Reduction of NF-κB Signals in Platelets and Prolongation of Platelet Plug Formation against High Shear Flow in Whole Blood on Human Subject by Columbianadin

 , , , and
, , , and 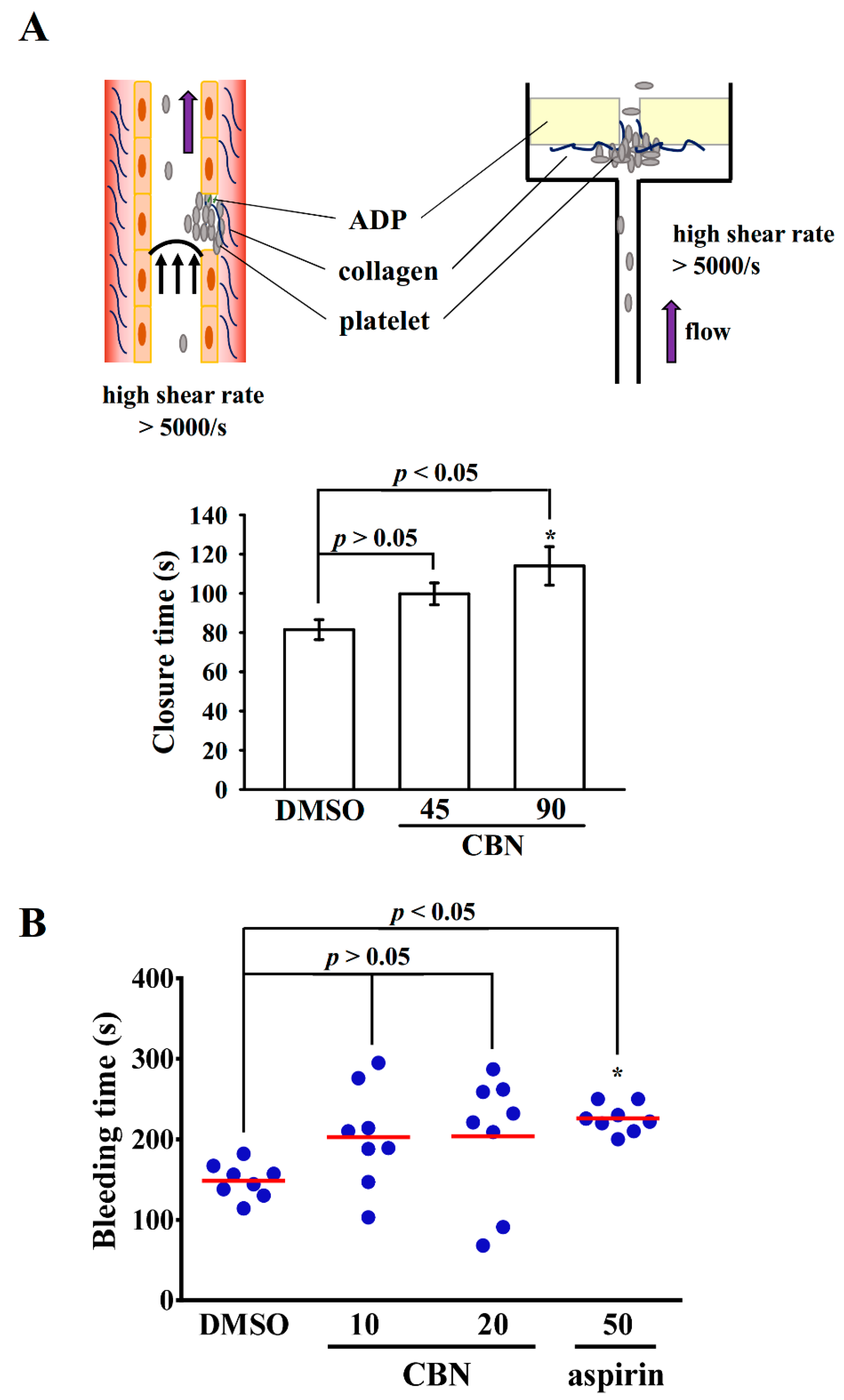{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Platelet Function Analysis in Human Whole Blood
2.3. Tail Bleeding Time
2.4. Platelet Aggregation
2.5. Immunoblotting Study
2.6. Measurement of Hydroxyl Radicals
2.7. Data Analysis
3. Results
3.1. Effect of CBN in Platelet Plug Formation in Human Whole Blood and Tail Bleeding in Experimental Mice
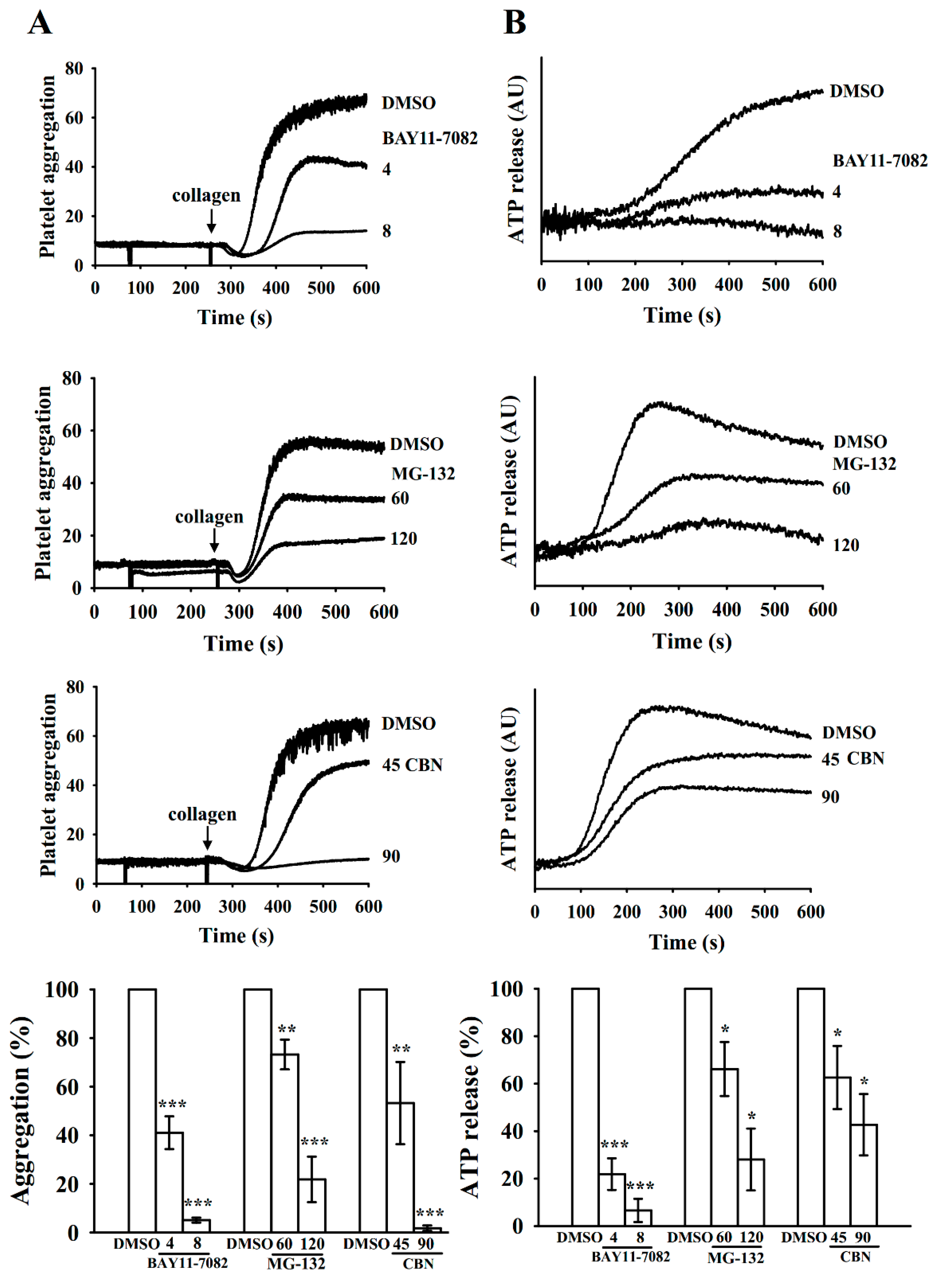
3.2. Inhibition of Platelet Aggregation and ATP-Release by BAY11-7082, MG-132, and CBN in Collagen-Stimulated Platelets
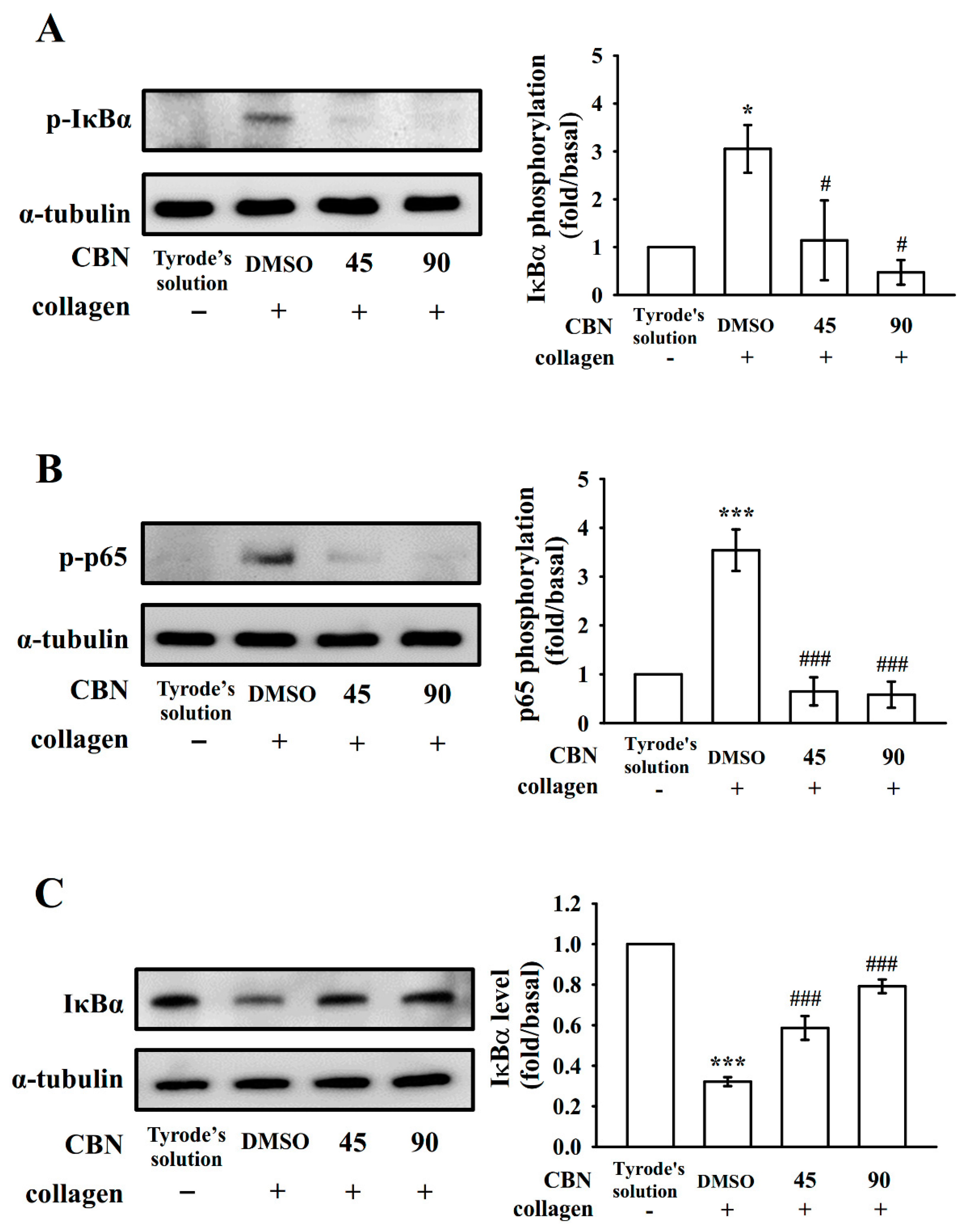
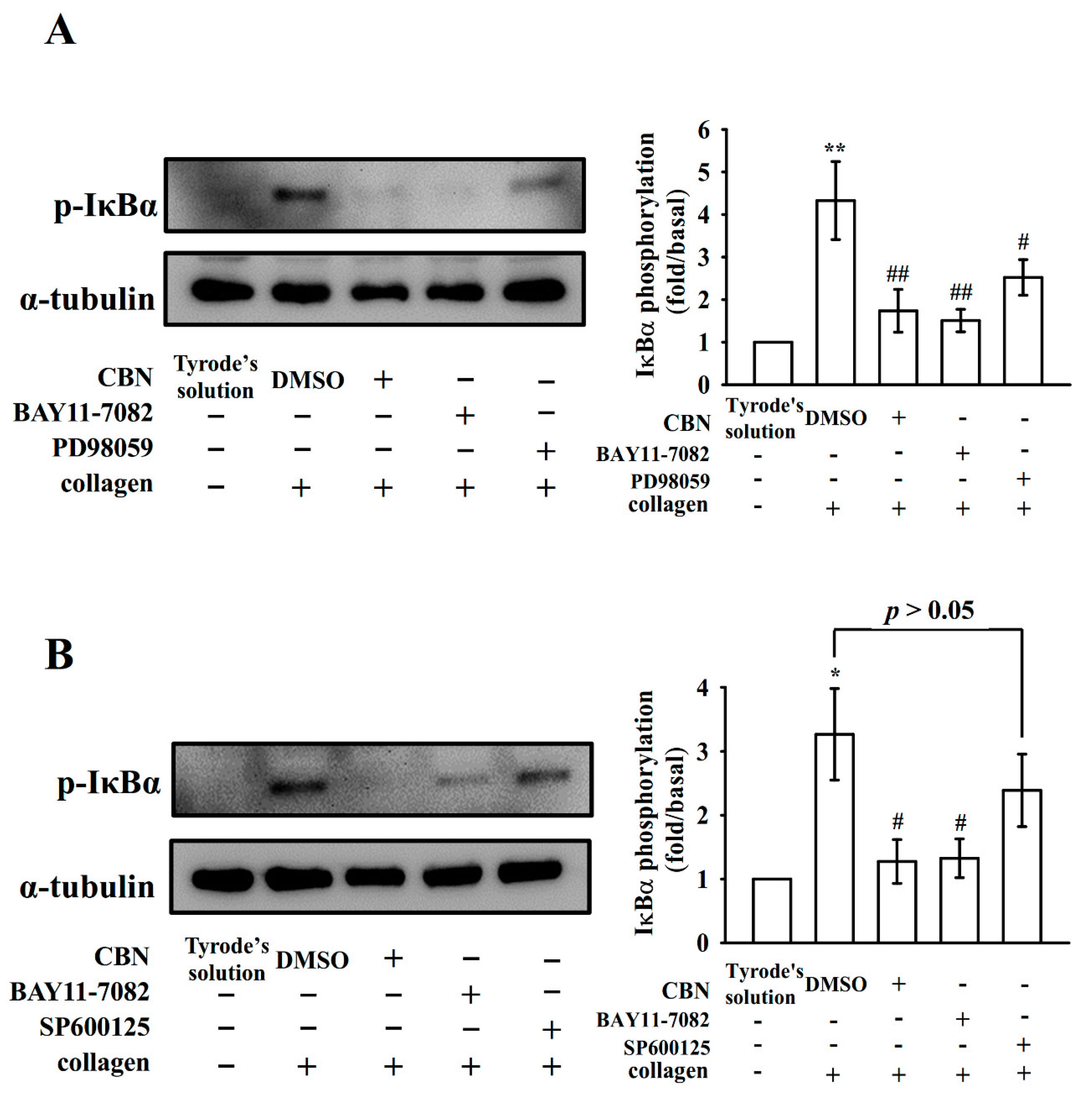
3.3. Reduction of NF-κB Signals by CBN in Platelets
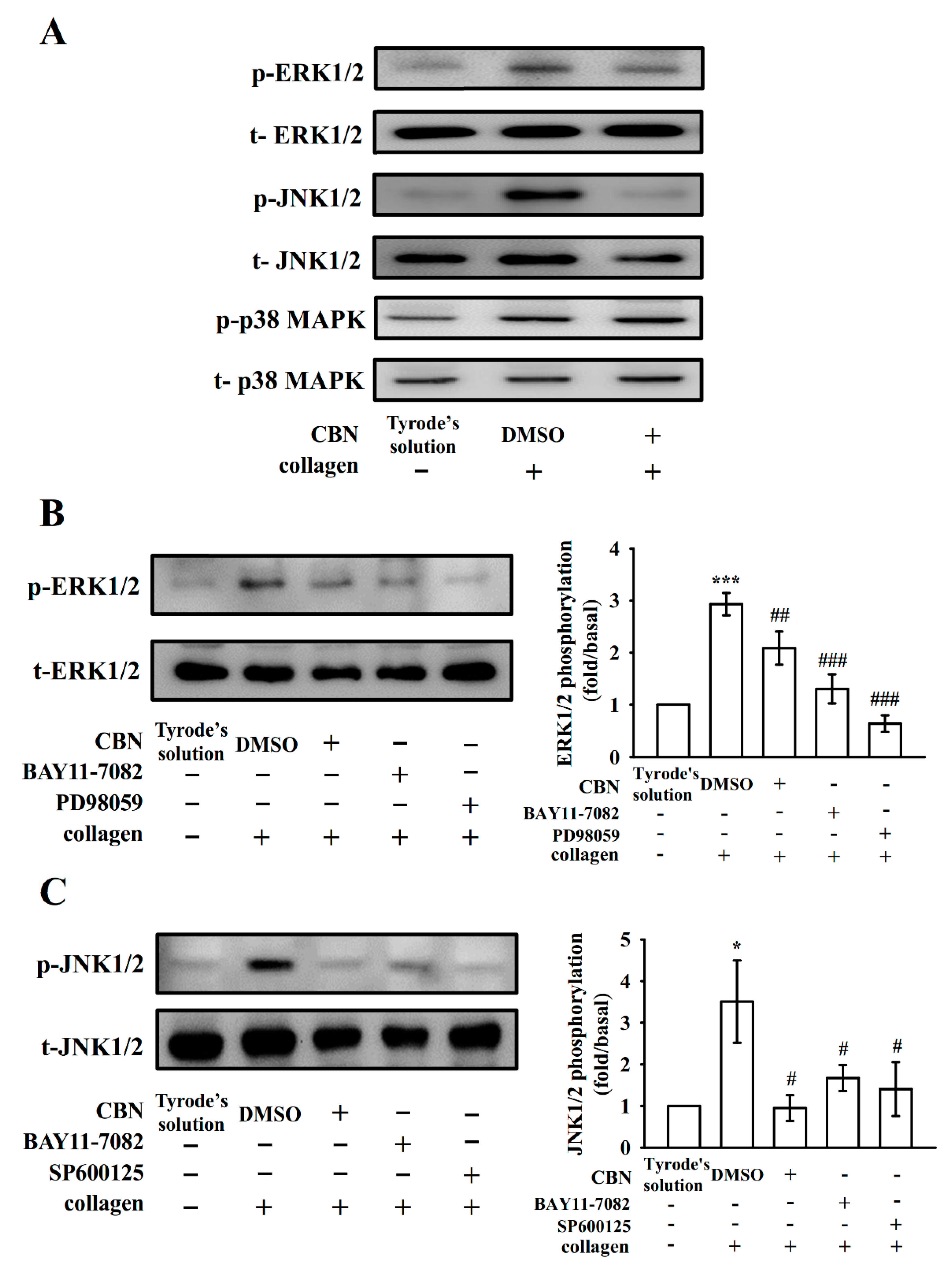
3.4. Connection Between NF-κB Signaling and MAPK Phosphorylation
3.5. Effect of CBN and BAY11-7082 on HO● Formation
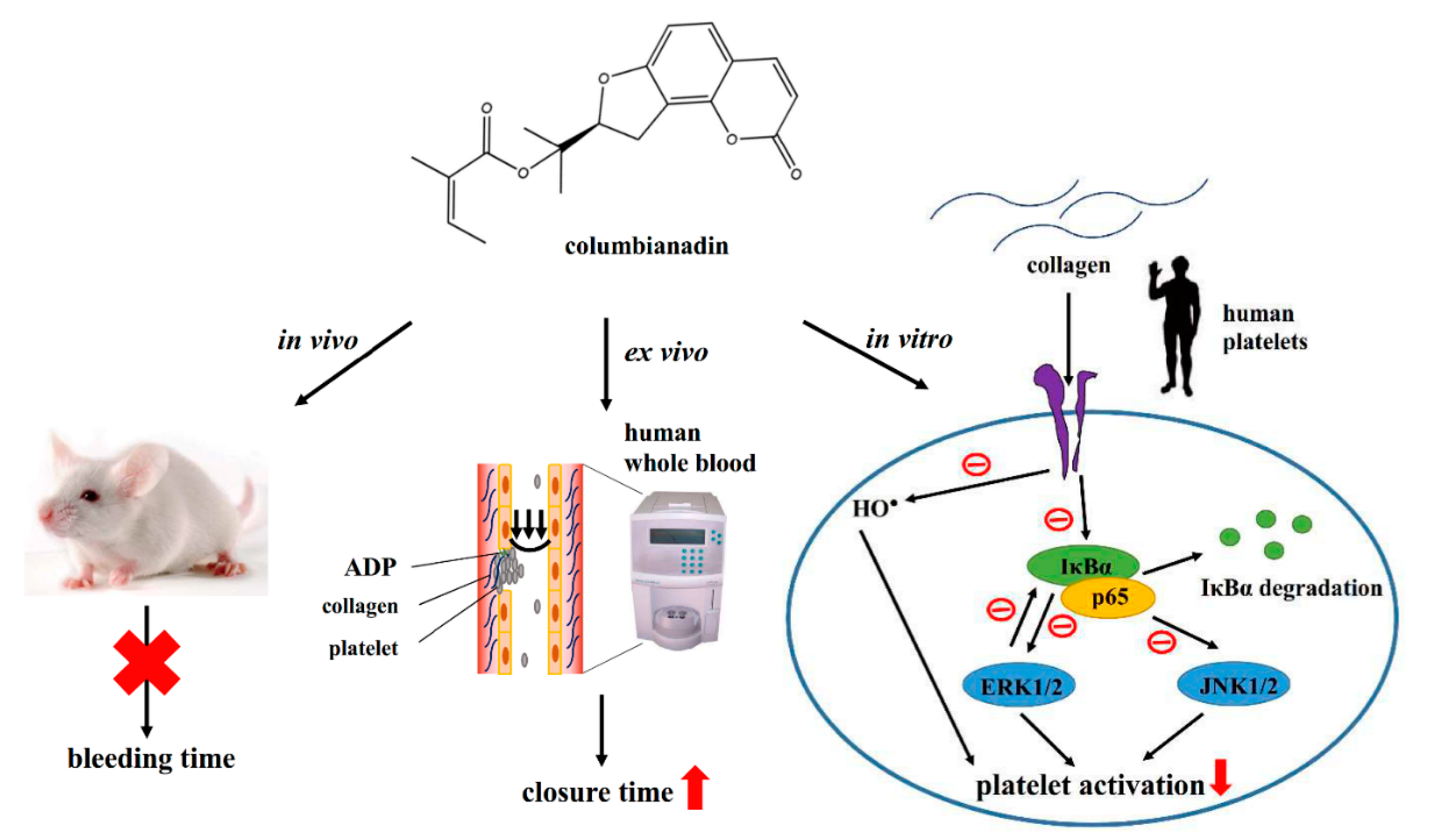
