Foxn1 Control of Skin Function

Institute of Animal Reproduction and Food Research, Polish Academy of Sciences, 10-748 Olsztyn, Poland
Appl. Sci. 2020, 10(16), 5685; https://0-doi-org.brum.beds.ac.uk/10.3390/app10165685
Submission received: 28 July 2020
/
Revised: 12 August 2020
/
Accepted: 13 August 2020
/
Published: 16 August 2020
(This article belongs to the Special Issue Skin Tissue Engineering)
{kind=link}
{kind=link}
Abstract
:The forkhead box N1 (Foxn1) transcription factor regulates biological processes of the thymus and skin. Loss-of-function mutations in Foxn1 cause the nude phenotype in humans, mice, and rats, which is characterized by hairless skin and a lack of thymus. This review focuses on the role of Foxn1 in skin biology, including epidermal, dermal, and dermal white adipose tissue (dWAT) skin components. In particular, the role of Foxn1 in the scar-forming skin wound healing process is discussed, underscoring that Foxn1 inactivity in nude mice is permissive for scar-less cutaneous wound resolution.
1. Introduction
Skin, as the outermost organ, creates a barrier that protects the body from the external environment while acting as a sensing and responding organ to internal and external stimuli [1,2]. The skin is composed of three layers: epidermis, dermis, and subcutaneous tissue. The epidermis, the most superficial part of the skin, consists of several layers of keratinocytes, among which melanocytes (melanin-producing cells), Langerhans cells (part of the adaptive immune response), and Merkel cells (neuroendocrine cells with sensory function) are spread out. The basal layer of the epidermis, which is made up of proliferating, self-renewing keratinocytes, is attached to the basement membrane (BM) that separates the epidermis from the dermis. Keratinocytes in the basal layer undergo continuous asymmetric cell division, differentiate, and move continuously upwards, changing their phenotype to form stratum spinosum, followed by stratum granulosum to finally reach the outermost cornified layer (stratum corneum) [3].
The dermis is composed of collagens, elastin, and glycosaminoglycans to form the extracellular matrix (ECM), which is produced by fibroblasts, the major cellular component of the dermis [4]. Two major populations of dermal fibroblasts (DFs) have been characterized according to their developmental origin, differences in gene expression, positioning/location in the dermis and function: (1) papillary dermis, which is located in proximity to the epidermis and (2) reticular dermis which is situated below [5,6]. Recently, it has been recognized that cells with adipogenic potential form a third layer of the dermis, dermal white adipose tissue (dWAT), representing different fat depots from the subcutaneous layer [7,8,9,10].
The processes of keratinocyte proliferation, differentiation, and acquisition of progressively developed features are tightly controlled by transcription factors that regulate epidermal activities through downstream genes [3,11,12]. Increasing data suggest that epidermally expressed transcription factors act beyond host tissue, reaching dermal fibroblasts and dWAT in the dermal part of the skin [13,14,15]. AP-1, Pparβ/δ, HoxA3, HoxD3, Foxo1-3, Ovol1/2, and Wnt signalling, among others, have been shown to regulate intact and post-wounded skin [16,17,18,19]. These transcription factors are expressed in the epidermis but have also been shown to be expressed in other locations. In this review, we focus on Foxn1, a transcription factor, whose expression is limited to epithelial cells of the thymus and the skin, and its function in skin physiology.
2. Foxn1 Discovery, Structure, and Localization
Foxn1 belongs to the superclass Helix-turn-helix, class Fork head/winged helix of transcription factors. Named after the Drosophila melanogaster gene fork head (fkh), this family of transcriptional regulators received the name Forkhead box (Fox) [20].
There are 44 (mouse) and 50 (human) members of the Fox family that are divided into 19 subfamilies, from A to S. The common feature of Fox proteins is the highly conserved ~100-residue fork head DNA-binding domain [21]. The Fox gene family is an ancient class of transcription factors with broad functions in development, physiology, and pathology (i.e., cancer) [20]. Most Fox transcription factors act as transcriptional activators, although some (i.e., the FoxP family) has dual activator/suppressor functions [22]. The Foxn group consists of transcription factors named from 1 to 4.
The mapping and sequencing of the transcription factor Foxn1 in 1994, was preceded 30 years earlier by the spontaneous appearance of phenotypically (hairless) nude mice in an albino stock at the Virus Laboratory, Ruchill Hospital, Glasgow, (UK) [23,24]. The mice were characterized by a lack of visible hair and a lack of thymus, resulting in T-cell deficiency. Because of their immunodeficiency, nude mice rapidly became the ideal host for xenograft studies investigating predominantly human cancerous tumors. In 1994, Nehls et al. showed that a single-base-pair (G) deletion in exon 3 of the whn gene (the former name of Foxn1) causes a frameshift resulting in aberrant protein that lack the DNA-binding domain [25]. The results of the study provided strong evidence that the nude mouse phenotype is the result of the mutation in the Foxn1 (whn) gene. Further study revealed the presence of a nude phenotype due to a Foxn1 mutation in humans [26,27] and rats [25,28]. In mice, Foxn1 is located on chromosome 11, and in humans, it is located on chromosome 17. In humans and mice, the Foxn1 gene consists of eight coding exons with two alternative first exons: 1a and 1b. Interestingly, a study by Schropp et al., revealed tissue-specific alternative first exon usage [29]. While 1a-2 transcripts were detected in mouse thymus and skin, transcript 1b-2 was detected only in the skin. The authors concluded that these results indicate the presence of two separate promotors: one upstream of exon 1a and the second upstream of exon 1b [29].
The predicted protein product of the Foxn1 gene consists of 648 amino acids with 85% homology between humans and mice [29]. Tissue-specific Foxn1 expression is located in the epithelium of the thymus and the skin, and recessive nude alleles lead to abnormalities in cutaneous and thymic epithelial development (nude phenotype) [24,30,31]. The skin of nude mice contains the same number and normally cycling hair follicles as wild-type mice, but the defect in keratinization of the hair shaft makes them non-growing above the skin surface and causes impairment in the nail matrix [32]. In the thymus, Foxn1 is expressed in thymic epithelial cells (TECs), in which it is involved in thymus development and TEC differentiation [33]. The expression and functional study of Foxn1 in the thymus was meticulously conducted by several groups of investigators [33,34,35,36]. Foxn1 in the skin has attracted less attention; however, comprehensive experiments consistently show the importance of Foxn1 in skin physiology [37,38,39,40].
