
Comparative Evaluation of Apoptosis Induction Using Needles, Bark, and Pollen Extracts and Essential Oils of Pinus eldarica in Lung Cancer Cells

 ,
, 
Abstract
:1. Introduction
2. Results
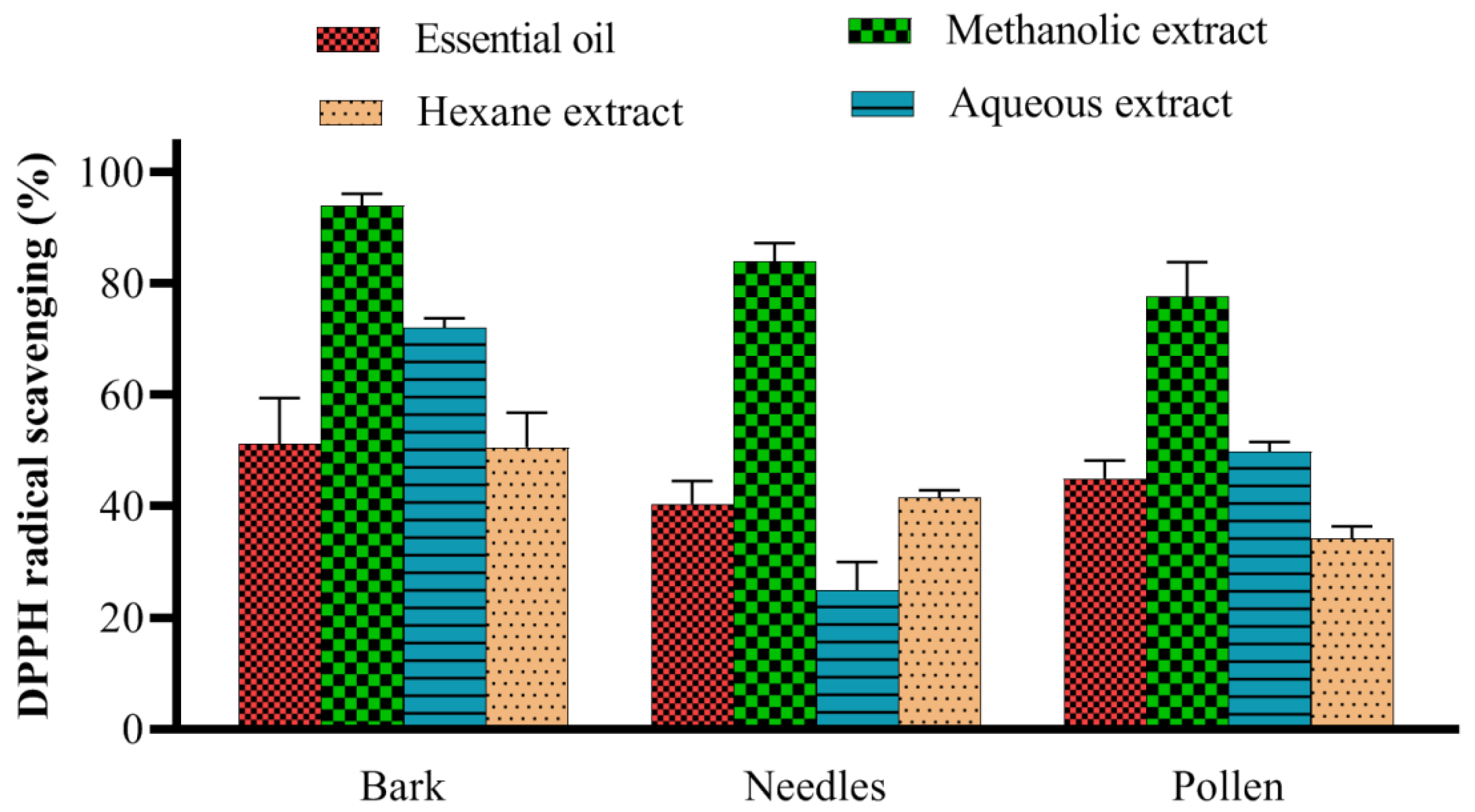
2.1. DPPH Radical Scavenging Assay
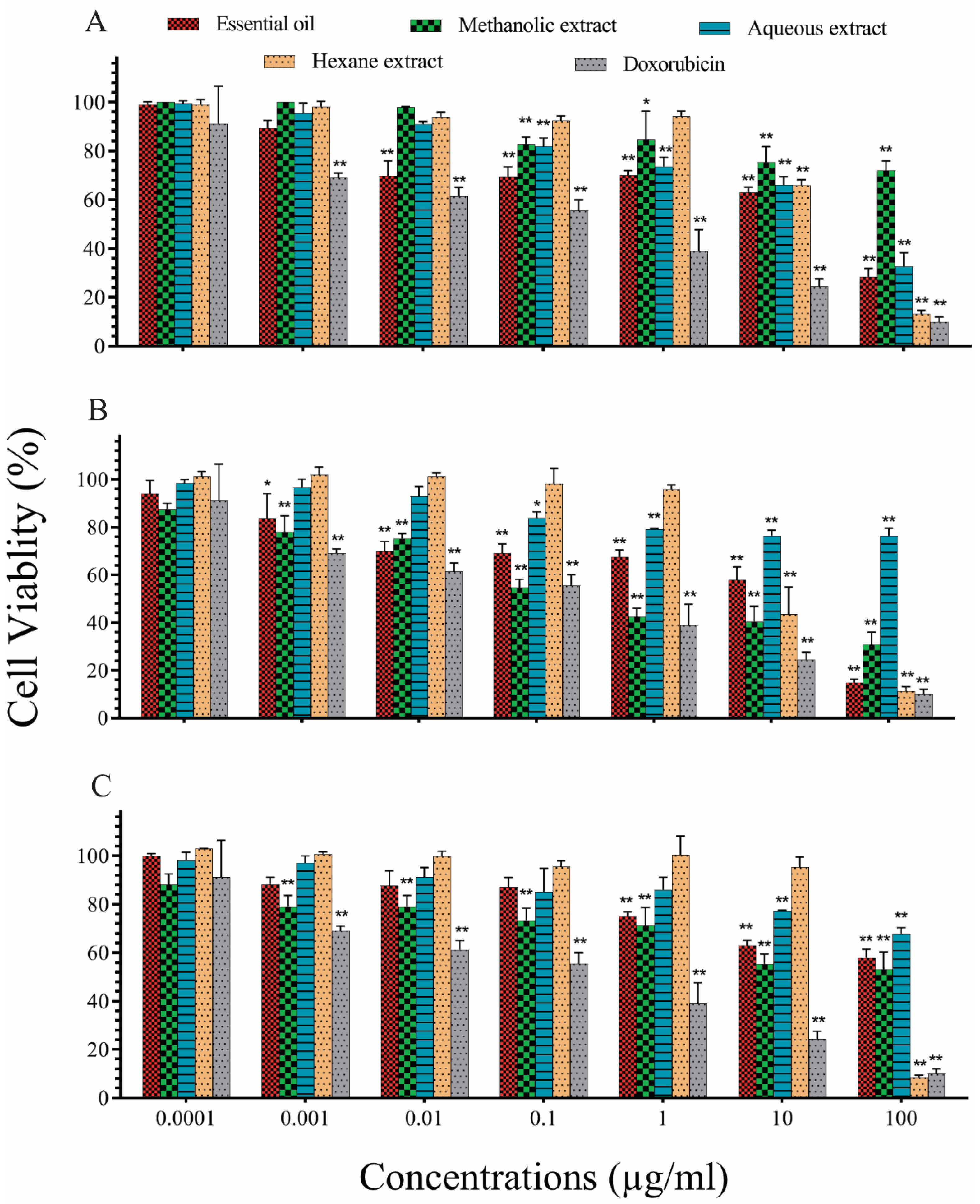
2.2. In Vitro Evaluation of Cytotoxic Effects of P. eldarica Extracts and Essential Oils on A549 Lung Cancer Cells
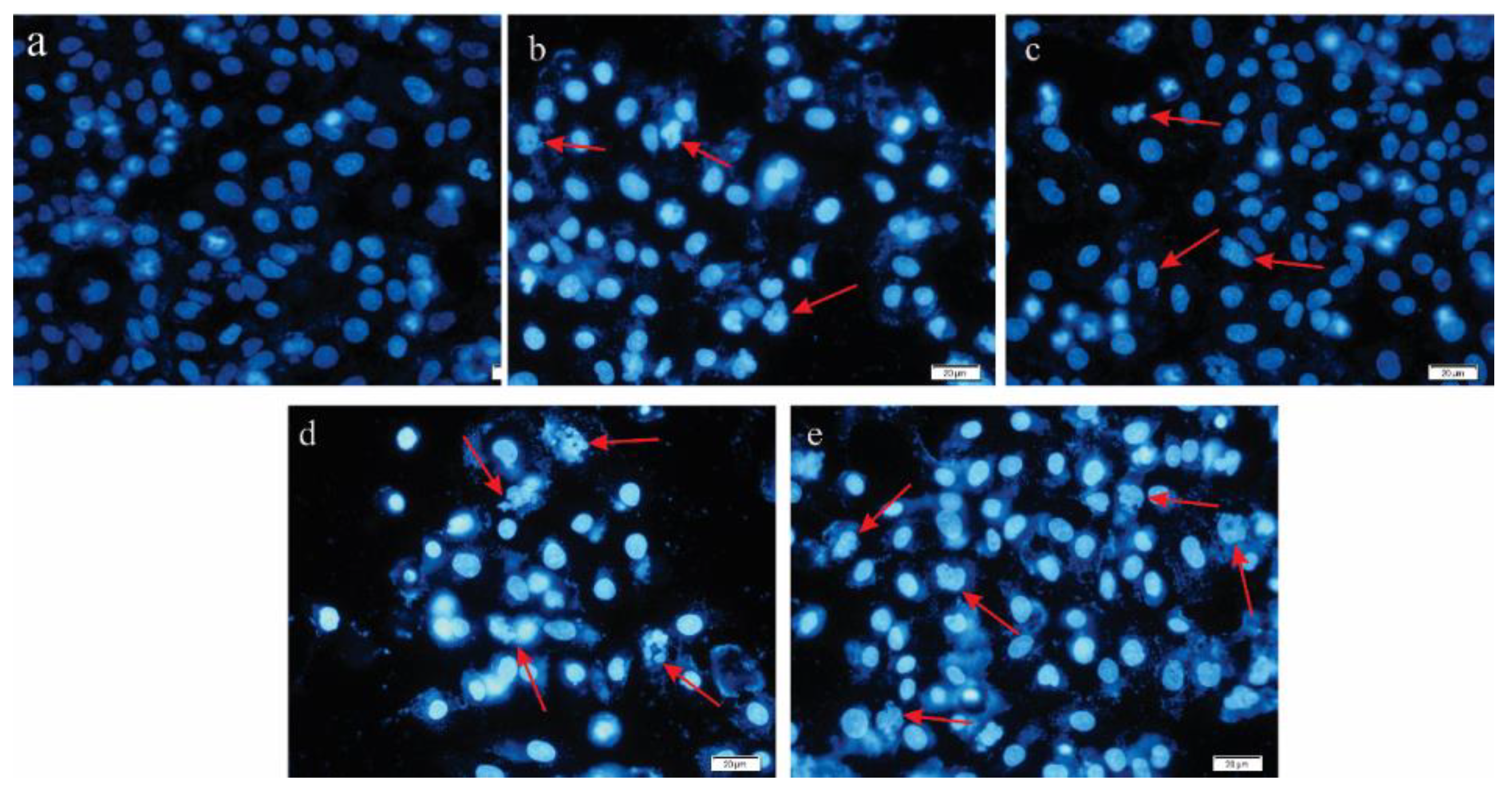
2.3. Evaluating Apoptosis Induction by DAPI Staining

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (µg/mL) a | Essential Oil | Methanolic Extracts | Aqueous Extracts | Hexane Extracts |
|---|---|---|---|---|
| Pollen | 88.9 ± 36 | 49.4 ± 24 | 174.5 ± 46.7 | 31.7 ± 7.2 |
| Needles | 10.3 ± 4.1 | 0.3 ± 0.15 | 262.4 ± 43.87 | 8.7 ± 0.9 |
| Bark | 17.9 ± 7.2 | 209 ± 64 | 25.1 ± 6.8 | 18.0 ± 1.5 |
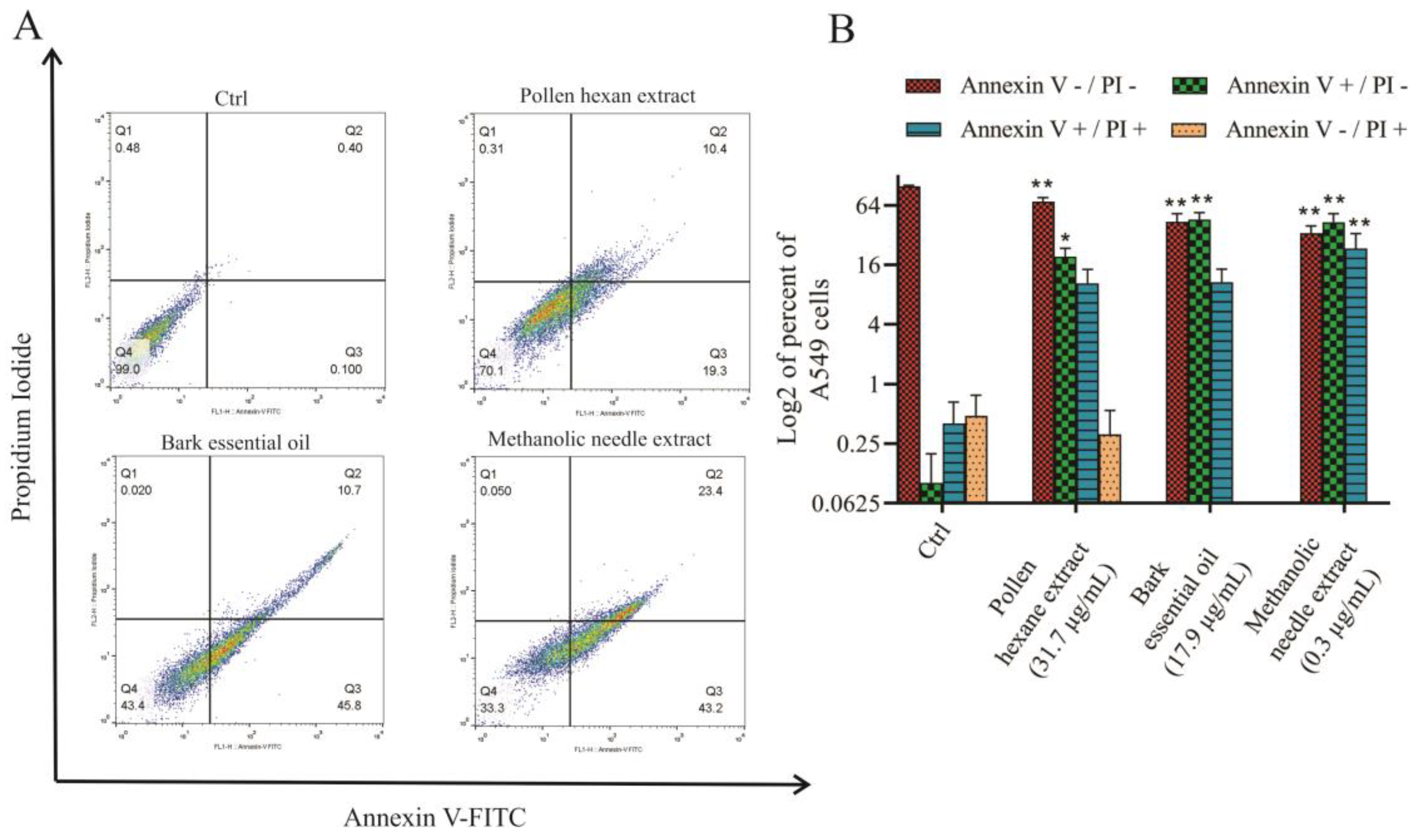
2.4. Analysis of Apoptosis Using Flow Cytometry
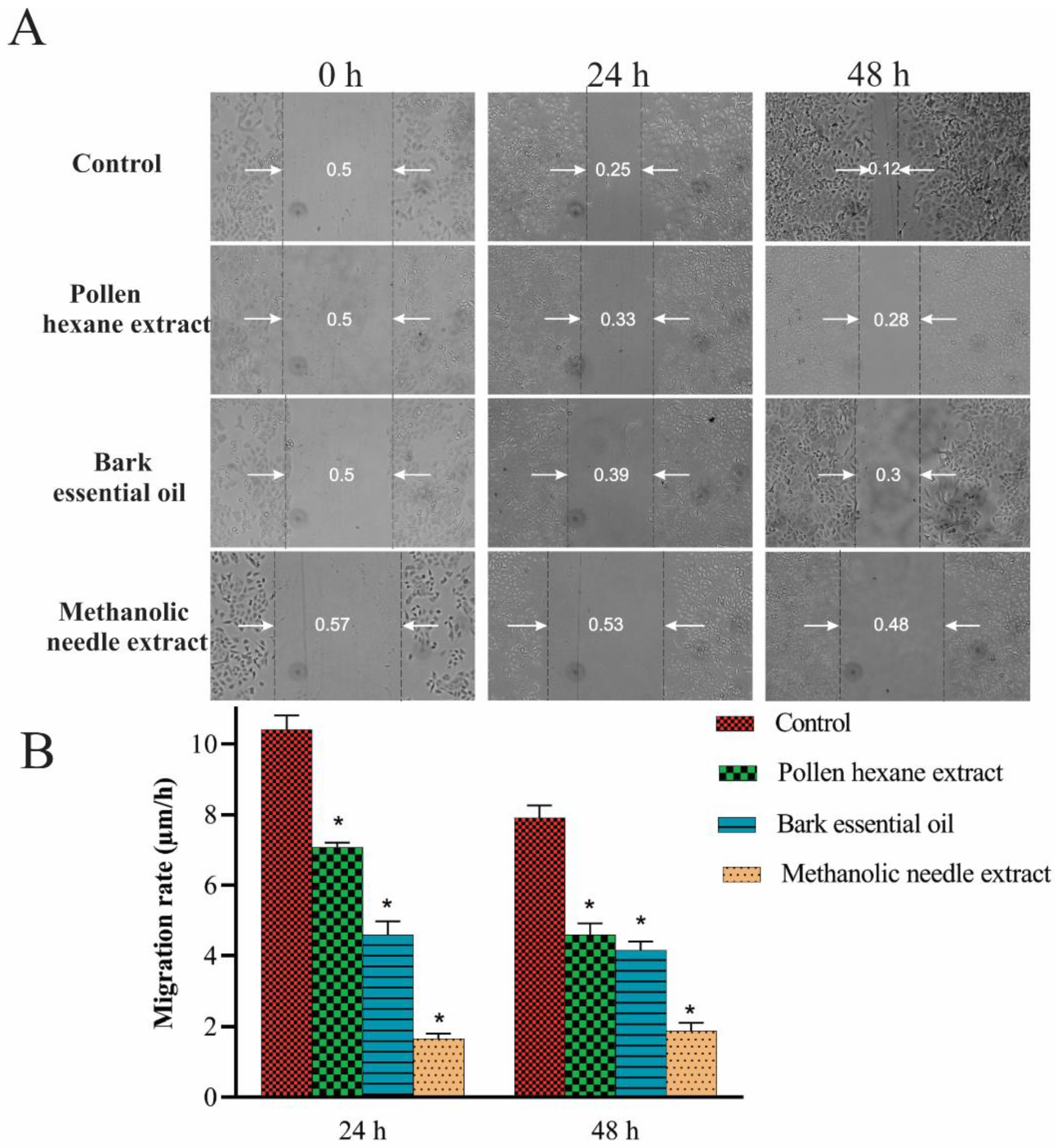
2.5. Scratch Migration Assay
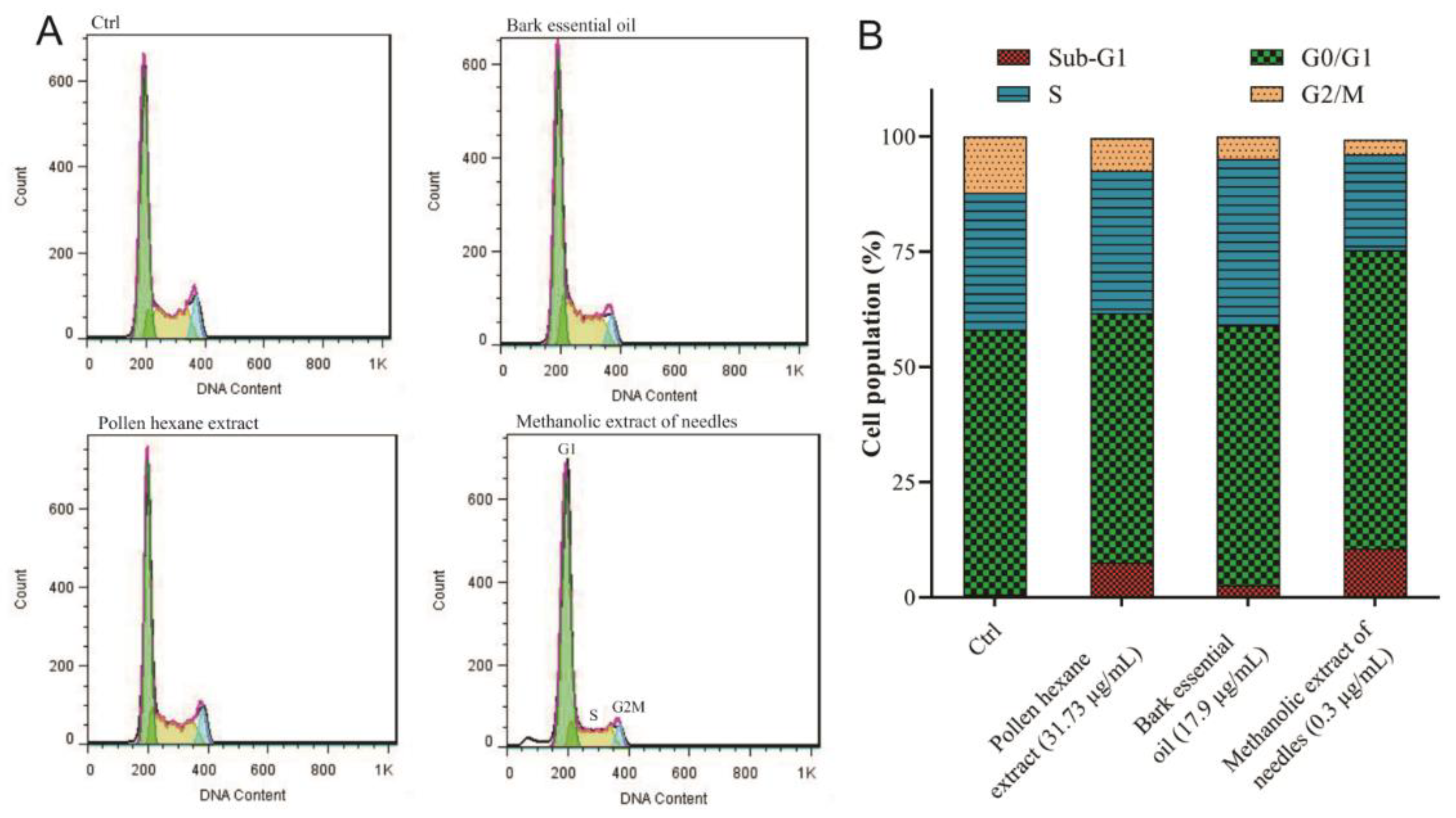
2.6. Cell Cycle Analysis and Apoptosis Evaluation
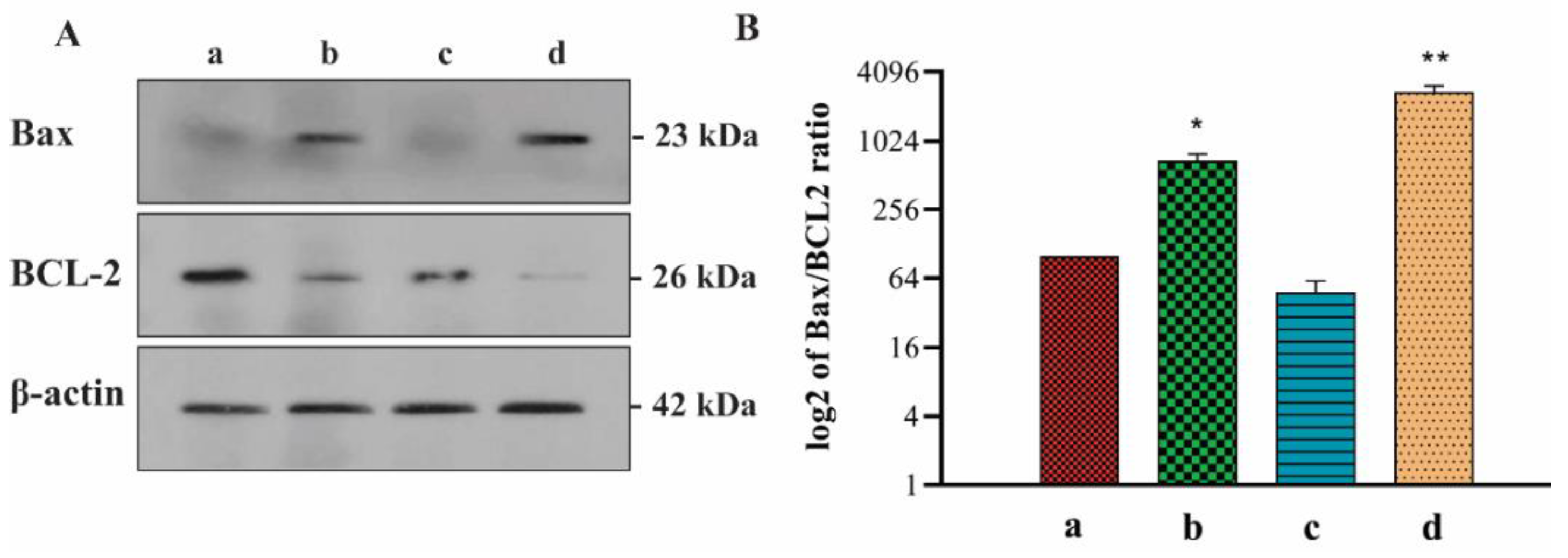
2.7. Western Blot Analysis: Effects of P. eldarica Extracts and Essential Oil on Bax/Bcl 2 Ratio
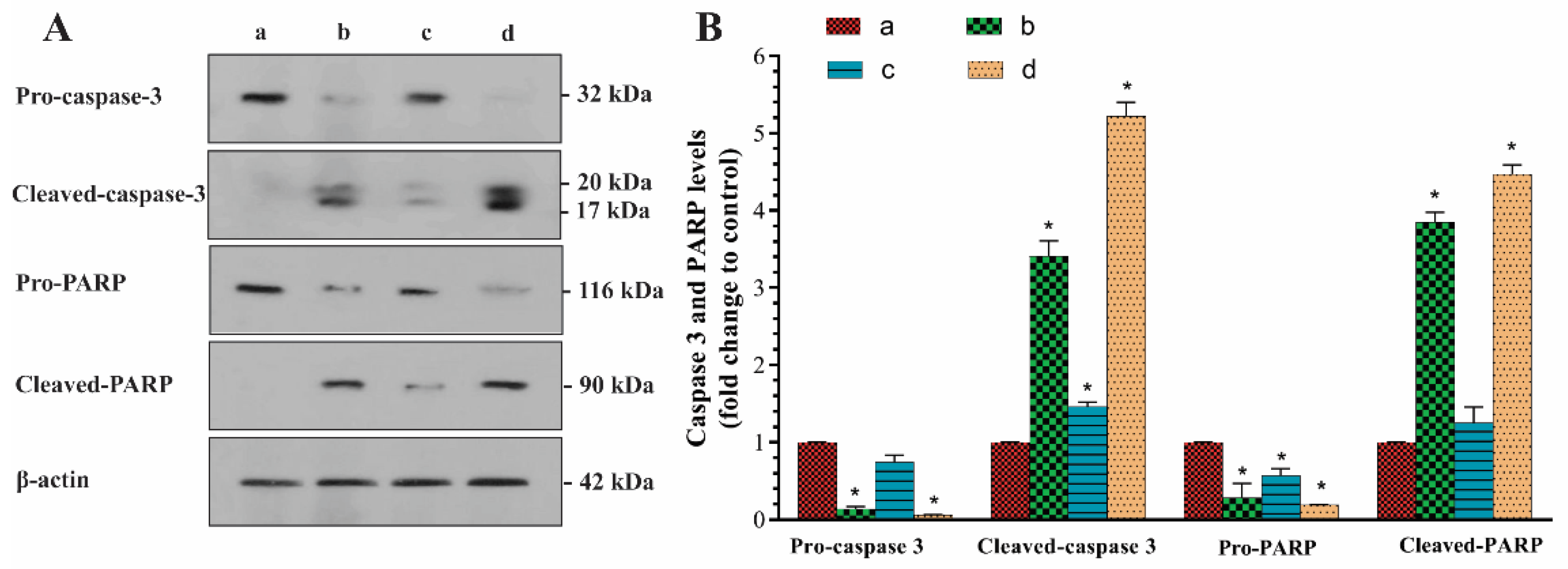
2.8. Western Blot Analysis: Effects of P. eldarica Extracts and Essential Oil on Caspase-3 Activation and PARP Cleavage
2.9. Western Blot Analysis: Effects of P. eldarica Extracts and Essential Oil on p53
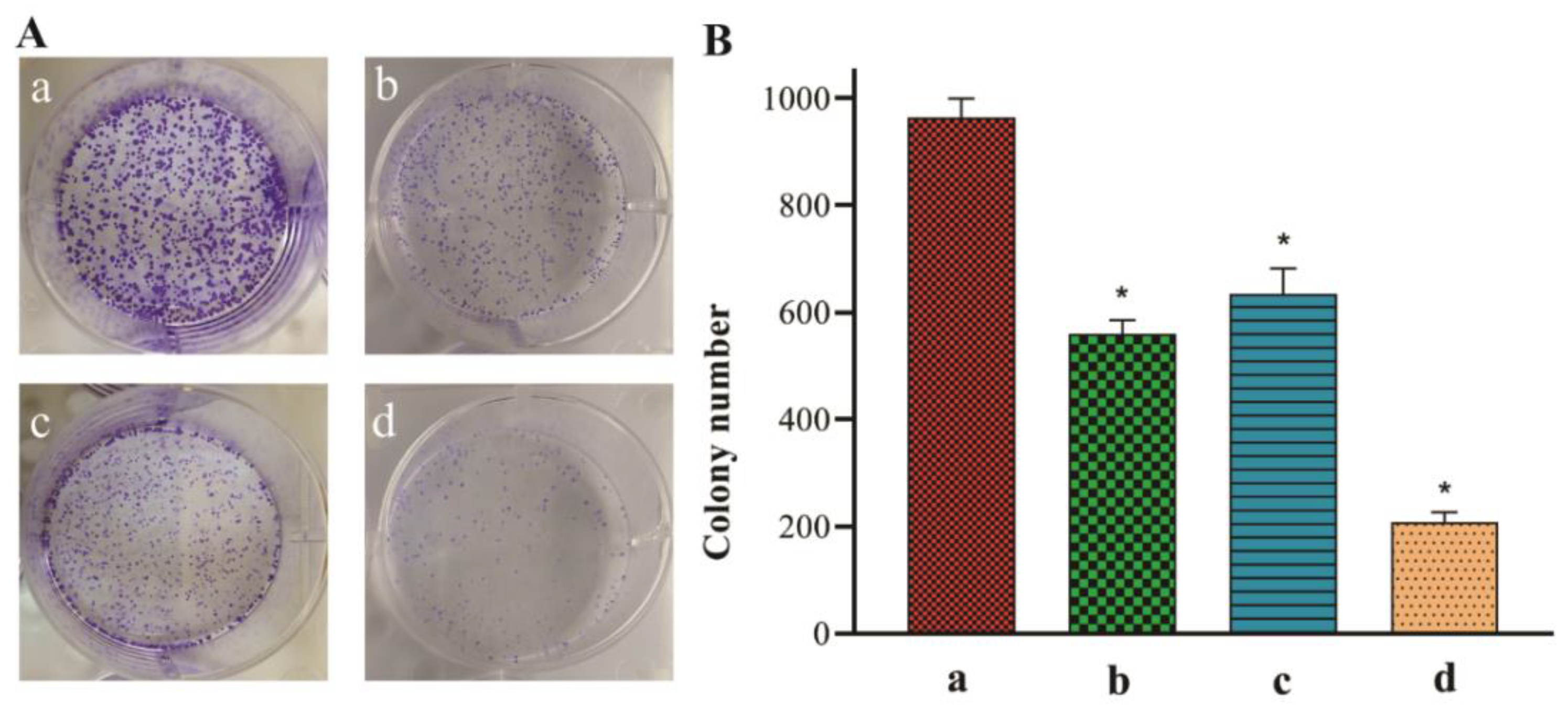
2.10. Colony Formation Assay
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Plant Material
4.3. DPPH Radical Scavenging Assay
4.4. Cell Culture
4.5. MTT-Based Cytotoxicity Assay
4.6. DAPI Staining
4.7. Annexin V/PI Flow Cytometry
4.8. Scratch Migration Assay
4.9. Cell Cycle Analysis
4.10. Western Blot Analysis
4.11. Colony Formation Assay
