
Lactobacillus plantarum SK15 as a Starter Culture for Prevention of Biogenic Amine Accumulation in Fermented Beverage Containing Hericium erinaceus Mushroom

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.2. Starter Culture Preparation
2.3. Evaluation of L. plantarum SK15 on Amino Acid Decarboxylase Production
2.4. Fermented Mushroom Beverage (FMB) Preparation
2.5. Determination of Biogenic Amines
2.6. Determination of pH and Total Acidity
2.7. Determination of Methanol and Ethanol
2.8. Determination of Amino Acids
2.9. Determination of Reducing Sugar
2.10. Determination of Total Phenolic Content (TPC)
2.11. Determination of Antioxidant Activity
2.12. Enumeration of Microbial Population
2.13. Statistics Analysis
3. Results
3.1. Evaluation of L. plantarum SK15 on Amino Acid Decarboxylase Production
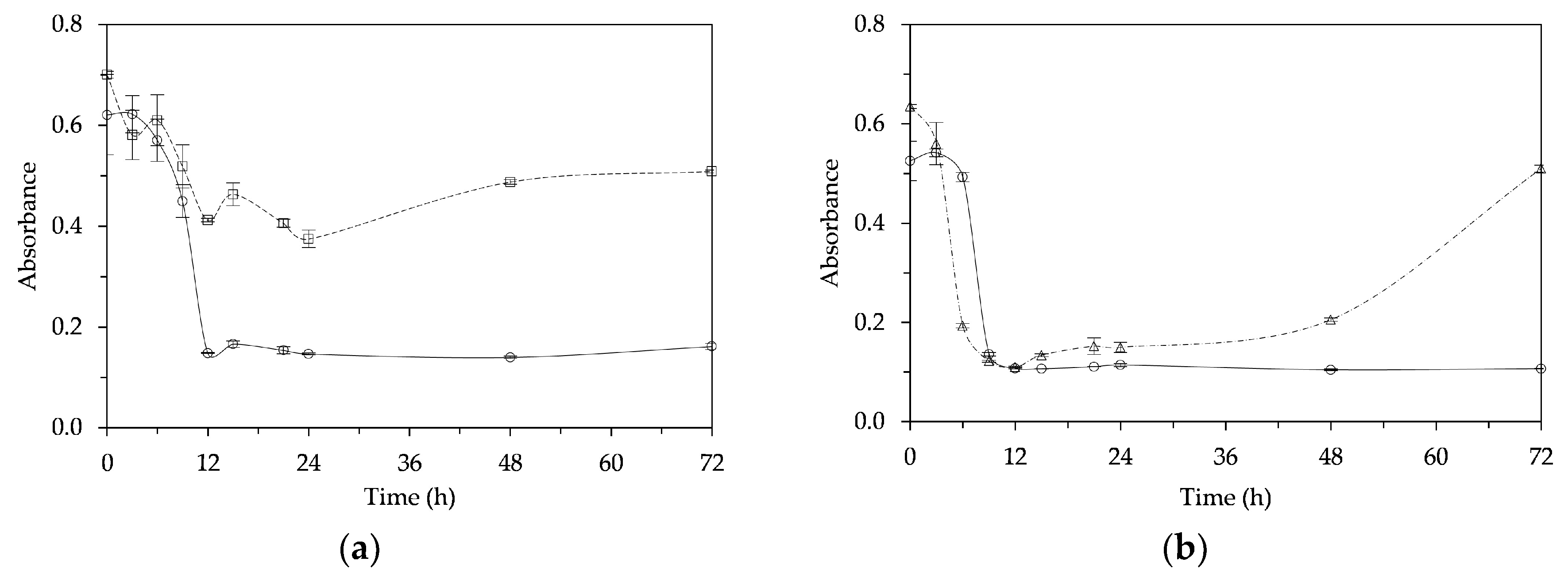
3.2. Physical Observations of a Fermented Mushroom Beverage (FMB)
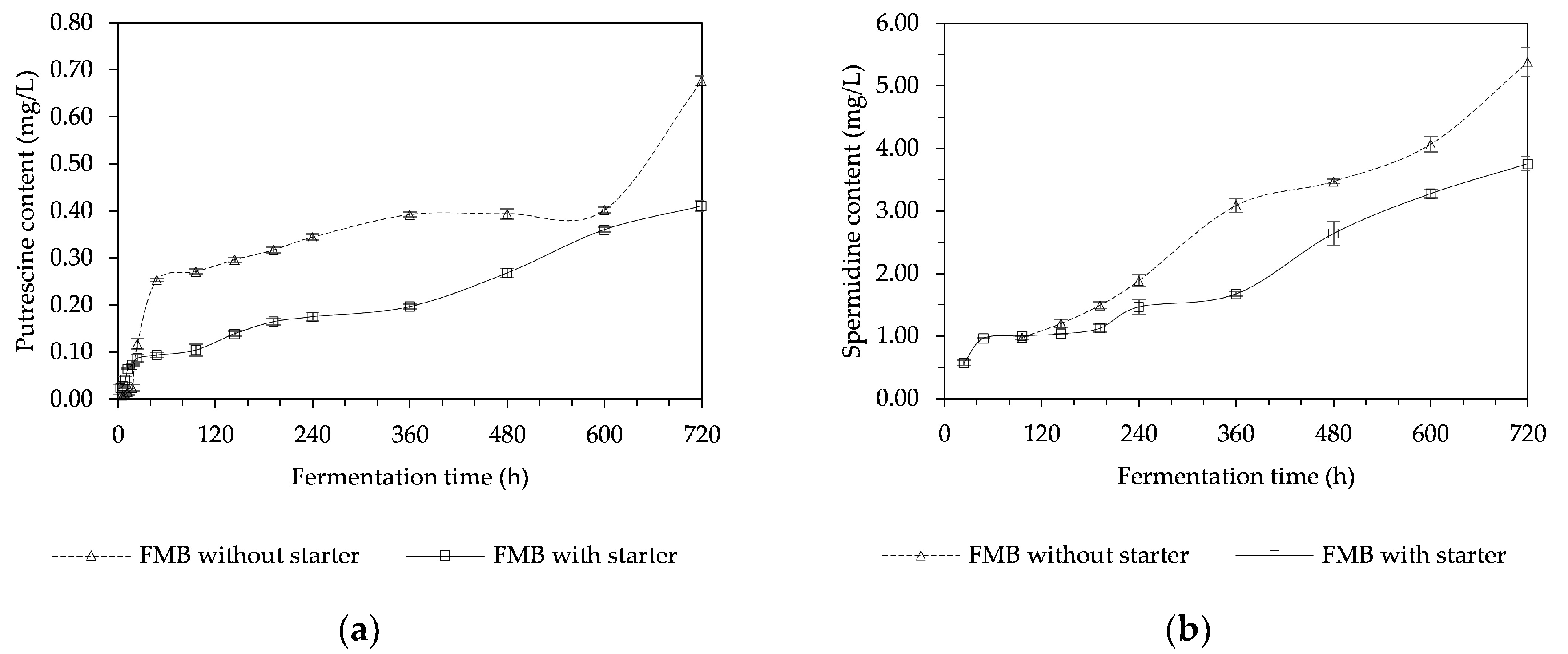
3.3. Biogenic Amines Content of a Fermented Mushroom Beverage (FMB)
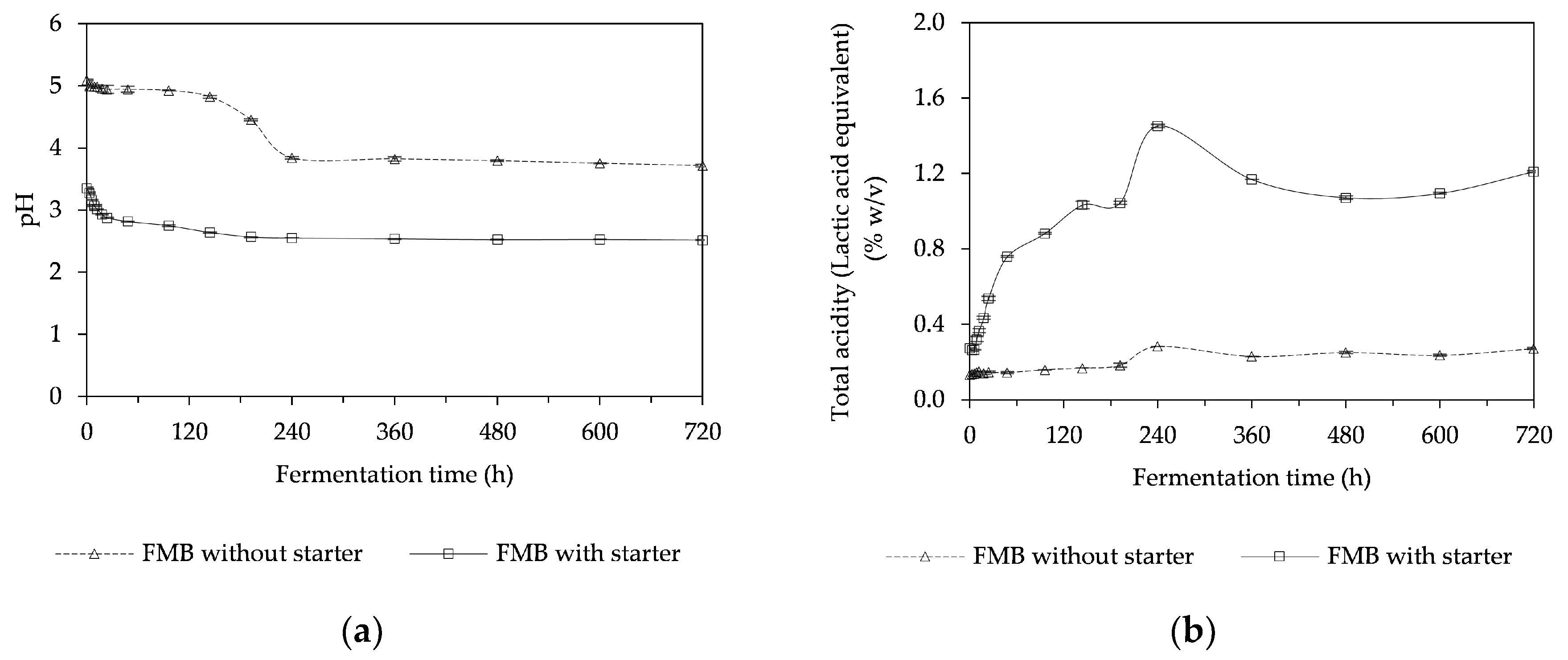
3.4. pH and Total Acidity of a Fermented Mushroom Beverage (FMB)
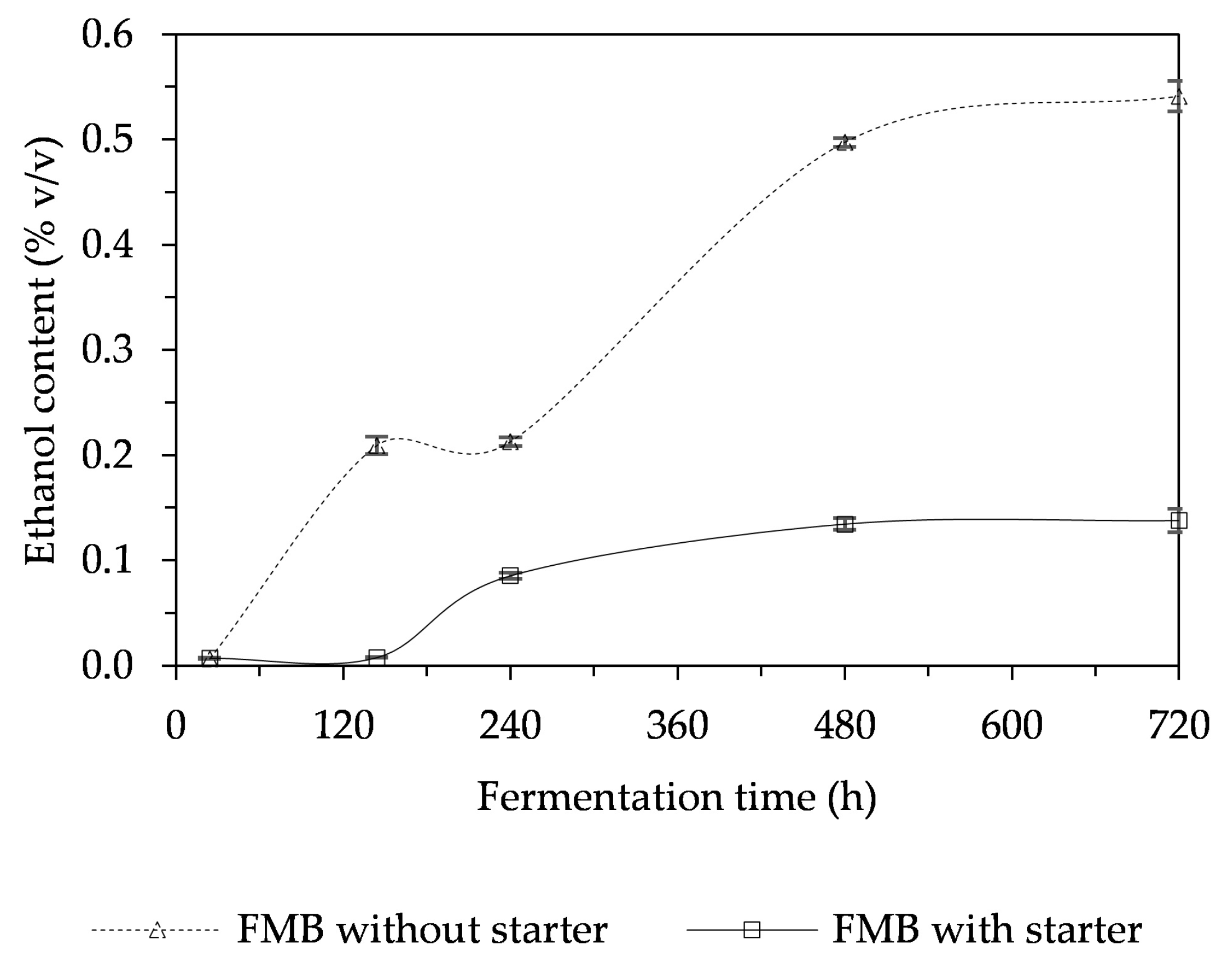
3.5. Alcohol Content in a Fermented Mushroom Beverage (FMB)
3.6. Amino Acid Content of a Fermented Mushroom Beverage (FMB)
3.7. Reducing Sugar Content of a Fermented Mushroom Beverage (FMB)
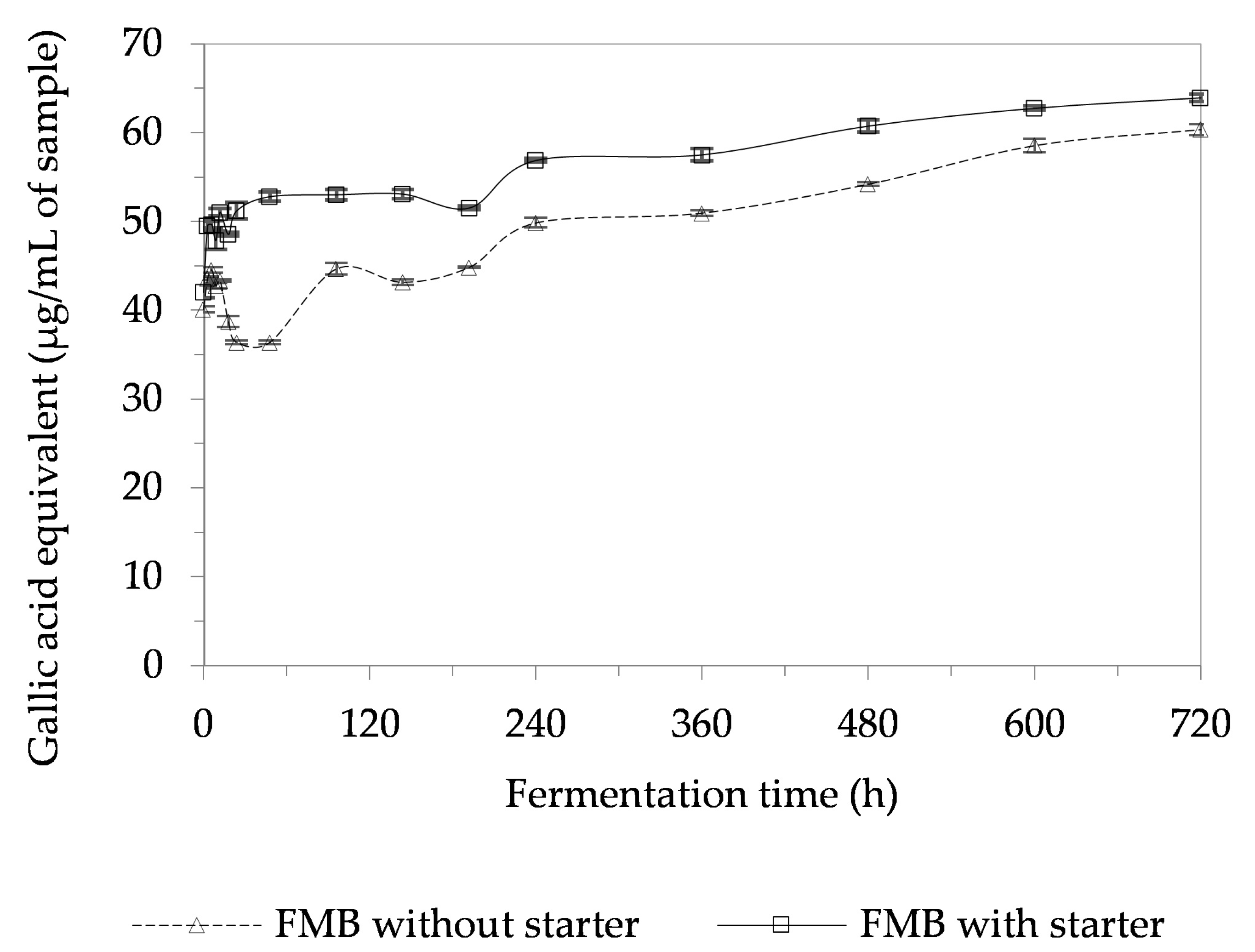
3.8. Total Phenolic Content (TPC) of a Fermented Mushroom Beverage (FMB)
