
Environmental Conditions and Agronomical Factors Influencing the Levels of Phytochemicals in Brassica Vegetables Responsible for Nutritional and Sensorial Properties


 ,
,  and
and
Abstract
:1. Introduction
2. Nutritional Quality
2.1. Antioxidant Compounds
2.1.1. Water-Soluble Antioxidants
- Phenolic compounds
- Flavonols: together with anthocyanins, they are the main represented flavonoids in Brassica species; they can be found in internal and external parts of leaves, seeds, shoots and sprouts leaves [22,23]. The most represented flavonols in Brassica vegetables are quercetin (up to 23 mg/100 g fresh product in kale), kaempferol (up to 47 mg/100 g fresh product in kale) and isorhamnetin (up to 24 mg/100 g fresh product in kale) [24]. Quercetin, found mainly in kale, is characterized by a strong antioxidant power (higher than vitamin C); it exerts its activity against free oxygen radicals and acts on the prevention of cardiovascular diseases and cancer, atherosclerosis and chronic inflammation, and the induction of enzymes that detoxify carcinogens [25,26]. Kaempferol 3-O-sophoroside is the main represented flavonol in broccoli florets; its high intake is linked with a lower risk of coronary heart disease [27]. Kaempferol and quercetin, and in less amounts, myricetin, are the main represented flavonols in B. rapa subsp. sylvestris.
- Flavones: Apigenin and luteolin are the only flavones detected in hydrolysed extracts of different Brassica vegetables (up to 45 and 12 µg/g of fresh weight in Chinese cabbage, respectively), excluding broccoli, where they were not detected [28].
- Anthocyanins were detected in Brassica vegetables and described by several authors [13,29]. They are present only in bright coloured species and varieties with red, orange and purple pigmentation, such as some kales, purple broccoli, and red and black cabbage. These compounds show an interesting antioxidant activity. The 80% of anthocyanins present in Brassica species are in the acylated form, more stable and easily absorbable by the organism. The main represented anthocyanins in cruciferous are cyanidin derivatives. In particular, red cabbage possesses eight main types of anthocyanins (for a total of up to 190 mg Cyanidin−3-Glucoside equivalents/100 g of fresh weight) [30]; cyanidin−3-diglucoside is the most represented [31]. In broccoli, more than 17 anthocyanins were detected [29].
- Vitamin C and vitamin B9 (Folic Acid)
2.1.2. Lipo-Soluble Antioxidants
- Carotenoids
- Vitamin E
2.2. Micro- and Macro-Elements
2.3. Glucosinolates (GLS) and Isothiocyanates (ITCS)
3. Sensorial Quality
- Firmness, which indicates the resistance of vegetables to mechanical damages; it assumes a great importance during the post-harvest management.
- Colour, which indicates the freshness of the product and the quality of the storage conditions; it visually attracts the consumers [76].
- Sweetness, which is linked to the presence of glucose, fructose, and sucrose, and provides the sweet sensation to the consumer.
- Acidity, which indicates the acid sensation that the product stimulates in the consumer.