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Gurib-Fakim, A. Medicinal plants: Traditions of yesterday and drugs of tomorrow. Mol. Asp. Med. 2006, 27, 1–93. [Google Scholar] [CrossRef]
- Kumar, M.A.S.; Nair, M.; Hema, P.S.; Mohan, J.; Santhoshkumar, T.R. Pinocembrin triggers Bax-dependent mitochondrial apoptosis in colon cancer cells. Mol. Carcinog. 2007, 46, 231–241. [Google Scholar] [CrossRef]
- Kim, J.H.; Yoon, E.K.; Chung, H.J.; Park, S.Y.; Hong, K.M.; Lee, C.H.; Lee, Y.S.; Choi, K.; Yang, Y.; Kim, K.; et al. P53 acetylation enhances Taxol-induced apoptosis in human cancer cells. Apoptosis 2013, 18, 110–120. [Google Scholar] [CrossRef]
- Sun, N.K.; Huang, S.L.; Chang, T.C.; Chao, C.C.K. TLR4 and NFκB signaling is critical for taxol resistance in ovarian carcinoma cells. J. Cell. Physiol. 2018, 233, 2489–2501. [Google Scholar] [CrossRef]
- Ran, J.H.; Shen, T.T.; Wu, H.; Gong, X.; Wang, X.Q. Phylogeny and evolutionary history of Pinaceae updated by transcriptomic analysis. Mol. Phylogenet. Evol. 2018, 129, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, D.W. Gymnosperms. In The Evolution of Plant Form; John Wiley & Sons: Hoboken, NJ, USA, 2012; Volume 45, pp. 141–161. [Google Scholar]
- Kaushik, D.; Kumar, A.; Kaushik, P.; Rana, A.C. Analgesic and anti-inflammatory activity of pinus roxburghii sarg. Adv. Pharmacol. Sci. 2012, 2012, 245431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parihar, P.; Parihar, L.; Bohra, A. Antibacterial activity of extracts of Pinus roxburghii sarg. Bangladesh J. Bot. 2006, 35, 85–86. [Google Scholar]
- Huseini, F.H.; Mehrzadi, S.; Ghaznavi, H.; Tajallizadehkhoob, Y.; Fakhrzadeh, H. Effects of pinus eldarica Medw. nut extract on blood glucose and cholesterol levels in hypercholesterolemic alloxan-induced diabetic rats. J. Med. Plants 2013, 12, 68–74. [Google Scholar]
- Kwak, C.S.; Moon, S.C.; Lee, M.S. Antioxidant, antimutagenic, and antitumor effects of pine needles (Pinus densiflora). Nutr. Cancer 2006, 56, 162–171. [Google Scholar] [CrossRef]