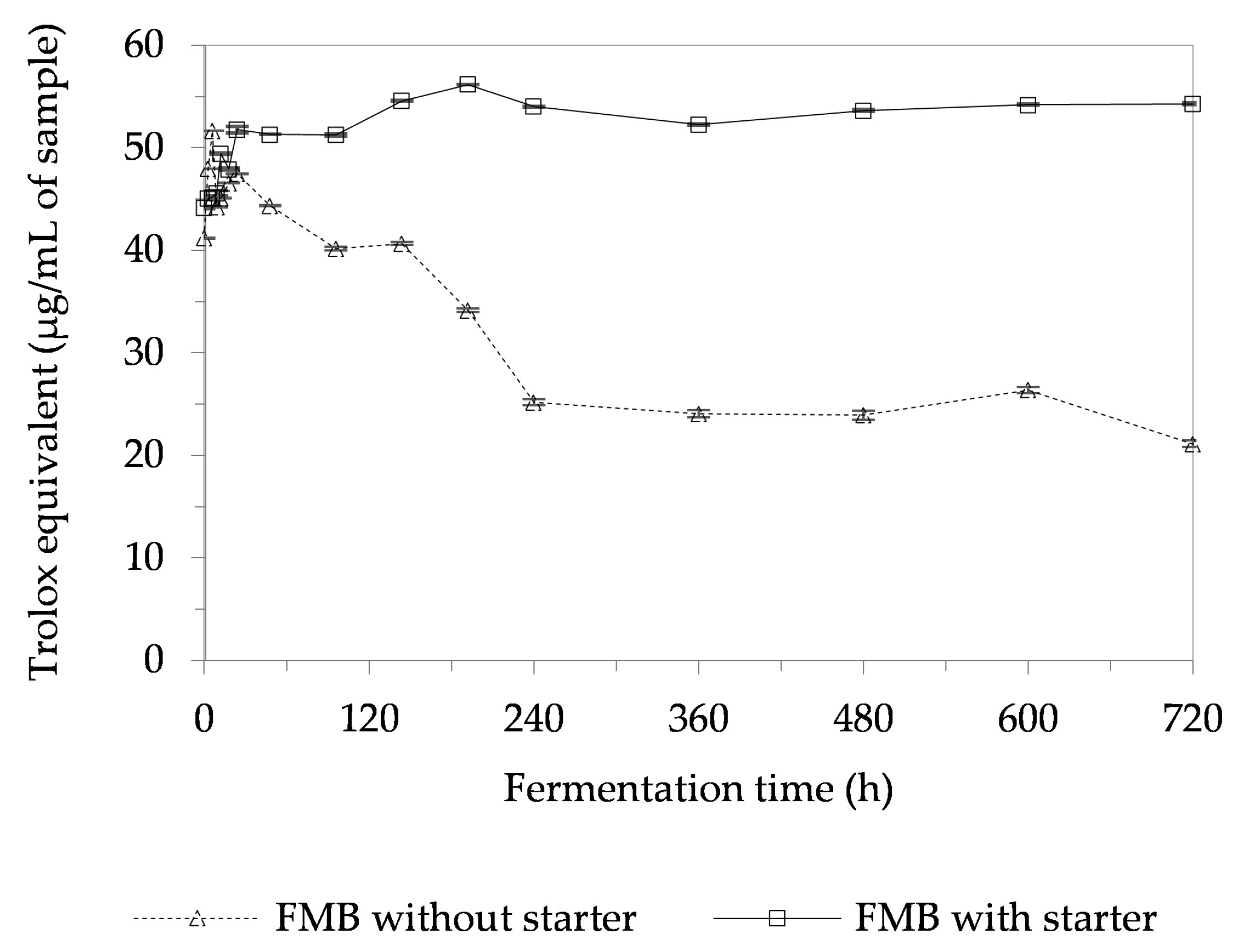
3.9. Antioxidant Activity of a Fermented Mushroom Beverage (FMB)
3.10. Microbial Load of a Fermented Mushroom Beverage (FMB)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, M.; Tania, M.; Liu, R.; Rahman, M.M. Hericium erinaceus: An edible mushroom with medicinal values. J. Complement. Integr. Med. 2013, 10, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.M.; Freitas, A.C.; Rocha-Santos, T.A.; Vasconcelos, M.W.; Roriz, M.; Rodríguez-Alcalá, L.M.; Gomes, A.M.; Duarte, A.C. Chemical composition and nutritive value of Pleurotus citrinopileatus var cornucopiae, P. eryngii, P. salmoneo stramineus, Pholiota nameko and Hericium erinaceus. J. Food Sci. Technol. 2015, 52, 6927–6939. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry nutrition and health-promoting properties of Hericium erinaceus (lion’s mane) mushroom fruiting bodies and mycelia and their bioactive compounds. J. Agric. Food Chem. 2015, 63, 7108–7123. [Google Scholar] [CrossRef]
- Chaiyasut, C.; Makhamrueang, N.; Peerajan, S.; Sivamaruthi, B. Assessment of organic acid content, and brix value of representative indigenous fermented plant beverages of Thailand. Assessment 2017, 10, 350–354. [Google Scholar] [CrossRef]
- Shalaby, A.R. Significance of biogenic amines to food safety and human health. Food Res. Int. 1996, 29, 675–690. [Google Scholar] [CrossRef]
