Anti-Inflammatory Properties of Mineral-Balanced Deep Sea Water in In-Vitro and In-Vivo Models of Inflamed Intestinal Epithelium


Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of DSW and DSW-Derived Mineral Waters
2.3. Quantification of Minerals in DSW Using Inductively Coupled Plasma Atomic Emission Spectroscopy
2.4. Cell Culture
2.5. Coculture for Inducing Inflammatory Reaction
2.6. Transepithelial Electrical Resistance (TEER) Measurement
2.7. Permeability Measurement
2.8. Total RNA Extraction and Quantitative Reverse Transcription (RT-q) PCR
2.9. Animals
2.10. Colitis Model and Disease
2.11. DAI Scoring
2.12. Histological Analysis
2.13. In-Vivo Intestinal Permeability Measurement
2.14. Statistical Analysis
2.15. Total RNA Sequencing
2.16. Bioinformatics Analysis
2.17. Functional Enrichment Analysis
3. Results
3.1. Mineral Contents of DSW-Derived Water Samples
3.2. Effect of DSW-Derived Mineral Waters on Intestinal Permeability
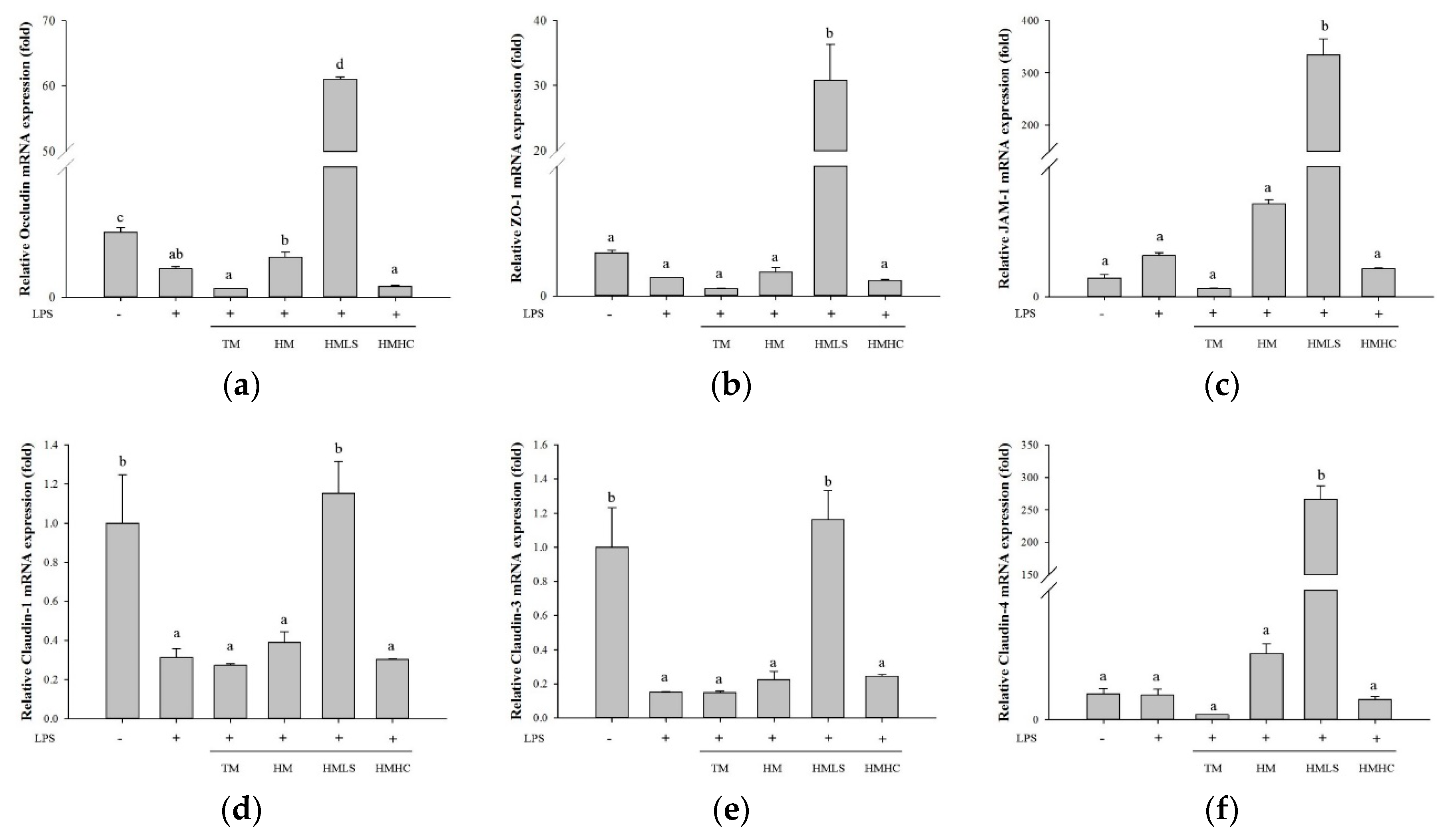
3.3. Effects of DSW-Derived Mineral Waters on the Expression of TJ-Related Proteins
3.4. DSW-Derived Mineral Waters Improve Clinical Symptoms of DSS-Induced Colitis
3.5. Mineral Water Intake Reduces DSS-Induced Colonic Histopathological Changes
3.6. Intestinal Permeability
3.7. Differential Gene Expression Analysis
3.8. PPI Network Analysis
3.9. GO and KEGG Enrichment Analyses of DEG
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Podolsky, D.K. Inflammatory bowel disease. N. Engl. J. Med. 1991, 325, 1008–1016. [Google Scholar] [CrossRef]
- Guo, T.; Song, D.; Cheng, L.; Zhang, X. Interactions of tea catechins with intestinal microbiota and their implication for human health. Food Sci. Biotechnol. 2019, 28, 1617–1625. [Google Scholar] [CrossRef]
- Mowat, C.; Cole, A.; Windsor, A.; Ahmad, T.; Arnott, I.; Driscoll, R.; Mitton, S.; Orchard, T.; Rutter, M.; Younge, L. Guidelines for the management of inflammatory bowel disease in adults. Gut 2011, 60, 571–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhave, M.; Winter, H.S.; Grand, R.J. Azathioprine in the treatment of children with inflammatory bowel disease. J. Pediatrics 1990, 117, 809–814. [Google Scholar] [CrossRef]
- Neurath, M.F. Cytokines in inflammatory bowel disease. Nat. Rev. Immunol. 2014, 14, 329–342. [Google Scholar] [CrossRef]
- Strober, W.; Fuss, I.J.; Blumberg, R.S. The immunology of mucosal models of inflammation. Annu. Rev. Immunol. 2002, 20, 495–549. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A. Leaky gut and autoimmune diseases. Clin. Rev. Allergy Immunol. 2012, 42, 71–78. [Google Scholar] [CrossRef]
