62,70]. The Marine Scotland Act 2010 and Marine and Coastal Access Act 2009 are followed by the Scottish authorities to ensure the implantation of Scotland’s National Marine Plan governs both the deep and shallow water areas of Scotland per under the EU Directive 2014/89/EU on nautical location designing (Marine 2015).
4. Phytochemistry of Seaweeds
Humans and animals have consumed multicellular algae or macroalgae because they contain abundant levels of proteins, carbohydrates, minerals, and vitamins [157,158]. These advantages, in addition to the advantages mentioned previously, make them a competitive alternative to plant for food and feed production [159]. The novel phytochemicals which are found in marine algae have unique stereochemistry, many of which are expected to be ‘natural leads’ to the discovery of various beneficial drugs [160] and, potentially, a wide array of other useful compounds [161] are worthy of being mentioned in this review. In the following section, we describe various metabolites of algae with their structures and some biological activities of the chemical structures were sketched using Chem Draw® professional from PerkinElmer. The most important extraction methods used to extract materials from algae are also discussed.
4.1. Overview of Some Seaweed Derived Phytochemicals and Their Biological Activity
F. vesiculosus is incorporated in many food supplements to supply our bodies with some nutrients, minerals and vitamins and in other natural formulations to control body weight [98,162,163]. InSea2 is a natural formulation commercially available in many countries, where it is used to control the blood sugar and metabolism of carbohydrates. This formulation is formed from a combination of two natural extracts from seaweeds F. vesiculosus and A. nodosum. These two natural extracts were recorded to have both α-amylase and α-glucosidase inhibition activities [164]. The pharmacological effects on α-amylase and α-glucosidase explained the role of F. vesiculosus and A. nodosum in the optimisation of blood glucose and inhibition of these two enzymes in vivo [164].
The consumption of two hard gelatine capsules containing 500 mg of F. vesiculosus produced some effects on glucose and insulin levels in blood; however, that effect was not significant enough to indicate that this seaweed might have a controlling mechanism on the postprandial insulin and sugar levels, and this study confirmed that F. vesiculosus is a safe seaweed that has no side effects [165]. The presence of polyphenols in F. vesiculosus give this seaweed an antioxidant property. Several nutraceutical products have been developed from F. vesiculosus, which is rich in dietary elements, fibres, and phenolic entities.
F. vesiculosus has a high concentration of the antioxidants phlorotannins that encouraged many scientists to investigate its antioxidant properties. A study tested the antioxidant capacity and polysaccharide composition of F. vesiculosus and compared these capacities and antioxidants’ effects with those of some common commercial nutraceuticals. The percentage of fibre was the highest in F. vesiculosus, being around 55% compared to the commercial nutraceuticals used in this study. Interestingly the antioxidant effect of F. vesiculosus was higher than the commercial extracts and fucoidan 1. It was confirmed that the phlorotannins are the active constituents exerting the antioxidant activities in the raw seaweed and in all other materials tested [166]. Table 1 shows several compounds from different seaweeds with their associated biological effects.
The fucoidan 1 extract has an important percentage of phenolic compounds, indicating that the antioxidants activities of the fucoidan 1 extract belongs to the effects of the polysaccharides and phenolic compounds combined together [206]. Fucoidan was detected in F. vesiculosus, F. serratus, and A. nodosum [207,208,209].
Some of the phytochemicals found widely spread in algae are the natural pigments carotenoids. These compounds are used as natural food colourings in many kinds of food [210]. Carotenoids are synthesised by algae via the photosynthesis pathway [211]. β-carotene 2 was detected in fresh F. vesiculosus following maceration with methylated spirit and several sequences of purification steps [212].
Fucoxanthin 3 is another carotenoid detected in F. vesiculosus, F. serratus, and A. nodosum [191]. Fucoxanthin 3 and its metabolite Fucoxanthinol 4 showed anticancer effects against several cancer cells [173]. F. vesiculosus and F. serratus biosynthesised isofucoxanthin 5 [176,190]. Isofucoxanthinol 6 is separated from F. vesiculosus [176]. Neoxanthin 7 is another carotenoid isolated from F. vesiculosus [174]. In general, carotenoids have several medicinal effects, as they reduce the hepatic and cardiovascular diseases in addition to their effects against cancers [213] and in addition to their effects in controlling of weight and insulin levels [214]. Figure 6 shows the structures of the compounds from 1–7.
Several benzene derivatives 8–20 have been isolated from a controlled cleavage process of polyphenolic compounds extracted from F. vesiculosus [178]. Compounds 21–24 mentioned in Table 1 are known to have potential antioxidant activities, and they are isolated from the endophyte fungus Epicoccum sp. derived from the brown algae F. vesiculosus [179]. Trans-4-Hydroxymellein 21 was isolated from Seimatosporium sp., an endophyte of Epilobium hirsutum [215]. 1,3,5-trimethoxybenzene 8 has a pleasant odour and gives sedative effects [216]. Figure 7 shows the compounds from 8–24.
Phytosterols present in brown seaweeds have a important medicinal effects in lowering total cholesterol and harmful low-density lipoproteins [182]. A galactolipid compound 27 and 3,7,11-Trimethyl-2-dodecen-1-ol 28 were detected from F. vesiculosus, and other glycolipids detected from F. vesiculosus are compounds 29 and 30 [185,186] Figure 8 shows the compounds from 25–30.
Three polyphenolic compounds from F. vesiculosus L. fucotriphlorethol A 31, trifucodiphlorethol A 32, and trifucotriphlorethol A 33 [187] Figure 9 shows the compounds from 31–33.
Carotenoids and flavonoids such as quercetin 34, curcumin 35, and low molecular weight phlorotannins were detected in F. vesiculosus extract [188]. The polar extracts extracted by water and ethanol from five rock seaweeds (F. vesiculosus, F. serratus, F. spiralis, A. nodosum, P. canaliculata) have α-amylase and α-glucosidase inhibitory activities that decreased the glycaemic response. A. nodosum is the most effective inhibitor of α-amylase among this set of seaweed [217,218].
F. vesiculosus was sequentially extracted with water at variable temperatures, and with HCl (0.1 M) and KOH (2 M). 40% of the fractions were soluble and composed mainly of sugars and uronic acids and small quantities of sulfate, proteins and polyphenols. The main sugars were fucose 36, glucose 37, galactose 38, and xylose 39. The most effective antioxidant fraction was the acidic fraction followed by alkali and aqueous fractions. The natural antioxidants, sulphated polysaccharides from brown seaweed, can be used as potential antioxidants in food supplements [189]. These food supplements have been tried in vivo on animals to prove their hypoglycaemic effects; however, that effect needs to be proved on human. Manufacturing companies indicated that these food supplements are contraindicated in people with thyroid hormone imbalance as they can alter the thyroid hormones levels, also they are contraindicated in cases of hypersensitivity to iodine or F. vesiculosus, bleeding, blood clots, kidney diseases, BP, diabetes, stroke or if the user is pregnant or breast feeder [219].
F. vesiculosus is sold commercially under different trade names and it is used for variable purposes, such as for anticoagulant effects, to improve the skin firmness by increasing collagen thickness, and to enhance the fibroblast expression of integrins, which increase collagen thickness. Increasing the thickness of the collagen seems to treat cellulite that is the herniation of subcutaneous fat within fibrous connective tissue that manifests as skin dimpling and nodularity, often on the pelvic region (specifically the buttocks), lower limbs, and abdomen, so the seaweed has cosmetic effects. F. serratus contains zeaxanthin 40 [190], chlorophyll c1 41 [177], and all-trans-fucoxanthin 42 [220]. Figure 10 shows the compounds from 34–42.
Some of the phytochemicals detected in the rock seaweeds are phlorotannins. Phlorotannins are polymers, complex in nature and formed of several monomers named phloroglucinol 43 [192,221]. It showed also antithrombotic and profibrinolytic effects [194]. The aqueous ethanolic and aqueous extract from F. spiralis, A. nodosum, and P. canaliculata were analysed by UPLC-MS. F. spiralis showed the lower molecular weight phlorotannins of less than six monomers, while the other two showed higher molecular weight phlorotannins [222]. Other compounds from F. spiralis are 7-methoxycoumarin 44 [223], 1-monooleoyl glycerol 45 [197], β-carotene 5,8-epoxide (Citroxanthin) 46 [224], and 5-hydroxyramulosin 47 [223]. Figure 11 shows the compounds from 43–47.
Aqueous ethanol was used for the extraction of A. nodosum, and the extract showed antidiabetic activity by testing the extracts for an α-glucosidase inhibitory effect and glucose uptake stimulatory activity [225]. The polyphenols in the extract were responsible for this effect, and this was proved by a bioassay-guided separation, and purification scheme. Streptozotocin-diabetic mice models were designed to test the antidiabetic effects of crude phenols extract, purified phenols fraction, and purified polysaccharide fraction from A. nodosum. The results showed that phenols extract, and phenols fraction improved the fasting glucose and cholesterol levels, and the purified phenols fraction was effective in the sucrose tolerance test as the glucose level was maintained as low. The phenols extract, purified phenol fraction, and the polysaccharide fraction showed antioxidant activities [226].
A. nodosum is a source of nutrients, trace elements and minerals, amino acids, vitamins, phyto-hormones, and natural antioxidant material (phlorotannins, fucosterol 26, and fucoxanthin 3); all these materials proved to improve the skin health. P. canaliculata extract improves the brightness of the skin [227]. L. digitata and A. nodosum are rich in vitamins, minerals, amino-acids, and essential fatty acids which maintain moisture and a polysaccharide named alginic acid 48 that locks in the moisture of the skin (Sarfati, 2017). The wrinkling or aging of the skin at cellular compartment is attributed to the level of oxidative stress, inflammation, and cell senescence. A. nodosum extract contains large quantities of phlorotannins that have an activity opposed to oxidative stress, inflammation, and senescence [228]. The role of macromolecules extracted from seaweed in drug delivery is well documented. The colloidal polysaccharide alginate 49 proved itself to be safe, non-immunogenic, biocompatible and biodegradable; these properties are important for a compound as an anticancer drug vehicle for treatment of cancers [202]. A. nodosum was extracted to yield ascophyllan (a sulphated polysaccharide) whose full structure is unavailable but composed mainly of the sugars fucose 36, xylose 39, glucuronic acid 50, and sulphated half-ester in approximately equal amounts. The composition is clearly different from the A-fucoidan, for which the structure is also not available and isolated from A. nodosum, while S-fucoidan 1 is isolated from Fucus vesiculosus. Fucoidan 1 has lower quantities of uronic 50 acid and xylose 39 than those of ascophyllan [203,229]. Finally, 1,2,3,5-tetrahydroxybenzol-2,5-disulfate ester 51 is detected from A. nodosum [204] and bis-fucopentaphlorethol A nonadecaactate 52 from H. elongata [205]. Figure 12 shows the compounds from 48-52.
Lectins are a group of proteins isolated from macroalgae or seaweeds, and they have many industrial applications, as they have high recognition abilities to carbohydrates without the induction of structural modifications. Their mediation in protein–protein interaction turns them into antivirus, biomarkers, or drug targeting candidates [230,231,232,233]. New research indicated that lectin could serve as an inhibitor to coronavirus [234].
4.2. Symbiotic Microorganisms
F. vesiculosus has some microbial populations that live with it. This seaweed has epibiotic biofilms that have the potential to control F. vesiculosus’ life and environment. The effect of different salt levels on these biofilms have been investigated, with non-living stones used as a control. The microbiota was analysed and sequenced using 454 pyrosequencing of 16S rRNA gene sequences. The results showed that salt level is important in the determination of the epiphytic and epilithic colonies and growth [235]. Phoma tropica is an endophyte separated from F. spiralis; it was cultivated on a large scale to yield two important compounds, 5-hydroxyramulosin 47 and 7-methoxycoumarin 44. Structures of these compounds were characterised by NMR and by single crystal X-ray diffraction for the novel 5-hydroxyramulosin 47 [223]. Several endophytes were isolated from A. nodosum; around 800 bacterial isolates were isolated from this seaweed, and some of them have polysaccharides hydrolytic activities [236]. We mentioned above that microorganisms play an important role in the composting process of green and food waste; probably, bacterial cells present in algae play a positive role in the digestion of the waste material and the composting process when the algae are used to promote the composting process.
5. Extraction Techniques for Algal Derived Compounds
5.1. Extraction of Proteins
Alginate 49 is a polymer that is primarily separated from the cell wall and the cells of L. hyperborea, Macrocystis pyrifera, and A. nodosum [237]. The algal cell wall contains polysaccharides macromolecules such as the alginates 49 in brown seaweed and carrageenan 53 in the red ones. Other polysaccharides such as laminarin 54, cellulose 55, xylan 56, galactan 57, and fucoidan 1, were also reported in seaweed; those compounds decrease the availability of the proteins and other molecules for the extraction solvents; nevertheless, this availability increased with the use of mechanical shredders, and ultrasound destructive waves [238,239]. Using ultrasound waves in an extraction process conserves cost, time, and energy. This technique is successfully used in the extraction of natural compounds, which are heat-sensitive [8,240]. Other additives such as buffers may be used in conjunction with the mechanical destructive methods; those materials were used primarily to keep the structure of the extracted materials such as proteins intact during extraction [241].
A useful combination of extraction methods is the use of polysaccharides enzymes like cellulase, xylanase, κ-carrageenase, and β-agarase with alkali [242]. Proteins are usually extracted by the effect of acid and bases followed by the chromatographic technique or ultracentrifugation; an example of this is the extraction of proteins from A. nodosum using acid–base extraction solvent [243]. Other adaptations such as homogenisers and osmotic shocks were also used to augment the extraction process of proteins from algae [238,244]. Pulsed electrical waves formed holes in the algal cell walls to increase the protein extraction [245,246]. The quantities of proteins extracted from Spirulina sp. were reportedly increased by more than 10% after the use of a pulsed electric field at 15 kV/cm and 100 kJ/kg [247]. The extraction of natural materials from algae using membrane technologies were used to separate algal cell wall macromolecules and bacteria by a method of microfiltration, then, proteins were collected by ultrafiltration, and salts by nanofiltration. In other words, a sequential separation technique, separating compounds from seaweed depending on the particle size of the compound, was adapted to extract most of chemicals available in the bio-matrix of the seaweed [248,249].
5.2. Microwave-Assisted Extraction
The application of electromagnetic radiation in the microwave frequency range is an effective method for the extraction of chemicals from the biomass. The microwave heating process plays its role efficiently in extraction, hydrolysis, and pyrolysis of the targeted tissues [250]. The temperature control in this kind of extraction processes is an important factor in defining the type of chemicals produced [251]. Phenolics are extracted by a hydro-alcoholic solvent using a mixture of water and methanol [252]; other work indicated that water or ethanol with water extracted more phenolic materials from brown algae than acetone mixed with 20 parts of water using solid liquid extraction [253]. The phenolic materials of A. nodosum was extracted using the microwave-assisted extraction method. Using a heating temperature of 110 °C for 15 min, the yield of the process was higher in quantity and the time of the process was shorter than the ordinary extraction methods that extends for 16 times longer [254].
5.3. Green Method to Extract Sodium Alginate by Ultra-Sonication
The extraction of chemicals from macroalgae can be achieved using ultrasound waves; this method in principle uses the ultrasound waves that vibrate the sample molecules and separate them from the biomass into solvent of extraction. To extract sodium alginate 49, the Japanese wireweed or Sargassum muticum seaweed were mixed as one part to 20 parts of water. The mixture then extracted in an ultrasonic bath at room temperature for half an hour using a high-frequency wave as a green method and avoiding the use of acid [255].
5.4. Extraction of Alginates Using Formaldehyde to Remove Phenolic Compounds
Sodium alginate 49 was separated from Sargassum natans after washing the seaweed several times to remove all unwanted debris and mud, and the biomass was mixed with 2% (w/v) of formaldehyde and stored overnight to remove excess phenolics escaped from the acidic treatment [256]. Formaldehyde was previously reported to have high affinity to phenolic compounds distributed in the cell walls of macroalgae [257,258]. The phenolic fraction was separated from the biomass and the biomass was treated later to yield sodium alginate 49, which is characterised using HPLC [258,259].
6. Characterization of Alginate
The analysis of the alginates 49 can be accomplished using MALDI-TOF [260], FTIR [255], and HPLC [258]. Alginic acid 48 is isolated from algae and industrially isolated from brown algae. It is a polysaccharide formed of two polymers, G and M. Those two forms (1→4)-β-D-mannuronic acid (M) 58 and (1→4)-α-L-guluronic acid (G) 59 units are distributed differently on different alginates 49 structures depending on the species, stage of growth, and geographical place [261,262,263]. Those macromolecules are important in the manufacturing of medicines, the cosmetics industry, food industry, and biotechnology [259]. Proton NMR spectroscopy is used in the determination of the composition and percent of G and M moieties in the structure of alginates [264]. The percentage of each block can affect the gelling activities of alginates 49 as more M blocks increase the elasticity of the gel [265]. The uniform distribution of GG blocks turns the alginate 49 into a rigid and brittle structure, while the MM blocks turn the alginate 49 into the soft structure as the MG mixture improve the flexibility of the alginate 49 matrix [266]. Figure 13 shows the structure of compounds from 53–59. Figure 14 shows the connection between G and M blocks in alginate.
7. Conclusions and Future Vision
Scotland’s large and deep-water seashore hosts a vast array of micro and macro organisms which form a huge reservoir of sustainable resources. These resources need proper management and an ambitious investment plan to deploy new projects, research, and develop associated business growth, supporting the job market and developing the coastal societies. With a global shift from animal proteins to green proteins, the seaweed inhabiting the Scottish seashores provides a great opportunity for sustainable healthy foods, which do not need land, or fresh water for cultivation.
Scotland is firmly driving towards a circular economy to decrease its carbon footprint. This creates more opportunities for scientists, small businesses, and ambitious investors to discover new things, and start profitable businesses. Sustainable exploitation opportunities do exist for this resource.