- Mohedano, M.L.; Spano, G.; Spano, G.; López, P.; Russo, P. Controlling the formation of biogenic amines in fermented foods. In Advances in Fermented Foods and Beverages, 1st ed.; Holzapfel, W., Ed.; Woodhead Publishing: Cambridge, UK, 2014; pp. 273–310. [Google Scholar]
- Dadáková, E.; Pelikánová, T.; Kalač, P. Content of biogenic amines and polyamines in some species of European wild-growing edible mushrooms. Eur. Food Res. Technol. 2009, 230, 163–171. [Google Scholar] [CrossRef]
- Moret, S.; Smela, D.; Populin, T.; Conte, L.S. A survey on free biogenic amine content of fresh and preserved vegetables. Food Chem. 2005, 89, 355–361. [Google Scholar] [CrossRef]
- Papageorgiou, M.; Lambropoulou, D.; Morrison, C.; Kłodzinska, E.; Namiesnik, J.; Płotka-Wasylka, J. Literature update of analytical methods for biogenic amines determination in food and beverages. Trends Anal. Chem. 2018, 98, 128–142. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific opinion on risk based control of biogenic amine formation in fermented foods. EFSA J. 2011, 9, 2393. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, M.A.; Moreno-Arribas, M.V. The problem of biogenic amines in fermented foods and the use of potential biogenic amine-degrading microorganisms as a solution. Trends Food Sci. Technol. 2014, 39, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Visciano, P.; Schirone, M.; Paparella, A. An overview of histamine and other biogenic amines in fish and fish products. Foods 2020, 9, 1795. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Capillas, C.; Herrero, A.M. Impact of biogenic amines on food quality and safety. Foods 2019, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.S. Biogenic amines: Their importance in foods. Int. J. Food Microbiol. 1996, 29, 213–231. [Google Scholar] [CrossRef]