- Edelblum, K.L.; Turner, J.R. The tight junction in inflammatory disease: Communication breakdown. Curr. Opin. Pharmacol. 2009, 9, 715–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Mariscal, L.; Contreras, R.; Bolivar, J.; Ponce, A.; Chavez De Ramirez, B.; Cereijido, M. Role of calcium in tight junction formation between epithelial cells. Am. J. Physiol. Cell Physiol. 1990, 259, C978–C986. [Google Scholar] [CrossRef]
- Ma, T.Y.; Tran, D.; Hoa, N.; Nguyen, D.; Merryfield, M.; Tarnawski, A. Mechanism of extracellular calcium regulation of intestinal epithelial tight junction permeability: Role of cytoskeletal involvement. Microsc. Res. Tech. 2000, 51, 156–168. [Google Scholar] [CrossRef]
- Lechuga, S.; Ivanov, A.I. Disruption of the epithelial barrier during intestinal inflammation: Quest for new molecules and mechanisms. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2017, 1864, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, B.J.; Tuft, B.; Elfrey, J.E.; Smith, A.; Zhao, A.; Morimoto, M.; Chmielinska, J.J.; Tejero-Taldo, M.I.; Mak, I.T.; Weglicki, W.B. Intestinal inflammation caused by magnesium deficiency alters basal and oxidative stress-induced intestinal function. Mol. Cell. Biochem. 2007, 306, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Weglicki, W.B.; Mak, I.T.; Chmielinska, J.J.; Tejero-Taldo, M.I.; Komarov, A.; Kramer, J.H. The role of magnesium deficiency in cardiovascular and intestinal inflammation. Magnes. Res. Off. Organ Int. Soc. Dev. Res. Magnes. 2010, 23, S199. [Google Scholar]
- Heaney, R.; Dowell, M. Absorbability of the calcium in a high-calcium mineral water. Osteoporos. Int. 1994, 4, 323–324. [Google Scholar] [CrossRef] [PubMed]
- Kiss, S.A.; Forster, T.; Dongó, Á. Absorption and effect of the magnesium content of a mineral water in the human body. J. Am. Coll. Nutr. 2004, 23, 758S–762S. [Google Scholar] [CrossRef]
- Böhmer, H.; Müller, H.; Resch, K.-L. Calcium supplementation with calcium-rich mineral waters: A systematic review and meta-analysis of its bioavailability. Osteoporos. Int. 2000, 11, 938–943. [Google Scholar] [CrossRef]
- Sabatier, M.; Arnaud, M.J.; Kastenmayer, P.; Rytz, A.; Barclay, D.V. Meal effect on magnesium bioavailability from mineral water in healthy women. Am. J. Clin. Nutr. 2002, 75, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Aamodt, G.; Bukholm, G.; Jahnsen, J.; Moum, B.; Vatn, M.H.; Group, I.S. The association between water supply and inflammatory bowel disease based on a 1990–1993 cohort study in southeastern Norway. Am. J. Epidemiol. 2008, 168, 1065–1072. [Google Scholar] [CrossRef]
- Nani, M.; Zura, S.; Majid, F.A.A.; Jaafar, A.B.; Mahdzir, A.; Musa, M.N. Potential health benefits of deep sea water: A review. Evid. Based Complement. Altern. Med. 2016, 2016. [Google Scholar] [CrossRef]