Author Contributions
All authors contributed to the conception and design of the review. Data collection and analysis were performed by O.A.-D. The first draft of the manuscript was written by O.A.-D. and all authors commented on previous versions of the manuscript. All authors contributed to the conceptualization of the manuscript. The original draft was written by O.A.-D. M.E.R., G.T., A.S.H., and M.Y. reviewed the draft. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Innovate UK KTN Knowledge Transfer Partnership KTP—No. 11510. between Odour and Dust Solutions Limited and the University of the West of Scotland.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to express our special thanks and gratitude to the KTP team, including Stuart McKay, Carol Thomson at UWS and Gerry Black from KTN Limited and Stephanie McInroy, and Odour and Dust Solutions Limited.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rebours, C.; Marinho-Soriano, E.; Zertuche-González, J.A.; Hayashi, L.; Vásquez, J.A.; Kradolfer, P.; Soriano, G.; Ugarte, R.; Abreu, M.H.; Bay-Larsen, I. Seaweeds: An opportunity for wealth and sustainable livelihood for coastal communities. J. Appl. Phycol. 2014, 26, 1939–1951. [Google Scholar] [CrossRef] [Green Version]
- Evangelista, V.; Barsanti, L.; Frassanito, A.M.; Passarelli, V.; Gualtieri, P. Algal Toxins: Nature, Occurrence, Effect and Detection; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Factories 2018, 17, 36. [Google Scholar] [CrossRef] [PubMed]
- van Ginneken, V. Some mechanism seaweeds employ to cope with salinity stress in the harsh euhaline oceanic environment. Am. J. Plant Sci. 2018, 9, 1191–1211. [Google Scholar] [CrossRef] [Green Version]
- Fernández, P.A.; Roleda, M.Y.; Rautenberger, R.; Hurd, C.L. Carbonic anhydrase activity in seaweeds: Overview and recommendations for measuring activity with an electrometric method, using Macrocystis pyrifera as a model species. Mar. Biol. 2018, 165, 1–12. [Google Scholar] [CrossRef]
- Baweja, P.; Sahoo, D. Classification of algae. In The Algae World; Springer: Berlin/Heidelberg, Germany, 2015; pp. 31–55. [Google Scholar]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Application of Novel Extraction Technologies for Bioactives from Marine Algae. J. Agric. Food Chem. 2013, 61, 4667–4675. [Google Scholar] [CrossRef] [PubMed]
- Vilkhu, K.; Mawson, R.; Simons, L.; Bates, D. Applications and opportunities for ultrasound assisted extraction in the food industry—A review. Innov. Food Sci. Emerg. Technol. 2008, 9, 161–169. [Google Scholar] [CrossRef]
- Davenport, J.; Davenport, J.L. Effects of shore height, wave exposure and geographical distance on thermal niche width of intertidal fauna. Mar. Ecol. Prog. Ser. 2005, 292, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Ounsley, J.P.; Gallego, A.; Morris, D.J.; Armstrong, J.D. Regional variation in directed swimming by Atlantic salmon smolts leaving Scottish waters for their oceanic feeding grounds—A modelling study. ICES J. Mar. Sci. 2020, 77, 315–325. [Google Scholar] [CrossRef]
- Scotland, M. Marine Scotland Information. Available online: http://marine.gov.scot/data/facts-and-figures-about-scotlands-sea-area-coastline-length-sea-area-sq-kms (accessed on 20 May 2021).
- Rae, G.H. Sea louse control in Scotland, past and present. Pest Manag. Sci. Former. Pestic. Sci. 2002, 58, 515–520. [Google Scholar] [CrossRef]
- Howard, P.M. Environment, Labour and Capitalism at Sea:‘Working the Ground’ in Scotland; Manchester University Press: Manchester, UK, 2017. [Google Scholar]
- Directorate, M.S. Scotland’s Marine Economic Statistics 2016. Available online: https://www.gov.scot/publications/scotlands-marine-economic-statistics/pages/7/ (accessed on 20 May 2021).
- Roberts, T.; Upham, P. Prospects for the use of macro-algae for fuel in Ireland and the UK: An overview of marine management issues. Mar. Policy 2012, 36, 1047–1053. [Google Scholar] [CrossRef]
- Rasmussen, R.S.; Morrissey, M.T. Marine biotechnology for production of food ingredients. Adv. Food Nutr. Res. 2007, 52, 237–292. [Google Scholar]
- Lucarini, M.; Zuorro, A.; Di Lena, G.; Lavecchia, R.; Durazzo, A.; Benedetti, B.; Lombardi-Boccia, G. Sustainable Management of Secondary Raw Materials from the Marine Food-Chain: A Case-Study Perspective. Sustainability 2020, 12, 8997. [Google Scholar] [CrossRef]
- Coull, J.R. Fish farming in the Highlands and Islands: Boom industry of the 1980s. Scott. Geogr. Mag. 1988, 104, 4–13. [Google Scholar] [CrossRef]
- Wippelhauser, G.S. Ecology and Management of Maine’s Eelgrass, Rockweeds, and Kelps; US Department of Commerce NOAA Coastal Services Center Library: Charleston, SC, USA, 1996. [Google Scholar]
- Hill, J.; White, N. Marine Life Information Network: Biology and Sensitivity Key Information Sub-programme. Available online: https://www.marlin.ac.uk/ (accessed on 20 April 2021).
- Carlson, L. Seasonal variation in growth, reproduction and nitrogen content of Fucus vesiculosus L. in the Öresund, Southern Sweden. Bot. Mar. 1991, 34, 447–454. [Google Scholar] [CrossRef]
- Morrissey, J.; Kraan, S.; Guiry, M.D. A Guide to Commercially Important Seaweeds on the Irish Coast; Irish Bord Iascaigh Mhara/Iirish Sea Fisheries Board: Dublin, Ireland, 2001. [Google Scholar]
- McLachlan, J. Seaweed Resources in Europe: Uses and Potential; Taylor & Francis: Boca Raton, FL, USA, 1992. [Google Scholar]
- Guiry, M.D.; Blunden, G. Seaweed Resources in Europe: Uses and Potential; John Wiley & Sons: Hoboken, NJ, USA, 1991. [Google Scholar]
- Williams, G.A. Seasonal variation in a low shore Fucus serratus (Fucales, Phaeophyta) population and its epiphytic fauna. Hydrobiology 1996, 326, 191–197. [Google Scholar] [CrossRef]
- Anderson, C.; Scott, G. The occurrence of distinct morphotypes within a population of Fucus spiralis. J. Mar. Biol. Assoc. UK 1998, 78, 1003–1006. [Google Scholar] [CrossRef]
- Bond, P.; Brown, M.; Moate, R.; Gledhill, M.; Hill, S.; Nimmo, M. Arrested development in Fucus spiralis (Phaeophyceae) germlings exposed to copper. Eur. J. Phycol. 1999, 34, 513–521. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Teng, D.; Zhang, Y.; Mao, R.; Xi, D.; Wang, J. Candidacidal mechanism of the arenicin-3-derived peptide NZ17074 from Arenicola marina. Appl. Microbiol. Biotechnol. 2014, 98, 7387–7398. [Google Scholar] [CrossRef] [PubMed]
- Chock, J.S.; Mathieson, A.C. Physiological ecology of Ascophyllum nodosum (L.) Le Jolis and its detached ecad scorpioides (Hornemann) Hauck (Fucales, Phaeophyta). Bot. Mar. 1979, 22, 21–26. [Google Scholar] [CrossRef]
- Gibb, D.C. The free-living forms of Ascophyllum nodosum (L.) Le Jol. J. Ecol. 1957, 45, 49–83. [Google Scholar] [CrossRef]
- Pereira, L.; Morrison, L.; Shukla, P.S.; Critchley, A.T. A concise review of the brown macroalga Ascophyllum nodosum (Linnaeus) Le Jolis. J. Appl. Phycol. 2020, 32, 1–24. [Google Scholar] [CrossRef]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum-based biostimulants: Sustainable applications in agriculture for the stimulation of plant growth, stress tolerance, and disease management. Front. Plant Sci. 2019, 10, 655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundene, O. Growth and Reproduction in Ascophyllum Nodosum (Phaeophyceae). Nor. J. Bot. 1973, 20, 249–255. [Google Scholar]
- Halat, L.; Galway, M.E.; Garbary, D.J. Cell wall structural changes lead to separation and shedding of biofouled epidermal cell wall layers by the brown alga Ascophyllum nodosum. Protoplasma 2020, 257, 1–13. [Google Scholar] [CrossRef]
- Filion-Myklebust, C. Epidermis shedding in the brown seaweed Ascophyllum nodosum (L.) Le Jolis and its ecological significance. Mar. Biol. Lett. 1981, 2, 45–51. [Google Scholar]
- Boaden, P.J.; Dring, M. A quantitative evaluation of the effects of Ascophyllum harvesting on the littoral ecosystem. Helgoländer Meeresunters. 1980, 33, 700–710. [Google Scholar] [CrossRef] [Green Version]
- Hill, J. Ascophyllum Nodosum. Knotted Wrack. In Marine Life Information Network: Biology and Sensitivity Key Information Reviews [on-line]; Tyler-Walters, H., Hiscock, K., Eds.; Marine Biological Association of the United Kingdom: Plymouth, UK, 2008. [Google Scholar] [CrossRef]
- Bush, L.; Davies, A.; Maggs, C.A.; Yesson, C.; Brodie, J.A. A Review for the Crown Estate July 2013. Available online: https://macroalgalresearchgroupcom.files.wordpress.com/2017/03/bush_2013_reviewofloss.pdf (accessed on 20 April 2021).
- White, N. Pelvetia Canaliculata. Channelled Wrack. In Marine Life Information Network: Biology and Sensitivity Key Information Reviews [on-line]; Tyler-Walters, H., Hiscock, K., Eds.; Marine Biological Association of the United Kingdom: Plymouth, UK, 2008. [Google Scholar]
- Stengel, D.; Wilkes, R.; Guiry, M. Seasonal growth and recruitment of Himanthalia elongata Fucales, Phaeophycota) in different habitats on the Irish west coast. Eur. J. Phycol. 1999, 34, 213–221. [Google Scholar] [CrossRef]
- De Schryver, A.M.; Brakkee, K.W.; Goedkoop, M.J.; Huijbregts, M.A. Characterization Factors for Global Warming in Life Cycle Assessment Based on Damages to Humans and Ecosystems; ACS Publications: Washington, DC, USA, 2009. [Google Scholar]
- Gårdmark, A.; Huss, M. Individual variation and interactions explain food web responses to global warming. Philos. Trans. R. Soc. B 2020, 375, 20190449. [Google Scholar] [CrossRef]
- Botkin, D.B.; Saxe, H.; Araujo, M.B.; Betts, R.; Bradshaw, R.H.; Cedhagen, T.; Chesson, P.; Dawson, T.P.; Etterson, J.R.; Faith, D.P. Forecasting the effects of global warming on biodiversity. Bioscience 2007, 57, 227–236. [Google Scholar] [CrossRef]
- Egan, S.; Harder, T.; Burke, C.; Steinberg, P.; Kjelleberg, S.; Thomas, T. The seaweed holobiont: Understanding seaweed–bacteria interactions. Fems Microbiol. Rev. 2013, 37, 462–476. [Google Scholar] [CrossRef] [Green Version]
- Suryanarayanan, T. Fungal endosymbionts of seaweeds. In Biology of Marine Fungi; Springer: Berlin/Heidelberg, Germany, 2012; pp. 53–69. [Google Scholar]
- Nybakken, J.W. Marine Biology: An Ecological Approach; Benjamin Cummings: San Francisco, CA, USA, 2001. [Google Scholar]
- Burrows, M.T.; Smale, D.; Connor, N.O.; Van Rein, H.; Moore, P. Marine Strategy Framework Directive Indicators for UK Kelp Habitats Part 1: Developing Proposals for Potential Indicators; Joint Nature Conservation Committee: Peterborough, UK, 2014. [Google Scholar]
- Jones, L.A.; Hiscock, K.; Connor, D.W. Marine Habitat Reviews: A summary of Ecological Requirements and Sensitivity Characteristics for the Conservation and Management of Marine SACs; Joint Nature Conservation Committee: Peterborough, UK, 2000. [Google Scholar]
- Christie, H.; Jørgensen, N.M.; Norderhaug, K.M.; Waage-Nielsen, E. Species distribution and habitat exploitation of fauna associated with kelp (Laminaria hyperborea) along the Norwegian coast. Mar. Biol. Assoc. UK J. Mar. Biol. Assoc. 2003, 83, 687. [Google Scholar] [CrossRef]
- Adams, J.; Toop, T.; Donnison, I.S.; Gallagher, J.A. Seasonal variation in Laminaria digitata and its impact on biochemical conversion routes to biofuels. Bioresour. Technol. 2011, 102, 9976–9984. [Google Scholar] [CrossRef] [PubMed]
- Kain, J.M. A view of the genus Laminaria. Oceanogr. Mar. Biol. 1979, 17, 101–161. [Google Scholar]
- Pavia, H.; Carr, H.; Åberg, P. Habitat and feeding preferences of crustacean mesoherbivores inhabiting the brown seaweed Ascophyllum nodosum (L.) Le Jol. and its epiphytic macroalgae. J. Exp. Mar. Biol. Ecol. 1999, 236, 15–32. [Google Scholar] [CrossRef]
- Wilkinson, M. Information Review on the Impact of Kelp Harvesting; Scottish Natural Heritage: Edinburgh, UK, 1995. [Google Scholar]
- Marzinelli, E.M.; Leong, M.R.; Campbell, A.H.; Steinberg, P.D.; Vergés, A. Does restoration of a habitat-forming seaweed restore associated faunal diversity? Restor. Ecol. 2016, 24, 81–90. [Google Scholar] [CrossRef]
- Pereira, R.C.; Da Gama, B.A.P.; Teixeira, V.L.; Yoneshigue-Valentin, Y. Ecological roles of natural products of the Brazilian red seaweed Laurencia obtusa. Braz. J. Biol. 2003, 63, 665–672. [Google Scholar] [CrossRef]
- McClintock, J.B.; Baker, B.J. Marine Chemical Ecology; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Kumari, P.; Reddy, C. Antimicrobial compounds from seaweeds-associated bacteria and fungi. Appl. Microbiol. Biotechnol. 2015, 99, 1571–1586. [Google Scholar] [CrossRef]
- Pringgenies, D.; Retnowati, E.I.; Ariyanto, D.; Dewi, K.; Viharyo, M.A.; Susilowati, R. Symbiotic microbes from various seaweeds with antimicrobial and fermentative properties. Aquac. Aquar. Conserv. Legis. 2020, 13, 2211–2217. [Google Scholar]
- Hoey, G.v.; Drent, J.; Ysebaert, T.; Herman, P. The Benthic Ecosystem Quality Index (BEQI), Intercalibraton and Assessment of Dutch Coastal and Transitional Waters for the Water Frame Directive. Final Report. 2008. Available online: https://www.semanticscholar.org/paper/The-Benthic-Ecosystem-Quality-Index-(BEQI)%2C-and-of-Hoey-Drent/fda18bba6b2c10924aa62d96d9a2f4768b5422a6 (accessed on 20 April 2021).
- Alonso, P.D. Water European Law And The Watershed Management. Int. Bus. Econ. Res. J. 2012, 11, 1545–1548. [Google Scholar] [CrossRef] [Green Version]
- The Scottish, G. Wild Seaweed Harvesting: Strategic Environmental Assessment Environmental Report; APS Group Scotland: Edinburgh, UK, 2016. [Google Scholar]
- Juanes, J.; Guinda, X.; Puente, A.; Revilla, J. Macroalgae, a suitable indicator of the ecological status of coastal rocky communities in the NE Atlantic. Ecol. Indic. 2008, 8, 351–359. [Google Scholar] [CrossRef]
- Alami, A.H.; Alasad, S.; Ali, M.; Alshamsi, M. Investigating algae for CO2 capture and accumulation and simultaneous production of biomass for biodiesel production. Sci. Total Environ. 2021, 759, 143529. [Google Scholar] [CrossRef] [PubMed]
- Tsai, D.D.-W.; Chen, P.H.; Ramaraj, R. The potential of carbon dioxide capture and sequestration with algae. Ecol. Eng. 2017, 98, 17–23. [Google Scholar] [CrossRef]
- Muraoka, D. Seaweed resources as a source of carbon fixation. Bull. Fish. Res. Agency Jpn. 2004, (Supplement No. 1), 59–64. [Google Scholar]
- Zou, D.; Gao, K. Physiological responses of seaweeds to elevated atmospheric CO2 concentrations. In Seaweeds and their Role In Globally Changing Environments; Springer: Berlin/Heidelberg, Germany, 2010; pp. 115–126. [Google Scholar]
- Fernández, P.A.; Hurd, C.L.; Roleda, M.Y. Bicarbonate uptake via an anion exchange protein is the main mechanism of inorganic carbon acquisition by the giant kelp M acrocystis pyrifera (L aminariales, P haeophyceae) under variable pH. J. Phycol. 2014, 50, 998–1008. [Google Scholar] [CrossRef] [PubMed]
- Change.org. Do Not Allow Mechanical Kelp Dredging in Scottish Waters. Available online: https://www.change.org/p/scottish-parliament-ensure-that-mechanical-kelp-dredging-does-not-happen-in-scotland (accessed on 20 May 2021).
- Oaten, J.; Hull, S.; Roberts, C.; Brooks, T.; San Martin, E.; Smedley, M. Wild Seaweed Harvesting; ABP mer: Southampton, UK, 2018. [Google Scholar]
- Scotland, M. Seaweed Cultivation Policy Statement. 2017. Available online: https://www.gov.scot/binaries/content/documents/govscot/publications/speech-statement/2017/03/seaweed-cultivation-policy-statement-2017/documents/00515518-pdf/00515518-pdf/govscot%3Adocument/00515518.pdf (accessed on 20 April 2021).