- Ghaffari, T.; Kafil, H.S.; Asnaashari, S.; Farajnia, S.; Delazar, A.; Baek, S.C.; Hamishehkar, H.; Kim, K.H. Chemical Composition and Antimicrobial Activity of Essential Oils from the Aerial Parts of Pinus eldarica Grown in Northwestern Iran. Molecules 2019, 24, 3203. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Park, J.H.; Park, E. Effect of Sulgidduk containing pine needle juice on lipid metabolism in high fat-cholesterol diet induced dyslipidemic rats. J. Nutr. Health 2019, 51, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Rohdewald, P. A review of the French maritime pine bark extract (Pycnogenol®), a herbal medication with a diverse clinical pharmacology. Int. J. Clin. Pharmacol. Ther. 2002, 40, 158–168. [Google Scholar] [CrossRef]
- Guri, A.; Kefalas, P.; Roussis, V. Antioxidant potential of six pine species. Phytother. Res. 2006, 20, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S. Evaluation of in vitro antioxidant activity of the water extract obtained from dried pine needle (pinus densiflora). Prev. Nutr. Food Sci. 2018, 23, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.B.; Seo, E.W.; Jeong, H.J. Effect of extracts from pine needle against oxidative DNA damage and apoptosis induced by hydroxyl radical via antioxidant activity. Food Chem. Toxicol. 2009, 47, 2135–2141. [Google Scholar] [CrossRef] [PubMed]
- Iravani, S.; Zolfaghari, B. Phytochemical analysis of Pinus eldarica bark. Res. Pharm. Sci. 2014, 9, 243–250. [Google Scholar]
- Borner, C. The Bcl-2 protein family: Sensors and checkpoints for life-or-death decisions. Mol. Immunol. 2003, 39, 615–647. [Google Scholar] [CrossRef]
- Lee, T.K.; Roh, H.S.; Yu, J.S.; Baek, J.; Lee, S.; Ra, M.; Kim, S.Y.; Baek, K.H.; Kim, K.H. Pinecone of Pinus koraiensis Inducing Apoptosis in Human Lung Cancer Cells by Activating Caspase-3 and its Chemical Constituents. Chem. Biodivers. 2017, 14. [Google Scholar] [CrossRef]
