1. Introduction
It is almost taken for granted that the days of incurable diseases and the uncontrollable spread of infections are a thing of the past. However, the rising tide of multiresistant pathogens serves as a wake-up call to researchers that new sources of antimicrobial compounds are an urgent priority [
1,
2]. The world health organisation (WHO) has described the current shortage of new antibiotics as a threat to the containment of drug resistant infections, stating that it is insufficient to tackle the challenge of antimicrobial resistance.
There have been several attempts to revive the antibiotic discovery pipeline after the lull of the past few decades by using automated processes. This mainly took the form of combinatorial chemistry which synthesized large repositories of small compounds derived from antibiotic core structures and screened these against an array of pathogenic bacterial antigens [
3,
4]. The investment in this technology did not produce as many lead compounds as expected, perhaps due to the limited number of core structures. Although chemical modification of existing antibiotic core units is the current failsafe of antimicrobial chemotherapy [
5], major investment in antibiotic discoveries was abandoned by major drug companies mainly due to high cost and limited returns scenarios [
4]. As a consequence, most of the new antibiotic discoveries are now in the hands of small to medium enterprises (SMEs) that have fewer resources [
2].
The discovery of new antibiotics is now heavily reliant on innovative solutions. Some researchers have returned to isolation of antimicrobial-producing organisms from the natural environment as it offers the prospect of increased chemical diversity of core antibiotic structures [
6]. This has included an examination of traditional medicine as a new source of lead compounds [
7,
8]. One of the inspirations for research into traditional medicine may have come from Geoffrey Cordell, who devised a series of systematic searches of plant metabolites for anticancer medicines. This included an ethnomedical approach, which evaluated and highlighted the importance of written or historical evidence from traditional medicinal practice [
9]. Additionally, there have been a number of reports of epiphytic and endophytic species of
Streptomyces associated with traditional medicine plants from many continents [
10,
11]. Even more interesting is the discovery of traditional medicine soils [
11]. Unlike the antimicrobial discoveries during the peak of post war antibiotic discovery, traditional medical soils are quite specific in their character and location. These have been reported for the Red Soil of Jordan, [
12] and glacial clay from British Columbia, Canada [
13,
14].
Although traditional medicines are quite well known for Chinese and African traditions, many European traditions have almost disappeared or are the preserve of rural backwaters. Some of the last vestiges of these traditions can be found on the furthest fringes of Western Europe on the island of Ireland [
15]. Although some of this traditional medicine has been partially documented [
16,
17], many of the traditional cures were closely guarded secrets passed down through generations that have now almost faded from memory. However, these cures are still associated with specific habitats which enables us to unravel some of their common characteristics. The area of the West Fermanagh Scarplands is one such area. It constitutes the majority of limestone grassland in Northern Ireland, a consequence of the weathering of the underlying limestone bedrock which forms a thin layer of slightly alkaline soil. The area is well known for its caves, limestone pavement and upland bog, which provide unique habitats for a large diversity of flora. The area is also known for traditional medicines or “cures”, some of which are publicly known and others are secrets passed down through generations. According to anecdotal evidence, this soil has been used for millennia for treatment of various ailments including skin infections, ring worm and digestive problems. In previous research we isolated
Streptomyces sp. myrophorea from this very specific area of upland grassland that had inhibitory activity against several of the most common hospital multiresistant (ESKAPE) pathogens [
18].
However, respecting the strong sensitivities around this specific soil, which remains within consecrated ground, we feel this cannot be considered as an exploitable resource for scientists in any normal sense. Instead, we returned to an adjacent upland alkaline grassland region which is still associated with traditional medicine approximately a kilometer from our original discovery to examine whether the same type of soil could be a new resource for antimicrobial discovery.
2. Materials and Methods
2.1. Soil Samples
Soil sampling permissions were granted by a local landowner in Boho, County Fermanagh, Northern Ireland, at a distance of a few kilometers from a previous examination [
18].
2.2. Soil pH Measurement
Triplicate samples were used to measure the soil pH using a Jenway standard digital 3510 pH meter. Approximately 10 g of field soil was dissolved with 20 mL of water, shaken gently to form a slurry, incubated for 30 min and measured [
19].
2.3. Microorganisms
S. aureus MRSA (ATCC 43300), S. aureus (MSSA), Pseudomonas aeruginosa PA01, Starmerella bombicola, Bacillus subtilis (ATCC 6633), Escherichia coli (ATCC 25922) and Propionibacterium acne (DSM 1897) were kind gifts from Prof. Ibrahim Banat (Ulster University).