- Chakdar, H.; Jadhav, S.D.; Dhar, D.W.; Pabbi, S. Potential applications of blue green algae. J. Sci. Indust. Res. 2012, 71, 13–20. [Google Scholar]
- MacArtain, P.; Gill, C.I.R.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible seaweeds. Nutr. Rev. 2007, 65, 535–543. [Google Scholar] [CrossRef]
- Fleurence, J.; Morançais, M.; Dumay, J. Seaweed proteins. In Proteins in Food Processing; Elsevier: Amsterdam, The Netherlands, 2018; pp. 245–262. [Google Scholar]
- Sho, H. History and characteristics of Okinawan longevity food. Asia Pac. J. Clin. Nutr. 2001, 10, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Allen, V.G.; Pond, K.R.; Saker, K.E.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Brown, C.P.; Miller, M.F.; Montgomery, J.L. Tasco-Forage: III. Influence of a seaweed extract on performance, monocyte immune cell response, and carcass characteristics in feedlot-finished steers. J. Anim. Sci. 2001, 79, 1032–1040. [Google Scholar] [CrossRef]
- Montgomery, J.L.; Allen, V.G.; Pond, K.R.; Miller, M.F.; Wester, D.B.; Brown, C.P.; Evans, R.; Bagley, C.P.; Ivy, R.L.; Fontenot, J.P. Tasco-Forage: IV. Influence of a seaweed extract applied to tall fescue pastures on sensory characteristics, shelf-life, and vitamin E status in feedlot-finished steers. J. Anim. Sci. 2001, 79, 884–894. [Google Scholar] [CrossRef]
- Saker, K.E.; Allen, V.G.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Wester, D.B. Tasco-Forage: II. Monocyte immune cell response and performance of beef steers grazing tall fescue treated with a seaweed extract. J. Anim. Sci. 2001, 79, 1022–1031. [Google Scholar] [CrossRef] [Green Version]
- Brownlee, I.A.; Seal, C.J.; Wilcox, M.; Dettmar, P.W.; Pearson, J.P. Applications of alginates in food. In Alginates: Biology and Applications; Springer: Berlin/Heidelberg, Germany, 2009; pp. 211–228. [Google Scholar]
- Brownlee, I.; Allen, A.; Pearson, J.; Dettmar, P.; Havler, M.; Atherton, M.; Onsøyen, E. Alginate as a source of dietary fiber. Crit. Rev. Food Sci. Nutr. 2005, 45, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.M.; Jorge, K.; Nogueira, L.C.; Silva, F.; Trugo, L.C. Effects of the combination of hydrophobic polypeptides, iso-α acids, and malto-oligosaccharides on beer foam stability. J. Agric. Food Chem. 2005, 53, 4976–4981. [Google Scholar] [CrossRef]
- Huang, X.; Kakuda, Y.; Cui, W. Hydrocolloids in emulsions: Particle size distribution and interfacial activity. Food Hydrocoll. 2001, 15, 533–542. [Google Scholar] [CrossRef]
- George, M.; Abraham, T.E. Polyionic hydrocolloids for the intestinal delivery of protein drugs: Alginate and chitosan—A review. J. Control Release 2006, 114, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Hwang, S.; Park, J.; Park, H.J. Preparation and release characteristics of polymer-coated and blended alginate microspheres. J. Microencapsul. 2003, 20, 179–192. [Google Scholar] [CrossRef] [PubMed]
- de Vos, P.; Faas, M.M.; Strand, B.; Calafiore, R. Alginate-based microcapsules for immunoisolation of pancreatic islets. Biomaterials 2006, 27, 5603–5617. [Google Scholar] [CrossRef]
- Uludag, H.; De Vos, P.; Tresco, P.A. Technology of mammalian cell encapsulation. Adv. Drug Deliv. Rev. 2000, 42, 29–64. [Google Scholar] [CrossRef]
- Wang, L.Z.; Liu, L.; Holmes, J.; Kerry, J.F.; Kerry, J.P. Assessment of film-forming potential and properties of protein and polysaccharide-based biopolymer films. Int. J. Food Sci. Technol. 2007, 42, 1128–1138. [Google Scholar] [CrossRef]
- Oussalah, M.; Caillet, S.; Salmieri, S.; Saucier, L.; Lacroix, M. Antimicrobial effects of alginate-based films containing essential oils on Listeria monocytogenes and Salmonella typhimurium present in bologna and ham. J. Food Prot. 2007, 70, 901–908. [Google Scholar] [CrossRef]
- Datta, S.; Janes, M.; Xue, Q.G.; Losso, J.; La Peyre, J. Control of Listeria monocytogenes and Salmonella anatum on the surface of smoked salmon coated with calcium alginate coating containing oyster lysozyme and nisin. J. Food Sci. 2008, 73, M67–M71. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Sathiyaseelan, A.; Mariadoss, A.V.A.; Xiaowen, H.; Wang, M.-H. Physical and bioactivities of biopolymeric films incorporated with cellulose, sodium alginate and copper oxide nanoparticles for food packaging application. Int. J. Biol. Macromol. 2020, 153, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Amjadi, S.; Nouri, S.; Yorghanlou, R.A.; Roufegarinejad, L. Development of hydroxypropyl methylcellulose/sodium alginate blend active film incorporated with Dracocephalum moldavica L. essential oil for food preservation. J. Thermoplast. Compos. Mater. 2020, 0892705720962153. [Google Scholar] [CrossRef]
- Fenoradosoa, T.A.; Ali, G.; Delattre, C.; Laroche, C.; Petit, E.; Wadouachi, A.; Michaud, P. Extraction and characterization of an alginate from the brown seaweed Sargassum turbinarioides Grunow. J. Appl. Phycol. 2010, 22, 131–137. [Google Scholar] [CrossRef]
- Gheorghita Puscaselu, R.; Lobiuc, A.; Dimian, M.; Covasa, M. Alginate: From Food Industry to Biomedical Applications and Management of Metabolic Disorders. Polymers 2020, 12, 2417. [Google Scholar] [CrossRef]
- Cherry, P. Seaweeds as a Source of Non-Digestible Complex Polysaccharide Components for the Development of Novel Prebiotic Ingredients for the Functional Food Industry; Ulster University: Coleraine, Ireland, 2020. [Google Scholar]
- Fao, F. Food and Agriculture Organisation of the United Nations. Available online: http://www.fao.org/home/en/ (accessed on 18 June 2021).
- Nisizawa, K.; Noda, H.; Kikuchi, R.; Watanabe, T. The main seaweed foods in Japan. Hydrobiologia 1987, 151, 5–29. [Google Scholar] [CrossRef]
- Fujiwara-Arasaki, T.; Mino, N.; Kuroda, M. The Protein Value in Human Nutrition of Edible Marine Algae in Japan; Springer: Berlin/Heidelberg, Germany, 1984; pp. 513–516. [Google Scholar]
- Hall, A.C.; Fairclough, A.C.; Mahadevan, K.; Paxman, J.R. Ascophyllum nodosum enriched bread reduces subsequent energy intake with no effect on post-prandial glucose and cholesterol in healthy, overweight males. A pilot study. Appetite 2012, 58, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Hoad, C.L.; Rayment, P.; Spiller, R.C.; Marciani, L.; Alonso, B.d.C.; Traynor, C.; Mela, D.J.; Peters, H.P.; Gowland, P.A. In vivo imaging of intragastric gelation and its effect on satiety in humans. J. Nutr. 2004, 134, 2293–2300. [Google Scholar] [CrossRef] [Green Version]
- Pelkman, C.L.; Navia, J.L.; Miller, A.E.; Pohle, R.J. Novel calcium-gelled, alginate-pectin beverage reduced energy intake in nondieting overweight and obese women: Interactions with dietary restraint status. Am. J. Clin. Nutr. 2007, 86, 1595–1602. [Google Scholar] [CrossRef]
- Holman, B.W.B.; Malau-Aduli, A.E.O. Spirulina as a livestock supplement and animal feed. J. Anim. Physiol. Anim. Nutr. 2013, 97, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Angell, A.R.; Angell, S.F.; de Nys, R.; Paul, N.A. Seaweed as a protein source for mono-gastric livestock. Trends Food Sci. Technol. 2016, 54, 74–84. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Milley, J.E.; Lall, S.P. Nutritional quality of some wild and cultivated seaweeds: Nutrient composition, total phenolic content and in vitro digestibility. J. Appl. Phycol. 2016, 28, 3575–3585. [Google Scholar] [CrossRef]
- de Beukelaar, M.F.A.; Zeinstra, G.G.; Mes, J.J.; Fischer, A.R.H. Duckweed as human food. The influence of meal context and information on duckweed acceptability of Dutch consumers. Food Qual. Prefer. 2019, 71, 76–86. [Google Scholar] [CrossRef]
- Van der Spiegel, M.; Noordam, M.Y.; Van der Fels-Klerx, H.J. Safety of novel protein sources (insects, microalgae, seaweed, duckweed, and rapeseed) and legislative aspects for their application in food and feed production. Compr. Rev. Food Sci. Food Saf. 2013, 12, 662–678. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food Security: The Challenge of Feeding 9 Billion People. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [Green Version]
- Sousa, I.; Gouveia, L.; Batista, A.P.; Raymundo, A.; Bandarra, N.M. Microalgae in novel food products. Food Chem. Res. Dev. 2008, 75–112. [Google Scholar]
- Van Krimpen, M.M.; Bikker, P.; Van der Meer, I.M.; Van der Peet-Schwering, C.M.C.; Vereijken, J.M. Cultivation, Processing and Nutritional Aspects for Pigs and Poultry of European Protein Sources as Alternatives for Imported Soybean Products; Wageningen UR Livestock Research: Wageningen, The Netherlands, 2013. [Google Scholar]
- Trentacoste, E.M.; Martinez, A.M.; Zenk, T. The place of algae in agriculture: Policies for algal biomass production. Photosynth. Res. 2015, 123, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Handler, R.M.; Shi, R.; Shonnard, D.R. Land use change implications for large-scale cultivation of algae feedstocks in the United States Gulf Coast. J. Clean. Prod. 2017, 153, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, D.; Houser, J.; Preiss, E.; White, O.; Fang, H.; Mesnick, L.; Barsky, T.; Tariche, S.; Schreck, J.; Alpert, S. Water resources: Agriculture, the environment, and society. BioScience 1997, 47, 97–106. [Google Scholar] [CrossRef]
- Iwamoto, H. Industrial production of microalgal cell-mass and secondary products-major industrial species. Handb. Microalgal Cult. Biotechnol. Appl. Phycol. 2004, 255, 263. [Google Scholar]
- Champenois, J.; Marfaing, H.; Pierre, R. Review of the taxonomic revision of Chlorella and consequences for its food uses in Europe. J. Appl. Phycol. 2015, 27, 1845–1851. [Google Scholar] [CrossRef]
- Buschmann, A.H.; Hernandez-Gonzalez, M.d.C.; Varela, D. Seaweed future cultivation in Chile: Perspectives and challenges. Int. J. Environ. Pollut. 2008, 33, 432–456. [Google Scholar] [CrossRef]
- Aaron-Amper, J.; Largo, D.B.; Handugan, E.R.B.; Nini, J.L.; Alingasa, K.M.A.; Gulayan, S.J. Culture of the tropical brown seaweed Sargassum aquifolium: From hatchery to field out-planting. Aquac. Rep. 2020, 16, 100265. [Google Scholar] [CrossRef]
- Joubert, Y.; Fleurence, J. Simultaneous extraction of proteins and DNA by an enzymatic treatment of the cell wall of Palmaria palmata (Rhodophyta). J. Appl. Phycol. 2008, 20, 55–61. [Google Scholar] [CrossRef]
- Abdel-Fattah, A.F.; Sary, H.H. Selective isolation of glycoprotein materials from the green seaweed Ulva lactuca. Pak. J. Biochem. 1987, 20, 61. [Google Scholar]
- Polprasert, C.; Koottatep, T. Organic Waste Recycling: Technology, Management and Sustainability; IWA Publishing: London, UK, 2017. [Google Scholar]
- Wosnitza, T.M.A.; Barrantes, J.G. Utilization of seaweed Ulva sp. in Paracas Bay (Peru): Experimenting with compost. J. Appl. Phycol. 2006, 18, 27. [Google Scholar] [CrossRef]
- Eyras, M.C.; Defosse, G.; Dellatorre, F. Seaweed compost as an amendment for horticultural soils in Patagonia, Argentina. Compos. Sci. Util. 2008, 16, 119–124. [Google Scholar] [CrossRef]
- Haug, R. The Practical Handbook of Compost Engineering; Routledge: London, UK, 2018. [Google Scholar]
- Chang, J.I.; Hsu, T.-E. Effects of compositions on food waste composting. Bioresour. Technol. 2008, 99, 8068–8074. [Google Scholar] [CrossRef]
- Eiland, F.; Klamer, M.; Lind, A.-M.; Leth, M.; Bååth, E. Influence of initial C/N ratio on chemical and microbial composition during long term composting of straw. Microb. Ecol. 2001, 41, 272–280. [Google Scholar] [CrossRef]
- Liu, C.-Y.; Paull, J.; Rathjen, A. Shoot mineral composition and yield of wheat genotypes grown on a sodic and a non-sodic soil. Aust. J. Exp. Agric. 2000, 40, 69–78. [Google Scholar] [CrossRef]
- EKOGEA. BCx for Compost. Available online: https://www.ekogea-int.com/compost (accessed on 20 May 2021).
- Hasznos, G. Compost/Biofertiliser Certification Schemes’ Annual Report 2018; Renewable Energy Assurance Ltd.: London, UK, 2019. [Google Scholar]
- Jayaraman, J.; Norrie, J.; Punja, Z.K. Commercial extract from the brown seaweed Ascophyllum nodosum reduces fungal diseases in greenhouse cucumber. J. Appl. Phycol. 2011, 23, 353–361. [Google Scholar] [CrossRef]
- Lola-Luz, T.; Hennequart, F.; Gaffney, M. Effect on yield, total phenolic, total flavonoid and total isothiocyanate content of two broccoli cultivars (Brassica oleraceae var italica) following the application of a commercial brown seaweed extract (Ascophyllum nodosum). Agric. Food Sci. 2014, 23, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Rayorath, P.; Jithesh, M.N.; Farid, A.; Khan, W.; Palanisamy, R.; Hankins, S.D.; Critchley, A.T.; Prithiviraj, B. Rapid bioassays to evaluate the plant growth promoting activity of Ascophyllum nodosum (L.) Le Jol. using a model plant, Arabidopsis thaliana (L.) Heynh. J. Appl. Phycol. 2008, 20, 423–429. [Google Scholar] [CrossRef]
- Somai-Jemmali, L.; Siah, A.; Randoux, B.; Magnin-Robert, M.; Halama, P.; Hamada, W.; Reignault, P. Brown alga Ascophyllum nodosum extract-based product, Dalgin Active®, triggers defense mechanisms and confers protection in both bread and durum wheat against Zymoseptoria tritici. J. Appl. Phycol. 2020, 32, 3387–3399. [Google Scholar] [CrossRef]
- De Saeger, J.; Van Praet, S.; Vereecke, D.; Park, J.; Jacques, S.; Han, T.; Depuydt, S. Toward the molecular understanding of the action mechanism of Ascophyllum nodosum extracts on plants. J. Appl. Phycol. 2019, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Ervin, E.H. Cytokinins and seaweed extracts for summer putting green health. Golfdom 2013, 69, 36–39. [Google Scholar]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Zodape, S.; Gupta, A.; Bhandari, S.; Rawat, U.; Chaudhary, D.; Eswaran, K.; Chikara, J. Foliar application of seaweed sap as biostimulant for enhancement of yieldand quality of tomato (Lycopersicon esculentum Mill.). J. Sci. Ind. Res. 2011, 70, 215–219. [Google Scholar]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Fan, D.; Hodges, D.M.; Zhang, J.; Kirby, C.W.; Ji, X.; Locke, S.J.; Critchley, A.T.; Prithiviraj, B. Commercial extract of the brown seaweed Ascophyllum nodosum enhances phenolic antioxidant content of spinach (Spinacia oleracea L.) which protects Caenorhabditis elegans against oxidative and thermal stress. Food Chem. 2011, 124, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Angus, S. Modern Seaweed Harvesting and Gathering in Scotland: The Legal and Ecological Context. Scott. Geogr. J. 2017, 133, 1–14. [Google Scholar] [CrossRef]
- VisitScotland. Scotland Map. Available online: https://www.visitscotland.com/destinations-maps/ (accessed on 18 May 2021).
- Angus, S. Dé tha cearr air a’mhachaire? Biodiversity issues for Scottish machair: An initial appraisal. Glasg. Nat. 2009, 25, 53–62. [Google Scholar]
- Marine, S. Draft Seaweed Policy Statement Consultation Paper; APS Group Scotland: Scotland, UK, 2013. [Google Scholar]
- Mac Monagail, M.; Cornish, L.; Morrison, L.; Araújo, R.; Critchley, A.T. Sustainable harvesting of wild seaweed resources. Eur. J. Phycol. 2017, 52, 371–390. [Google Scholar] [CrossRef]
- Radulovich, R.; Neori, A.; Valderrama, D.; Reddy, C.; Cronin, H.; Forster, J. Farming of seaweeds. In Seaweed Sustainability; Elsevier: Amsterdam, The Netherlands, 2015; pp. 27–59. [Google Scholar]
- Hurd, C.L.; Harrison, P.J.; Bischof, K.; Lobban, C.S. Seaweed Ecology and Physiology; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Valderrama, D.; Cai, J.; Hishamunda, N.; Ridler, N. Social and Economic Dimensions of Carrageenan Seaweed Farming; FAO: Rome, Italy, 2013. [Google Scholar]
- Burrows, M.T.; Macleod, M.; Orr, K. Mapping the Intertidal Seaweed Resources of the Outer Hebrides; Scottish Association for Marine Science and Hebridean Seaweed Company: Scotland, UK, 2010. [Google Scholar]
- MacLeod, A.K.; Orr, K.K.; Greenhill, L.; Burrows, M.T. Understanding the Potential Effects of Wave Energy Devices on Kelp Biotopes; Scottish Natural Heritage: Inverness, UK, 2014.