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beckman, M.G.; Hooper, W.C.; Critchley, S.E.; Ortel, T.L. Venous thromboembolism: A public health concern. Am. J. Prev. Med. 2010, 38, S495–S501. [Google Scholar]
- George, J.N. Platelets. Lancet 2000, 355, 1531–1539. [Google Scholar] [PubMed]
- Xu, X.R.; Zhang, D.; Oswald, B.E.; Carrim, N.; Wang, X.; Hou, Y.; Zhang, Q.; Lavalle, C.; McKeown, T.; Marshall, A.H.; et al. Platelets are versatile cells: New discoveries in hemostasis, thrombosis, immune responses, tumor metastasis and beyond. Crit. Rev. Clin. Lab. Sci. 2016, 53, 409–430. [Google Scholar] [PubMed]
- Pikarsky, E.; Porat, R.M.; Stein, I.; Abramovitch, R.; Amit, S.; Kasem, S. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature 2004, 431, 461–466. [Google Scholar]
- Kojok, K.; El-Kadiry, A.E.; Merhi, Y. Role of NF-κB in Platelet Function. Int. J. Mol. Sci. 2019, 20, 4185. [Google Scholar]
- Lee, H.S.; Kim, S.D.; Lee, W.M.; Endale, M.; Kamruzzaman, S.M.; Oh, W.J.; Cho, J.Y.; Kim, S.K.; Cho, H.J.; Park, H.J.; et al. A noble function of BAY 11-7082: Inhibition of platelet aggregation mediated by an elevated cAMP-induced VASP, and decreased ERK2/JNK1 phosphorylations. Eur. J. Pharmacol. 2010, 627, 85–91. [Google Scholar] [PubMed]