3.1. Brassica Aroma
3.2. Brassica Taste
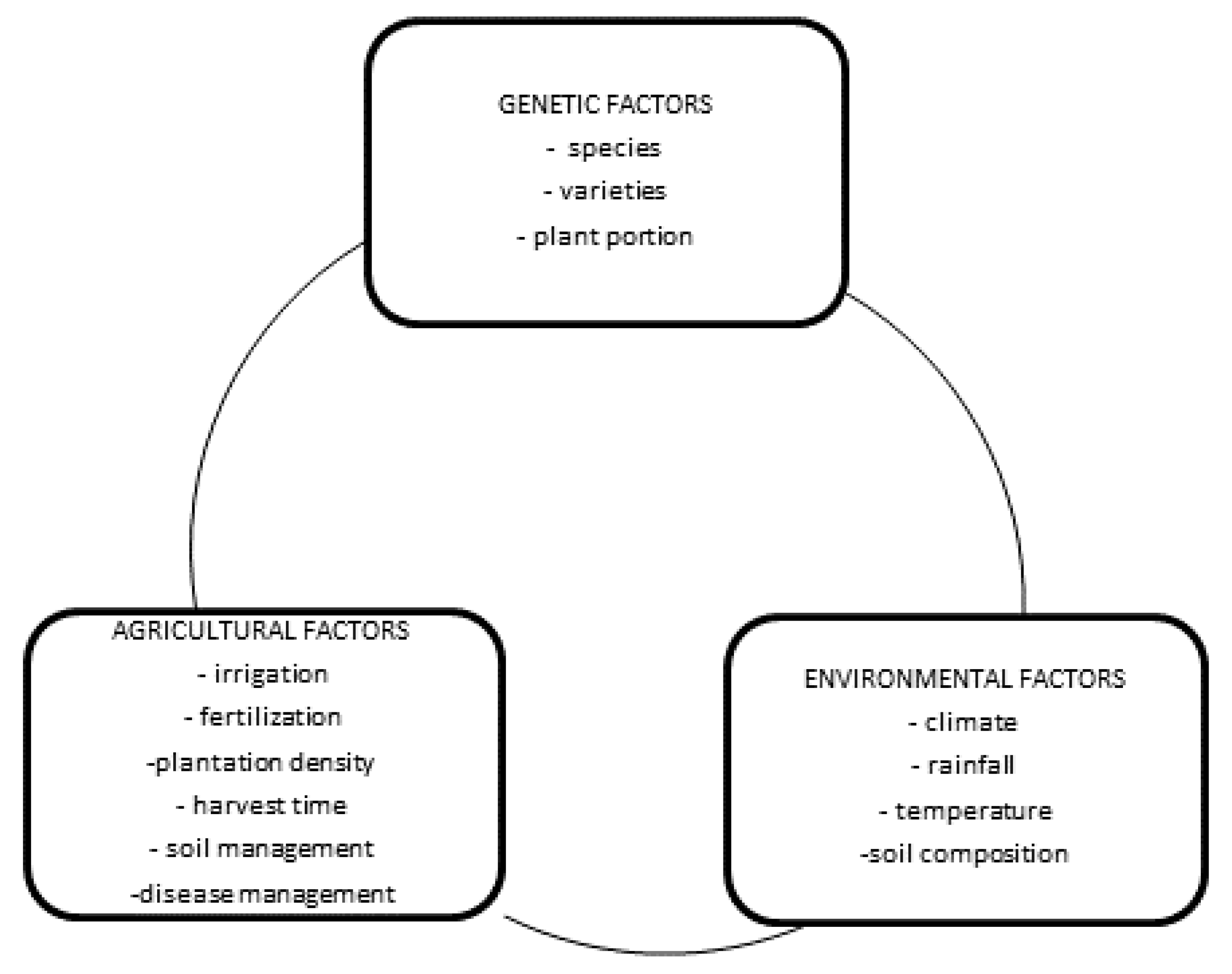
4. Factors Influencing the Phytochemical Compounds of Brassica Vegetables
4.1. Genetic Factors
4.1.1. Brassica Oleracea Species
4.1.2. Brassica rapa Species and Other Cruciferous Crops
4.1.3. Plant Portion and Plant Developmental Stage
4.2. Environmental and Agricultural Factors
4.2.1. Cultivation System and Soil Composition
4.2.2. Water Stress
4.2.3. Plant Density, Intercropping and Trap Cropping
4.2.4. Fertilization Practices
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Dauchet, L.; Amouyel, P.; Hercberg, S.; Dallongeville, J. Fruit and vegetable consumption and risk of coronary heart disease: A meta-analysis of cohort studies. J. Nutr. 2006, 136, 2588–2593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.J.; Nowson, C.A.; MacGregor, G.A. Fruit and vegetable consumption and stroke: Meta-analysis of cohort studies. Lancet 2006, 367, 320–326. [Google Scholar] [CrossRef]
- Farnham, M.W.; Wilson, P.E.; Stephenson, K.K.; Fahey, J.W. Genetic and environmental effects on glucosinolate content and chemoprotective potency of broccoli. Plant Breed. 2004, 123, 60–65. [Google Scholar] [CrossRef]
- FAOSTAT—Food and Agriculture Organization of the United Nations. Top exports of Cabbages and Other Brassicas. In FAOSTAT Database; FAOSTAT: Rome, Italy, 2011. [Google Scholar]
- FAOSTAT—Food and Agriculture Organization of the United Nations. Top exports of Cabbages and Other Brassicas. In FAOSTAT Database; FAOSTAT: Rome, Italy, 2019. [Google Scholar]
- Francisco, M.; Tortosa, M.; Martinez-Ballesta, M.; Velasco, P.; Garcia-Viguera, C.; Moreno, D.A. Nutritional and phytochemical value of Brassica crops from the agri-food perspective. Ann. Appl. Biol. 2017, 170, 273–285. [Google Scholar] [CrossRef]
- Ciancaleoni, S.; Chiarenza, G.L.; Raggi, L.; Branca, F.; Negri, V. Diversity characterisation of broccoli (Brassica oleracea L.var. italica Plenck) landraces for their on-farm (in situ) safeguard and use in breeding programs. Genet. Resour. Crop. Evol. 2014, 61, 451–464. [Google Scholar] [CrossRef]
- Fahey, J.W. Brassicas. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Cambridge, MA, USA, 2003; pp. 606–615. ISBN 9780122270550. [Google Scholar]
- Köpke, U. Organic foods: Do they have a role? Forum Nutr. 2005, 57, 62–72. [Google Scholar]
- Ou, B.X.; Huang, D.J.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Analysis of antioxidant activities of common vegetables employing oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays: A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Yu, L. Total phenolic contents and antioxidant properties of commonly consumed vegetables grown in Colorado. LWT Food Sci. Techol. 2006, 39, 1155–1162. [Google Scholar] [CrossRef]