- Gardini, F.; Özogul, Y.; Suzzi, G.; Tabanelli, G.; Özogul, F. Technological factors affecting biogenic amine content in foods: A review. Front. Microbiol. 2016, 7, 1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasini, F.; Soglia, F.; Petracci, M.; Caboni, M.F.; Marziali, S.; Montanari, C.; Gardini, F.; Grazia, L.; Tabanelli, G. Effect of fermentation with different lactic acid bacteria starter cultures on biogenic amine content and ripening patterns in dry fermented sausages. Nutrients 2018, 10, 1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, C.; Guerin, M.; Souidi, K.; Remize, F. Lactic fermented fruit or vegetable juices: Past, present and future. Beverages 2020, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Chen, Q.; Li, F.; Zheng, D.; Kong, B. Biogenic amine inhibition and quality protection of Harbin dry sausages by inoculation with Staphylococcus xylosus and Lactobacillus plantarum. Food Control 2016, 68, 358–366. [Google Scholar] [CrossRef]
- Naila, A.; Flint, S.; Fletcher, G.; Bremer, P.; Meerdink, G. Control of biogenic amines in food—existing and emerging approaches. J. Food Sci. 2010, 75, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.C.; Kung, H.F.; Huang, Y.L.; Wu, C.H.; Huang, Y.R.; Tsai, Y.H. Reduction of biogenic amines during miso fermentation by Lactobacillus plantarum as a starter culture. J. Food Prot. 2016, 79, 1556–1561. [Google Scholar] [CrossRef] [PubMed]