- Li, K.; Li, Q.; Li, J.; Gao, D.; Zhang, T.; Han, Z.; Zheng, F. Effect of procyanidins from Pinus koraiensis bark on growth inhibition and expression of PCNA and TNF-α in mice with U14 cervical cancer. Therapy 2007, 4, 685–690. [Google Scholar] [CrossRef]
- Kang, H.; Lim, J.W.; Kim, H. Inhibitory effect of Korean Red Ginseng extract on DNA damage response and apoptosis in Helicobacter pylori-infected gastric epithelial cells. J. Ginseng Res. 2020, 44, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Sharifan, A.; Etebari, M.; Zolfaghari, B.; Aliomrani, M. Investigating the effects of bark extract and volatile oil of Pinus eldarica against cisplatin-induced genotoxicity on HUVECs cell line. Toxicol. Res. 2021, 10, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Sarvmeili, N.; Jafarian-Dehkordi, A.; Zolfaghari, B. Cytotoxic effects of Pinus eldarica essential oil and extracts on HeLa and MCF-7 cell lines. Res. Pharm. Sci. 2016, 11, 476–483. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Zhao, M.; Wang, J.s.; Cui, C.; Yang, B.; Jiang, Y.; Zhao, Q. Antioxidant, immunomodulatory and anti-breast cancer activities of phenolic extract from pine (Pinus massoniana Lamb) bark. Innov. Food Sci. Emerg. Technol. 2008, 9, 122–128. [Google Scholar] [CrossRef]
- Dudonné, S.; Vitrac, X.; Coutiére, P.; Woillez, M.; Mérillon, J.M. Comparative study of antioxidant properties and total phenolic content of 30 plant extracts of industrial interest using DPPH, ABTS, FRAP, SOD, and ORAC assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef]
- Miserocchi, G.; Mercatali, L.; Liverani, C.; De Vita, A.; Spadazzi, C.; Pieri, F.; Bongiovanni, A.; Recine, F.; Amadori, D.; Ibrahim, T. Management and potentialities of primary cancer cultures in preclinical and translational studies. J. Transl. Med. 2017, 15, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Zhou, X.; Wang, S.; Trinkle, C. Tumor organoid models in precision medicine and investigating cancer-stromal interactions. Pharmacol. Ther. 2020, 218, 107668. [Google Scholar] [CrossRef] [PubMed]