- Hwang, H.S.; Kim, S.H.; Yoo, Y.G.; Chu, Y.S.; Shon, Y.H.; Nam, K.S.; Yun, J.W. Inhibitory effect of deep-sea water on differentiation of 3T3-L1 adipocytes. Mar. Biotechnol. 2009, 11, 161–168. [Google Scholar] [CrossRef]
- Ha, B.; Shin, E.; Park, J.-E.; Shon, Y. Anti-diabetic effect of balanced deep-sea water and its mode of action in high-fat diet induced diabetic mice. Mar. Drugs 2013, 11, 4193–4212. [Google Scholar] [CrossRef]
- Hsu, T.-C.; Chiu, C.-C.; Lin, H.-L.; Kao, T.-W.; Chen, L.-J.; Wu, L.-Y.; Huang, C.-Y.; Tzang, B.-S. Attenuated effects of deep-sea water on hepatic apoptosis in STZ-induced diabetic rats. Chin. J. Physiol. 2015, 58, 197–205. [Google Scholar] [PubMed] [Green Version]
- Bak, J.-P.; Kim, Y.-M.; Son, J.; Kim, C.-J.; Kim, E.-H. Application of concentrated deep sea water inhibits the development of atopic dermatitis-like skin lesions in NC/Nga mice. BMC Complement. Altern. Med. 2012, 12, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimata, H.; Tai, H.; Nakagawa, K.; Yokoyama, Y.; Nakajima, H.; Ikegami, Y. Improvement of skin symptoms and mineral imbalance by drinking deep sea water in patients with atopic eczema/dermatitis syndrome (AEDS). Acta Med. (Hradec Kralove) 2002, 45, 83–84. [Google Scholar] [CrossRef]
- Kimata, H.; Tai, H.; Nakajima, H. Reduction of allergic skin responses and serum allergen-specific IgE and IgE-inducing cytokines by drinking deep-sea water in patients with allergic rhinitis. Oto-Rhino-Laryngol. Nova 2001, 11, 302–303. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Jung, I.-S.; Song, H.-J.; Choi, E.-Y.; Choi, I.-S.; Choi, Y.-J. Study of deep ground sea-like water on antioxidant activity and the immune response in RAW264. 7 Macrophages. J. Life Sci. 2008, 18, 329–335. [Google Scholar] [CrossRef]
- Jung, S.; Joo, E.; Yoo, J.; Kim, Y.; Cho, Y.; Yoon, B.; Cho, J.; Nam, K.; Kwang, S. Effect of the supply of natural water from deep sea rock on the immune response and antioxidant activity in rats. J. Anim. Sci. Technol. 2006. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.S.; Chun, S.Y.; Kwon, Y.S.; Kim, S.; Nam, K.S. Deep sea water improves hypercholesterolemia and hepatic lipid accumulation through the regulation of hepatic lipid metabolic gene expression. Mol. Med. Rep. 2017, 15, 2814–2822. [Google Scholar] [CrossRef]
- Ha, B.G.; Park, J.-E.; Cho, H.-J.; Shon, Y.H. Stimulatory effects of balanced deep sea water on mitochondrial biogenesis and function. PLoS ONE 2015, 10, e0129972. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.-J.; Don, T.-M.; Lin, C.-W.; Mi, F.-L. Delivery of berberine using Chitosan/Fucoidan-Taurine conjugate nanoparticles for treatment of defective intestinal epithelial tight junction barrier. Mar. Drugs 2014, 12, 5677–5697. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2- ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Cooper, H.S.; Murthy, S.; Shah, R.; Sedergran, D. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Investig. A J. Tech. Methods Pathol. 1993, 69, 238–249. [Google Scholar]