- Azuma, K.; Ippoushi, K.; Ito, H.; Higashio, H.; Terao, J. Evaluation of antioxidative activity of vegetable extracts in linoleic acid emulsion and phospholipid bilayers. J. Sci. Food Agric. 1999, 79, 2010–2016. [Google Scholar] [CrossRef]
- Wu, X.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Gebhardt, S.E.; Prior, R.L. Lipophilic and hydrophilic antioxidant capacities of common foods in the United States. J. Agric. Food Chem. 2004, 52, 4026–4037. [Google Scholar] [CrossRef] [PubMed]
- Podsedek, A. Natural antioxidants and antioxidant capacity of Brassica vegetables: A review. LWT Food Sci. Technol. 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Soengas, P.; Sotelo, T.; Velasco, P.; Cartea, M.E. Antioxidant Properties of Brassica Vegetables. Funct. Plant Sci. Biotechnol. 2011, 43, 55. [Google Scholar]
- Kurilich, A.C.; Jeffery, E.H.; Juvik, J.A.; Wallig, M.A.; Klein, B.P. Antioxidant capacity of different broccoli (Brassica oleracea) genotypes using the oxygen radical absorbance capacity (ORAC) assay. J. Agric. Food Chem. 2002, 50, 5053–5057. [Google Scholar] [CrossRef]
- Jahangir, M.; Abdel-Farida, I.B.; Kima, H.K.; Choia, Y.H.; Verpoortea, R. Healthy and unhealthy plants: The effect of stress on the metabolism of Brassicaceae. Environ. Exp. Bot. 2009, 67, 23–33. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, D.A.; Carvajal, M.; Lopez-Berenguer, C.; García-Viguera, C. Chemical and biological characterisation of nutraceutical compounds of broccoli. J. Pharm. Biomed. Anal. 2006, 41, 1508–1522. [Google Scholar] [CrossRef] [PubMed]
- Heimler, D.; Vignolini, P.; Dini, M.G.; Vincieri, F.F.; Romani, A. Antiradical activity and polyphenol composition of local Brassicaceae edible varieties. Food Chem. 2006, 99, 464–469. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic Compounds in Brassica Vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Fernandes, F.; Sousa, C.; Valentão, P.; Pereira, J.A.; Andrade, P.B. Metabolic and Bioactivity Insights into Brassica oleracea var. acephala. J. Agric. Food Chem. 2009, 57, 8884–8892. [Google Scholar] [CrossRef]
- Haytowitz, D.B.; Wu, X.; Bhagwat, S. USDA Database for the Flavonoid Content of Selected Foods, Release 3.3. U.S. Department of Agriculture, Agricultural Research Service. Nutrient Data Laboratory Home Page. 2018. Available online: http://www.ars.usda.gov/nutrientdata/flav (accessed on 3 May 2020).