- Fuentes, E.; Rojas, A.; Palomo, I. NF-κB signaling pathway as target for antiplatelet activity. Blood Rev. 2016, 30, 309–315. [Google Scholar] [PubMed]
- Rivadeneyra, L.; Carestia, A.; Etulain, J.; Pozner, R.G.; Fondevila, C.; Negrotto, S.; Schattner, M. Regulation of platelet responses triggered by Toll-like receptor 2 and 4 ligands is another non-genomic role of nuclear factor-kappaB. Thromb. Res. 2014, 133, 235–243. [Google Scholar]
- Kang, J.I.; Hong, J.Y.; Choi, J.S.; Lee, S.K. Columbianadin inhibits cell proliferation by inducing apoptosis and necroptosis in HCT116 colon cancer cells. Biomol. Ther. 2016, 24, 320–327. [Google Scholar]
- Zhang, C.; Hsu, A.C.; Pan, H.; Gu, Y.; Zuo, X.; Dong, B.; Wang, Z.; Zheng, J.; Lu, J.; Zheng, R.; et al. Columbianadin Suppresses Lipopolysaccharide (LPS)-Induced Inflammation and Apoptosis through the NOD1 Pathway. Molecules 2019, 24, 549. [Google Scholar]
- Lim, H.J.; Lee, J.H.; Choi, J.S.; Lee, S.K.; Kim, Y.S.; Kim, H.P. Inhibition of airway inflammation by the roots of Angelica decursiva and its constituent, columbianadin. J. Ethnopharmacol. 2014, 155, 1353–1361. [Google Scholar]
- Hou, S.M.; Hsia, C.W.; Tsai, C.L.; Hsia, C.H.; Jayakumar, T.; Velusamy, M.; Sheu, J.R. Modulation of human platelet activation and in vivo vascular thrombosis by columbianadin: Regulation by integrin αIIbβ3 inside-out but not outside-in signals. J. Biomed. Sci. 2020, 27, 60. [Google Scholar] [PubMed]
- Jilma, B. Platelet function analyzer (PFA-100): A tool to quantify congenital or acquired platelet dysfunction. J. Lab. Clin. Med. 2001, 138, 152–163. [Google Scholar]
- Chen, W.F.; Lee, J.J.; Chang, C.C.; Lin, K.H.; Wang, S.H.; Sheu, J.R. Platelet protease-activated receptor (PAR)4, but not PAR1, associated with neutral sphingomyelinase responsible for thrombin-stimulated ceramide-NF-κB signaling in human platelets. Haematologica 2013, 98, 793–801. [Google Scholar]
- Chou, D.S.; Hsiao, G.; Shen, M.Y.; Tsai, Y.J.; Chen, T.F.; Sheu, J.R. ESR spin trapping of a carbon-centered free radical from agonist-stimulated human platelets. Free Radic. Biol. Med. 2005, 39, 237–248. [Google Scholar] [PubMed]