- Byrnes, J.E.; Reed, D.C.; Cardinale, B.J.; Cavanaugh, K.C.; Holbrook, S.J.; Schmitt, R.J. Climate-driven increases in storm frequency simplify kelp forest food webs. Glob. Chang. Biol. 2011, 17, 2513–2524. [Google Scholar] [CrossRef] [Green Version]
- Burrows, M.T.; Fox, C.J.; Moore, P.; Smale, D.; Sotheran, I.; Benson, A.; Greenhill, L.; Martino, S.; Parker, A.; Thompson, E.; et al. Wild Seaweed Harvesting as a Diversification Opportunity for Fishermen; A Report by SRSL for Highlands and Islands Enterprise: Scotland, UK, 2018. [Google Scholar]
- Kain, J.M.; Dawes, C. Useful European seaweeds: Past hopes and present cultivation. In Proceedings of the Twelfth International Seaweed Symposium; Springer: Dordrecht, The Netherlands, 1987; pp. 173–181. [Google Scholar]
- Andreakis, N.; Schaffelke, B. Invasive marine seaweeds: Pest or prize? In Seaweed Biology; Springer: Berlin/Heidelberg, Germany, 2012; pp. 235–262. [Google Scholar]
- Angulo-Valdés, J.A.; Hatcher, B.G. A new typology of benefits derived from marine protected areas. Mar. Policy 2010, 34, 635–644. [Google Scholar] [CrossRef]
- Tyler-Walters, H.; James, B.; Carruthers, M.; Wilding, C.; Durkin, O.; Lacey, C.; Philpott, E.; Adams, L.; Chaniotis, P.D.; Wilkes, P.T.V.; et al. Descriptions of Scottish Priority Marine Features (PMFs); Scottish Natural Heritage: Scotland, UK, 2016. [Google Scholar]
- Marine, S. Scotland’s National Marine Plan A Single Framework for Managing Our Seas; The Scottish Government: Edinburgh, UK, 2015. [Google Scholar]
- Wood, D.; Capuzzo, E.; Kirby, D.; Mooney-McAuley, K.; Kerrison, P. UK macroalgae aquaculture: What are the key environmental and licensing considerations? Mar. Policy 2017, 83, 29–39. [Google Scholar] [CrossRef]
- Kelly, C.; Gray, L.; Shucksmith, R.; Tweddle, J.F. Review and evaluation of marine spatial planning in the Shetland Islands. Mar. Policy 2014, 46, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, M.; Pico, Y. Chapter 2—Gas Chromatography and Mass Spectroscopy Techniques for the Detection of Chemical Contaminants and Residues in Foods. In Chemical Contaminants and Residues in Food, 2nd ed.; Schrenk, D., Cartus, A., Eds.; Woodhead Publishing: Cambridge, UK, 2017; pp. 15–50. [Google Scholar] [CrossRef]
- Chakraborty, K.; Joseph, D.; Praveen, N.K. Antioxidant activities and phenolic contents of three red seaweeds (Division: Rhodophyta) harvested from the Gulf of Mannar of Peninsular India. J. Food Sci. Technol. 2015, 52, 1924–1935. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Macquarrie, D. Microwave assisted extraction of sulfated polysaccharides (fucoidan) from Ascophyllum nodosum and its antioxidant activity. Carbohydr. Polym. 2015, 129, 101–107. [Google Scholar] [CrossRef]
- Gordaliza, M. Natural products as leads to anticancer drugs. Clin. Transl. Oncol. 2007, 9, 767–776. [Google Scholar] [CrossRef]
- Abel, U.; Koch, C.; Speitling, M.; Hansske, F.G. Modern methods to produce natural-product libraries. Curr. Opin. Chem. Biol. 2002, 6, 453–458. [Google Scholar] [CrossRef]
- Ranger, S.; Rose, C. Seaweed in the Daily Diet and Nutrition. Available online: http://www.seagreens.co.uk/Documents/Daily_Diet_and_Nutrtion_article_plus_products_10.13.pdf (accessed on 18 May 2021).
- Restani, P.; Persico, A.; Ballabio, C.; Moro, E.; Fuggetta, D.; Colombo, M.L. Analysis of food supplements containing iodine: A survey of Italian market. Clin. Toxicol. 2008, 46, 282–286. [Google Scholar] [CrossRef]
- Roy, M.-C.; Anguenot, R.; Fillion, C.; Beaulieu, M.; Bérubé, J.; Richard, D. Effect of a commercially-available algal phlorotannins extract on digestive enzymes and carbohydrate absorption in vivo. Food Res. Int. 2011, 44, 3026–3029. [Google Scholar] [CrossRef]
- Paradis, M.-E.; Couture, P.; Lamarche, B. A randomised crossover placebo-controlled trial investigating the effect of brown seaweed (Ascophyllum nodosum and Fucus vesiculosus) on postchallenge plasma glucose and insulin levels in men and women. Appl. Physiol. Nutr. Metab. 2011, 36, 913–919. [Google Scholar] [CrossRef]
- Díaz-Rubio, M.E.; Pérez-Jiménez, J.; Saura-Calixto, F. Dietary fiber and antioxidant capacity in Fucus vesiculosus products. Int. J. Food Sci. Nutr. 2009, 60, 23–34. [Google Scholar] [CrossRef]
- Ale, M.T.; Maruyama, H.; Tamauchi, H.; Mikkelsen, J.D.; Meyer, A.S. Fucoidan from Sargassum sp. and Fucus vesiculosus reduces cell viability of lung carcinoma and melanoma cells in vitro and activates natural killer cells in mice in vivo. Int. J. Biol. Macromol. 2011, 49, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Liang, H.; Ji, X.; Zhou, Z.; Liu, Y.; Sun, T.; Zhang, L. Effects of fucoidan on gut flora and tumor prevention in 1, 2-dimethylhydrazine-induced colorectal carcinogenesis. J. Nutr. Biochem. 2020, 82, 108396. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.-C.; Huang, R.-Y.; Chou, T.-C. Oligo-Fucoidan Improves Diabetes-Induced Renal Fibrosis via Activation of Sirt-1, GLP-1R, and Nrf2/HO-1: An In Vitro and In Vivo Study. Nutrients 2020, 12, 3068. [Google Scholar] [CrossRef] [PubMed]
- Zahra, N.; Hina, S.; Masood, S.; Kalim, I.; Saeed, M.K.; Ahmad, I.; Arshad, M. Exploration of Locally Grown Yellow and Green Pumpkin as a Potential Source of b-Carotene and Vitamin A. Biol. Sci. Pjsir 2020, 63, 238–241. [Google Scholar] [CrossRef]
- Coronel, J.; Pinos, I.; Amengual, J. β-carotene in obesity research: Technical considerations and current status of the field. Nutrients 2019, 11, 842. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Chen, X.; Nakamura, Y.; Yu, C.; Qi, H. Fucoxanthin activities motivate its nanoencapsulation for food or nutraceutical application: A review. Food Funct. 2020, 11, 9338–9358. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, M.; Takahashi, S.; Nishimura, R.; Kubota, A.; Kojima, H.; Ohta, T.; Hamada, J.; Kuramitsu, Y.; Maeda, H.; Miyashita, K. A Marine Carotenoid of Fucoxanthinol Accelerates the Growth of Human Pancreatic Cancer PANC-1 Cells. Nutr. Cancer 2021, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, H. Neoxanthin and fucoxanthinol in Fucus vesiculosus. Biochim. Biophys. Acta Gen. Subj. 1974, 338, 572–576. [Google Scholar] [CrossRef]
- Jin, W.; Yang, L.; Yi, Z.; Fang, H.; Chen, W.; Hong, Z.; Zhang, Y.; Zhang, G.; Li, L. Anti-Inflammatory Effects of Fucoxanthinol in LPS-Induced RAW264. 7 Cells through the NAAA-PEA Pathway. Mar. Drugs 2020, 18, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnett, R.; Mallams, A.; Spark, A.; Tee, J.; Weedon, B.; McCormick, A. Carotenoids and related compounds. Part XX. Structure and reactions of fucoxanthin. J. Chem. Soc. C Org. 1969, 3, 429–454. [Google Scholar] [CrossRef]
- Budzikiewicz, H.; Taraz, K. Chlorophyll c. Tetrahedron 1971, 27, 1447–1460. [Google Scholar] [CrossRef]
- Glombitza, K.-W.; Lentz, G. Antibiotics from algae—XXVIII: Cleavage of high molecular phlorotannin derivatives from the brown alga fucus vesiculosus L. Tetrahedron 1981, 37, 3861–3866. [Google Scholar] [CrossRef]
- Abdel-Lateff, A.; Fisch, K.M.; Wright, A.D.; König, G.M. A new antioxidant isobenzofuranone derivative from the algicolous marine fungus Epicoccum sp. Planta Med. 2003, 69, 831–834. [Google Scholar]
- McInnes, A.; Walter, J.; Wright, J. 13C NMR Spectra of Δ 24 (28) phytosterols. Org. Magn. Reson. 1980, 13, 302–303. [Google Scholar] [CrossRef]
- Ioannou, E.; Zervou, M.; Ismail, A.; Ktari, L.; Vagias, C.; Roussis, V. 2, 6-Cyclo-xenicanes from the brown algae Dilophus fasciola and Dilophus spiralis. Tetrahedron 2009, 65, 10565–10572. [Google Scholar] [CrossRef]
- Song, L.; Qu, D.; Zhang, Q.; Zhou, H.; Jiang, R.; Li, Y.; Zhang, Y.; Yan, H. Phytosterol esters attenuate hepatic steatosis in rats with non-alcoholic fatty liver disease rats fed a high-fat diet. Sci. Rep. 2017, 7, 1–18. [Google Scholar] [CrossRef]
- Heilbron, I.; Phipers, R.; Wright, H. 343. The chemistry of the algæ. Part I. The algal sterol fucosterol. J. Chem. Soc. 1934, 1572–1576. [Google Scholar] [CrossRef]
- Knapp, F.; Greig, J.; Goad, L.; Goodwin, T. The conversion of 24-ethylidene-sterols into poriferasterol by Ochromonas malhamensis. J. Chem. Soc. D Chem. Commun. 1971, 707–709. [Google Scholar] [CrossRef]
- Buedenbender, L.; Astone, F.A.; Tasdemir, D. Bioactive Molecular Networking for Mapping the Antimicrobial Constituents of the Baltic Brown Alga Fucus vesiculosus. Mar. Drugs 2020, 18, 311. [Google Scholar] [CrossRef] [PubMed]
- de Souza, N.J.; Nes, W.R. The presence of phytol in brown and blue-green algae and its relationship to evolution. Phytochemistry 1969, 8, 819–822. [Google Scholar] [CrossRef]
- Parys, S.; Kehraus, S.; Krick, A.; Glombitza, K.-W.; Carmeli, S.; Klimo, K.; Gerhäuser, C.; König, G.M. In vitro chemopreventive potential of fucophlorethols from the brown alga Fucus vesiculosus L. by anti-oxidant activity and inhibition of selected cytochrome P450 enzymes. Phytochemistry 2010, 71, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Gite, S.; Ross, R.P.; Kirke, D.; Guihéneuf, F.; Aussant, J.; Stengel, D.B.; Dinan, T.G.; Cryan, J.F.; Stanton, C. Nutraceuticals to promote neuronal plasticity in response to corticosterone-induced stress in human neuroblastoma cells. Nutr. Neurosci. 2019, 22, 551–568. [Google Scholar] [CrossRef]
- Rupérez, P.; Ahrazem, O.; Leal, J.A. Potential antioxidant capacity of sulfated polysaccharides from the edible marine brown seaweed Fucus vesiculosus. J. Agric. Food Chem. 2002, 50, 840–845. [Google Scholar] [CrossRef]
- Budzikiewicz, H.; Brzezinka, H.; Johannes, B. Zur photosynthese grüner pflanzen, 2. Mitt.: Massenspektroskopische untersuchungen an carotinoiden. Mon. Für Chem. Chem. Mon. 1970, 101, 579–609. [Google Scholar] [CrossRef]
- Haugan, J.A.; Liaaen-Jensen, S. Isolation and characterisation of four allenic (6′ S)-isomers of fucoxanthin. Tetrahedron Lett. 1994, 35, 2245–2248. [Google Scholar] [CrossRef]
- Kim, M.-M.; Kim, S.-K. Effect of phloroglucinol on oxidative stress and inflammation. Food Chem. Toxicol. 2010, 48, 2925–2933. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.-H.; Qian, Z.-J.; Ryu, B.; Karadeniz, F.; Kim, D.; Kim, S.-K. Antioxidant peptides from protein hydrolysate of microalgae Navicula incerta and their protective effects in HepG2/CYP2E1 cells induced by ethanol. Phytother. Res. 2012, 26, 1555–1563. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.-S. Antithrombotic and profibrinolytic activities of phloroglucinol. Food Chem. Toxicol. 2011, 49, 1572–1577. [Google Scholar] [CrossRef] [PubMed]
- Zingue, S.; Michel, T.; Nde, C.B.M.; Njuh, A.N.; Cisilotto, J.; Ndinteh, D.T.; Clyne, C.; Fernandez, X.; Creczynski-Pasa, T.B.; Njamen, D. Estrogen-like and tissue-selective effects of 7-methoxycoumarin from Ficus umbellata (Moraceae): An in vitro and in vivo study. BMC Complementary Altern. Med. 2017, 17, 383. [Google Scholar] [CrossRef] [Green Version]
- Sancheti, S.; Sancheti, S.; Seo, S.-Y. Ameliorative effects of 7-methylcoumarin and 7-methoxycoumarin against CCl4-induced hepatotoxicity in rats. Drug Chem. Toxicol. 2013, 36, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Lopes, G.; Daletos, G.; Proksch, P.; Andrade, P.B.; Valentão, P. Anti-inflammatory potential of monogalactosyl diacylglycerols and a monoacylglycerol from the edible brown seaweed Fucus spiralis Linnaeus. Mar. Drugs 2014, 12, 1406–1418. [Google Scholar] [CrossRef] [PubMed]
- Fathalipour, M.; Fathalipour, H.; Safa, O.; Nowrouzi-Sohrabi, P.; Mirkhani, H.; Hassanipour, S. The therapeutic role of carotenoids in diabetic retinopathy: A systematic review. DiabetesMetab. Syndr. Obes. Targets Ther. 2020, 13, 2347. [Google Scholar] [CrossRef]
- Rai, M.; Rathod, D.; Ingle, A.; Proksch, P.; Kon, K. 5 Biocidal Metabolites from Endophytes that Occur in Medicinal Plants. In Natural Antioxidants and Biocides from Wild Medicinal Plants; CAB International: Wallingford, UK, 2013. [Google Scholar]
- Santiago, C.; Fitchett, C.; Munro, M.H.; Jalil, J.; Santhanam, J. Cytotoxic and antifungal activities of 5-hydroxyramulosin, a compound produced by an endophytic fungus isolated from Cinnamomum mollisimum. Evid. Based Complementary Altern. Med. 2012, 2012, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Sarfati, L. Stopping the Clock: Antiaging treatments and care for clients of every age. Ski. Deep 2017, 16, 48–53. [Google Scholar]
- He, L.; Shang, Z.; Liu, H.; Yuan, Z.-X. Alginate-Based Platforms for Cancer-Targeted Drug Delivery. Biomed Res. Int. 2020, 1–17. [Google Scholar] [CrossRef]
- Jiang, Z.; Okimura, T.; Yamaguchi, K.; Oda, T. The potent activity of sulfated polysaccharide, ascophyllan, isolated from Ascophyllum nodosum to induce nitric oxide and cytokine production from mouse macrophage RAW264.7 cells: Comparison between ascophyllan and fucoidan. Nitric Oxide 2011, 25, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Jensen, A.; Ragan, M.A. 1, 2, 3, 5-tetrahydroxybenzene 2, 5-disulfate ester: The “phenolic precursor” in gelbstoff-forming exudates from the marine brown alga Ascophyllum nodosum (L.) Lejol. Tetrahedron Lett. 1978, 19, 847–850. [Google Scholar] [CrossRef]
- Grosse-Damhues, J.; Glombitza, K.-W.; Schulten, H.-R. An eight-ring phlorotannin from the brown alga Himanthalia elongata. Phytochemistry 1983, 22, 2043–2046. [Google Scholar] [CrossRef]
- Liu, J.; Luthuli, S.; Wu, Q.; Wu, M.; Choi, J.-I.; Tong, H. Pharmaceutical and Nutraceutical Potential Applications of Sargassum fulvellum. Biomed Res. Int. 2020, 1–12. [Google Scholar] [CrossRef]
- Borst, E.M.; Ständker, L.; Wagner, K.; Schulz, T.F.; Forssmann, W.-G.; Messerle, M. A peptide inhibitor of cytomegalovirus infection from human hemofiltrate. Antimicrob. Agents Chemother. 2013, 57, 4751–4760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilan, M.I.; Grachev, A.A.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure of a fucoidan from the brown seaweed Fucus serratus L. Carbohydr. Res. 2006, 341, 238–245. [Google Scholar] [CrossRef]
- Kim, K.-T.; Rioux, L.-E.; Turgeon, S.L. Alpha-amylase and alpha-glucosidase inhibition is differentially modulated by fucoidan obtained from Fucus vesiculosus and Ascophyllum nodosum. Phytochemistry 2014, 98, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Britton, G. Carotenoids. In Natural Food Colorants; Springer: Berlin/Heidelberg, Germany, 1996; pp. 197–243. [Google Scholar]
- Takaichi, S. Carotenoids in algae: Distributions, biosyntheses and functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef] [PubMed]
- Heilbron, I.M.; Phipers, R.F. The algae: The lipochromes of Fucus vesiculosus. Biochem. J. 1935, 29, 1369–1375. [Google Scholar] [CrossRef] [Green Version]