- Rees, V. Chronic experimental colitis induced by dextran sulphate sodium (DSS) is characterized by Th1 and Th2 cytokines. Clin. Exp. Immunol. 1998, 114, 385–391. [Google Scholar]
- Tambuwala, M.M.; Cummins, E.P.; Lenihan, C.R.; Kiss, J.; Stauch, M.; Scholz, C.C.; Fraisl, P.; Lasitschka, F.; Mollenhauer, M.; Saunders, S.P. Loss of prolyl hydroxylase-1 protects against colitis through reduced epithelial cell apoptosis and increased barrier function. Gastroenterology 2010, 139, 2093–2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562. [Google Scholar] [CrossRef] [Green Version]
- Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.-L.; Ideker, T. Cytoscape 2.8: New features for data integration and network visualization. Bioinformatics 2010, 27, 431–432. [Google Scholar] [CrossRef] [Green Version]
- Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar]
- Moon, D.-S.; Kim, K.S.; Gi, H.; Choi, M.Y.; Jung, H.J.; Kim, H.J. The hardness water production by RO/NF/ED linking process from deep seawater. J. Korean Soc. Mar. Environ. Energy 2013, 16, 227–238. [Google Scholar] [CrossRef]
- Wu, C.; Yosef, N.; Thalhamer, T.; Zhu, C.; Xiao, S.; Kishi, Y.; Regev, A.; Kuchroo, V.K. Induction of pathogenic T H 17 cells by inducible salt-sensing kinase SGK1. Nature 2013, 496, 513–517. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Pollock, N.K.; Kotak, I.; Gutin, B.; Wang, X.; Bhagatwala, J.; Parikh, S.; Harshfield, G.A.; Dong, Y. Dietary sodium, adiposity, and inflammation in healthy adolescents. Pediatrics 2014, 133, e635–e642. [Google Scholar] [CrossRef] [Green Version]
- Arora, M. Cell culture media: A review. Mater Methods 2013, 3, 24. [Google Scholar] [CrossRef]
- Tanoue, T.; Nishitani, Y.; Kanazawa, K.; Hashimoto, T.; Mizuno, M. In Vitro model to estimate gut inflammation using co-cultured Caco-2 and RAW264. 7 cells. Biochem. Biophys. Res. Commun. 2008, 374, 565–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, T.Y.; Boivin, M.A.; Ye, D.; Pedram, A.; Said, H.M. Mechanism of TNF-α modulation of Caco-2 intestinal epithelial tight junction barrier: Role of myosin light-chain kinase protein expression. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G422–G430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKay, D.M.; Baird, A.W. Cytokine regulation of epithelial permeability and ion transport. Gut 1999, 44, 283–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, V.W.; Goodenough, D.A. Paracellular ion channel at the tight junction. Biophys. J. 2003, 84, 1660–1673. [Google Scholar] [CrossRef] [Green Version]