2.4. Soil Sample Pretreatment
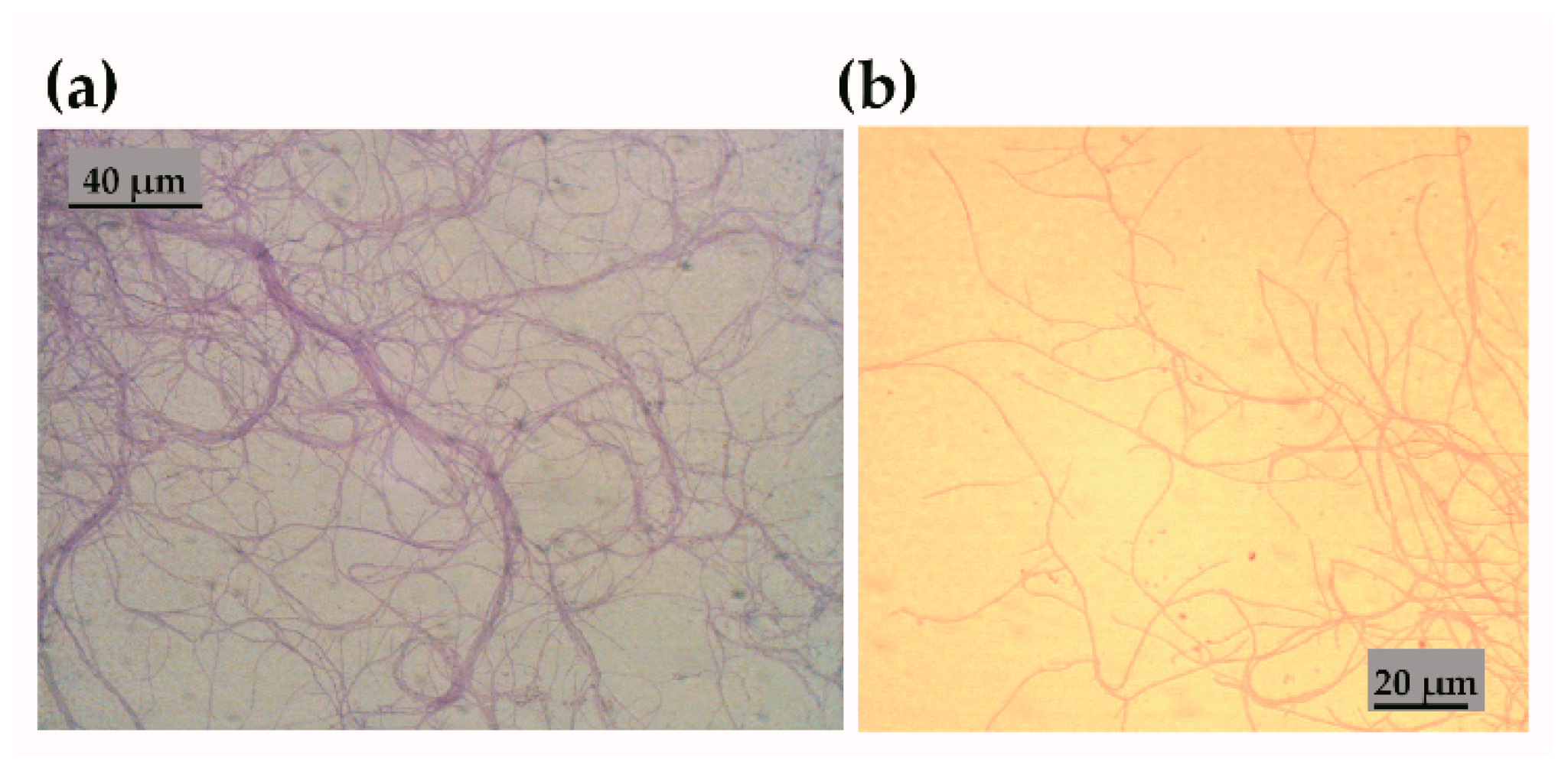
Soil samples were air dried in sterile laminar flow cabinet. Soil samples were then incubated for one hour at 60 °C, dry heat. A small fraction of this soil was removed and dissolved in 1 mL of sterile water. This was mixed on a rotary shaker for 30 min before removing 10 µL aliquots and inoculating onto one-fifth nutrient strength ISP2 containing 0.03% chlorhexidine gluconate (to discourage fungi) and original mineral extract from soil. These Petri dishes were then incubated for 2 h at room temperature followed by 2 weeks at 8 °C.