- Elvira-Torales, L.I.; García-Alonso, J.; Periago-Castón, M.J. Nutritional importance of carotenoids and their effect on liver health: A review. Antioxidants 2019, 8, 229. [Google Scholar] [CrossRef] [Green Version]
- Mounien, L.; Tourniaire, F.; Landrier, J.-F. Anti-obesity effect of carotenoids: Direct impact on adipose tissue and adipose tissue-driven indirect effects. Nutrients 2019, 11, 1562. [Google Scholar] [CrossRef] [Green Version]
- Hussain, H.; Krohn, K.; Schulz, B.; Draeger, S.; Nazir, M.; Saleem, M. Two new antimicrobial metabolites from the endophytic fungus, Seimatosporium sp. Nat. Prod. Commun. 2012, 7, 1934578X1200700305. [Google Scholar] [CrossRef] [Green Version]
- Lei, G.; Song, C.; Luo, Y. Chemical composition of hydrosol volatiles of flowers from ten Paeonia× suffruticosa Andr. cultivars from Luoyang, China. Nat. Prod. Res. 2020, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Naveen, J.; Baskaran, V. Antidiabetic plant-derived nutraceuticals: A critical review. Eur. J. Nutr. 2018, 57, 1275–1299. [Google Scholar] [CrossRef]
- Lordan, S.; Smyth, T.J.; Soler-Vila, A.; Stanton, C.; Ross, R.P. The α-amylase and α-glucosidase inhibitory effects of Irish seaweed extracts. Food Chem. 2013, 141, 2170–2176. [Google Scholar] [CrossRef]
- Ulbricht, C. Focus: Diabetes. J. Diet. Suppl. 2011, 8, 239–256. [Google Scholar] [CrossRef]
- Haugan, J.A.; Aakermann, T.; Liaaen-Jensen, S. [20] Isolation of fucoxanthin and peridinin. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1992; Volume 213, pp. 231–245. [Google Scholar]
- Kang, K.A.; Lee, K.H.; Chae, S.; Zhang, R.; Jung, M.S.; Ham, Y.M.; Baik, J.S.; Lee, N.H.; Hyun, J.W. Cytoprotective effect of phloroglucinol on oxidative stress induced cell damage via catalase activation. J. Cell. Biochem. 2006, 97, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Tierney, M.S.; Soler-Vila, A.; Rai, D.K.; Croft, A.K.; Brunton, N.P.; Smyth, T.J. UPLC-MS profiling of low molecular weight phlorotannin polymers in Ascophyllum nodosum, Pelvetia canaliculata and Fucus spiralis. Metabolomics 2014, 10, 524–535. [Google Scholar] [CrossRef]
- Osterhage, C.; König, G.M.; Jones, P.G.; Wright, A.D. 5-Hydroxyramulosin, a new natural product produced by Phoma tropica, a marine-derived fungus isolated from the alga Fucus spiralis. Planta Med. 2002, 68, 1052–1054. [Google Scholar] [CrossRef]
- Williams, R.; Goodwin, T. The occurrence of mutatochrome in green tissues. Phytochemistry 1967, 6, 1037–1039. [Google Scholar] [CrossRef]
- Apostolidis, E.; Lee, C. In vitro potential of Ascophyllum nodosum phenolic antioxidant-mediated α-glucosidase and α-amylase inhibition. J. Food Sci. 2010, 75, H97–H102. [Google Scholar] [CrossRef]
- Zhang, J.; Tiller, C.; Shen, J.; Wang, C.; Girouard, G.S.; Dennis, D.; Barrow, C.J.; Miao, M.; Ewart, H.S. Antidiabetic properties of polysaccharide- and polyphenolic-enriched fractions from the brown seaweed Ascophyllum nodosum. Can. J. Physiol. Pharmacol. 2007, 85, 1116–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarfati, L. AGE Beautifully. Ski. Deep 2018, 17, 70–77. [Google Scholar]
- Dutot, M.; Fagon, R.; Hemon, M.; Rat, P. Antioxidant, anti-inflammatory, and anti-senescence activities of a phlorotannin-rich natural extract from brown seaweed Ascophyllum nodosum. Appl. Biochem. Biotechnol. 2012, 167, 2234–2240. [Google Scholar] [CrossRef]
- Nakayasu, S.; Soegima, R.; Yamaguchi, K.; Oda, T. Biological activities of fucose-containing polysaccharide ascophyllan isolated from the brown alga Ascophyllum nodosum. Biosci. Biotechnol. Biochem. 2009, 73, 961–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakubo, A.; Makino, H.; Ohnishi, J.-i.; Hirohara, H.; Hori, K. The marine red alga Eucheuma serra J. Agardh, a high yielding source of two isolectins. J. Appl. Phycol. 1997, 9, 331. [Google Scholar] [CrossRef]
- Weis, W.I.; Drickamer, K. Structural basis of lectin-carbohydrate recognition. Annu. Rev. Biochem. 1996, 65, 441–473. [Google Scholar] [CrossRef]
- Ziółkowska, N.E.; Wlodawer, A. Structural studies of algal lectins with anti-HIV activity. Acta Biochim. Pol. 2006, 53, 617–626. [Google Scholar] [CrossRef] [Green Version]
- Naeem, A.; Saleemuddin, M.; Hasan Khan, R. Glycoprotein targeting and other applications of lectins in biotechnology. Curr. Protein Pept. Sci. 2007, 8, 261–271. [Google Scholar] [CrossRef]
- Alavi, M.; Asare-Addo, K.; Nokhodchi, A. Lectin Protein as a Promising Component to Functionalize Micelles, Liposomes and Lipid NPs against Coronavirus. Biomedicines 2020, 8, 580. [Google Scholar] [CrossRef] [PubMed]
- Stratil, S.B.; Neulinger, S.C.; Knecht, H.; Friedrichs, A.K.; Wahl, M. Salinity affects compositional traits of epibacterial communities on the brown macroalga Fucus vesiculosus. Fems Microbiol. Ecol. 2014, 88, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Ihua, M.W.; Guihéneuf, F.; Mohammed, H.; Margassery, L.M.; Jackson, S.A.; Stengel, D.B.; Clarke, D.J.; Dobson, A.D. Microbial population changes in decaying Ascophyllum nodosum result in macroalgal-polysaccharide-degrading bacteria with potential applicability in enzyme-assisted extraction technologies. Mar. Drugs 2019, 17, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agüero, L.; Zaldivar-Silva, D.; Peña, L.; Dias, M.L. Alginate microparticles as oral colon drug delivery device: A review. Carbohydr. Polym. 2017, 168, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Harnedy, P.A.; FitzGerald, R.J. Extraction of protein from the macroalga Palmaria palmata. Lwt Food Sci. Technol. 2013, 51, 375–382. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Chemat, F.; Zille, H.; Khan, M.K. Applications of ultrasound in food technology: Processing, preservation and extraction. Ultrason. Sonochem. 2011, 18, 813–835. [Google Scholar] [CrossRef]
- Kuhavichanan, A.; Kusolkumbot, P.; Sirisattha, S.; Areeprasert, C. Mechanical Extraction of Protein Solution from Microalgae by Ultrasonication. IOP Conf. Ser. Earth Environ. Sci. 2018, 159, 12009. [Google Scholar] [CrossRef]
- Mæhre, H.; Jensen, I.-J.; Eilertsen, K.-E. Enzymatic pre-treatment increases the protein bioaccessibility and extractability in Dulse (Palmaria palmata). Mar. Drugs 2016, 14, 196. [Google Scholar] [CrossRef] [Green Version]
- Kadam, S.U.; Álvarez, C.; Tiwari, B.K.; O’Donnell, C.P. Extraction and characterization of protein from Irish brown seaweed Ascophyllum nodosum. Food Res. Int. 2017, 99, 1021–1027. [Google Scholar] [CrossRef]
- Barbarino, E.; Lourenço, S.O. An evaluation of methods for extraction and quantification of protein from marine macro-and microalgae. J. Appl. Phycol. 2005, 17, 447–460. [Google Scholar] [CrossRef]
- Vanthoor-Koopmans, M.; Wijffels, R.H.; Barbosa, M.J.; Eppink, M.H.M. Biorefinery of microalgae for food and fuel. Bioresour. Technol. 2013, 135, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Goettel, M.; Eing, C.; Gusbeth, C.; Straessner, R.; Frey, W. Pulsed electric field assisted extraction of intracellular valuables from microalgae. Algal Res. 2013, 2, 401–408. [Google Scholar] [CrossRef]
- Töpfl, S. Pulsed Electric Fields (PEF) for Permeabilization of Cell Membranes in Food-and Bioprocessing–Applications, Process and Equipment Design and Cost Analysis. Ph.D. Thesis, Technische Universität Berlin, Fakultät III, Berlin, Germnay, 2006. [Google Scholar]
- Barba, F.J.; Grimi, N.; Vorobiev, E. New approaches for the use of non-conventional cell disruption technologies to extract potential food additives and nutraceuticals from microalgae. Food Eng. Rev. 2015, 7, 45–62. [Google Scholar] [CrossRef]
- Kumar, P.; Sharma, N.; Ranjan, R.; Kumar, S.; Bhat, Z.F.; Jeong, D.K. Perspective of membrane technology in dairy industry: A review. Asian Australas. J. Anim. Sci. 2013, 26, 1347. [Google Scholar] [CrossRef] [Green Version]
- Macquarrie, D.J.; Clark, J.H.; Fitzpatrick, E. The microwave pyrolysis of biomass. BiofuelsBioprod. Biorefining 2012, 6, 549–560. [Google Scholar] [CrossRef]
- Michalak, I.; Tuhy, Ł.; Chojnacka, K. Seaweed extract by microwave assisted extraction as plant growth biostimulant. Open Chem. 2015, 13, 1183–1195. [Google Scholar] [CrossRef]
- van Hees, D.H.; Olsen, Y.S.; Wernberg, T.; Van Alstyne, K.L.; Kendrick, G.A. Phenolic concentrations of brown seaweeds and relationships to nearshore environmental gradients in Western Australia. Mar. Biol. 2017, 164, 74. [Google Scholar] [CrossRef] [Green Version]
- Tierney, M.S.; Smyth, T.J.; Hayes, M.; Soler-Vila, A.; Croft, A.K.; Brunton, N. Influence of pressurised liquid extraction and solid–liquid extraction methods on the phenolic content and antioxidant activities of I rish macroalgae. Int. J. Food Sci. Technol. 2013, 48, 860–869. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, J.; Fan, J.; Clark, J.; Shen, P.; Li, Y.; Zhang, C. Microwave assisted extraction of phenolic compounds from four economic brown macroalgae species and evaluation of their antioxidant activities and inhibitory effects on alpha-amylase, alpha-glucosidase, pancreatic lipase and tyrosinase. Food Res. Int. 2018, 113, 288–297. [Google Scholar] [CrossRef]
- Flórez-Fernández, N.; Domínguez, H.; Torres, M.D. A green approach for alginate extraction from Sargassum muticum brown seaweed using ultrasound-assisted technique. Int. J. Biol. Macromol. 2019, 124, 451–459. [Google Scholar] [CrossRef]
- Fertah, M. Chapter 2—Isolation and Characterization of Alginate from Seaweed. In Seaweed Polysaccharides; Venkatesan, J., Anil, S., Kim, S.-K., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 11–26. [Google Scholar] [CrossRef]
- Fertah, M.; Belfkira, A.; Dahmane, E.m.; Taourirte, M.; Brouillette, F. Extraction and characterization of sodium alginate from Moroccan Laminaria digitata brown seaweed. Arab. J. Chem. 2017, 10, S3707–S3714. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, A.; Bissoon, R.; Bajnath, E.; Mohammed, K.; Lee, T.; Bissram, M.; John, N.; Jalsa, N.K.; Lee, K.-Y.; Ward, K. Multistage extraction and purification of waste Sargassum natans to produce sodium alginate: An optimization approach. Carbohydr. Polym. 2018, 198, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Hernández-carmona, G.; McHugh, D.J.; Arvizu-Higuera, D.L.; Rodríguez-montesinos, Y.E. Pilot plant scale extraction of alginate from Macrocystis pyrifera. 1. Effect of pre-extraction treatments on yield and quality of alginate. J. Appl. Phycol. 1998, 10, 507–513. [Google Scholar] [CrossRef]
- McHugh, D.J. Production and Utilization of Products from Commercial Seaweeds; FAO: Rome, Italy, 1987; pp. 1553–1561. [Google Scholar]
- Chee, S.Y.; Wong, P.K.; Wong, C.L. Extraction and characterisation of alginate from brown seaweeds (Fucales, Phaeophyceae) collected from Port Dickson, Peninsular Malaysia. J. Appl. Phycol. 2011, 23, 191–196. [Google Scholar] [CrossRef]
- Rioux, L.E.; Turgeon, S.L.; Beaulieu, M. Characterization of polysaccharides extracted from brown seaweeds. Carbohydr. Polym. 2007, 69, 530–537. [Google Scholar] [CrossRef]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyraud, A.; Gey, C.; Leonard, C.; Rochas, C.; Girond, S.; Kloareg, B. NMR spectroscopy analysis of oligoguluronates and oligomannuronates prepared by acid or enzymatic hydrolysis of homopolymeric blocks of alginic acid. Application to the determination of the substrate specificity of Haliotis tuberculata alginate lyase. Carbohydr. Res. 1996, 289, 11–23. [Google Scholar] [CrossRef]
- Torres, M.R.; Sousa, A.P.A.; Silva Filho, E.A.T.; Melo, D.F.; Feitosa, J.P.A.; de Paula, R.C.M.; Lima, M.G.S. Extraction and physicochemical characterization of Sargassum vulgare alginate from Brazil. Carbohydr. Res. 2007, 342, 2067–2074. [Google Scholar] [CrossRef] [Green Version]
- Draget, K.I.; Strand, B.; Hartmann, M.; Valla, S.; Smidsrød, O.; Skjåk-Bræk, G. Ionic and acid gel formation of epimerised alginates; the effect of AlgE4. Int. J. Biol. Macromol. 2000, 27, 117–122. [Google Scholar] [CrossRef]
Figure 1.
The taxonomic order of the brown seaweeds, “the wracks”, which are widely distributed in Scotland.
Figure 1.
The taxonomic order of the brown seaweeds, “the wracks”, which are widely distributed in Scotland.

Figure 2.
The distribution of certified compost makers throughout the UK; data extracted from Hasznos (2019) shows that the highest number of compost makers are in England as 137 units, Scotland, 23, Wales, 12, and Northern Ireland, 7 [127].
Figure 2.
The distribution of certified compost makers throughout the UK; data extracted from Hasznos (2019) shows that the highest number of compost makers are in England as 137 units, Scotland, 23, Wales, 12, and Northern Ireland, 7 [127].

Figure 3.
The type of waste materials processed at certified waste centres in Scotland; data extracted from Hasznos (2019) shows that the waste centres in Scotland are processing green waste [127].
Figure 3.
The type of waste materials processed at certified waste centres in Scotland; data extracted from Hasznos (2019) shows that the waste centres in Scotland are processing green waste [127].

Figure 4.
The weight of waste in tonnes processed by certified plants in the countries of the UK; data extracted from Hasznos (2019) shows that England proceed the highest amount of waste per year [127].
Figure 4.
The weight of waste in tonnes processed by certified plants in the countries of the UK; data extracted from Hasznos (2019) shows that England proceed the highest amount of waste per year [127].

Figure 5.
The industrial seaweed harvesting areas in Scotland; the map has been produced by Omar Al-Dulaimi using a map of Scotland downloaded from https://www.visitscotland.com/destinations-maps/ accessed on 20 April 2021.
Figure 5.
The industrial seaweed harvesting areas in Scotland; the map has been produced by Omar Al-Dulaimi using a map of Scotland downloaded from https://www.visitscotland.com/destinations-maps/ accessed on 20 April 2021.

Figure 6.
Compounds from 1–7 (The fucoidan and several carotenoids).

Figure 7.
Compounds from 8–24 (Polyphenolic, isochromane, 2-furoic acid derivatives).

Figure 8.
Compounds from 25–28 (phytosterols, glycolipid, and farnesol) and compounds 29 and 30 (Diacylglyceroglycolipids).
Figure 8.
Compounds from 25–28 (phytosterols, glycolipid, and farnesol) and compounds 29 and 30 (Diacylglyceroglycolipids).

Figure 9.
Compounds from 31–33 (Polyphenolic compounds).

Figure 10.
Compounds from 34–42 (Carotenoids, flavonoids, sugars, and carotene).

Figure 11.
Compounds from 43–47 (phloroglucinol, coumarin, ester, carotene, and hydroxyramulosin).

Figure 12.
Compounds from 48–52 (Acids, sugars, and esters).

Figure 13.
Compounds from 53–59 (polysaccharides and sugars).

Figure 14.
A molecules formed of several G and M.