3. Foxn1 Regulates Epidermis and Hair Follicle Physiology
Brissette et al., were the first to investigate Foxn1 function in the skin [32,37,41]. A developmental study using transgenic mice in which a β-galactosidase reporter gene was placed under the control of the wild-type Foxn1 promoter revealed Foxn1 expression in epithelial cells of skin, thymus, nails, nasal passages, tongue, palate, and teeth [32]. Whole mounts of heterozygous (whn-lacZ fusion; whn+/−) embryos stained for β-galactosidase activity showed staining (Foxn1 activity) in the developing nasal region at day 13 of gestation, developing mouth, hair follicles of the eyebrows and lashes, and epidermis of the ear and tail at day 14.5. The entire epidermis, including developing hair follicles of the fur coat, expressed β-galactosidase at gestational days E15.5–16.5 [32]. In the epidermis of heterozygous whn-lacZ mice, β-galactosidase activity was predominantly found in the first suprabasal layer, with occasionally observed positive cells in the basal layer of keratinocytes [32]. Location of Foxn1 in the suprabasal layer that contains keratinocytes at the early stage of differentiation suggested Foxn1 involvement in the process of interfollicular keratinocyte differentiation. Hair follicular whn (Foxn1) expression is primarily associated with the anagen stage (the period of hair growth) when differentiating cells are continuously produced. During hair follicular regression (catagen), Foxn1 expression decreases, becoming undetectable at the telogen (resting) stage [42]. The location of Foxn1-positive cells in the interfollicular suprabasal layer of the epidermis and in hair follicles was further confirmed in Foxn1::eGFP mice (Figure 1), in which the enhanced green fluorescent protein transgene is driven by the Foxn1 regulatory sequence [43,44].
In a subsequent study, Prowse et al., used transgenic mice that express whn (Foxn1) from the promoter of involucrin, which in normal skin is expressed in postmitotic (differentiated) epidermal cells [41]. The results of the study suggest that Foxn1 promotes the differentiation of Foxn1-expressing cells and stimulates the proliferation of neighboring cells through paracrine mechanisms [41,45]. A study by Kur-Piotrowska et al., further confirmed the role of Foxn1 as a transcription factor that activates and stimulates the first stages of keratinocyte differentiation [46]. The overexpression of Foxn1 in cultured keratinocytes stimulated the increase in keratin10-positive cells (marker of differentiated keratinocytes located in the suprabasal layer of epidermis) at the expense of a decrease in the percentage of keratin14-positive cells (marker of proliferative keratinocytes—located in the basal layer of the epidermis) [46].
Recent data have identified Foxn1 as a factor required for skin developmental programs and skin maturation [47]. Detailed comparative analysis of transcriptomes from the skin of adult Foxn1+/+ and Foxn1−/− mice and the skin of mouse fetuses at embryonic day 14 (Foxn1−/−) and at embryonic day 18 (Foxn1+/+) revealed substantial similarities in the gene expression profiles between E14 and Foxn1−/− mice. Genes associated with differentiation and skin development were downregulated in the skin of E14 and Foxn1−/− mice, whereas those involved in tissue remodeling, cytoskeletal rearrangements, wound healing, and the immune response showed upregulation [47]. Among the downregulated genes, there were genes encoding transcription factors associated with the homeodomain: Hox genes (Hoxb3, Hoxb5, Hoxc6, and Hoxd13), extended Hox genes (Gbx2), paired domain (Pax1), paired-like domain (Phox2b and Pitx1), and six/sine homeobox (Six1 and Six2). These transcription factors are involved in embryonic development and have been shown to regulate stem cell differentiation [48]. These data suggest that the lack of Foxn1 (Foxn1−/−) maintains the skin of adult (nude) mice in an undeveloped state, which is a rare example of neoteny phenomenon [47]. Presumably, these features of Foxn1−/− (nude) mice permit regenerative skin wound healing, as was observed for mouse embryos at up to 16.5 days of development [49,50,51]. Among genes that are exclusively related to Foxn1−/− deficiency in adult skin, members of the Wnt, Bmp, and Notch families were significantly altered [47]. Genes of the Notch group (Notch2 and 4 receptors, Notch ligands: Dll1, Dll4, and Dlk1) and several genes in the bone morphogenetic protein (Bmp) family (Bmp2 and Bmp4, receptor Bmpr1b) in the skin of Foxn1−/− mice were downregulated. In contrast, most of the members of Wnt family genes, namely, the Wnt ligands Wisp2, Wnt10a, and Wnt11, the Wnt mediator β-catenin (Ctnnb1) and Wnt receptors frizzled 1 (Fzd1) and frizzled 6 (Fzd6), were upregulated in the skin of Foxn1−/− mice with concurrently downregulated Wnt pathway inhibitors, namely, dickkopf 1 (Dkk1), frizzled-related protein (Frzb), and secreted frizzled-related protein 2 (Sfrp2) [47].
The participation of Wnt signaling in the regulation of Foxn1 expression has been observed in the thymus [52], but little is known about epidermal Foxn1 regulation. Hu et al. showed that Wnt5a is a direct Notch signaling target in dermal papilla cells, with Foxn1 acting as a downstream target in keratinocytes [53]. Cai et al. demonstrated that Foxn1 and Msx2, which function downstream of Bmp4 signaling, are required for the maintenance of Notch1 expression during the development of hair follicles [54]. A study by Potter et al., initially based on phenotypic similarities between Foxn1−/− and Hoxc13−/− mice due to their nude appearance and nail development, defects points to HOXC13-dependent activation of Foxn1 in the control of hair follicles and nail differentiation [55]. However, regulators of Foxn1 activity, particularly in the interfollicular part of the epidermis, are largely unidentified.
Although Foxn1 function in the skin and hair follicles has been analysed in wild-type mice as well as consequences of the defects caused by Foxn1 deficiency in nude mice, the precise role of Foxn1 in skin physiology and its target genes are still unclear. It has been shown that Foxn1 regulates the expression of hair keratin genes, particularly mHa3 and mHa5 [42,56,57]. HeLa cells (human epithelial cells that do not express hair keratins) transfected with the Foxn1 gene showed strong induction of mHa3, mHb3, and mHb5 by Foxn1 stimulation [56]. However, transient activation of Foxn1 in keratinocytes followed by gene microarray analyses did not show any keratin gene regulation due to Foxn1 stimulation [39]. In contrast, proteomics analysis of primary keratinocytes obtained from nude mice (Foxn1−/−) and transduced with Ad-Foxn1 showed upregulation of type II keratin 5 and Krt6b protein due to Foxn1 expression [46]. The prominent upregulation of some keratin (Krt) genes (Krt23, -73, -82, -16, and -17) in the total skin and Krt6a and -6b in the epidermis of Foxn1−/− (nude) mice was also reported [46].