2.5. Media
The core media used in these experiments were plain “base” agar, ISP-2 agar, TSA, Muller Hinton and Sabouraud dextrose agar (Oxoid, Thermo Fisher Scientific Inc, Basingstoke, Hampshire, UK). Sterilized washes of original soil were also used in some media as a mineral supplement. These were prepared by taking a small portion of the original soil from the isolation area, mixing with sterile water (5 g per 20 mL w/v) and vortexed for several minutes (to supplement with essential minerals that might not be found in laboratory media). This was centrifuged in small aliquots at 14,000× g. It was then passed through a 0.2 µm filter. Five mL of this wash may be added to 200 mL of agar.
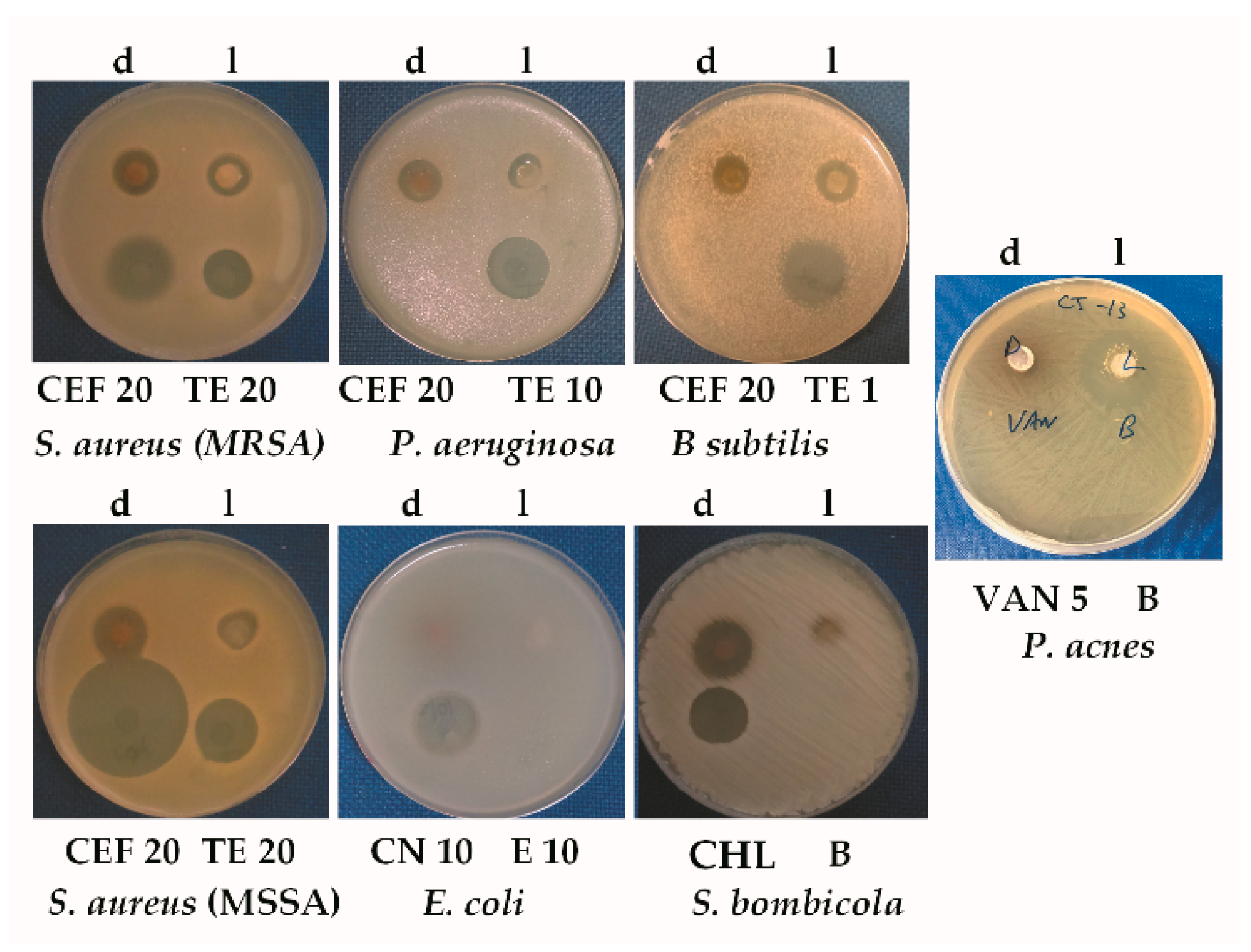
2.6. Antibiotic Tests
Antimicrobial tests were based on a modification of an ultrasensitive assay developed by Lehrer [
20] and a
Streptomyces overlay method [
21]. Briefly, several wells were cored from sterilized base agar without nutrients once it had cooled. These were filled with either antibiotic solutions or agar cores from
Streptomyces cultivation (killed by inverting
Streptomyces agar plate over 4 mL chloroform in a watch glass for 1 h). Once the antibiotics had absorbed into the test wells and surrounding agar, several drops of cooled base agar were added to seal the wells that contained liquid (lest there was any spillover once the top layer of agar was added) and to hold the
Streptomyces agar plugs in position. Test bacteria were added to 15 mL of hand warm (no more than 42 °C) Muller Hinton agar. This vessel was rotated to mix the organisms evenly and poured over the base agar. In cases where the organisms were not amenable to this method, such as
S. bombicola, the organisms were streaked over the surface of the agar using a sterile swab. In cases where test pathogen growth was weak on Muller Hinton agar, this was substituted by TSA in the case of
S. aureus or Sabouraud agar in the case of