- Vermette, D.; Hu, P.; Canarie, M.F.; Funaro, M.; Glover, J.; Pierce, R.W. Tight junction structure, function, and assessment in the critically ill: A systematic review. ICMx 2018, 6, 37. [Google Scholar] [CrossRef] [Green Version]
- Itallie, C.V.; Rahner, C.; Anderson, J.M. Regulated expression of claudin-4 decreases paracellular conductance through a selective decrease in sodium permeability. J. Clin. Investig. 2001, 107, 1319–1327. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H. Intestinal permeability regulation by tight junction: Implication on inflammatory bowel diseases. Intest. Res. 2015, 13, 11. [Google Scholar] [CrossRef] [Green Version]
- Pachikian, B.D.; Neyrinck, A.M.; Deldicque, L.; De Backer, F.C.; Catry, E.; Dewulf, E.M.; Sohet, F.M.; Bindels, L.B.; Everard, A.; Francaux, M. Changes in intestinal bifidobacteria levels are associated with the inflammatory response in magnesium-deficient mice. J. Nutr. 2010, 140, 509–514. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Shajib, M.S.; Manocha, M.M.; Khan, W.I. Investigating intestinal inflammation in DSS-induced model of IBD. JoVE J. Vis. Exp. 2012, e3678. [Google Scholar] [CrossRef] [Green Version]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran sulfate sodium (DSS)-induced colitis in mice. Curr. Protoc. Immunol. 2014, 104, 15.25. 1–15.25. 14. [Google Scholar] [CrossRef]
- Woo, J.K.; Choi, S.; Kang, J.-H.; Kim, D.E.; Hurh, B.-S.; Jeon, J.-E.; Kim, S.Y.; Oh, S.H. Fermented barley and soybean (BS) mixture enhances intestinal barrier function in dextran sulfate sodium (DSS)-induced colitis mouse model. Bmc Complement. Altern. Med. 2016, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Kim, J.Y. Ginger attenuates inflammation in a mouse model of dextran sulfate sodium-induced colitis. Food Sci. Biotechnol. 2018, 27, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Al-Sadi, R.; Said, H.M.; Ma, T.Y. Lipopolysaccharide causes an increase in intestinal tight junction permeability In Vitro and In Vivo by inducing enterocyte membrane expression and localization of TLR-4 and CD14. Am. J. Pathol. 2013, 182, 375–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeRoche, T.C.; Xiao, S.-Y.; Liu, X. Histological evaluation in ulcerative colitis. Gastroenterol. Rep. 2014, 2, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Erben, U.; Loddenkemper, C.; Doerfel, K.; Spieckermann, S.; Haller, D.; Heimesaat, M.M.; Zeitz, M.; Siegmund, B.; Kühl, A.A. A guide to histomorphological evaluation of intestinal inflammation in mouse models. Int. J. Clin. Exp. Pathol. 2014, 7, 4557. [Google Scholar] [PubMed]