- Lin, Y.; Bai, L.; Chen, W.; Xu, S. The NF-kappaB activation pathways, emerging molecular targets for cancer prevention and therapy. Expert Opin. Ther. Targets 2010, 14, 45–55. [Google Scholar]
- Lee, D.H.; Goldberg, A.L. Proteasome inhibitors: Valuable new tools for cell biologists. Trends Cell Biol. 1998, 8, 397–403. [Google Scholar]
- Ghosh, S.; Hayden, M.S. New regulators of NF-kappaB in inflammation. Nat. Rev. Immunol. 2008, 8, 837–848. [Google Scholar]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar]
- Ali, F.Y.; Davidson, S.J.; Moraes, L.A.; Traves, S.L.; Paul-Clark, M.; Bishop-Bailey, D.; Warner, T.D.; Mitchell, J.A. Role of nuclear receptor signaling in platelets: Antithrombotic effects of PPARbeta. FASEB J. 2006, 20, 326–328. [Google Scholar] [PubMed]
- Moraes, L.A.; Paul-Clark, M.J.; Rickman, A.; Flower, R.J.; Goulding, N.J.; Perretti, M. Ligand-specific glucocorticoid receptor activation in human platelets. Blood 2005, 106, 4167–4175. [Google Scholar] [PubMed] [Green Version]