- Bover-Cid, S.; Izquierdo-Pulido, M.; Vidal-Carou, M.C. Effectiveness of a Lactobacillus sakei starter culture in the reduction of biogenic amine accumulation as a function of the raw material quality. J. Food Prot. 2001, 64, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zheng, J.; Huang, J.; Zhou, R. Reduced nitrite and biogenic amine concentrations and improved flavor components of Chinese sauerkraut via co-culture of Lactobacillus plantarum and Zygosaccharomyces rouxii. Ann. Microbiol. 2014, 64, 847–857. [Google Scholar] [CrossRef]
- Woraharn, S.; Lailerd, N.; Sivamaruthi, B.S.; Wangcharoen, W.; Sirisattha, S.; Peerajan, S.; Chaiyasut, C. Evaluation of factors that influence the l-glutamic and γ-aminobutyric acid production during Hericium erinaceus fermentation by lactic acid bacteria. CyTA J. Food. 2016, 14, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Chaiyasut, C.; Woraharn, S.; Sivamaruthi, B.S.; Lailerd, N.; Kesika, P.; Peerajan, S. Lactobacillus fermentum HP3–mediated fermented Hericium erinaceus juice as a health promoting food supplement to manage diabetes mellitus. J. Evid.-Based Integr. Med. 2018, 23, 2515690X18765699. [Google Scholar] [CrossRef] [Green Version]
- Makhamrueang, N.; Sirilun, S.; Sirithunyalug, J.; Chaiyana, W.; Wangcharoen, W.; Peerajan, S.; Chaiyasut, C. Effect of pretreatment processes on biogenic amines content and some bioactive compounds in Hericium erinaceus extract. Foods 2021, 10, 996. [Google Scholar] [CrossRef]