- Sousa, C.; Lopes, G.; Pereira, D.M.; Taveira, M.; Valentao, P.; Seabra, R.M.; Pereira, J.A. Screening of antioxidant compounds during sprouting of Brassica oleracea L. var. costata DC. Comb. Chem. High Throughput Screen. 2007, 10, 377–386. [Google Scholar] [CrossRef]
- Ackland, M.L.; Van De Waarsenburg, S.; Jones, R. Synergistic antiproliferative action of the flavanols quercetin and kaempferol in cultured human cancer cell lines. In Vivo 2005, 19, 69–76. [Google Scholar] [PubMed]
- Kim, D.O.; Padilla-Zakour, O.I.; Griffiths, P.D. Flavonoids and antioxidant capacity of various cabbage genotypes at juvenile stage. J. Food Sci. 2004, 69, C685–C689. [Google Scholar] [CrossRef]
- Calderon-Montano, J.M.; Burgos-Moron, E.; Pérez-Guerrero, C.; Lòpez-Lazaro, M. A review on the dietary flavonoid kaempferol. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef] [PubMed]
- Bahorun, T.; Luximon-Ramma, A.; Crozier, A.; Aruoma, O.I. Total phenol; flavonoid; proanthocyanidin and vitamin C levels and antioxidant activities of Mauritian vegetables. J. Sci. Food Agric. 2004, 84, 1553–1561. [Google Scholar] [CrossRef]
- Moreno, D.A.; Pérez-Balibrea, S.; Ferreres, F.; Gil-Izquierdo, A.; García-Viguera, C. Acylated anthocyanins in broccoli sprouts. Food Chem. 2010, 123, 358–363. [Google Scholar] [CrossRef]
- Ahmadiani, N.; Robbins, R.J.; Collins, T.M.; Giusti, M.M. Anthocyanins Contents, Profiles, and Color Characteristics of Red Cabbage Extracts from Different Cultivars and Maturity Stages. J. Agric. Food Chem. 2014, 62, 7524–7531. [Google Scholar] [CrossRef]
- Dyrby, M.; Westergaard, N.; Stapelfeldt, H. Light and heat sensitivity of red cabbage extract in soft drink model system. Food Chem. 2001, 72, 431–437. [Google Scholar] [CrossRef]
- Milder, I.E.J.; Arts, I.C.W.; Van De Putte, B.; Venema, D.P.; Hollman, P.C.H. Lignin contents of Dutch plant foods: A database including lariciresinol; pinoresinol; secoisolariciresinol and matairesinol. Br. J. Nutr. 2005, 93, 393–402. [Google Scholar] [CrossRef]
- Kurilich, A.C.; Tsau, G.J.; Brown, A.; Howard, L.; Klein, B.P.; Jeffery, E.H.; Kushad, M.; Walling, M.A.; Juvik, J.A. Carotene; tocopherol; and ascorbate contents in subspecies of Brassica oleracea. J. Agric. Food Chem. 1999, 47, 1576–1581. [Google Scholar] [CrossRef]
- Vallejo, F.; Tomas-Barberan, F.A.; Garcia-Viguera, C. Potential bioactive compounds in health promotion from broccoli cultivars grown in Spain. J. Sci. Food Agric. 2002, 82, 1293–1297. [Google Scholar] [CrossRef]
- Davey, M.W.; Van Montagu, M.; Inze, D.; Sanmartin, M.; Kanellis, A.; Smirnoff, N. Plant L-ascorbic acid: Chemistry; function; metabolism; bioavailability and effects of processing. J. Sci. Food Agric. 2000, 80, 825–860. [Google Scholar] [CrossRef]
- Sanlier, N.; Guler Saban, M. The Benefits of Brassica Vegetables on Human Health. J. Hum. Health Res. 2018, 1, 104. [Google Scholar]
- Bailey, L.B.; Rampersaud, G.C.; Kauwell, G.P.A. Folic acid supplements and fortification affect the risk for neural tube defects; vascular disease and cancer: Evolving science. J. Nutr. 2003, 133, 1961S–1968S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atteia, B.M.R.; El-Kak, A.E.-A.A.; Lucchesi, P.A.; Delafontane, P. Antioxidant activity of folic acid: From mechanism of action to clinical application. FASEB J. 2009, 23, 103–107. [Google Scholar]
- Quang, D.D. Antioxidant properties of folic acid: A DFT study. Vietnam J. Sci. Technol. 2018, 56, 39. [Google Scholar] [CrossRef] [Green Version]