S. bombicola. Plates were incubated overnight at 28 °C. The presence of a clear zone in the test pathogen indicated inhibitory activity. The procedure was similar for anaerobic organisms such as
Propionibacterium acnes except that the final incubation conditions were 24 h in an anaerobic cabinet.
2.7. DNA Extraction
A single colony of Streptomyces was mixed in 200 μL sterile PBS and inoculated into 25 mL of sterile ISP2 media. The broth was shaken at 28 °C for two days. A subsample was taken to determine the purity of the bacteria. The rest of the sample was pelleted for 10 min at 4000× g. The pellet was suspended in 500 µL of the cryopreservant from a barcoded bead tube supplied by MicrobesNG. The whole genome sequencing was performed by Microbes NG (Units 1–2 First Floor, The BioHub, Birmingham Research Park, UK).
2.8. Sequencing Protocol
Three beads from the cryopreservant tube were washed with extraction buffer containing lysozyme (or lysostaphin for
Staphylococcus sp.) and RNase A, incubated for 25 min at 37 °C. Proteinase K and RNaseA were added and incubated for 5 min at 65 °C. Genomic DNA was purified using an equal volume of SPRI beads and resuspended in EB buffer. DNA was quantified in triplicate using the Quantit dsDNA HS assay in an Eppendorf AF2200 plate reader. Genomic DNA libraries were prepared using Nextera XT Library Prep Kit (Illumina, San Diego, CA, USA) following the manufacturer’s protocol with the following modifications: two nanograms of DNA instead of one were used as input, and PCR elongation time was increased to 1 min from 30 s. DNA quantification and library preparation were carried out on a Hamilton Microlab STAR automated liquid handling system. Pooled libraries were quantified using the Kapa Biosystems Library Quantification Kit for Illumina on a Roche light cycler 96 qPCR machine. Libraries were sequenced on the Illumina HiSeq using a 250 bp paired end protocol. Reads were adapter-trimmed using Trimmomatic 0.30 with a sliding window quality cut off of Q15 [
22]. De novo assembly was performed on samples using SPAdes version 3.7 [
23] and contigs were annotated using Prokka 1.11 [
24].