- Mao, P.; Zhang, E.; Chen, Y.; Liu, L.; Rong, D.; Liu, Q.; Li, W. Pinus massoniana bark extract inhibits migration of the lung cancer A549 cell line. Oncol. Lett. 2017, 13, 1019–1023. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Bai, J.; Jiang, G.; Li, X.; Wang, J.; Wu, D.; Owusu, L.; Zhang, E.; Li, W. Anti-tumor effect of Pinus massoniana bark proanthocyanidins on ovarian cancer through induction of cell apoptosis and inhibition of cell migration. PLoS ONE 2015, 10, e0142157. [Google Scholar] [CrossRef]
- Nair, H.K.; Rao, K.V.; Aalinkeel, R.; Mahajan, S.; Chawda, R.; Schwartz, S.A. Inhibition of prostate cancer cell colony formation by the flavonoid quercetin correlates with modulation of specific regulatory genes. Clin. Diagn. Lab. Immunol. 2004, 11, 63–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalilzadeh, N.; Samadi, N.; Salehi, R.; Dehghan, G.; Iranshahi, M.; Dadpour, M.R.; Hamishehkar, H. Novel nano-vehicle for delivery and efficiency of anticancer auraptene against colon cancer cells. Sci. Rep. 2020, 10, 1606. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Lee, Y.H.; Lee, K.H.; Lee, B.S.; Alishir, A.; Ko, Y.-J.; Kang, K.S.; Kim, K.H. Aviculin isolated from Lespedeza cuneata induce apoptosis in breast cancer cells through mitochondria-mediated caspase activation pathway. Molecules 2020, 25, 1708. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Lee, S.R.; Kang, K.S.; Ko, Y.; Pang, C.; Yamabe, N.; Kim, K.H. Betulinic acid suppresses ovarian cancer cell proliferation through induction of apoptosis. Biomolecules 2019, 9, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Lai, F.; Xie, H.; Wang, J.; Hongbin, W. Involvement of the Bcl-2 family members in pinus massoniana bark extract induced apoptosis in HeLa cells. Phytother. Res. 2008, 22, 1472–1476. [Google Scholar] [CrossRef] [PubMed]