- Trapani, V.; Petito, V.; Di Agostini, A.; Arduini, D.; Hamersma, W.; Pietropaolo, G.; Luongo, F.; Arena, V.; Stigliano, E.; Lopetuso, L.R. Dietary magnesium alleviates experimental murine colitis through upregulation of the transient receptor potential melastatin 6 channel. Inflamm. Bowel Dis. 2018, 24, 2198–2210. [Google Scholar] [CrossRef] [PubMed]
- Poritz, L.S.; Garver, K.I.; Green, C.; Fitzpatrick, L.; Ruggiero, F.; Koltun, W.A. Loss of the tight junction protein ZO-1 in dextran sulfate sodium induced colitis. J. Surg. Res. 2007, 140, 12–19. [Google Scholar] [CrossRef]
- Scharl, M.; R Vavricka, S.; Rogler, G. New anti-cytokines for IBD: What is in the pipeline? Curr. Drug Targets 2013, 14, 1405–1420. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.-M.; An, J. Cytokines, inflammation and pain. Int. Anesthesiol. Clin. 2007, 45, 27. [Google Scholar] [CrossRef] [Green Version]
- Rutgeerts, P.; Van Assche, G.; Vermeire, S. Optimizing anti-TNF treatment in inflammatory bowel disease. Gastroenterology 2004, 126, 1593–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, J.; Romani, A.M.; Valentin-Torres, A.M.; Luciano, A.A.; Kitchen, C.M.R.; Funderburg, N.; Mesiano, S.; Bernstein, H.B. Magnesium decreases inflammatory cytokine production: A novel innate immunomodulatory mechanism. J. Immunol. 2012, 188, 6338–6346. [Google Scholar] [CrossRef] [Green Version]
- Mazzone, R.; Zwergel, C.; Artico, M.; Taurone, S.; Ralli, M.; Greco, A.; Mai, A. The emerging role of epigenetics in human autoimmune disorders. Clin. Epigenetics 2019, 11, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Mahon, B.D.; Froicu, M.; Cantorna, M.T. Calcium and 1α, 25-dihydroxyvitamin D3 target the TNF-α pathway to suppress experimental inflammatory bowel disease. Eur. J. Immunol. 2005, 35, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Gao, G.; Schneider, S.; Helt, C.; Weiss, C.; O’Reilly, M.A.; Bohmann, D.; Zhao, J. DNA damage induces downregulation of histone gene expression through the G1 checkpoint pathway. Embo J. 2004, 23, 1133–1143. [Google Scholar] [CrossRef] [Green Version]
- Mariño-Ramírez, L.; Kann, M.G.; Shoemaker, B.A.; Landsman, D. Histone structure and nucleosome stability. Expert Rev. Proteom. 2005, 2, 719–729. [Google Scholar] [CrossRef]
- Becker, K.A.; Stein, J.L.; Lian, J.B.; Van Wijnen, A.J.; Stein, G.S. Establishment of histone gene regulation and cell cycle checkpoint control in human embryonic stem cells. J. Cell. Physiol. 2007, 210, 517–526. [Google Scholar] [CrossRef]
- Beckett, E.L.; Yates, Z.; Veysey, M.; Duesing, K.; Lucock, M. The role of vitamins and minerals in modulating the expression of microRNA. Nutr. Res. Rev. 2014, 27, 94–106. [Google Scholar] [CrossRef] [Green Version]