The stimulatory effect of Foxn1 on the first steps of the keratinocyte differentiation program and its suppressive effect on the late stages was examined in detail in primary keratinocyte cultures [58]. The study revealed that the transcriptional activity of Foxn1 can be mediated through indirect mechanisms. Protein kinase C (PKC) has been identified as a key target of Foxn1 during the keratinocyte differentiation program. Applying loss and gain of Foxn1 function methodology showed that inactivation of Foxn1 stimulates PKC protein levels, whereas its overexpression reduces PKC signaling [58]. The activation of Foxn1 in primary human keratinocytes [39] and primary nude mouse keratinocytes [46] caused an increase in phospholipase δ1 (PLCδ1, 1-phosphatidylinositol 4,5-bisphosphate phosphodiesterase delta-1) mRNA [39] and protein levels [46]. Stimulated by Foxn1, an increase in the level of PLCδ1, the key molecule in the phosphoinositide signaling pathway, further supports the role of Foxn1 in PKC [58] and AKT [39] signaling. Moreover, a study by Nakamura showed that PLCδ1 KO mice displayed a nude phenotype similar to Foxn1−/− mice and that the expression of PLCδ1 was induced in Foxn1-transfected cells, indicating that PLCδ1 is a molecule downstream of Foxn1 [59].
The broad range of Foxn1 functions in the epidermis is also delineated in its role in the pigmentation process. Foxn1 stimulates epithelial cells to release fibroblast growth factor 2 (Fgf2), a signal that is recognized by melanocytes for pigment transfer to Foxn1-positive cells [38,60].
The multiple effects of Foxn1 on epidermal physiology were also analysed through proteomic profiling of cultured keratinocytes transfected with Ad-Foxn1 [46]. A proteomics analysis identified proteins that can act as factors controlling the balance between cell proliferation, differentiation, and apoptosis in response to Foxn1. Interestingly, control (Ad-GFP transduced) keratinocytes were characterized by the presence of pro-survival proteins. 3-phosphoinositide-dependent protein kinase 1 (Pdpk1), the protein that acts upstream of the Akt (protein kinase B) pathway, and Hsp-90 and -27 proteins have been shown to be involved in pro-proliferative and pro-survival pathways, respectively. In contrast, some of the proteins detected in Foxn1-overexpressing keratinocytes are related to pro-apoptotic functions, such as prolyl 4-hydroxylase, beta polypeptide (P4hb), galectin-7 (Lgals7), and voltage-dependent anion-selective channel protein 1 (Vdac1). Furthermore, flow cytometry analysis revealed an increase in apoptotic cells in the Ad-Foxn1 population [46]. However, the most striking results of proteomic analysis point to Foxn1 as a factor regulating the levels of proteins (i.e., thioredoxin and prolyl 4-hydroxylase) involved in adaptation to normoxic/hypoxic conditions [46].
Interestingly, the comparison between the skin transcriptomes of Foxn1−/− and Foxn1+/+ mice revealed considerable differences in the genes associated with skin immuno- and proteolytic-defense [47]. As expected, compared to that of Foxn1+/+ mice, the skin of Foxn1−/− mice displayed downregulated transcripts of CD3 (T lymphocytes). In contrast, Foxn1−/− mouse skin showed upregulation of the transcripts of proteolytic molecules such as kallikreins and kallikrein-related peptidases, including Klk 6, Klk13, Klk7, Klk8, and Klk10, protease serine 27 (Prss27), which is involved in the skin barrier proteolytic cascade, and langerin (CD207, marker of dendritic cells), among others [47,61].
These data broaden the potential range of Foxn1 action in the skin but may also indicate the presence of a compensatory mechanism that already exists at the transcriptome level that provides a form of protection in nude mice as described by Miroslav Holub [62]. Considering that the major locations of Foxn1 are confined to the epithelium of the two immunocompetent organs, the thymus and the skin, the role of Foxn1 in the immunodefense guard in the skin has gained little interest [63].
Taken together, these investigations underline the multiple, both direct and indirect, effects of Foxn1 on the skin, particularly its epidermal layer.
4. Foxn1 as a Skin Wound Healing Regulator
The first premise that Foxn1 may regulate the skin wound healing process came from experiments performed on Foxn1−/− mice [64,65,66,67]. Foxn1 deficiency resulted in an unusual skin wound healing process in nude mice. While scar forming wound healing (reparative healing) is generally characteristic of mammals, nude mice appear to be remarkable in scar-less (regenerative) healing. Until that discovery, regenerative skin wound healing was detected exclusively in mammalian fetuses, although it was limited to the first two trimesters of intrauterine life [68]. Injury generated after the so-called transitional period, which takes place after the first two trimesters and before the third and last trimester of gestation, resulted in scar-forming skin healing observed in adults [51]. The analysis of post-injured skin of Foxn1−/− mice showed typical features of regenerative healing, including accelerated re-epithelialization process, absence of scars, high levels of Tgfβ3, the anti-scarring cytokine, low pro-scarring Tgfβ1 expression, high levels of hyaluronic acid, and expression of two matrix metalloproteinases (Mmps), Mmp9, and Mmp13, altered to favor the regenerative pathway, during the skin wound healing process [65,66]. Particularly, the differences in the pattern of Mmp9 expression during skin wound healing between Foxn1−/− and Foxn1+/+ mice has attracted some interest [65]. Low levels of Mmp9 expression in the epidermis of Foxn1−/− mice compared to high levels of Mmp9 expression in the total skin content have been reported [47]. Since low epidermal expression of Mmp9 coincided with epidermal Foxn1 deficiency in nude mice, it seemed conceivable that there is a direct interaction between them. So far, there are no data that may support the direct link between those two factors. However, it was shown that both Mmp9 and Foxn1 expression in keratinocytes is stimulated by hypoxic conditions [46].
All of the above features characterize the scar-free healing process detected in skin injuries of mammalian fetuses. Interestingly, the transition from scar-free to scar-forming healing during in utero life in mouse fetuses coincides with skin Foxn1 appearance, which takes place at embryonic day 16.5. It is conceivable that Foxn1 stimulates skin development and initiates the transition between scar-free and scar-forming skin wound healing during intrauterine life. This hypothesis is still under investigation [40].