{kind=link}
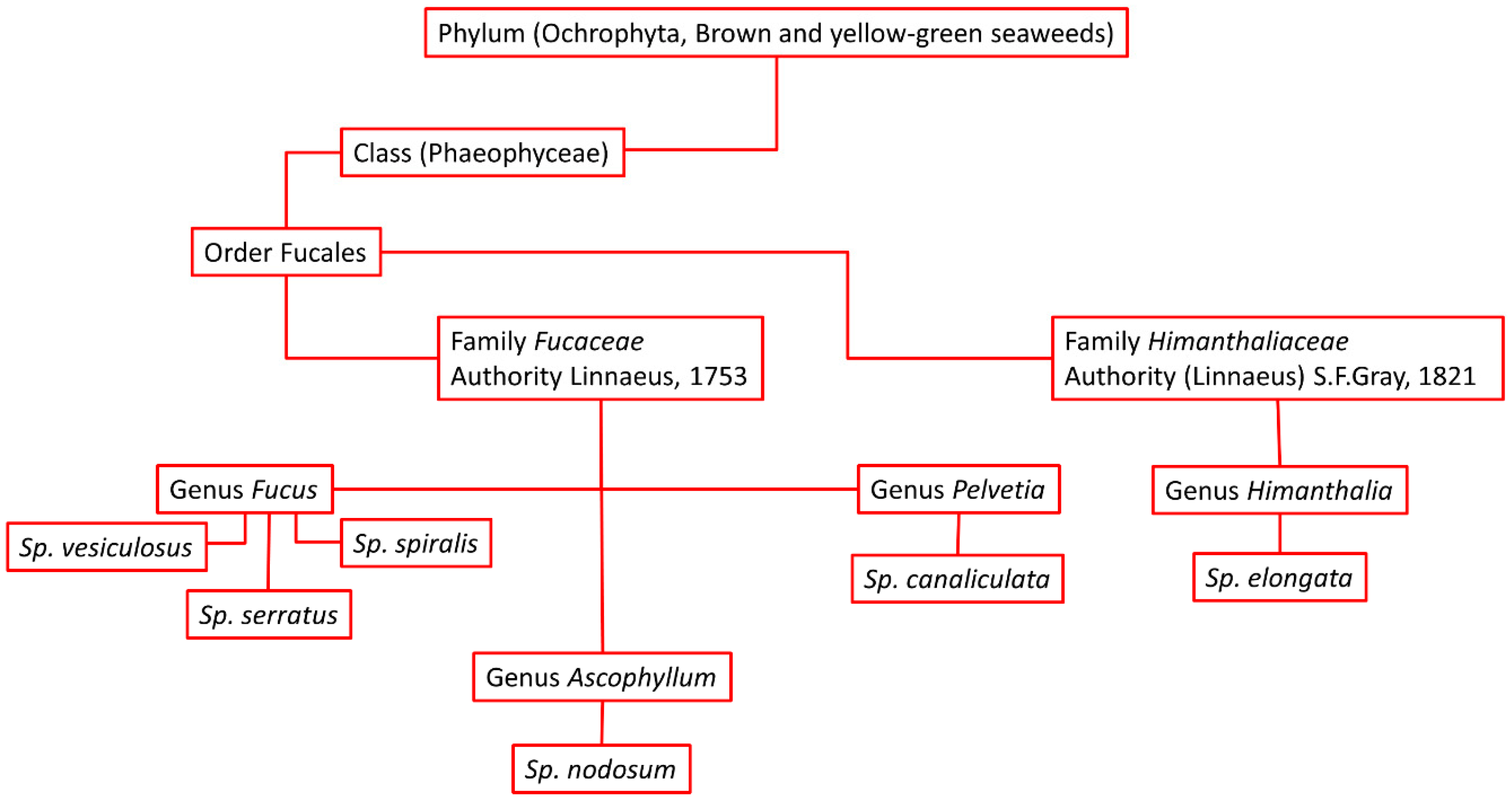{kind=link}
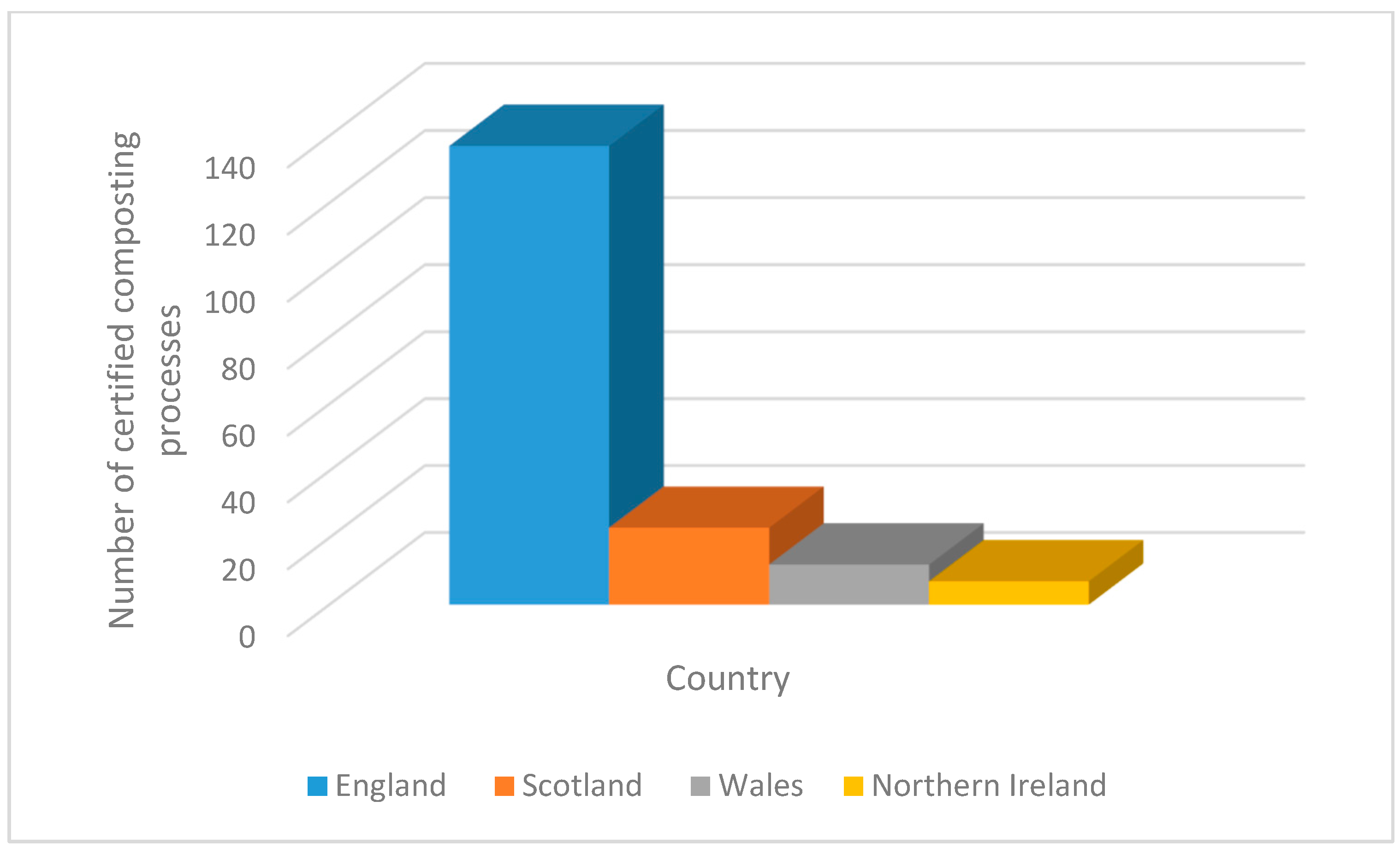{kind=link}
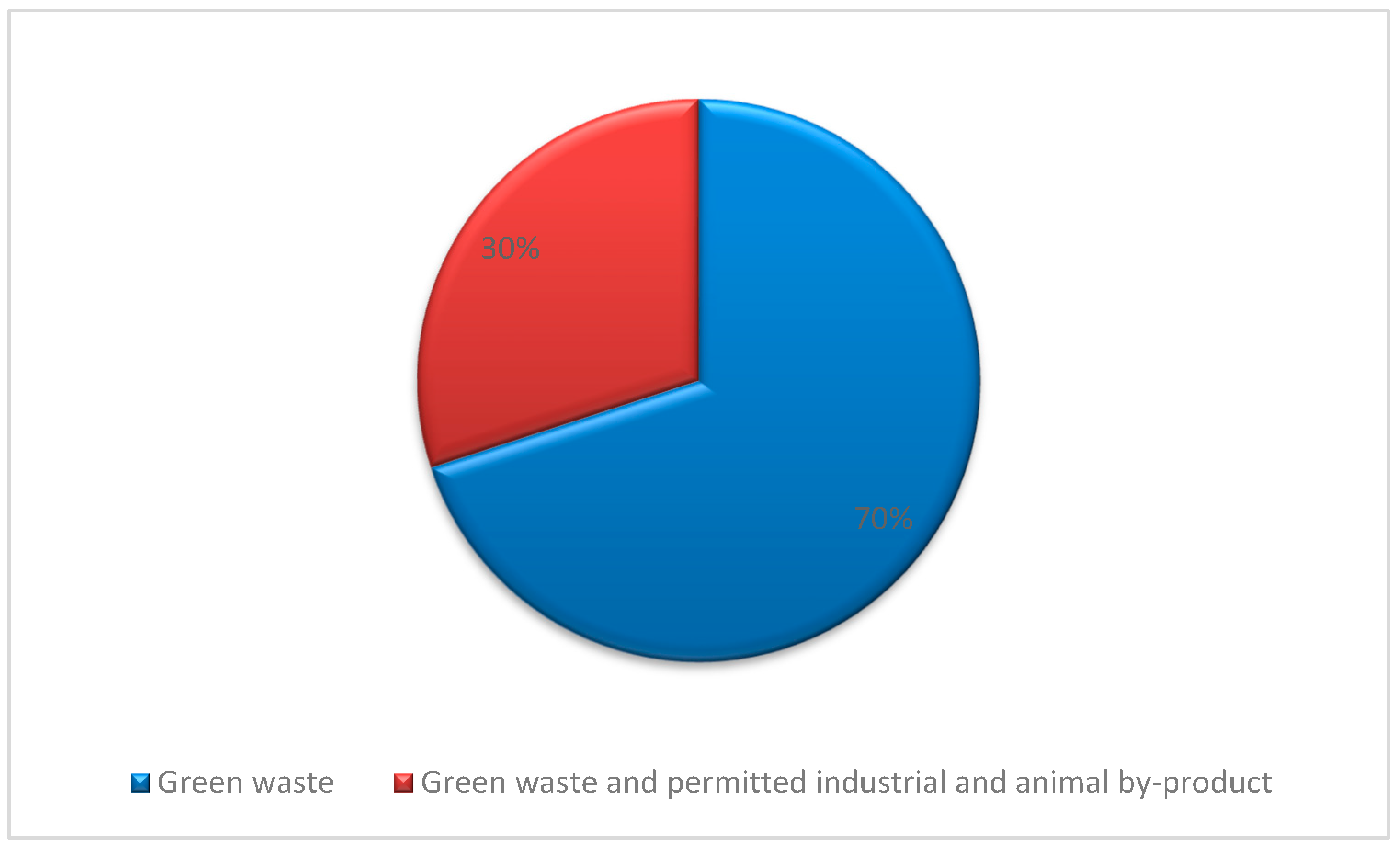{kind=link}
{kind=link}
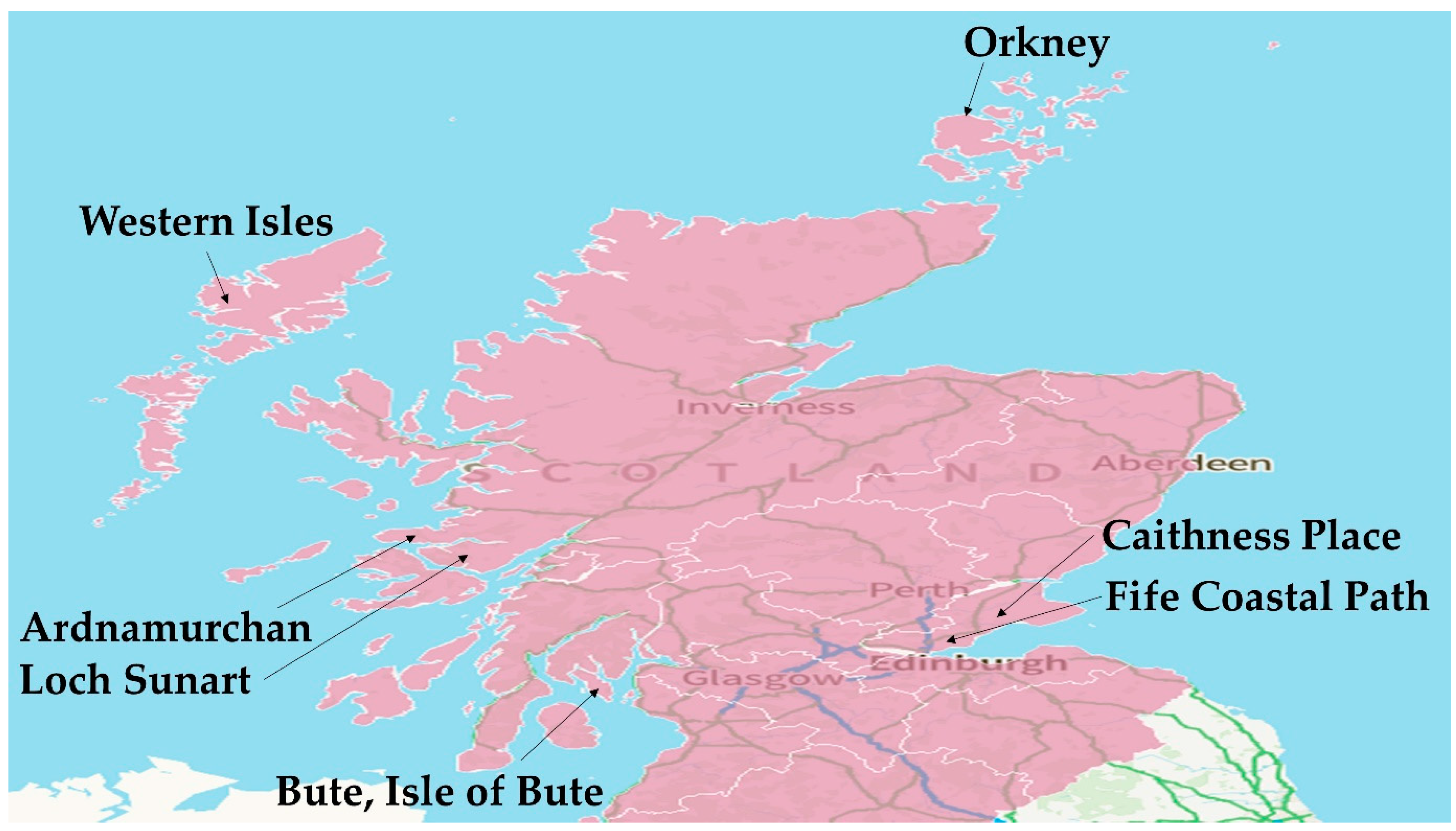{kind=link}
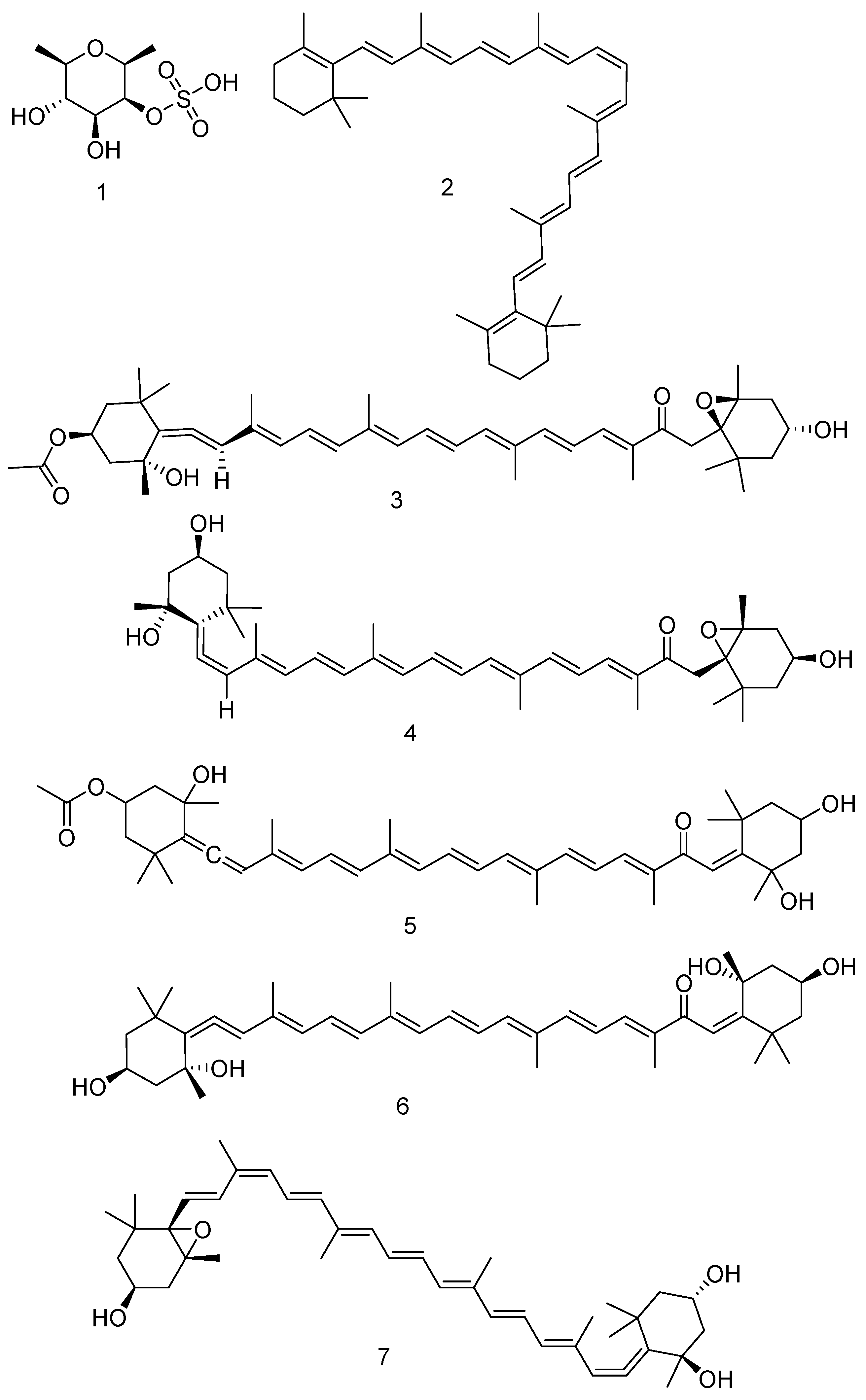{kind=link}
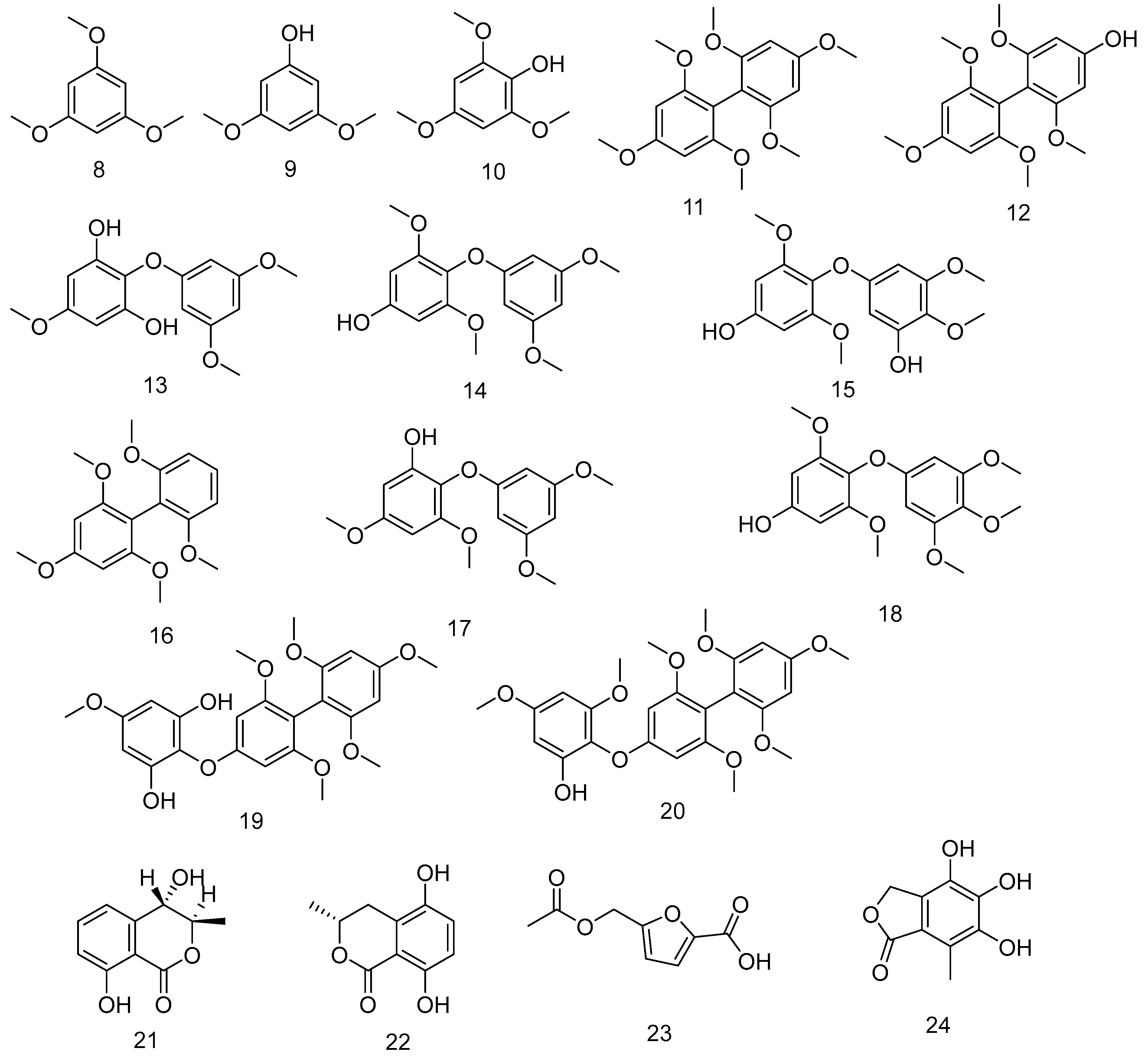{kind=link}
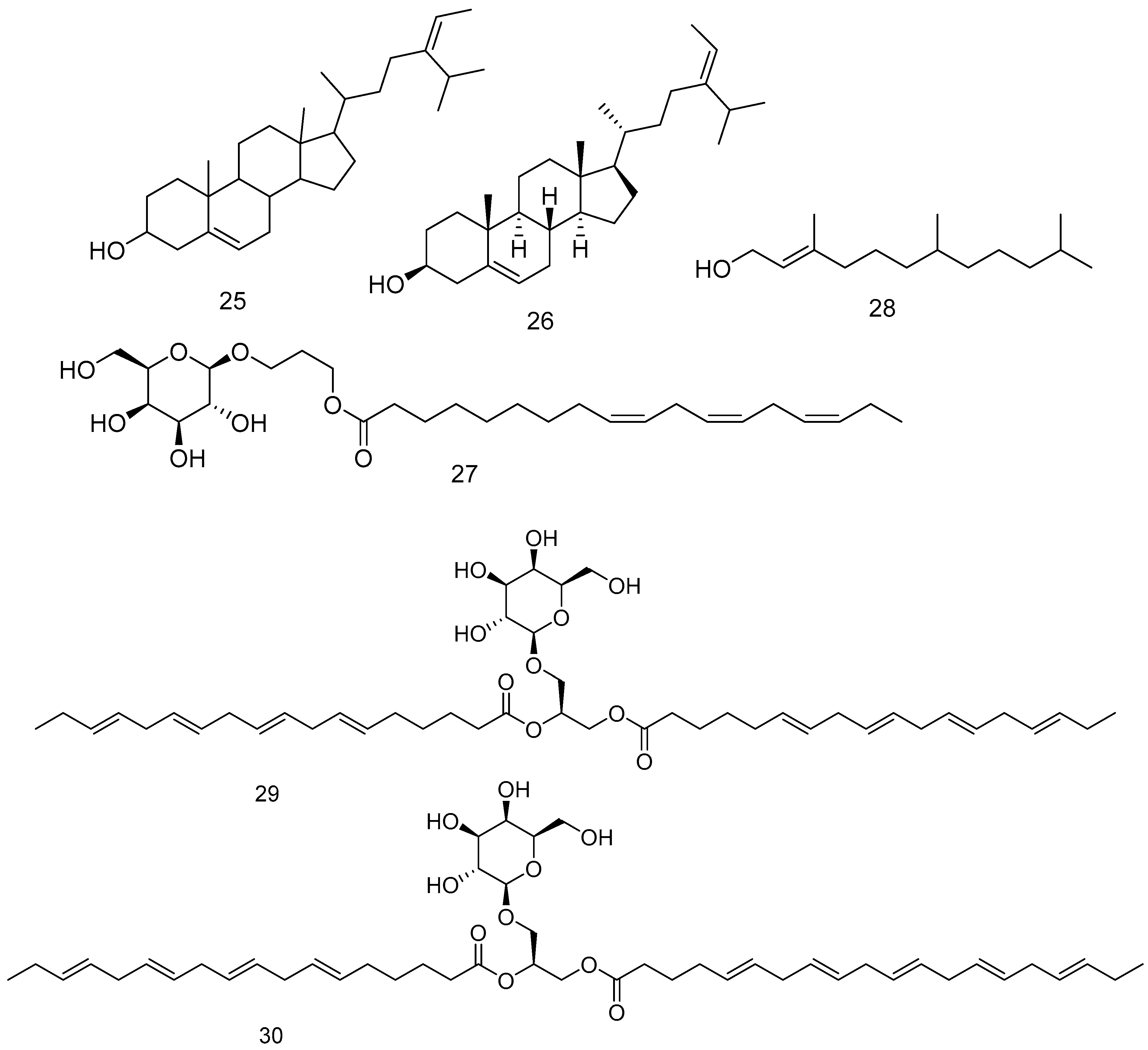{kind=link}
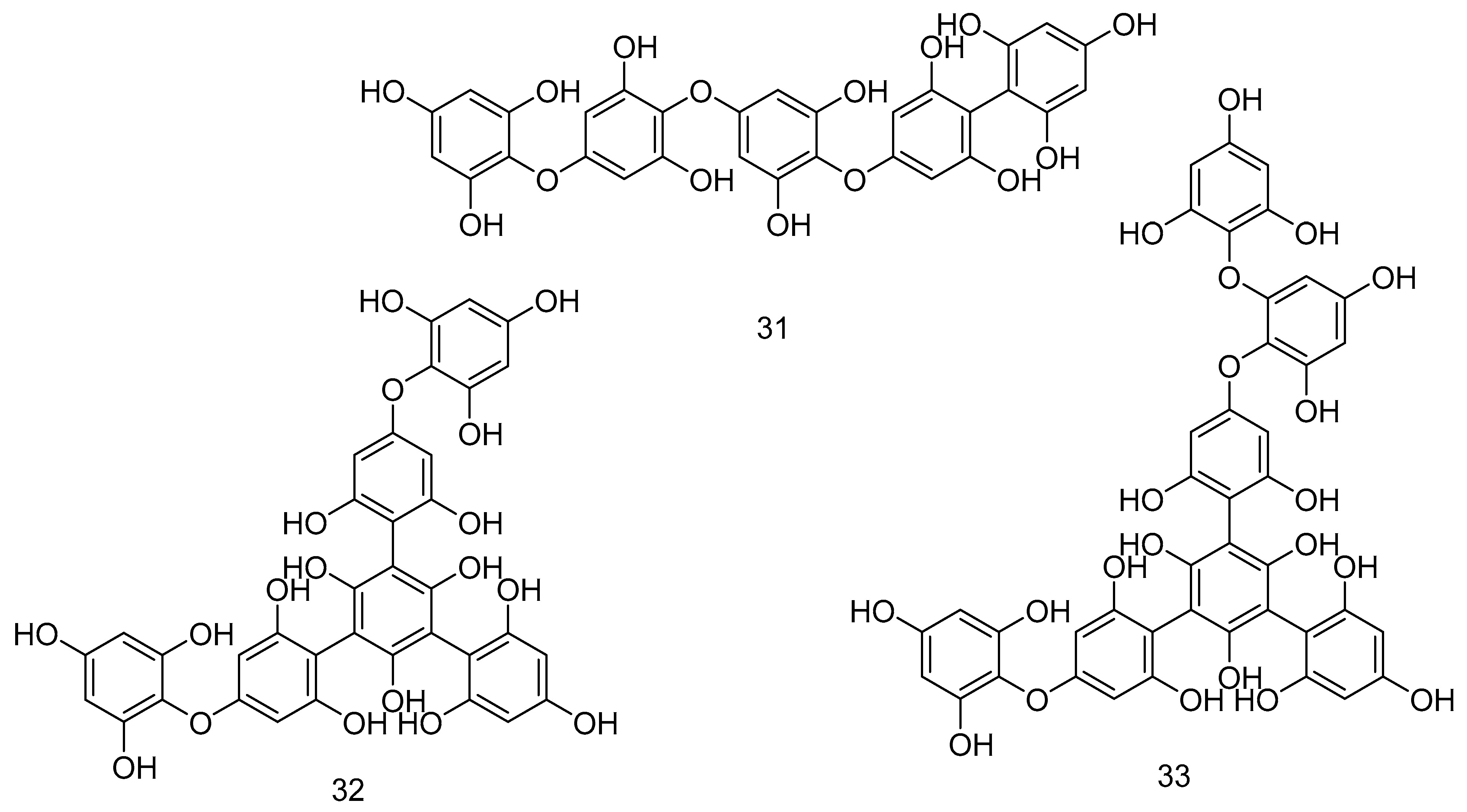{kind=link}
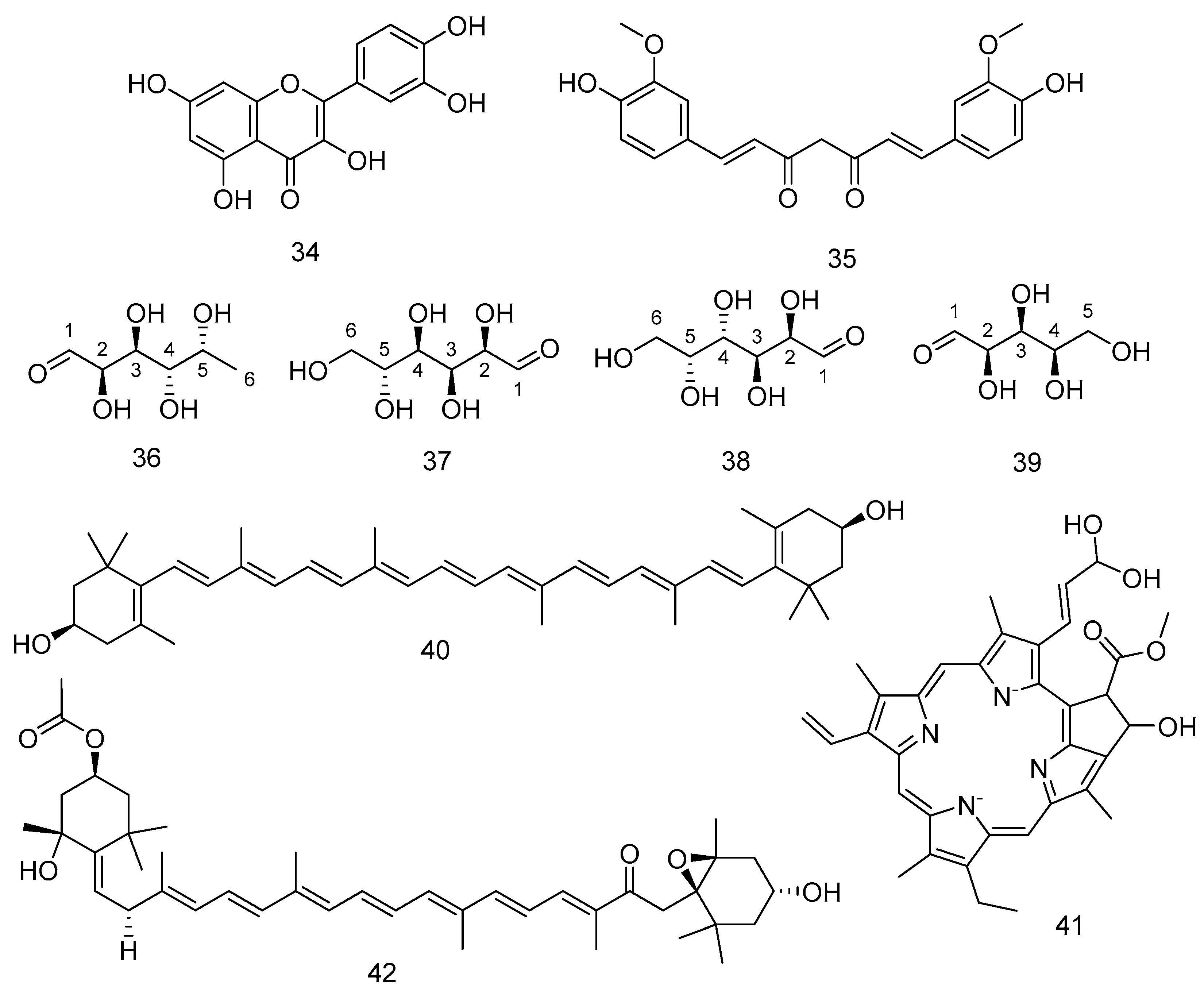{kind=link}
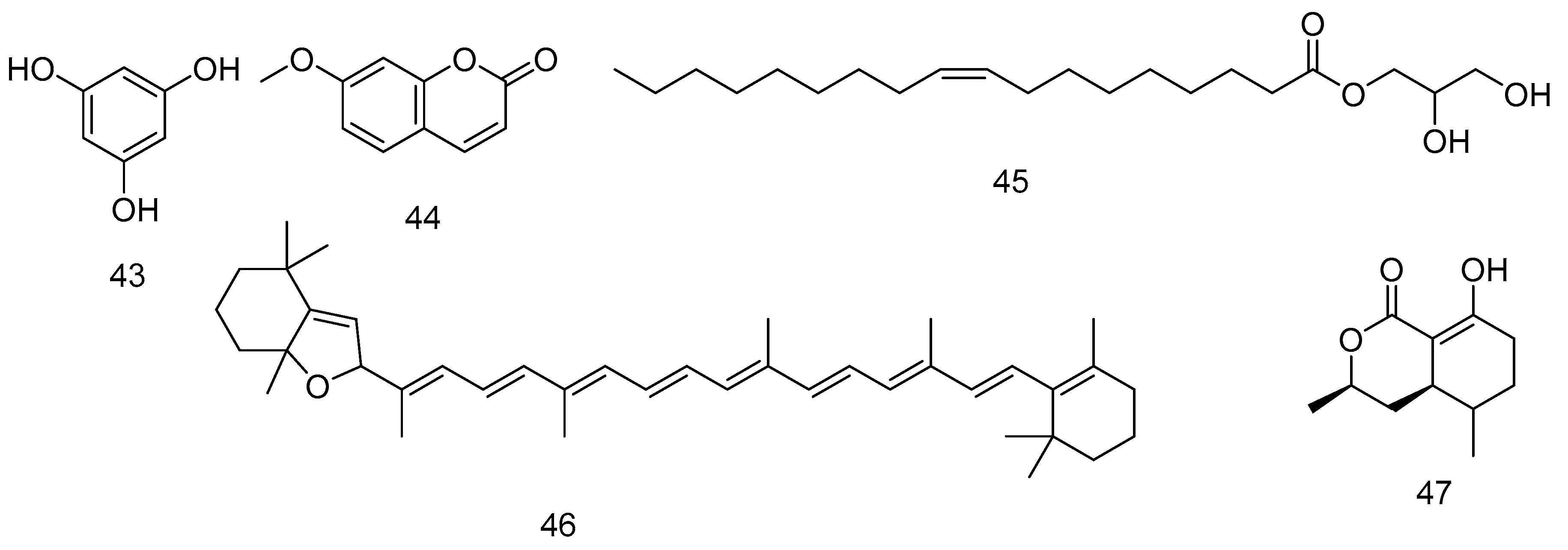{kind=link}
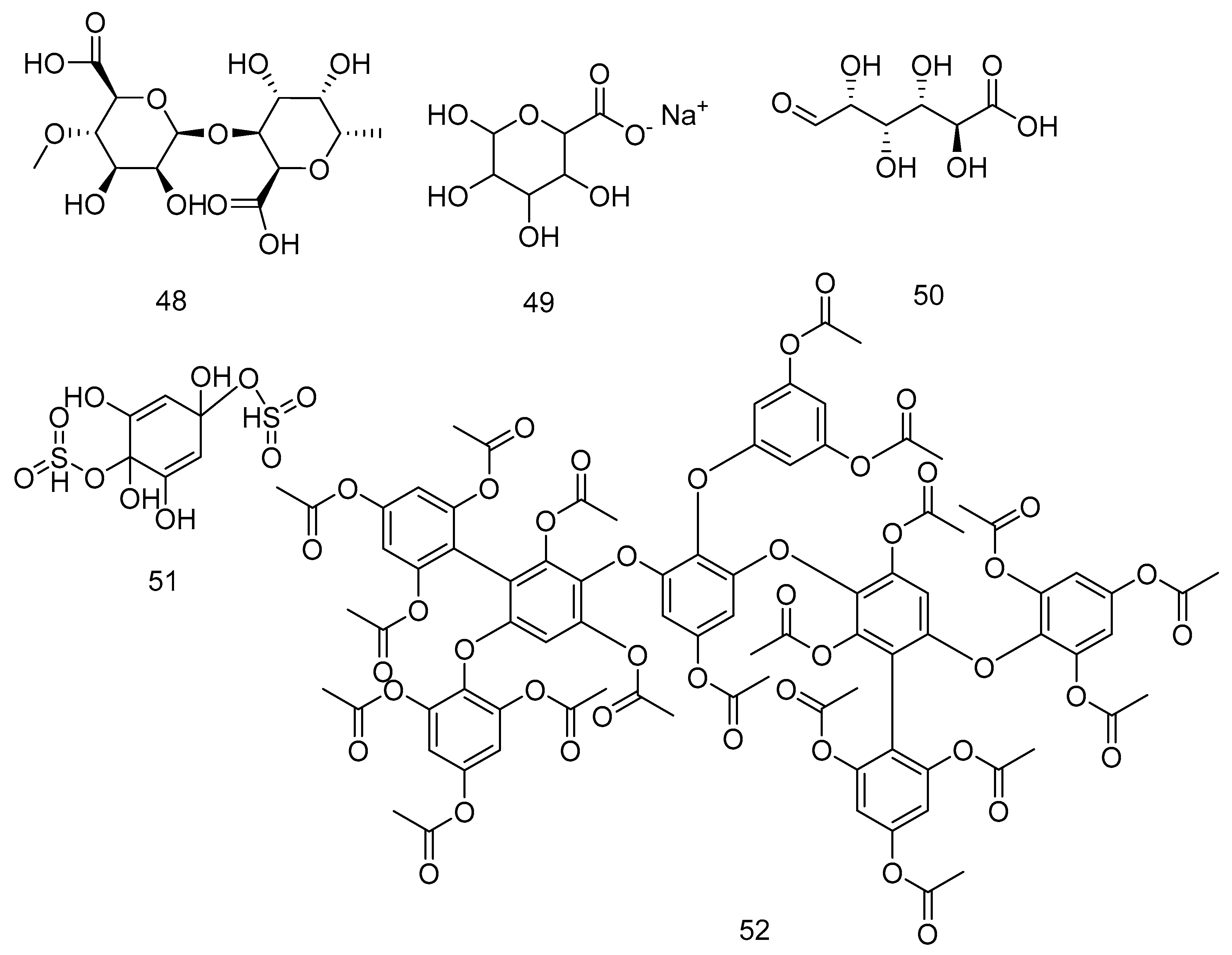{kind=link}
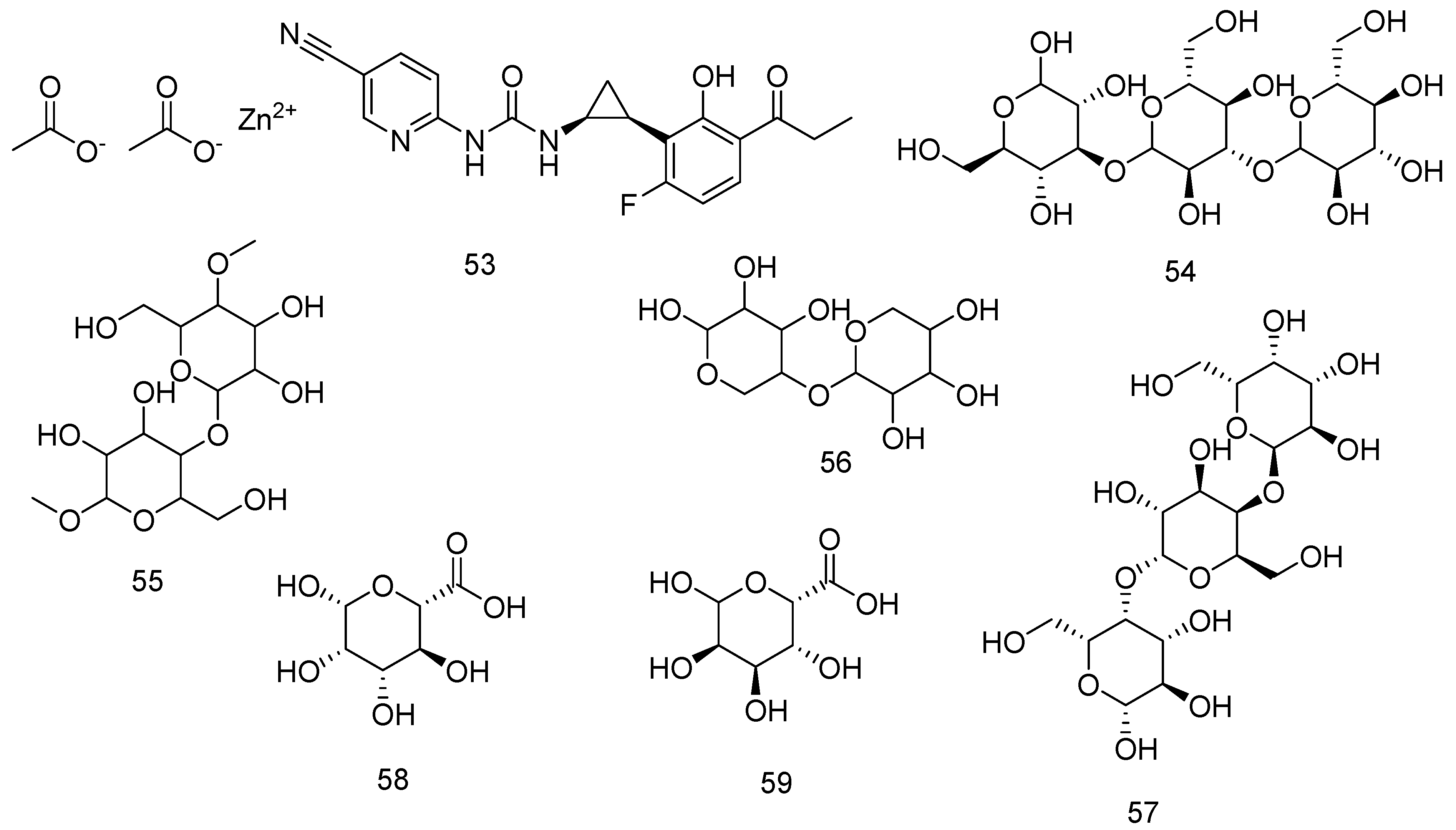{kind=link}
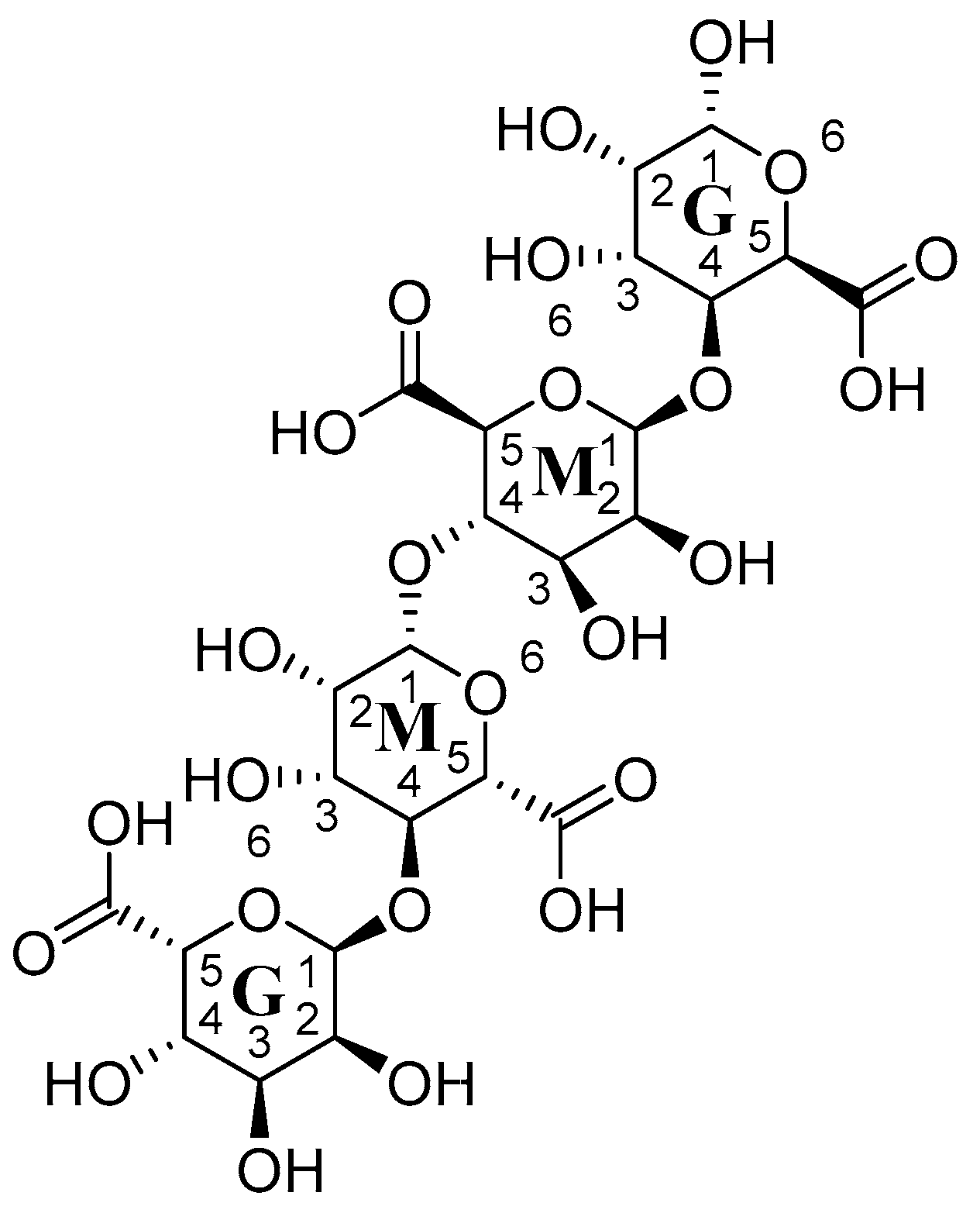{kind=link}
Table 1.
Compounds identified in Brown seaweeds (rockweeds) with their biological activity.
| Code | Compound Name | Original Algal Source | Biological Activities | Ref. |
|---|---|---|---|---|
| 1 | Fucoidan | F. vesiculosus | Effects on Lewis lung carcinoma cells (LCCs), melanoma B16 cells. Activated the natural killer cell activity in vivo in mice model. | [167] |
| In vivo effect as a suppressor to induced colorectal carcinogenesis in rats. | [168] | |||
| Low-molecular-weight fucoidan showed protective effect against renal fibrosis in animal in vitro and in vivo studies. | [169] | |||
| 2 | β-carotene | F. vesiculosus | It is the pro-vitamin A with high antioxidant activities. | [170] |
| Important role in metabolism of fat and regulation of body weight. | [171] | |||
| 3 | Fucoxanthin | F. vesiculosus | Good intervention with some types of tumours. | [172] |
| Anticancer effects. | [173] | |||
| 4 | Fucoxanthinol | F. vesiculosus | Anti-inflammatory. | [174,175] |
| Anticancer effects. | [173] | |||
| 5 | Isofucoxanthin | F. vesiculosus and F. serratus | - | [176,177] |
| 6 | Isofucoxanthinol | F. vesiculosus | - | [176] |
| 7 | Neoxanthin | F. vesiculosus | - | [174] |
| 8 | 1,3,5-trimethoxybenzene | Controlled cleavage process of polyphenolic compounds extracted from F. vesiculosus | - | [178] |