- Rice-Evans, C.; Sampson, J.; Bramley, P.M.; Holloway, D.E. Why do we expect carotenoids to be antioxidants in vivo. Free Radic. Res. 1997, 26, 381–398. [Google Scholar] [CrossRef]
- Muller, H. Determination of the carotenoid content in selected vegetables and fruit by HPLC and photodiode array detection. Food Res. Technol. 1997, 204, 88–94. [Google Scholar] [CrossRef]
- Wills, R.B.H.; Rangga, A. Determination of carotenoids in Chinese vegetables. Food Chem. 1996, 56, 451–455. [Google Scholar] [CrossRef]
- Piironen, V.; Syvaoja, E.L.; Varo, P.; Salminen, K.; Koivistoinen, P. Tocopherols and tocotrienols in Finnish foods: Vegetables; fruits; and berries. J. Agric. Food Chem. 1986, 34, 742–746. [Google Scholar] [CrossRef]
- Stampfer, M.J.; Rimm, E.B. Epidemiologic evidence for vitamin E in prevention of cardiovascular disease. Am. J. Clin. Nutr. 1995, 62, 1365–1369. [Google Scholar] [CrossRef]
- Ali, Z.; Waheed, H.; Gul, A.; Afzal, F.; Anwaar, K.; Imran, S. Brassicaceae plants. In Oilseed Crops; Ahmad, P., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar] [CrossRef]
- Thavarajah, D.; Lawrence, T.; Powers, S.; Jones, B.; Johnson, N.; Kay, J.; Bandaranayake, A.; Shipe, E.; Thavarajah, P. Genetic variation in the prebiotic carbohydrate and mineral composition of kale (Brassica oleracea L. var. acephala) adapted to an organic cropping system. J. Food Comp. Anal. 2021, 96, 103718. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.-E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef] [PubMed]
- Sisti, M.; Amagliani, G.; Brandi, G. Antifungal activity of Brassica oleracea var. botrytis fresh aqueous juice. Fitoterapia 2003, 74, 453–458. [Google Scholar]
- Branca, F.; Ragusa, L.; Tribulato, A.; Velasco, P.; Cartea, M.E. Glucosinolate profile in different mediterranean brassica species (n = 9). Acta Hortic. 2013, 1005, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Velasco, P.; Cartea, M.E.; Gonzalez, C.; Vilar, M.; Ordàs, A. Factors affecting the glucosinolate content of kale (Brassica oleracea acephala group). J. Agric. Food Chem. 2007, 55, 955–962. [Google Scholar] [CrossRef] [PubMed]
- López-Berenguer, C.; Martínez-Ballesta, M.C.; García-Viguera, C.; Carvajal, M. Leaf water balance mediated by aquaporins under salt stress and associated glucosinolate synthesis in broccoli. Plant Sci. 2008, 174, 321–328. [Google Scholar] [CrossRef]
- Mithen, R. Glucosinolates—Biochemistry; Genetics and biological activity. Plant Growth Regul. 2001, 34, 91–103. [Google Scholar] [CrossRef]
- Verkerk, R.; Schreiner, M.; Krumbein, A.; Ciska, E.; Holst, B.; Rowland, I.; de Schrijver, R.; Hansen, M.; Gerhäuser, C.; Mithen, R.; et al. Glucosinolates in Brassica vegetables: The influence of the food supply chain on intake, bioavailability and human health. Mol. Nutr. Food Res. 2009, 53, 219–265. [Google Scholar] [CrossRef] [PubMed]
- Blažević, I.; Montaut, S.; Burčul, F.; Rollin, P. Glucosinolates: Novel sources and biological potential. In Glucosinolates; Mérillon, J.-M., Ramawat, K.G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 3–60. [Google Scholar]
- Giamoustaris, A.; Mithen, R. The effect of modifying the glucosinolate content of leaves of oilseed rape (Brassica napus ssp. oleifera) on its interaction with specialist and generalist pests. Ann. Appl. Biol. 1995, 126, 347–363. [Google Scholar] [CrossRef]