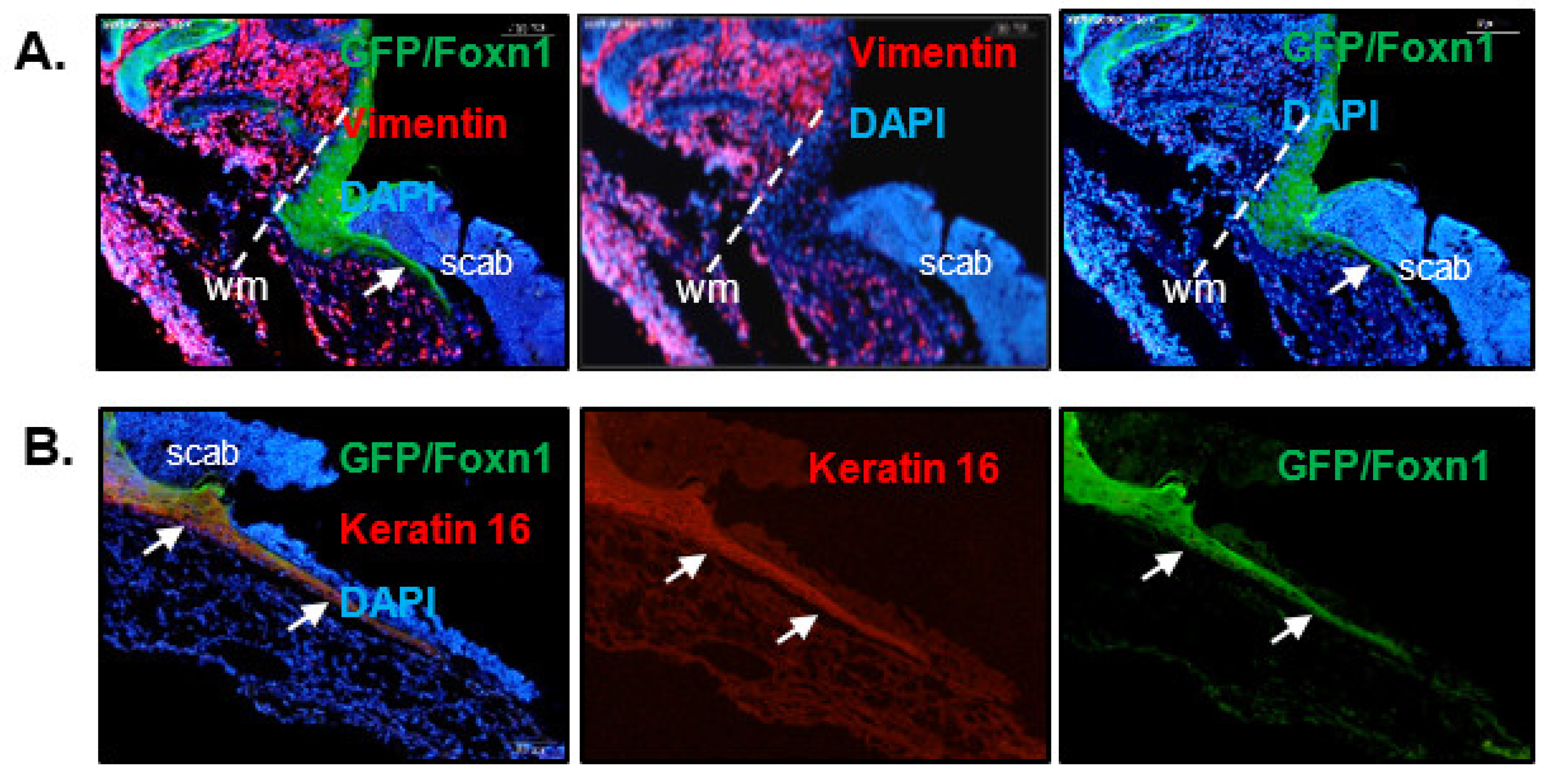
A study that aimed to analyze the role of Foxn1 in skin wound healing was performed on transgenic Foxn1::eGFP mice [43,44,46,69]. Post-wounded skin of Foxn1::eGFP mice showed the accumulation of cells bearing an eGFP (Foxn1) signal around the wound (wound margin) and in cells forming the leading epithelial tongue migrating underneath the scab. The entire extent and thickness of the migrating epithelium showed co-localization of Foxn1-eGFP with keratin 16 (a marker of activated, hyperproliferative epithelia; Figure 2) or with E-cadherin (a marker of epidermal cell characteristics; [43,44]).
However, the presence of Foxn1-positive cells was also observed in the dermal layer of post-wounded skin, the signal that co-localized with Snail1, a transcription factor involved in the epithelial-mesenchymal transition process (EMT; [70]) and Mmp9 (marker of the EMT) [43]. The participation of Foxn1-expressing cells in the EMT process, which leads to scar formation, was further confirmed with a flow cytometry assay, indicating an increase in N-cadherin (a marker of mesenchymal cell characteristics) and E-cadherin + N-cadherin-positive cells within the Foxn1-eGFP isolated cell population at post-wounded day 7 [44]. The presence of Foxn1-eGFP cells was also detected in post-wounded dermis (papillary dermis) during the remodeling phase of skin wound healing [44]. Foxn1-eGFP positive cells presented in the papillary dermis colocalized with α smooth muscle actin (αSMA) the myofibroblast marker [44]. Collectively, the data implicate Foxn1 as a transcription activator contributing to the activation of keratins (i.e., keratin16) required for epithelial resurfacing of post-wounded skin tissues and as a factor stimulating the EMT process, which leads to scar formation [43].
Little is known about how Foxn1 is controlled in the skin, including its regulation during the skin wound healing process. A study by Bukowska et al., suggested that, similar to its role in the thymus, the Wnt pathway is involved in Foxn1 regulation during skin wound healing [69]. In particular, β-catenin-dependent Wnt10a and β-catenin-independent Wnt11 signaling seem to regulate Foxn1 in the process of reparative (scar-forming) skin wound healing. Similar patterns of Foxn1, Wnt10a, Wnt11, and β-catenin mRNA expression during excisional wound healing in mice together with co-localization of their protein products to the epidermis and epidermal leading tongue indicate a possible interaction, although the precise mechanistic explanation awaits exploration [69].
The observed differences between Foxn1+/+ and Foxn1−/− mice in the process of skin wound healing suggest that Foxn1 acts beyond the epidermis, its tissue of origin. Indeed, a comparative analysis of dermal fibroblasts isolated from Foxn1+/+ and Foxn1−/− mice showed vast differences between them [65,71,72]. Dermal fibroblasts from Foxn1−/− mice showed features of neoteny. The population of Foxn1−/− DFs contained a high percentage of cells expressing stem cell markers CD117 and Oct3/4, which are low (CD117) or absent (Oct3/4) in Foxn1+/+ DFs [71]. Foxn1−/− DFs exhibited a higher cumulative number of population doublings (cumulative PD), increased adipogenic differentiation capacity, higher expression of Tgfβ3, and greater sensitivity to TGFβ3 stimulation compared to Foxn1+/+ DFs [72]. Moreover, dermal fibroblasts from Foxn1−/− mice produce higher levels of Mmp3, -9, and 13 and collagen III than Foxn1+/+ DFs [65,72]. All these features are common among sites prone to regenerative wound healing, namely, skin of mammalian fetuses and gingival tissues (scar-less healing of privileged tissues) [73,74]. Since strong evidence exists that epidermal Foxn1 plays a critical role in differentiation, developmental processes, and maturation, it is conceivable that a lack of Foxn1 maintains keratinocytes and DFs in the immature (fetal) regenerative period of development.