- Malaver, E.; Romaniuk, M.A.; D’Atri, L.P.; Pozner, R.G.; Negrotto, S.; Benzadon, R.; Schattner, M. NF-kappaB inhibitors impair platelet activation responses. J. Thromb. Haemost. 2009, 7, 1333–1343. [Google Scholar]
- Liu, F.; Morris, S.; Epps, J.; Carroll, R. Demonstration of an activation regulated NF-kappaB/I-kappaBalpha complex in human platelets. Thromb. Res. 2002, 106, 199–203. [Google Scholar] [PubMed]
- Salanova, B.; Choi, M.; Rolle, S.; Wellner, M.; Luft, F.C.; Kettritz, R. Beta2-integrins and acquired glycoprotein IIb/IIIa (GPIIb/IIIa) receptors cooperate in NF-kappaB activation of human neutrophils. J. Biol. Chem. 2007, 282, 27960–27969. [Google Scholar] [PubMed] [Green Version]
- Bugaud, F.; Nadal-Wollbold, F.; Levy-Toledano, S.; Rosa, J.P.; Bryckaert, M. Regulation of c-jun-NH2 terminal kinase and extracellular-signal regulated kinase in human platelets. Blood 1999, 94, 3800–3805. [Google Scholar] [PubMed]
- Kauskot, A.; Adam, F.; Mazharian, A.; Ajzenberg, N.; Berrou, E.; Bonnefoy, A.; Rosa, J.P.; Hoylaerts, M.F.; Bryckaert, M. Involvement of the mitogen-activated protein kinase c-Jun NH2-terminal kinase 1 in thrombus formation. J. Biol. Chem. 2007, 282, 31990–31999. [Google Scholar] [PubMed] [Green Version]
- Roger, S.; Pawlowski, M.; Habib, A.; Jandrot-Perrus, M.; Rosa, J.P.; Bryckaert, M. Costimulation of the Gi-coupled ADP receptor and the Gq-coupled TXA2 receptor is required for ERK2 activation in collagen-induced platelet aggregation. FEBS. Lett. 2004, 556, 227–235. [Google Scholar]
- Falker, K.; Lange, D.; Presek, P. ADP secretion and subsequent P2Y12 receptor signalling play a crucial role in thrombin-induced ERK2 activation in human platelets. Thromb. Haemost. 2004, 92, 114–123. [Google Scholar]
- Toth-Zsamboki, E.; Oury, C.; Cornelissen, H.; De Vos, R.; Vermylen, J.; Hoylaerts, M.F. P2X1-mediated ERK2 activation amplifies the collagen-induced platelet secretion by enhancing myosin light chain kinase activation. J. Biol. Chem. 2003, 278, 46661–46667. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsia, C.-W.; Yang, C.-H.; Sheu, J.-R.; Hsia, C.-H.; Tsai, C.-L.; Huang, W.-C.; Chen, T.