- Mithen, R.; Faulkner, K.; Magrath, R.; Rose, P.; Williamson, G.; Marquez, J. Development of isothiocyanate-enriched broccoli and its enhanced ability to induce phase 2 detoxification enzymes in mammalian cells. Theor. Appl. Gen. 2003, 106, 727–734. [Google Scholar] [CrossRef]
- Sikorska-Zimny, K.; Beneduce, L. The glucosinolates and their bioactive derivatives in Brassica: A review on classification, biosynthesis and content in plant tissues, fate during and after processing, effect on the human organism and interaction with the gut microbiota. Crit. Rev. Food Sci. Nutr. 2020, 25, 1–24. [Google Scholar] [CrossRef] [PubMed]
- VanEtten, C.H.; Daxenbichler, M.E.; Tookey, H.L.; Kwolek, W.F.; Williams, P.H.; Yoder, O.C. Glucosinolates: Potential toxicants in cabbage cultivars. J. Am. Soc. Hort. Sci. 1980, 105, 710–714. [Google Scholar]
- Sones, K.; Heaney, R.K.; Fenwick, G.R. The glucosinolate content of UK vegetables: Cabbage (Brassica oleracea); swede (B. napus) and turnip (B. campestris). Food Addit. Contam. 1984, 3, 289–296. [Google Scholar] [CrossRef]
- Cartea, M.E.; Velasco, P.; Obregòn, S.; del Rìo, M.; Padilla, G.; de Haro, A. Seasonal variation in glucosinolate content in Brassica oleracea crops grown in northwestern Spain. Phytochemistry 2008, 69, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Ciska, E.; Martyniak-Przybyszewska, B.; Kozlowska, H. Content of glucosinolates in cruciferous vegetables grown at the same site for two years under different climatic conditions. J. Agric. Food Chem. 2000, 48, 2862–2867. [Google Scholar] [CrossRef] [PubMed]
- Carlson, D.G.; Daxenbichler, M.E.; Tookey, H.L. Glucosinolates in turnip tops and roots: Cultivars grown for greens and/or roots. J. Am. Soc. Hort. Sci. 1987, 112, 179–183. [Google Scholar]
- Rosa, E.A.S.; Heaney, R.K. Seasonal variation in protein; mineral and glucosinolate composition of Portuguese cabbage and kale. Anim. Feed Sci. Technol. 1996, 57, 111–127. [Google Scholar] [CrossRef]
- Kushad, M.M.; Brown, A.F.; Kurilich, A.C.; Juvik, J.A.; Klein, B.; Wallig, M.A.; Jeffery, E.H. Variation of glucosinolates in vegetable subspecies of Brassica oleracea. J. Food Agric. Chem. 1999, 47, 1541–1548. [Google Scholar] [CrossRef]
- Padilla, G.; Cartea, M.E.; Velasco, P.; de Haro, A.; Ordàs, A. Variation of glucosinolates in vegetable crops of Brassica rapa. Phytochemistry 2007, 68, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Rosa, E.A.S. Glucosinolates from flower buds of Portuguese Brassica crops. Phytochemistry 1997, 44, 1415–1419. [Google Scholar] [CrossRef]
- Cartea, M.E.; Rodrìguez, V.M.; Velasco, P.; de Haro, A.; Ordàs, A. Variation of glucosinolates and nutritional value in nabicol (Brassica napus pabularia group). Euphytica 2008, 159, 111–122. [Google Scholar] [CrossRef]
- Angelino, D.; Dosz, E.B.; Sun, J.; Hoeflinger, J.L.; Van Tassell, M.L.; Chen, P.; Harnly, J.M.; Miller, M.J.; Jeffery, E.H. Myrosinase-dependent and -independent formation and control of isothiocyanate products of glucosinolate hydrolysis. Front. Plant. Sci. 2015, 6, 831. [Google Scholar] [CrossRef] [Green Version]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Grubb, C.D.; Abel, S. Glucosinolate metabolism and its control. Trends Plant. Sci. 2006, 11, 89–100. [Google Scholar] [CrossRef]
- Keum, Y.S.; Jeong, W.S.; Kong, A.N.T. Chemoprevention by isothiocyanates and their underlying molecular signaling mechanisms. Mutat. Res. 2004, 555, 191–202. [Google Scholar] [CrossRef]