5. Foxn1 in Permeability Barrier Function, dWAT Physiology, and Resistance to Obesity
The epidermis serves as a barrier to pathogen entry and a permeability barrier protecting from water and heat loss; thus, providing thermoregulatory functions. All these functions are mediated by keratinocytes and lipid-containing extracellular matrix. Lipids in the epidermis consist of ceramides (50% by weight), cholesterol (27%), and fatty acids (10%) that are synthesized by keratinocytes but in part come from circulation (essential fatty acids) [75]. A detailed comparative analysis of the lipidome of skin biopsies from Foxn1−/− and Foxn1+/+ mice showed that mutation in the Foxn1 gene modifies the lipid profile [76]. Non-activity of Foxn1 in nude mouse skin leads to an increase in fatty acids, cholesterol sulphate, phospholipids, and sphingolipids with a concurrent decrease in triacylglycerols and diacylglycerols. The detected differences may explain the high rate (72%) of newborn inv-whn transgenic mouse mortality due to the inability to retain water [41]. Moreover, downregulation of PLCδ1, which is the target gene of Foxn1, impairs the barrier function of the epidermis due to dysregulation of Ca2+ and p38 mitogen-activated protein kinase (MAPK) signaling [77].
The modification of the skin lipidome profile in Foxn1−/− mice suggests dysregulation of thermoregulatory skin function, which at least partially may explain the resistance of Foxn1−/− mice to diet-induced obesity. A study by Stemmer et al. showed that even mild cold stress (ambient housing temperature) increased energy expenditure, protecting Foxn1−/− mice from high-fat diet-induced obesity, compared with obesity-prone Foxn1+/+ mice [78]. Similar observations were made by Hirata et al., examining PLCδ1 KO resistance to diet-induced obesity [79]. Both Foxn1−/− and PLCδ1 KO mice are hairless, which causes changes in thermoregulatory mechanisms to maintain body temperature [59]. It requires an increase in oxygen consumption and heat production that consequently may affect their resistance to diet-induced obesity. Although a central (hypothalamic) role of Foxn1 in controlling food intake should also be considered [78].
However, a study performed on Foxn1+/+ and Foxn1+/− mice, which have two (Foxn1+/+) or one (Foxn1+/−) active allele of Foxn1 but both have regular fur coats, revealed substantial differences between them in susceptibility to diet-induced obesity [80]. Foxn1+/− mice fed a high-fat diet for a period of eight weeks gained significantly less body weight than Foxn1+/+ mice regardless of age or sex. Analysis of skin isolated from Foxn1+/+ and Foxn1+/− mice showed abrogated expression of master regulators of adipogenesis (Pparγ, Fabp4, leptin, and Mest) in the skin of Foxn1+/− mice [80]. Moreover, the differences in dWAT adipogenesis between Foxn1+/+ and Foxn1+/− mice were detected particularly during the skin wound healing process, supporting the important role of skin adipocytes in the restoration of skin integrity [80,81]. Currently, it is known that the epidermal Wnt/β-catenin signaling pathway is a key initiator of the cascade that includes the Bmp (Bmp2/Bmp6) and insulin (Igf2) signaling pathways and stimulates the process of adipogenesis in the skin [82]. Recent data showed that gradual inactivation of Foxn1 in the skin of Foxn1+/+, Foxn1+/−, and Foxn1−/− mice is accompanied by a gradual decrease in Bmp2, Igf2, and Mest mRNA expression levels [80]. Whether Foxn1 is a mediator in the adipogenic cascade associated with Wnt/β-catenin and Bmp2/Igf2; thereby, affecting skin homeostasis, wound healing, and susceptibility to diet-induced obesity, is currently under investigation.
6. Conclusions
Foxn1 regulates skin development, renewal, and response to cutaneous insults. Expressed in the epidermis, Foxn1 contributes to the control of interfollicular and hair follicular epidermal renewal and pigmentation processes. In the dermal layer of the skin it affects dermal fibroblasts and dWAT biology. During skin wound healing, Foxn1 plays a role in re-epithelization and EMT processes; thereby, contributing to scar formation. Furthermore, a role for Foxn1 in the skin immunodefense is highly conceivable. Considering that Foxn1 expression is limited to the skin and thymus, novel targeted therapeutics regulating skin ageing and wound healing management will likely prove to be both beneficial and safe.
Funding
This research was funded by the National Science Centre, Poland (Grant OPUS 14 No. 2017/27/B/NZ5/02610).
Conflicts of Interest
The author declares no conflict of interest.
References
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the environment: Regulation of local and global homeostasis by the skin’s neuroendocrine system. Adv. Anat. Embryol. Cell Biol. 2012, 212, 1–115. [Google Scholar]
- Nejati, R.; Kovacic, D.; Slominski, A. Neuro-immune-endocrine functions of the skin: An overview. Expert Rev. Dermatol. 2013, 8, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, A.; Cho, R.J.; Luu, A.; Harirchian, P.; Lee, J.; Cheng, J.B.; Song, J.S. Single-Cell Transcriptomics Reveals Spatial and Temporal Turnover of Keratinocyte Differentiation Regulators. Front. Genet. 2019, 10, 775. [Google Scholar] [CrossRef] [PubMed]
- Clark, R. The Molecular and Cellular Biology of Wound Repair; Plenum Press: New York, NY, USA, 1996. [Google Scholar]
- Driskell, R.R.; Lichtenberger, B.M.; Hoste, E.; Kretzschmar, K.; Simons, B.D.; Charalambous, M.; Ferron, S.R.; Herault, Y.; Pavlovic, G.; Ferguson-Smith, A.C.; et al. Distinct fibroblast lineages determine dermal architecture in skin development and repair. Nature 2013, 504, 277–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognoni, E.; Watt, F.M. Skin Cell Heterogeneity in Development, Wound Healing, and Cancer. Trends Cell Biol. 2018, 28, 709–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojciechowicz, K.; Gledhill, K.; Ambler, C.A.; Manning, C.B.; Jahoda, C.A. Development of the mouse dermal adipose layer occurs independently of subcutaneous adipose tissue and is marked by restricted early expression of FABP4. PLoS ONE 2013, 8, e59811. [Google Scholar] [CrossRef]