| 9 | 3,5-dimethoxyphenol | |||
| 10 | 2,4,6-trimethoxyphenol | |||
| 11 | 2,2′,4,4′,6,6′-hexamethoxy-1,1′-biphenyl | |||
| 12 | 4-hydroxy-2,6,2′,4′,6′-pentamethoxybiphenyl | |||
| 13 | 2,6-dihydroxy-4,3′,5′-trimethoxydiphenyl ether | |||
| 14 | 4-hydroxy-2,6,3′,5′-tetramethoxydiphenyl ether | |||
| 15 | 4,3′-dihydroxy-2,6,4′,5′-tetramethoxydiphenyl ether | |||
| 16 | 2,4,6,2′,6′-pentamethoxybiphenyl | |||
| 17 | 2-hydroxy-4,6,3′,5′-tetramethoxydiphenyl ether | |||
| 18 | 4-hydroxy-2,6,3′,4′,5′-pentamethoxydiphenyl ether | |||
| 19 | 2,6,2′,4′,6′-pentamethoxy-4-(2,6-dihydroxy-4-methoxyphenoxy) biphenyl | |||
| 20 | 2,6,2′,4′,6′-pentamethoxy-4-(2-hydroxy-4,6-dimethoxyphenoxy) biphenyl | |||
| 21 | Trans-4-Hydroxymellein | From the endophyte Epicoccum sp. derived from the brown algae F. vesiculosus | Antioxidant activities | [179] |
| 22 | (3R)-5-hydroxymellein | |||
| 23 | 5-(acetoxymethyl)-furan-2-carboxylic acid | |||
| 24 | 4,5,6-trihydroxy-7-methylphthalide | |||
| 25 | (3β,24E)-stigmasta-5,24(28)-dien-3-ol (Isofucosterol) | F. vesiculosus and F. spiralis | Lowering total cholesterol and harmful low-density lipoproteins | [180,181,182] |
| 26 | Fucosterol | F. vesiculosus, F. spiralis, and P. canaliculata | [182,183,184] | |
| 27 | 1,2-Di-O-α-linolenoyl-3-O-β-galactopyranosyl-sn-glycerol | F. vesiculosus | - | [185] |
| 28 | 3,7,11-Trimethyl-2-dodecen-1-ol | F. vesiculosus | - | [186] |
| 29 | Heterosigma-glycolipid I | F. vesiculosus | - | [185] |
| 30 | Heterosigma-glycolipid II | |||
| 31 | Fucotriphlorethol A | F. vesiculosus L. | Free radical scavenging activities. | [187] |
| 32 | Trifucodiphlorethol A | F. vesiculosus L. | Free radical scavenging activities. | [187] |
| 33 | Trifucotriphlorethol A | F. vesiculosus L. | Free radical scavenging activities. | [187] |
| 34 | Quercetin | F. vesiculosus | Neuroprotective promoting the neuronal plasticity. | [188] |
| 35 | Curcumin | F. vesiculosus | Neuroprotective promoting the neuronal plasticity. | [188] |