- Demidenko, Z.; Kalurupalle, S.; Hanko, C.; Lim, C.; Broude, E.; Blagosklonny, M. Mechanism of G1-like arrest by low concentrations of paclitaxel: Next cell cycle p53-dependent arrest with sub G1 DNA content mediated by prolonged mitosis. Oncogene 2008, 27, 4402–4410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pflaum, J.; Schlosser, S.; Müller, M. P53 family and cellular stress responses in cancer. Front. Oncol. 2014, 4, 285. [Google Scholar] [CrossRef]
- Cadwell, C.; Zambetti, G.P. The effects of wild-type p53 tumor suppressor activity and mutant p53 gain-of-function on cell growth. Gene 2001, 277, 15–30. [Google Scholar] [CrossRef]
- Lu, Z.; Xiao, Y.; Liu, X.; Zhang, Z.; Xiao, F.; Bi, Y. Matrine reduces the proliferation of A549 cells via the p53/p21/PCNA/eIF4E signaling pathway. Mol. Med. Rep. 2017, 15, 2415–2422. [Google Scholar] [CrossRef] [Green Version]
- Festjens, N.; Van Gurp, M.; Van Loo, G.; Saelens, X.; Vandenabeele, P. Bcl-2 family members as sentinels of cellular integrity and role of mitochondrial intermembrane space proteins in apoptotic cell death. Acta Haematol. 2004, 111, 7–27. [Google Scholar] [CrossRef]
- Giotakis, A.I.; Kontos, C.K.; Manolopoulos, L.D.; Sismanis, A.; Konstadoulakis, M.M.; Scorilas, A. High BAX/BCL2 mRNA ratio predicts favorable prognosis in laryngeal squamous cell carcinoma, particularly in patients with negative lymph nodes at the time of diagnosis. Clin. Biochem. 2016, 49, 890–896. [Google Scholar] [CrossRef]
- Jo, J.R.; Park, J.S.; Park, Y.K.; Chae, Y.Z.; Lee, G.H.; Park, G.Y.; Jang, B.C. Pinus densiflora leaf essential oil induces apoptosis via ROS generation and activation of caspases in YD-8 human oral cancer cells. Int. J. Oncol. 2012, 40, 1238–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazebnik, Y.A.; Kaufmann, S.H.; Desnoyers, S.; Poirier, G.G.; Earnshaw, W.C. Cleavage of poly(ADP-ribose) polymerase by a proteinase with properties like ICE. Nature 1994, 371, 346–347. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Chen, H.M.; Shiau, C.Y. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int. 2003, 36, 949–957. [Google Scholar] [CrossRef]