- Dai, Q.; Shu, X.-O.; Deng, X.; Xiang, Y.-B.; Li, H.; Yang, G.; Shrubsole, M.J.; Ji, B.; Cai, H.; Chow, W.-H. Modifying effect of calcium/magnesium intake ratio and mortality: A population-based cohort study. BMJ Open 2013, 3, e002111. [Google Scholar] [CrossRef] [Green Version]
- Rowe, W.J. Calcium-magnesium-ratio intake and cardiovascular risk. Am. J. Cardiol. 2006, 98, 140. [Google Scholar] [CrossRef]
- Zhao, J.; Giri, A.; Zhu, X.; Shrubsole, M.J.; Jiang, Y.; Guo, X.; Ness, R.; Seidner, D.L.; Giovannucci, E.; Edwards, T.L. Calcium: Magnesium intake ratio and colorectal carcinogenesis, results from the prostate, lung, colorectal, and ovarian cancer screening trial. Br. J. Cancer 2019, 121, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Dai, Q.; Sandler, R.S.; Barry, E.L.; Summers, R.W.; Grau, M.V.; Baron, J.A. Calcium, magnesium, and colorectal cancer. Epidemiology (Camb. Mass.) 2012, 23, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, M.H.; Lee, D.G.; Go, E.B.; Cho, M.; Park, Y.S.; Chung, N. Anti-diabetic effect of magnesium salt extracts from deep-sea water in C57BLKS/J-db/db mice. Appl. Biol. Chem. 2017, 60, 95–99. [Google Scholar] [CrossRef]
- Maxwell, T.; Lee, K.-S.; Chun, S.-Y.; Nam, K.-S. Mineral-balanced deep sea water enhances the inhibitory effects of chitosan oligosaccharide on atopic dermatitis-like inflammatory response. Biotechnol. Bioprocess. Eng. 2017, 22, 120–128. [Google Scholar] [CrossRef]
- Yokota, J.; Kitaoka, T.; Jobu, K.; Takuma, D.; Hamada, A.; Onogawa, M.; Yoshioka, S.; Kyotani, S.; Miyamura, M. Eriobotrya japonica seed extract and deep sea water protect against indomethacin-induced gastric mucosal injury in rats. J. Nat. Med. 2011, 65, 9–17. [Google Scholar] [CrossRef]
- Yuan, H.; Chung, S.; Ma, Q.; Ye, L.; Piao, G. Combination of deep sea water and Sesamum indicum leaf extract prevents high-fat diet-induced obesity through AMPK activation in visceral adipose tissue. Exp. Ther. Med. 2016, 11, 338–344. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.W.; Shim, J.M.; Kim, D.W.; Yao, Z.; Kim, J.A.; Kim, H.-J.; Kim, J.H. Effects of different types of salts on the growth of lactic acid bacteria and yeasts during kimchi fermentation. Food Sci. Biotechnol. 2018, 27, 489–498. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Mineral Contents (ppm) | Na/Mg | Mg/Ca | |||
|---|---|---|---|---|---|---|
| Na+ | Mg2+ | K+ | Ca2+ | |||
| DSW | 10,700 | 1380 | 480 | 390 | 7.75 | 3.5 |
| TM | 29.5 | 6.7 | 1.7 | 2.4 | 4.4 | 2.8 |
| HM | 39.9 | 248 | 45.2 | 0.78 | 0.16 | 318 |
| HMLS | 36 | 179 | 1.7 | 41.5 | 0.2 | 4.3 |
| HMHC | 133 | 211 | 48 | 200 | 0.63 | 1.1 |
| Gene ID | Gene Name | Fold Change | FDR |
|---|---|---|---|
| Downregulated | |||