| 36 | Fucose | F. vesiculosus | Sulfated polysaccharides have antioxidants properties | [189] |
| 37 | Glucose | |||
| 38 | Galactose | |||
| 39 | Xylose | |||
| 40 | zeaxanthin | F. serratus | - | [190] |
| 41 | Chlorophyll c1 | [177] | ||
| 42 | All-trans-fucoxanthin | [191] | ||
| 43 | Phloroglucinol | F. vesiculosus | Against the chronic inflammation consequences. | [178,192,193] |
| Antithrombotic and profibrinolytic. | [194] | |||
| 44 | 7-methoxycoumarin | F. spiralis | Estrogenic effects. | [195] |
| Hepatoprotective effects. | [196] | |||
| 45 | 1-monooleoyl glycerol | F. spiralis | [197] | |
| 46 | β-carotene 5,8-epoxide (Citroxanthin) | F. spiralis | Antidiabetic-retinopathy effect. | [198] |
| 47 | 5-hydroxyramulosin | Endophyte associated with F. spiralis | Strong antifungal effect IC50 1.56 µg/mL against Aspergillus niger. | [199,200] |
| 48 | Alginic acid | L. digitata and A. nodosum | Moisturise the skin | [201] |
| 49 | Alginate | A. nodosum | Anticancer drug vehicle | [202] |
| 50 | Glucuronic acid | A. nodosum | - | [203] |
| 51 | 1,2,3,5-tetrahydroxybenzol-2,5-disulfate ester | A. nodosum | - | [204] |
| 52 | Bis-fucopentaphlorethol A nonadecaactate | H. elongata | - | [205] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Al-Dulaimi, O.; Rateb, M.E.; Hursthouse, A.S.; Thomson, G.; Yaseen, M. The Brown Seaweeds of Scotland, Their Importance and Applications. Environments 2021, 8, 59. https://0-doi-org.brum.beds.ac.uk/10.3390/environments8060059
AMA Style
Al-Dulaimi O, Rateb ME, Hursthouse AS, Thomson G, Yaseen M. The Brown Seaweeds of Scotland, Their Importance and Applications. Environments. 2021; 8(6):59. https://0-doi-org.brum.beds.ac.uk/10.3390/environments8060059
Chicago/Turabian StyleAl-Dulaimi, Omar, Mostafa E. Rateb, Andrew S. Hursthouse, Gary Thomson, and Mohammed Yaseen. 2021. "The Brown Seaweeds of Scotland, Their Importance and Applications" Environments 8, no. 6: 59. https://0-doi-org.brum.beds.ac.uk/10.3390/environments8060059
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.