-Y.; Jayakumar, T.; Bhavan, P.S.; Chang, Y. Reduction of NF-κB Signals in Platelets and Prolongation of Platelet Plug Formation against High Shear Flow in Whole Blood on Human Subject by Columbianadin. Appl. Sci. 2020, 10, 7323. https://0-doi-org.brum.beds.ac.uk/10.3390/app10207323
Hsia C-W, Yang C-H, Sheu J-R, Hsia C-H, Tsai C-L, Huang W-C, Chen T-Y, Jayakumar T, Bhavan PS, Chang Y. Reduction of NF-κB Signals in Platelets and Prolongation of Platelet Plug Formation against High Shear Flow in Whole Blood on Human Subject by Columbianadin. Applied Sciences. 2020; 10(20):7323. https://0-doi-org.brum.beds.ac.uk/10.3390/app10207323
Chicago/Turabian StyleHsia, Chih-Wei, Chih-Hao Yang, Joen-Rong Sheu, Chih-Hsuan Hsia, Cheng-Lin Tsai, Wei-Chieh Huang, Ting-Yu Chen, Thanasekaran Jayakumar, Periyakali Saravana Bhavan, and Yi Chang. 2020. "Reduction of NF-κB Signals in Platelets and Prolongation of Platelet Plug Formation against High Shear Flow in Whole Blood on Human Subject by Columbianadin" Applied Sciences 10, no. 20: 7323. https://0-doi-org.brum.beds.ac.uk/10.3390/app10207323