| NOS2 | Nitric oxide synthase 2 | 0.129 | 1.13 × 10−3 |
| C2cd4b | C2 calcium dependent domain containing 4B | 0.163 | 1.13 × 10−3 |
| Ccl8 | C-C motif chemokine ligand 8 | 0.222 | 9.43 × 10−2 |
| Reg3g | Regenerating family member 3 gamma | 0.372 | 1.13 × 10−3 |
| Serpina3n | Serine (or cysteine) peptidase inhibitor, clade A, member 3N | 0.380 | 1.13 × 10−3 |
| Crlf2 | Cytokine receptor like factor 2 | 0.413 | 9.42 × 10−3 |
| Tac4 | Tachykinin 4 | 0.431 | 8.63 × 10−2 |
| Ido1 | Indoleamine 2,3-dioxygenase 1 | 0.434 | 1.13 × 10−3 |
| Gbp5 | Guanylate binding protein 5 | 0.442 | 3.10 × 10−2 |
| Hck | HCK proto-oncogene, Src family tyrosine kinase | 0.443 | 1.33 × 10−2 |
| Tnf | Tumor necrosis factor | 0.479 | 1.11 × 10−2 |
| Reg3a | Regenerating family member 3 alpha | 0.482 | 4.53 × 10−2 |
| Upregulated | |||
| Hmox1 | Heme oxygenase 1 | 2.027 | 5.79 × 10−3 |
| Ccl20 | C-C motif chemokine ligand 20 | 2.940 | 2.05 × 10−3 |
| Category | Term | ID | p | Benjamini |
|---|---|---|---|---|
| GO, cellular component | Nucleosome | GO:0000786 | 5.43 × 10−26 | 9.55 × 10−24 |
| Nuclear nucleosome | GO:0000788 | 9.14 × 10−21 | 8.04 × 10−19 | |
| Extracellular region | GO:0005576 | 1.04 × 10−11 | 6.10 × 10−10 | |
| Extracellular space | GO:0005615 | 9.96 × 10−10 | 4.38 × 10−8 | |
| KEGG pathways | Systemic lupus erythematosus | Mmu05322 | 2.95 × 10−24 | 4.34 × 10−22 |
| Alcoholism | Mmu05034 | 3.57 × 10−18 | 2.62 × 10−16 | |
| GO, biological process | Nucleosome assembly | GO:0006334 | 3.76 × 10−16 | 3.56 × 10−13 |
| Regulation of gene silencing | GO:0060968 | 4.25 × 10−12 | 2.27 × 10−9 | |
| DNA methylation on cytosine | GO:0032776 | 1.84 × 10−10 | 6.55 × 10−8 | |
| Xenophagy | GO:0098792 | 1.06 × 10−09 | 2.40 × 10−7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, J.; Kim, K.J.; Park, G.; Kim, B.G.; Jeong, G.-H.; Jeon, J.-e.; Hurh, B.S.; Kim, J.Y. Anti-Inflammatory Properties of Mineral-Balanced Deep Sea Water in In-Vitro and In-Vivo Models of Inflamed Intestinal Epithelium. Appl. Sci. 2020, 10, 5183. https://0-doi-org.brum.beds.ac.uk/10.3390/app10155183
Nam J, Kim KJ, Park G, Kim BG, Jeong G-H, Jeon J-e, Hurh BS, Kim JY. Anti-Inflammatory Properties of Mineral-Balanced Deep Sea Water in In-Vitro and In-Vivo Models of Inflamed Intestinal Epithelium. Applied Sciences. 2020; 10(15):5183. https://0-doi-org.brum.beds.ac.uk/10.3390/app10155183
Chicago/Turabian StyleNam, Jain, Kyeong Jin Kim, Geonhee Park, Byeong Goo Kim, Gwi-Hwa Jeong, Jong-eun Jeon, Byung Serk Hurh, and Ji Yeon Kim. 2020. "Anti-Inflammatory Properties of Mineral-Balanced Deep Sea Water in In-Vitro and In-Vivo Models of Inflamed Intestinal Epithelium" Applied Sciences 10, no. 15: 5183. https://0-doi-org.brum.beds.ac.uk/10.3390/app10155183