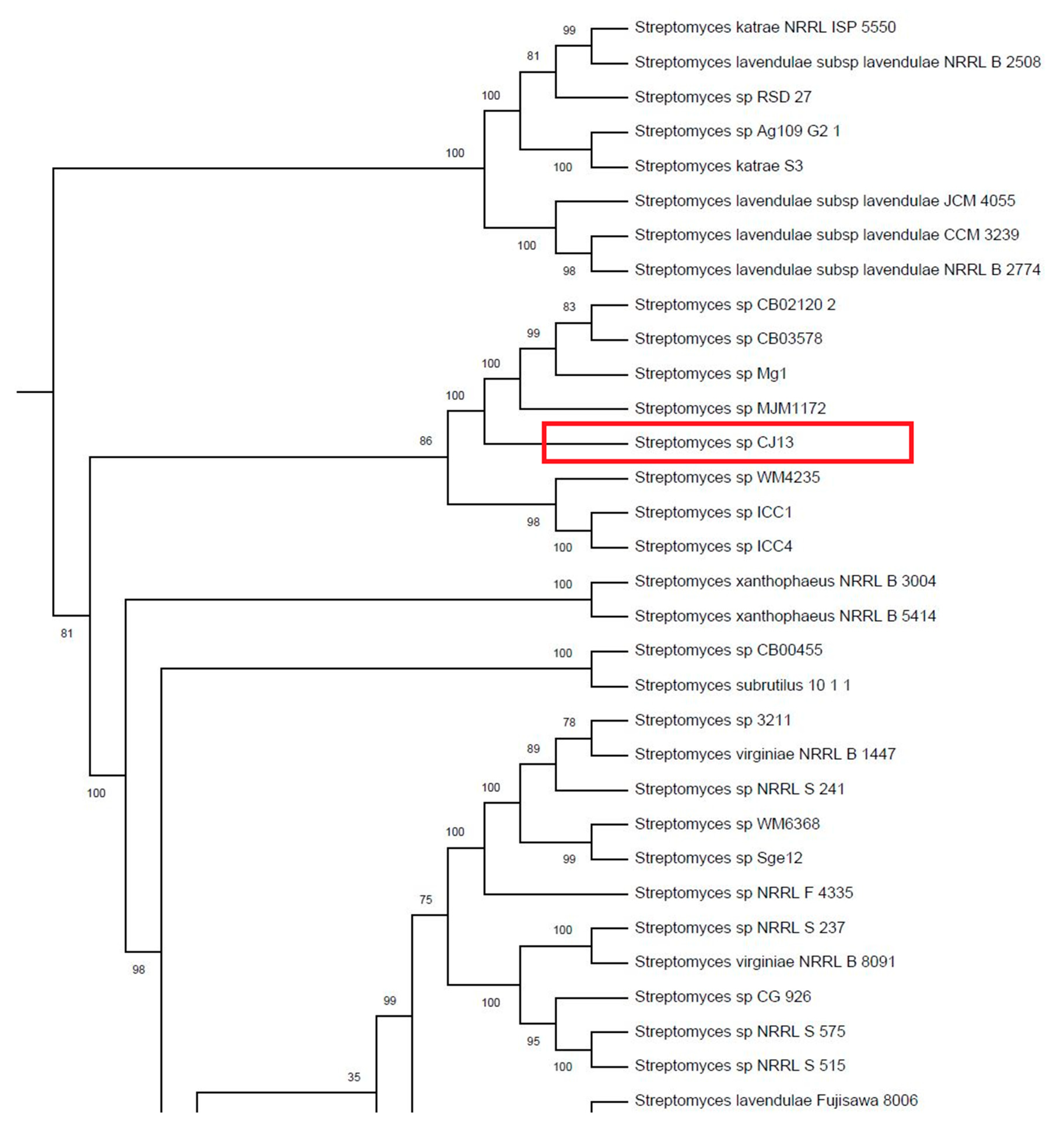
2.9. Phylogenetic Tree
Whole genome sequences (WGS) for all
Streptomycetes were retrieved autonomously from GenBank using a WGET script. Genomes were annotated in Prokka [
24] and the resulting translated CoDing Sequence (CDS) files (.faa files) were used to generate a multilocus, concatenated alignment of 400 proteins in PhyloPhlan [
25]. Proteins used for the alignment were shown to be conserved among bacterial genera [
25]. A maximum-likelihood phylogeny was reconstructed in FastTree [
26] and rendered in iTOL [
27]. The robustness of the phylogeny was assessed using 1000 bootstrap replications. Tree was outgroup-rooted along the lineage leading to
Frankia.
2.10. In Silico Secondary Metabolite Analysis
Genome sequences were analysed using subsystem technology (RAST) version 4.0 (
http://rast.nmpdr.org) and SEED [
28]. Similarities to known antibiotic gene clusters and other secondary metabolites were predicted by antiSMASH [
29,
30].
2.11. Accession Numbers
The whole genome of Streptomyces sp. CJ13 was deposited at DDBJ/ENA/GenBank under the accession JAAZTW000000000. The version described in this paper is version JAAZTW010000000, BioProject PRJNA433829 BioSample SAMN14520815.
4. Discussion
There have been several attempts over the last few decades to replenish the antibiotic discovery pipeline. This has become more urgent given the rise in reports of antimicrobial resistance [
45]. Rather than repeat older methods of antibiotic discovery, researchers have tried new innovations. One such strand is the examination of traditional medicines for antimicrobial producing bacteria or antimicrobials themselves. This follows from successful generation of lead compounds in the field of cancer [
9].