- Cartea, M.E.; Velasco, P. Glucosinolates in Brassica foods: Bioavailability in food and significance for human health. Phytochem. Rev. 2008, 7, 213–229. [Google Scholar] [CrossRef]
- Fahey, J.W.; Haristoy, X.; Dolan, P.M.; Kensler, T.W.; Scholtus, I.; Stephenson, K.K.; Talalay, P.; Lozniewski, A. Sulforaphane inhibits extracellular; intracellular; and antibiotic-resistant strains of Helicobacter pylori and prevents benzo[a]pyrene-induced stomach tumors. Proc. Natl. Acad. Sci. USA 2002, 99, 7610–7615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Matusheski, N.V.; Jeffery, E.H. Comparison of the bioactivity of two glucoraphanin hydrolysis products found in broccoli; sulforaphane and sulforaphane nitrile. J. Agric. Food Chem. 2001, 49, 5743–5749. [Google Scholar] [CrossRef] [PubMed]
- Bonasia, A.; Conversa, G.; Lazzizera, C.; Elia, A. Pre-harvest nitrogen and Azoxystrobin application enhances postharvest shelf-life in butterhead lettuce. Postharvest Biol. Technol. 2013, 85, 67–76. [Google Scholar] [CrossRef]
- Wieczorek, M.N.; Walczak, M.; Skrzypczak-Zielinska, M.; Jelen, H.H. Bitter taste of Brassica vegetables: The role of genetic factors; receptors; isothiocyanates; glucosinolates; and flavor context. Crit. Rev. Food Sci. Nutr. 2018, 58, 3130–3140. [Google Scholar] [CrossRef] [PubMed]
- Ghawi, S.K.; Shen, Y.; Niranjan, K.; Methven, L. Consumer acceptability and sensory profile of cooked broccoli with mustard seeds added to improve chemoprotective properties. J. Food Sci. 2014, 79, 1756–1762. [Google Scholar] [CrossRef]
- Stoewsand, S. Bioactive organosulfur phytochemicals in Brassica oleracea vegetables—A review. Food Chem. Toxicol. 1995, 33, 537. [Google Scholar] [CrossRef]
- Palermo, M.; Pellegrini, N.; Fogliano, V. The effect of cooking on the phytochemical content of vegetables. J. Sci. Food Agric. 2014, 94, 1057. [Google Scholar] [CrossRef] [PubMed]
- Kubec, R.; Drhova, V.; Velisek, J. Thermal Degradation of S-Methylcysteine and Its Sulfoxide Important Flavor Precursors of Brassica and Allium Vegetables. J. Agric. Food Chem. 1998, 46, 4334. [Google Scholar] [CrossRef]
- Hansen, M.; Laustsen, A.M.; Olsen, C.E.; Poll, L.; Sørensen, H. Chemical and sensory quality of broccoli (Brassica oleracea L. var italica). J. Food Qual. 1997, 20, 441–459. [Google Scholar] [CrossRef]
- Akpolat, H.; Barringer, S.A. The Effect of pH and Temperature on Cabbage Volatiles during Storage. J. Food Sci. 2015, 80, S1878–S1884. [Google Scholar] [CrossRef]
- Bell, L.; Oloyede, O.O.; Lignou, S.; Wagstaff, C.; Methven, L. Taste and Flavor Perceptions of Glucosinolates, Isothiocyanates, and Related Compounds. Mol. Nutr. Food Res. 2018, 62, e1700990. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, M.N.; Jelen, H.H. Volatile Compounds of Selected Raw and Cooked Brassica Vegetables. Molecules 2019, 24, 391. [Google Scholar] [CrossRef] [Green Version]
- Rosa, E.A.S.; Heaney, R.K.; Fenwick, G.R.; Portas, C.A.M. Glucosinolates in crop plants. Hortic. Rev. 1997, 19, 99–215. [Google Scholar]
- Schonhof, I.; Krumbein, A.; Brückner, B. Genotypic effects on glucosinolates and sensory properties of broccoli and cauliflower. Food Nahr. 2004, 48, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Beck, T.K.; Jensen, S.; Bjoern, G.K.; Kidmose, U. The Masking Effect of Sucrose on Perception of Bitter Compounds in Brassica Vegetables. J. Sens. Stud. 2014, 29, 190–200. [Google Scholar] [CrossRef]