- Saelee, M.; Sivamaruthi, B.S.; Sirilun, S.; Kesika, P.; Peerajan, S.; Chaiyasut, C. Effect of green tea extract during lactic acid bacteria mediated fermentation of Morinda citrifolia Linn. (Noni) fruit juice. Pak. J. Biol. Sci. 2019, 22, 486–493. [Google Scholar] [CrossRef] [Green Version]
- Falkow, S. Activity of lysine decarboxylase as an aid in the identification of Salmonellae and Shigellae. Am. J. Clin. Pathol. 1958, 29, 598–600. [Google Scholar] [CrossRef] [PubMed]
- Chaiyasut, C.; Jantavong, S.; Kruatama, C.; Peerajan, S.; Sirilun, S.; Shank, L. Factors affecting methanol content of fermented plant beverage containing Morinda citrifolia. Afr. J. Biotechnol. 2013, 12, 4356–4363. [Google Scholar]
- Kıvrak, İ.; Kıvrak, Ş.; Harmandar, M. Free amino acid profiling in the giant puffball mushroom (Calvatia gigantea) using UPLC–MS/MS. Food Chem. 2014, 158, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, C.; Rodriguez-Jasso, R.M.; Gomes, N.; Teixeira, J.A.; Belo, I. Adaptation of dinitrosalicylic acid method to microtiter plates. Anal. Methods 2010, 2, 2046–2048. [Google Scholar] [CrossRef] [Green Version]
- Adebo, O.A.; Njobeh, P.B.; Kayitesi, E. Fermentation by Lactobacillus fermentum strains (singly and in combination) enhances the properties of ting from two whole grain sorghum types. J. Cereal Sci. 2018, 82, 49–56. [Google Scholar] [CrossRef]