- Rivera-Gonzalez, G.; Shook, B.; Horsley, V. Adipocytes in skin health and disease. Cold Spring Harb. Perspect. Med. 2014, 4, a015271. [Google Scholar] [CrossRef]
- Kruglikov, I.L.; Scherer, P.E. Dermal Adipocytes: From Irrelevance to Metabolic Targets? Trends Endocrinol. Metab. 2016, 27, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Driskell, R.R.; Jahoda, C.A.; Chuong, C.M.; Watt, F.M.; Horsley, V. Defining dermal adipose tissue. Exp. Dermatol. 2014, 23, 629–631. [Google Scholar] [CrossRef] [Green Version]
- Bellavia, G.; Fasanaro, P.; Melchionna, R.; Capogrossi, M.C.; Napolitano, M. Transcriptional control of skin reepithelialization. J. Dermatol. Sci. 2014, 73, 3–9. [Google Scholar] [CrossRef]
- Dai, X.; Segre, J.A. Transcriptional control of epidermal specification and differentiation. Curr. Opin. Genet. Dev. 2004, 14, 485–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, C.A.; Kretzschmar, K.; Watt, F.M. Reprogramming adult dermis to a neonatal state through epidermal activation of beta-catenin. Development 2011, 138, 5189–5199. [Google Scholar] [CrossRef] [Green Version]
- Lichtenberger, B.M.; Mastrogiannaki, M.; Watt, F.M. Epidermal beta-catenin activation remodels the dermis via paracrine signalling to distinct fibroblast lineages. Nat. Commun. 2016, 7, 10537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallant-Behm, C.L.; Du, P.; Lin, S.M.; Marucha, P.T.; DiPietro, L.A.; Mustoe, T.A. Epithelial regulation of mesenchymal tissue behavior. J. Investig. Dermatol. 2011, 131, 892–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schafer, M.; Werner, S. Transcriptional control of wound repair. Annu. Rev. Cell Dev. Biol. 2007, 23, 69–92. [Google Scholar] [CrossRef] [PubMed]
- Haensel, D.; Sun, P.; MacLean, A.L.; Ma, X.; Zhou, Y.; Stemmler, M.P.; Brabletz, S.; Berx, G.; Plikus, M.V.; Nie, Q.; et al. An Ovol2-Zeb1 transcriptional circuit regulates epithelial directional migration and proliferation. EMBO Rep. 2019, 20, e46273. [Google Scholar] [CrossRef]
- Rajendran, N.K.; Kumar, S.S.D.; Houreld, N.N.; Abrahamse, H. Understanding the perspectives of forkhead transcription factors in delayed wound healing. J. Cell Commun. Signal. 2019, 13, 151–162. [Google Scholar] [CrossRef]
- Boudra, R.; Ramsey, M.R. Understanding Transcriptional Networks Regulating Initiation of Cutaneous Wound Healing. Yale J. Biol. Med. 2020, 93, 161–173. [Google Scholar]
- Hannenhalli, S.; Kaestner, K.H. The evolution of Fox genes and their role in development and disease. Nat. Rev. Genet. 2009, 10, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, E.; Knochel, W. Five years on the wings of fork head. Mech. Dev. 1996, 57, 3–20. [Google Scholar] [CrossRef]
- Golson, M.L.; Kaestner, K.H. Fox transcription factors: From development to disease. Development 2016, 143, 4558–4570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flanagan, S.P. ‘Nude’, a new hairless gene with pleiotropic effects in the mouse. Genet. Res. 1966, 8, 295–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mecklenburg, L.; Tychsen, B.; Paus, R. Learning from nudity: Lessons from the nude phenotype. Exp. Dermatol. 2005, 14, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Nehls, M.; Pfeifer, D.; Schorpp, M.; Hedrich, H.; Boehm, T. New member of the winged-helix protein family disrupted in mouse and rat nude mutations. Nature 1994, 372, 103–107. [Google Scholar] [CrossRef]
- Pignata, C.; Fiore, M.; Guzzetta, V.; Castaldo, A.; Sebastio, G.; Porta, F.; Guarino, A. Congenital Alopecia and nail dystrophy associated with severe functional T-cell immunodeficiency in two sibs. Am. J. Med. Genet. 1996, 65, 167–170. [Google Scholar] [CrossRef]
- Frank, J.; Pignata, C.; Panteleyev, A.A.; Prowse, D.M.; Baden, H.; Weiner, L.; Gaetaniello, L.; Ahmad, W.; Pozzi, N.; Cserhalmi-Friedman, P.B.; et al. Exposing the human nude phenotype. Nature 1999, 398, 473–474. [Google Scholar] [CrossRef] [Green Version]
- Segre, J.A.; Nemhauser, J.L.; Taylor, B.A.; Nadeau, J.H.; Lander, E.S. Positional cloning of the nude locus: Genetic, physical, and transcription maps of the region and mutations in the mouse and rat. Genomics 1995, 28, 549–559. [Google Scholar] [CrossRef]
- Schorpp, M.; Hofmann, M.; Dear, T.N.; Boehm, T. Characterization of mouse and human nude genes. Immunogenetics 1997, 46, 509–515. [Google Scholar] [CrossRef]
- Schlake, T.; Schorpp, M.; Nehls, M.; Boehm, T. The nude gene encodes a sequence-specific DNA binding protein with homologs in organisms that lack an anticipatory immune system. Proc. Natl. Acad. Sci. USA 1997, 94, 3842–3847. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Burnley, P.; Coder, B.; Su, D.M. Insights on FoxN1 biological significance and usages of the “nude” mouse in studies of T-lymphopoiesis. Int. J. Biol. Sci. 2012, 8, 1156–1167. [Google Scholar] [CrossRef]
- Lee, D.; Prowse, D.M.; Brissette, J.L. Association between mouse nude gene expression and the initiation of epithelial terminal differentiation. Dev. Biol. 1999, 208, 362–374. [Google Scholar] [CrossRef]
- Chen, L.; Xiao, S.; Manley, N.R. Foxn1 is required to maintain the postnatal thymic microenvironment in a dosage-sensitive manner. Blood 2009, 113, 567–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbeaux, T.; Hess, I.; Swann, J.B.; Kanzler, B.; Haas-Assenbaum, A.; Boehm, T. Thymopoiesis in mice depends on a Foxn1-positive thymic epithelial cell lineage. Proc. Natl. Acad. Sci. USA 2010, 107, 16613–16618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bredenkamp, N.; Nowell, C.S.; Blackburn, C.C. Regeneration of the aged thymus by a single transcription factor. Development 2014, 141, 1627–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, M.D.; Csomos, K.; Dias, L.P.; Prodan, Z.; Szerafin, T.; Savino, W.; Takacs, L. Decline of FOXN1 gene expression in human thymus correlates with age: Possible epigenetic regulation. Immun. Ageing 2015, 12, 18. [Google Scholar] [CrossRef] [Green Version]
- Brissette, J.L.; Li, J.; Kamimura, J.; Lee, D.; Dotto, G.P. The product of the mouse nude locus, Whn, regulates the balance between epithelial cell growth and differentiation. Genes Dev. 1996, 10, 2212–2221. [Google Scholar] [CrossRef] [Green Version]