- Molavi, O.; Torkzaban, F.; Jafari, S.; Asnaashari, S.; Asgharian, P. Chemical compositions and anti-proliferative activity of the aerial parts and rhizomes of squirting cucumber, Cucurbitaceae. Jundishapur J. Nat. Pharm. Prod. 2020, 15. [Google Scholar] [CrossRef]
- Lee, S.; Ryoo, R.; Choi, J.H.; Kim, J.-H.; Kim, S.-H.; Kim, K.H. Trichothecene and tremulane sesquiterpenes from a hallucinogenic mushroom Gymnopilus junonius and their cytotoxicity. Arch. Pharmacal. Res. 2020, 43, 214–223. [Google Scholar] [CrossRef]
- Trinh, T.A.; Park, E.-J.; Lee, D.; Song, J.H.; Lee, H.L.; Kim, K.H.; Kim, Y.; Jung, K.; Kang, K.S.; Yoo, J.-E. Estrogenic activity of sanguiin H-6 through activation of estrogen receptor α coactivator-binding site. Nat. Prod. Sci. 2019, 25, 28–33. [Google Scholar] [CrossRef]
- Lee, S.R.; Kang, H.; Yoo, M.J.; Yu, J.S.; Lee, S.; Yi, S.A.; Beemelmanns, C.; Lee, J.; Kim, K.H. Anti-adipogenic pregnane steroid from a Hydractinia-associated fungus, Cladosporium sphaerospermum SW67. Nat. Prod. Sci. 2020, 26, 230–235. [Google Scholar]
- Lee, K.; Seo, Y.-J.; Song, J.-H.; Chei, S.; Lee, B.-Y. Ginsenoside Rg1 promotes browning by inducing UCP1 expression and mitochondrial activity in 3T3-L1 and subcutaneous white adipocytes. J. Ginseng Res. 2019, 43, 589–599. [Google Scholar] [CrossRef]
- Sabzichi, M.; Ramezani, M.; Mohammadian, J.; Ghorbani, M.; Mardomi, A.; Najafipour, F.; Mehdizadeh, A. The synergistic impact of quinacrine on cell cycle and anti-invasiveness behaviors of doxorubicin in MDA-MB-231 breast cancer cells. Process Biochem. 2019, 81, 175–181. [Google Scholar] [CrossRef]
- Mahmoudi, S.; Ghorbani, M.; Sabzichi, M.; Ramezani, F.; Hamishehkar, H.; Samadi, N. Targeted hyaluronic acid-based lipid nanoparticle for apigenin delivery to induce Nrf2-dependent apoptosis in lung cancer cells. J. Drug Deliv. Sci. Technol. 2019, 49, 268–276. [Google Scholar] [CrossRef]