- Pumtes, P.; Rojsuntornkitti, K.; Kongbangkerd, T.; Jittrepotch, N. Effects of different extracting conditions on antioxidant activities of Pleurotus flabellatus. Int. Food Res. J. 2016, 23, 173–179. [Google Scholar]
- Sirilun, S.; Sivamaruthi, B.S.; Kesika, P.; Makhamrueang, N.; Chaiyasut, K.; Peerajan, S.; Chaiyasut, C. Development and evaluation of Mustard green pickled liquid as starter for Morinda citrifolia Linn. Fermentation. Int. Food Res. J. 2017, 24, 2170–2176. [Google Scholar]
- Arena, M.E.; Manca de Nadra, M.C. Biogenic amine production by Lactobacillus. J. Appl. Microbiol. 2001, 90, 158–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaman, M.Z.; Bakar, F.A.; Jinap, S.; Bakar, J. Novel starter cultures to inhibit biogenic amines accumulation during fish sauce fermentation. Int. J. Food Microbiol. 2011, 145, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Mah, J.H.; Hwang, H.J. Inhibition of biogenic amine formation in a salted and fermented anchovy by Staphylococcus xylosus as a protective culture. Food Control 2009, 20, 796–801. [Google Scholar] [CrossRef]
- Limsuwan, S.; Visessanguan, W.; Kongkiattikajorn, J. The effect of starter cultures on biogenic amines and free amino acid contents in Nham during fermentation. Kasetsart J. Nat. Sci. 2007, 41, 205–212. [Google Scholar]
- Thai Community Standard Product (TCSP). Thai Community Standard Product No. 481. Thai Industrial Standards Institute (TISI), 2007. Available online: https://www.tisi.go.th/home/en (accessed on 15 June 2021).