- Weiner, L.; Han, R.; Scicchitano, B.M.; Li, J.; Hasegawa, K.; Grossi, M.; Lee, D.; Brissette, J.L. Dedicated epithelial recipient cells determine pigmentation patterns. Cell 2007, 130, 932–942. [Google Scholar] [CrossRef]
- Janes, S.M.; Ofstad, T.A.; Campbell, D.H.; Watt, F.M.; Prowse, D.M. Transient activation of FOXN1 in keratinocytes induces a transcriptional programme that promotes terminal differentiation: Contrasting roles of FOXN1 and Akt. J. Cell Sci. 2004, 117, 4157–4168. [Google Scholar] [CrossRef] [Green Version]
- Bukowska, J.; Kopcewicz, M.; Walendzik, K.; Gawronska-Kozak, B. Foxn1 in Skin Development, Homeostasis and Wound Healing. Int. J. Mol. Sci. 2018, 19, 1956. [Google Scholar] [CrossRef] [Green Version]
- Prowse, D.M.; Lee, D.; Weiner, L.; Jiang, N.; Magro, C.M.; Baden, H.P.; Brissette, J.L. Ectopic expression of the nude gene induces hyperproliferation and defects in differentiation: Implications for the self-renewal of cutaneous epithelia. Dev. Biol. 1999, 212, 54–67. [Google Scholar] [CrossRef] [Green Version]
- Meier, N.; Dear, T.N.; Boehm, T. Whn and mHa3 are components of the genetic hierarchy controlling hair follicle differentiation. Mech. Dev. 1999, 89, 215–221. [Google Scholar] [CrossRef]
- Gawronska-Kozak, B.; Grabowska, A.; Kur-Piotrowska, A.; Kopcewicz, M. Foxn1 Transcription Factor Regulates Wound Healing of Skin through Promoting Epithelial-Mesenchymal Transition. PLoS ONE 2016, 11, e0150635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopcewicz, M.M.; Kur-Piotrowska, A.; Bukowska, J.; Gimble, J.M.; Gawronska-Kozak, B. Foxn1 and Mmp-9 expression in intact skin and during excisional wound repair in young, adult, and old C57Bl/6 mice. Wound Repair Regen. 2017, 25, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Baxter, R.M.; Brissette, J.L. Role of the nude gene in epithelial terminal differentiation. J. Investig. Dermatol. 2002, 118, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Kur-Piotrowska, A.; Bukowska, J.; Kopcewicz, M.M.; Dietrich, M.; Nynca, J.; Slowinska, M.; Gawronska-Kozak, B. Foxn1 expression in keratinocytes is stimulated by hypoxia: Further evidence of its role in skin wound healing. Sci. Rep. 2018, 8, 5425. [Google Scholar] [CrossRef] [Green Version]
- Kur-Piotrowska, A.; Kopcewicz, M.; Kozak, L.P.; Sachadyn, P.; Grabowska, A.; Gawronska-Kozak, B. Neotenic phenomenon in gene expression in the skin of Foxn1-deficient (nude) mice—A projection for regenerative skin wound healing. BMC Genom. 2017, 18, 56. [Google Scholar] [CrossRef] [Green Version]
- Boyer, L.A.; Plath, K.; Zeitlinger, J.; Brambrink, T.; Medeiros, L.A.; Lee, T.I.; Levine, S.S.; Wernig, M.; Tajonar, A.; Ray, M.K.; et al. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature 2006, 441, 349–353. [Google Scholar] [CrossRef]
- Ihara, S.; Motobayashi, Y.; Nagao, E.; Kistler, A. Ontogenetic transition of wound healing pattern in rat skin occurring at the fetal stage. Development 1990, 110, 671–680. [Google Scholar]
- Longaker, M.T.; Whitby, D.J.; Adzick, N.S.; Crombleholme, T.M.; Langer, J.C.; Duncan, B.W.; Bradley, S.M.; Stern, R.; Ferguson, M.W.; Harrison, M.R. Studies in fetal wound healing, VI. Second and early third trimester fetal wounds demonstrate rapid collagen deposition without scar formation. J. Pediatr. Surg. 1990, 25, 63–68, Discussion 68–69. [Google Scholar] [CrossRef]
- Lorenz, H.P.; Adzick, N.S. Scarless skin wound repair in the fetus. West. J. Med. 1993, 159, 350–355. [Google Scholar]
- Balciunaite, G.; Keller, M.P.; Balciunaite, E.; Piali, L.; Zuklys, S.; Mathieu, Y.D.; Gill, J.; Boyd, R.; Sussman, D.J.; Hollander, G.A. Wnt glycoproteins regulate the expression of FoxN1, the gene defective in nude mice. Nat. Immunol. 2002, 3, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Lefort, K.; Qiu, W.; Nguyen, B.C.; Rajaram, R.D.; Castillo, E.; He, F.; Chen, Y.; Angel, P.; Brisken, C.; et al. Control of hair follicle cell fate by underlying mesenchyme through a CSL-Wnt5a-FoxN1 regulatory axis. Genes Dev. 2010, 24, 1519–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, J.; Lee, J.; Kopan, R.; Ma, L. Genetic interplays between Msx2 and Foxn1 are required for Notch1 expression and hair shaft differentiation. Dev. Biol. 2009, 326, 420–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, C.S.; Pruett, N.D.; Kern, M.J.; Baybo, M.A.; Godwin, A.R.; Potter, K.A.; Peterson, R.L.; Sundberg, J.P.; Awgulewitsch, A. The nude mutant gene Foxn1 is a HOXC13 regulatory target during hair follicle and nail differentiation. J. Investig. Dermatol. 2011, 131, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Schlake, T.; Schorpp, M.; Maul-Pavicic, A.; Malashenko, A.M.; Boehm, T. Forkhead/winged-helix transcription factor Whn regulates hair keratin gene expression: Molecular analysis of the nude skin phenotype. Dev. Dyn. 2000, 217, 368–376. [Google Scholar] [CrossRef]
- Schorpp, M.; Schlake, T.; Kreamalmeyer, D.; Allen, P.M.; Boehm, T. Genetically separable determinants of hair keratin gene expression. Dev. Dyn. 2000, 218, 537–543. [Google Scholar] [CrossRef]
- Li, J.; Baxter, R.M.; Weiner, L.; Goetinck, P.F.; Calautti, E.; Brissette, J.L. Foxn1 promotes keratinocyte differentiation by regulating the activity of protein kinase C. Differentiation 2007, 75, 694–701. [Google Scholar] [CrossRef]
- Nakamura, Y.; Ichinohe, M.; Hirata, M.; Matsuura, H.; Fujiwara, T.; Igarashi, T.; Nakahara, M.; Yamaguchi, H.; Yasugi, S.; Takenawa, T.; et al. Phospholipase C-delta1 is an essential molecule downstream of Foxn1, the gene responsible for the nude mutation, in normal hair development. FASEB J. 2008, 22, 841–849. [Google Scholar] [CrossRef]
- Weiner, L.; Fu, W.; Chirico, W.J.; Brissette, J.L. Skin as a living coloring book: How epithelial cells create patterns of pigmentation. Pigment Cell Melanoma Res. 2014, 27, 1014–1031. [Google Scholar] [CrossRef] [Green Version]