- Grada, A.; Otero-Vinas, M.; Prieto-Castrillo, F.; Obagi, Z.; Falanga, V. Research Techniques Made Simple: Analysis of Collective Cell Migration Using the Wound Healing Assay. J. Investig. Dermatol. 2017, 137, e11–e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousefi, H.; Karimi, P.; Alihemmati, A.; Reza Alipour, M.; Habibi, P.; Ahmadiasl, N. Therapeutic potential of genistein in ovariectomy-induced pancreatic injury in diabetic rats: The regulation of mapk pathway and apoptosis. Iran. J. Basic Med. Sci. 2017, 20, 1009–1015. [Google Scholar] [PubMed]
- Alimova, I.N.; Liu, B.; Fan, Z.; Edgerton, S.M.; Dillon, T.; Lind, S.E.; Thor, A.D. Metformin inhibits breast cancer cell growth, colony formation and induces cell cycle arrest in vitro. Cell Cycle 2009, 8, 909–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghaffari, T.; Asnaashari, S.; Irannejad, E.; Delazar, A.; Farajnia, S.; Hong, J.-H.; Pang, C.; Hamishehkar, H.; Kim, K.H. Comparative Evaluation of Apoptosis Induction Using Needles, Bark, and Pollen Extracts and Essential Oils of Pinus eldarica in Lung Cancer Cells. Appl. Sci. 2021, 11, 5763. https://0-doi-org.brum.beds.ac.uk/10.3390/app11135763
Ghaffari T, Asnaashari S, Irannejad E, Delazar A, Farajnia S, Hong J-H, Pang C, Hamishehkar H, Kim KH. Comparative Evaluation of Apoptosis Induction Using Needles, Bark, and Pollen Extracts and Essential Oils of Pinus eldarica in Lung Cancer Cells. Applied Sciences. 2021; 11(13):5763. https://0-doi-org.brum.beds.ac.uk/10.3390/app11135763
Chicago/Turabian StyleGhaffari, Tayyebeh, Solmaz Asnaashari, Ebrahim Irannejad, Abbas Delazar, Safar Farajnia, Joo-Hyun Hong, Changhyun Pang, Hamed Hamishehkar, and Ki Hyun Kim. 2021. "Comparative Evaluation of Apoptosis Induction Using Needles, Bark, and Pollen Extracts and Essential Oils of Pinus eldarica in Lung Cancer Cells" Applied Sciences 11, no. 13: 5763. https://0-doi-org.brum.beds.ac.uk/10.3390/app11135763