- Peerajan, S.; Chaiyasut, C.; Sirilun, S.; Chaiyasut, K.; Kesika, P.; Sivamaruthi, B.S. Enrichment of nutritional value of Phyllanthus emblica fruit juice using the probiotic bacterium, Lactobacillus paracasei HII01 mediated fermentation. Food Sci. Technol. 2016, 36, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Kraut, J.A.; Kurtz, I. Toxic alcohol ingestions: Clinical features, diagnosis, and management. J. Am. Soc. Nephrol. 2008, 3, 208–225. [Google Scholar] [CrossRef] [PubMed]
- Poojary, M.M.; Orlien, V.; Passamonti, P.; Olsen, K. Enzyme-assisted extraction enhancing the umami taste amino acids recovery from several cultivated mushrooms. Food Chem. 2017, 234, 236–244. [Google Scholar] [CrossRef]
- Michlmayr, H.; Kneifel, W. β-Glucosidase activities of lactic acid bacteria: Mechanisms, impact on fermented food and human health. FEMS Microbiol. Lett. 2014, 352, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiranmai, E.; Suneetha, A.; Sethi, N. Preparation of tamarind jam blended with mango. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3365–3373. [Google Scholar] [CrossRef]
- Bautista-Expósito, S.; Tomé-Sánchez, I.; Martín-Diana, A.B.; Frias, J.; Peñas, E.; Rico, D.; Martínez-Villaluenga, C. Enzyme eelection and hydrolysis under optimal conditions improved phenolic acid solubility, and antioxidant and anti-inflammatory activities of wheat bran. Antioxidants 2020, 9, 984. [Google Scholar] [CrossRef] [PubMed]
- Tiram, C. Effect of different drying techniques on the nutritional values of oyster mushroom (Pleurotus sajor-caju). Sains Malays. 2013, 42, 937–941. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Amino Acid Content 1 (µg/L) | |||
|---|---|---|---|---|
| 0 h | 720 h | |||
| Without Starter | With Starter | Without Starter | With Starter | |
| Alanine | 138.65 ± 2.03 c | 133.21 ± 1.84 b | 145.03 ± 2.21 d | 69.60 ± 1.06 a |
| Arginine | 22.95 ± 1.72 a | 23.30 ± 1.50 a | 28.97 ± 1.97 b | 23.40 ± 1.45 a |
| Aspartic acid | 10.78 ± 2.85 b | 9.91 ± 0.87 ab | 11.86 ± 1.99 b | 6.05 ± 0.26 a |
| Cysteine | ND | ND | ND | ND |
| Glutamic acid | 65.09 ± 2.97 b | 56.85 ± 2.52 a | 69.28 ± 0.85 b | 54.78 ± 1.23 a |
| Glycine | 14.13 ± 0.84 a | 17.58 ± 1.37 ab | 19.40 ± 2.38 b | 20.54 ± 0.96 b |
| Histidine | 2.25 ± 0.20 b | 2.68 ± 0.14 b | 3.22 ± 0.24 c | 1.39 ± 0.10 a |
| Isoleucine | 173.99 ± 2.08 b | 176.22 ± 2.28 b | 178.86 ± 2.95 b | 165.44 ± 1.55 a |
| Leucine | 184.96 ± 2.87 b | 192.88 ± 2.08 c | 190.69 ± 1.76 c | 177.79 ± 1.64 a |
| Lysine | ND | ND | ND | ND |
| Methionine | ND | ND | ND | ND |
| Phenylalanine | 101.81 ± 2.73 b | 102.17 ± 1.59 b | 108.62 ± 1.17 c | 56.51 ± 1.55 a |
| Proline | 38.01 ± 0.89 a | 39.53 ± 0.59 ab | 42.07 ± 1.72 b | 47.50 ± 1.42 c |
| Serine | 7.24 ± 0.24 c | 5.72 ± 0.44 b | 5.29 ± 0.36 b | 2.30 ± 0.28 a |
| Tyrosine | 50.81 ± 1.11 c | 46.26 ± 0.47 b | 57.62 ± 1.07 d | 16.70 ± 0.55 a |
| Threonine | 18.62 ± 0.47 ab | 19.73 ± 2.00 b | 24.54 ± 1.15 c | 16.17 ± 0.98 a |
| Valine | 162.06 ± 1.51 a | 172.04 ± 3.48 b | 173.83 ± 0.93 b | 187.65 ± 0.91 c |
| Total | 991.35 ± 22.49 b | 998.05 ± 21.16 b | 1059.27 ± 20.75 c | 845.80 ± 13.93 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makhamrueang, N.; Sirilun, S.; Sirithunyalug, J.; Chaiyana, W.; Wangcharoen, W.; Peerajan, S.; Chaiyasut, C. Lactobacillus plantarum SK15 as a Starter Culture for Prevention of Biogenic Amine Accumulation in Fermented Beverage Containing Hericium erinaceus Mushroom. Appl. Sci. 2021, 11, 6680. https://0-doi-org.brum.beds.ac.uk/10.3390/app11156680
Makhamrueang N, Sirilun S, Sirithunyalug J, Chaiyana W, Wangcharoen W, Peerajan S, Chaiyasut C. Lactobacillus plantarum SK15 as a Starter Culture for Prevention of Biogenic Amine Accumulation in Fermented Beverage Containing Hericium erinaceus Mushroom. Applied Sciences. 2021; 11(15):6680. https://0-doi-org.brum.beds.ac.uk/10.3390/app11156680
Chicago/Turabian StyleMakhamrueang, Netnapa, Sasithorn Sirilun, Jakkapan Sirithunyalug, Wantida Chaiyana, Wiwat Wangcharoen, Sartjin Peerajan, and Chaiyavat Chaiyasut. 2021. "Lactobacillus plantarum SK15 as a Starter Culture for Prevention of Biogenic Amine Accumulation in Fermented Beverage Containing Hericium erinaceus Mushroom" Applied Sciences 11, no. 15: 6680. https://0-doi-org.brum.beds.ac.uk/10.3390/app11156680