- Eissa, A.; Amodeo, V.; Smith, C.R.; Diamandis, E.P. Kallikrein-related peptidase-8 (KLK8) is an active serine protease in human epidermis and sweat and is involved in a skin barrier proteolytic cascade. J. Biol. Chem. 2011, 286, 687–706. [Google Scholar] [CrossRef] [Green Version]
- Holub, M. Immunology of Nude Mice; CRC Press, Inc.: Boca Raton, FL, USA, 1989; p. 173. [Google Scholar]
- Yang, J.; Hu, S.; Zhao, L.; Kaplan, D.H.; Perdew, G.H.; Xiong, N. Selective programming of CCR10(+) innate lymphoid cells in skin-draining lymph nodes for cutaneous homeostatic regulation. Nat. Immunol. 2016, 17, 48–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gawronska-Kozak, B. Regeneration in the ears of immunodeficient mice: Identification and lineage analysis of mesenchymal stem cells. Tissue Eng. 2004, 10, 1251–1265. [Google Scholar] [CrossRef] [PubMed]
- Gawronska-Kozak, B. Scarless skin wound healing in FOXN1 deficient (nude) mice is associated with distinctive matrix metalloproteinase expression. Matrix Biol. 2011, 30, 290–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gawronska-Kozak, B.; Bogacki, M.; Rim, J.S.; Monroe, W.T.; Manuel, J.A. Scarless skin repair in immunodeficient mice. Wound Repair Regen. 2006, 14, 265–276. [Google Scholar] [CrossRef]
- Gawronska-Kozak, B.; Grabowska, A.; Kopcewicz, M.; Kur, A. Animal models of skin regeneration. Reprod. Biol. 2014, 14, 61–67. [Google Scholar] [CrossRef]
- Rowlatt, U. Intrauterine wound healing in a 20 week human fetus. Virchows Arch. A Pathol. Anat. Histol. 1979, 381, 353–361. [Google Scholar] [CrossRef]
- Bukowska, J.; Walendzik, K.; Kopcewicz, M.; Cierniak, P.; Gawronska-Kozak, B. Wnt signaling and the transcription factor Foxn1 contribute to cutaneous wound repair in mice. Connect. Tissue Res. 2019, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Gawronska-Kozak, B.; Kirk-Ballard, H. Cyclosporin A reduces matrix metalloproteinases and collagen expression in dermal fibroblasts from regenerative FOXN1 deficient (nude) mice. Fibrogenesis Tissue Repair 2013, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Bukowska, J.; Kopcewicz, M.; Kur-Piotrowska, A.; Szostek-Mioduchowska, A.Z.; Walendzik, K.; Gawronska-Kozak, B. Effect of TGFbeta1, TGFbeta3 and keratinocyte conditioned media on functional characteristics of dermal fibroblasts derived from reparative (Balb/c) and regenerative (Foxn1 deficient; nude) mouse models. Cell Tissue Res. 2018, 374, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.S.; Borrelli, M.R.; Hong, W.X.; Malhotra, S.; Cheung, A.T.M.; Ransom, R.C.; Rennert, R.C.; Morrison, S.D.; Lorenz, H.P.; Longaker, M.T. Embryonic skin development and repair. Organogenesis 2018, 14, 46–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szpaderska, A.M.; Zuckerman, J.D.; DiPietro, L.A. Differential injury responses in oral mucosal and cutaneous wounds. J. Dent. Res. 2003, 82, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Wertz, P.W. Lipids and the Permeability and Antimicrobial Barriers of the Skin. J. Lipids 2018, 2018, 5954034. [Google Scholar] [CrossRef] [Green Version]
- Lanzini, J.; Dargere, D.; Regazzetti, A.; Tebani, A.; Laprevote, O.; Auzeil, N. Changing in lipid profile induced by the mutation of Foxn1 gene: A lipidomic analysis of Nude mice skin. Biochimie 2015, 118, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Kanemaru, K.; Nakamura, Y.; Totoki, K.; Fukuyama, T.; Shoji, M.; Kaneko, H.; Shiratori, K.; Yoneda, A.; Inoue, T.; Iwakura, Y.; et al. Phospholipase Cdelta1 regulates p38 MAPK activity and skin barrier integrity. Cell Death Differ. 2017, 24, 1079–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stemmer, K.; Kotzbeck, P.; Zani, F.; Bauer, M.; Neff, C.; Muller, T.D.; Pfluger, P.T.; Seeley, R.J.; Divanovic, S. Thermoneutral housing is a critical factor for immune function and diet-induced obesity in C57BL/6 nude mice. Int. J. Obes. 2015, 39, 791–797. [Google Scholar] [CrossRef] [Green Version]
- Hirata, M.; Suzuki, M.; Ishii, R.; Satow, R.; Uchida, T.; Kitazumi, T.; Sasaki, T.; Kitamura, T.; Yamaguchi, H.; Nakamura, Y.; et al. Genetic defect in phospholipase Cdelta1 protects mice from obesity by regulating thermogenesis and adipogenesis. Diabetes 2011, 60, 1926–1937. [Google Scholar] [CrossRef] [Green Version]
- Walendzik, K.; Kopcewicz, M.; Bukowska, J.; Panasiewicz, G.; Szafranska, B.; Gawronska-Kozak, B. The Transcription Factor FOXN1 Regulates Skin Adipogenesis and Affects Susceptibility to Diet-Induced Obesity. J. Investig. Dermatol. 2020, 140, 1166–1175. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, B.A.; Horsley, V. Intradermal adipocytes mediate fibroblast recruitment during skin wound healing. Development 2013, 140, 1517–1527. [Google Scholar] [CrossRef] [Green Version]
- Donati, G.; Proserpio, V.; Lichtenberger, B.M.; Natsuga, K.; Sinclair, R.; Fujiwara, H.; Watt, F.M. Epidermal Wnt/beta-catenin signaling regulates adipocyte differentiation via secretion of adipogenic factors. Proc. Natl. Acad. Sci. USA 2014, 111, E1501–E1509. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Foxn1 expression in the skin of Foxn1::eGFP mice. hf-hair follicles, arrows indicate Foxn1 positivity in the epidermis.
Figure 1.
Foxn1 expression in the skin of Foxn1::eGFP mice. hf-hair follicles, arrows indicate Foxn1 positivity in the epidermis.

Figure 2.
Foxn1 expression in injured skin of Foxn1::eGFP mice at post-wounded day 2 (A) and day 3 (B). wm—wound margin, arrows indicate leading epithelial tongue.
Figure 2.
Foxn1 expression in injured skin of Foxn1::eGFP mice at post-wounded day 2 (A) and day 3 (B). wm—wound margin, arrows indicate leading epithelial tongue.

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gawronska-Kozak, B. Foxn1 Control of Skin Function. Appl. Sci. 2020, 10, 5685. https://0-doi-org.brum.beds.ac.uk/10.3390/app10165685
AMA Style
Gawronska-Kozak B. Foxn1 Control of Skin Function. Applied Sciences. 2020; 10(16):5685. https://0-doi-org.brum.beds.ac.uk/10.3390/app10165685
Chicago/Turabian StyleGawronska-Kozak, Barbara. 2020. "Foxn1 Control of Skin Function" Applied Sciences 10, no. 16: 5685. https://0-doi-org.brum.beds.ac.uk/10.3390/app10165685
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.