In this manuscript, we isolated a species of
Streptomyces (
Streptomyces sp. CJ13) from an alkaline grassland soil traditionally known for its healing properties. This had a broad range of inhibitory activity against multiresistant bacteria, anaerobes and fungi. There are many antibiotic producing species of
Streptomyces associated with traditional medicines, both as endophytes and epiphytes [
10,
11]. Many of the discoveries during the golden age of antibiotics also came from soils. The difference between the older types of antimicrobial surveys in soils and those that have been recently isolated from traditional medicine is their specific soil character and location. For instance, Kisameet clay is based on a glacial clay deposit found in British Columbia that inhibits ESKAPE pathogens [
13,
14]. Similarly, traditional soil discoveries in Jordan focus on red soil that inhibits various pathogens [
12]. Whilst our experiments were based on alkaline grassland soil, we did not test the whole soil for antimicrobial activity but rather tried to selectively isolate
Streptomyces from the soil. Other studies have shown that karst regions, i.e. regions where there is dissolution of the rocks, especially limestone, support the growth of many
Streptomyces [
46,
47,
48].
By coincidence, some of the closest phylogenetic matches to
Streptomyces sp. CJ13 were
Streptomyces sp. ICC 1 and ICC4 originating from the Iron Curtain Cave in Chilliwack, British Columbia [
32]. Both the West Fermanagh Scarplands and Iron Curtain Cave are regions of karst and share other botanical species including
Spiranthes romanzoffiana (Irish lady’s-tresses) and
Sisyrinchium bermudiana (blue-eyed grass). Additionally, both areas also have indigenous soil-based traditional medicines: in British Columbia from Kisameet Bay and in Fermanagh from the Boho area [
13,
18].
Streptomyces sp. CJ13 also shares phylogenetic similarities with the larger group of
S. lavendulae. This group was originally described as
Actinomyces lavendulae by Waksman and Henrici (1916), although relative distinctions between
Actinomycetes had not been formalised at that time [
33].
One of the world’s most pressing health issues is treating multidrug resistant Gram-negative bacteria. These can cause serious infections such as pneumoniae and urinary tract infections, which are often difficult to treat [
49]. Fortunately, there are many more antibiotic discoveries for regulatory approval than in the previous decades, but still not enough to meet projected patient needs [
2]. The inhibitory activity of
Streptomyces sp. CJ13 covered a surprisingly broad array of organisms, including Gram-negative, multiresistant
P. aeruginosa PA01.
The expression of antimicrobial activity varied between pigmentation types in our experiments. This phenomenon was also noted in the Iron Curtain Cave (ICC) strains from British Columbia [
32]. To maintain a consistently high level of antibiotic activity, the ICC group grew their isolates on minimal media at low temperatures over an extended period of time [
32]. This suggests that more antibiotics were induced by stressful growth conditions.
Streptomyces are known to have silent antibiotic gene synthesis clusters stimulated during stress, microbial competition or interspecies communication [
50,
51,
52,
53]. In contrast to the ICC strains of British Columbia,
Streptomyces sp. CJ13 grew quite rapidly (in a number of days) and also expressed antibiotic activity over that same time period. To maintain optimal antibiotic production, we followed a regime similar to the ICC group using very Spartan media. However, most of our antimicrobial experiments were derived from stocks of
Streptomyces sp. CJ13, which had been frozen at −80 °C in glycerol.
The phylogenetically related group
S. lavendulae produce many antimicrobial substances, including ileumycin, mitomycin C, eurymycin, glomecidin, saframycin, lavendamycin, angucycline, chlorocarcin, mimosamycin, streptothricin and streptomycin [
33,
54,
55,
56,
57]. Additionally, secondary metabolite prediction software showed similarities to the antibiotic gene synthesis clusters for salinichelins, mediomycin A, weishanmycin, combamide (macrolactam), heat stable antifungal factor and SAL-2242. However, preliminary studies of antimicrobial
Streptomyces sp. CJ13 extracts by mass spectrometry failed to find any identities with compounds in the
Streptomyces or small molecule databases (results not shown). This raises the possibility that the inhibitory activity detected in
Streptomyces sp. CJ13 could be from new antimicrobials. Further rounds of purification and analysis by mass spectrometry and NMR might reveal the identity of these compounds.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}