- Baik, H.Y.; Juvik, J.A.; Jeffery, E.H.; Wallig, M.A.; Kushad, M.; Klein, B.P. Relating glucosinolate content and flavor of broccoli cultivars. J. Food Sci. 2003, 68, 1043–1050. [Google Scholar] [CrossRef]
- Schatzer, M.; Rust, P.; Elmadfa, I. Fruit and vegetable intake in Austrian adults: Intake frequency; serving sizes; reasons for and barriers to consumption; and potential for increasing consumption. Public Health Nutr. 2009, 13, 480–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, D.N.; Melo, L.; Zabaras, D.; Delahunty, C.M. Acceptance of health-promoting Brassica vegetables: The influence of taste perception; information and attitudes. Public Health Nutr. 2012, 15, 1474–1482. [Google Scholar] [CrossRef] [Green Version]
- Reed, D.R.; Tanaka, T.; McDaniel, A.H. Diverse tastes: Genetics of sweet and bitter perception. Physiol. Behav. 2006, 88, 215–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drewnowski, A. The science and complexity of bitter taste. Nutr. Rev. 2001, 59, 163–169. [Google Scholar] [CrossRef]
- Branca, F.; Ragusa, L.; Tribulato, A. Diversity of Kale Growing in Europe as a Basis for Crop Improvement. Acta Hortic. 2013, 1005, 141–147. [Google Scholar] [CrossRef]
- Jagdish, S.; Upadhyay, A.K.; Singh, S.; Rai, M. Total phenolics content and free radical scavenging activity of Brassica vegetables. J. Food Sci. Tech. Mysore 2009, 46, 595–597. [Google Scholar]
- Perez-Balibrea, S.; Moreno, D.A.; Garcia-Viguera, C. Genotypic effects on the phytochemical quality of seeds and sprouts from commercial broccoli cultivar. Food Chem. 2011, 125, 348–354. [Google Scholar] [CrossRef]
- Volden, J.; Bengtsson, G.B.; Wicklund, T. Glucosinolates; L-ascorbic acid; total phenols; anthocyanins; antioxidant capacities and colour in cauliflower (Brassica oleracea L. ssp. botrytis); effects of long-term freezer storage. Food Chem. 2009, 112, 967–976. [Google Scholar] [CrossRef]
- Branca, F.; Chiarenza, G.L.; Pinio, M.; Alonso, M.; Argento, S. Characteristics and seed production of sicilian landraces of violet cauliflower. Acta Hortic. 2013, 1005, 519–524. [Google Scholar] [CrossRef]
- Singh, J.; Upadhyay, A.K.; Prasad, K.; Bahadur, A.; Rai, M. Variability of carotenes. vitamin C. E and phenolics in Brassica vegetables. J. Food Compos. Anal. 2007, 20, 106–112. [Google Scholar] [CrossRef]
- Francisco, M.; Moreno, D.A.; Cartea, M.E.; Ferreres, F.; Garcia-Viguera, C.; Velasco, P. Simultaneous identification of glucosinolates and phenolic compounds in a representative collection of vegetable Brassica rapa. J. Chromatogr. A 2009, 1216, 6611–6619. [Google Scholar] [CrossRef] [PubMed]
- Wachtel-Galor, S.; Wong, K.W.; Benzie, I.F.F. The effect of cooking on Brassica vegetables. Food Chem. 2008, 110, 706–710. [Google Scholar] [CrossRef]
- Martìnez-Sanchez, A.; Gil-Izquierdo, A.; Gil, M.I.; Ferreres, F. A comparative study of flavonoid compounds; vitamin C, and antioxidant properties of baby leaf Brassicaceae species. J. Agric. Food Chem. 2008, 56, 2330–2340. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Villaluenga, C.; Peñas, E.; Ciska, E.; Piskula, M.K.; Kozlowska, H.; Vidal-Valverde, C.; Frias, J. Time dependence of bioactive compounds and antioxidant capacity during germination of different cultivars of broccoli and radish seeds. Food Chem. 2010, 120, 710–716. [Google Scholar] [CrossRef]
- Francisco, M.; Cartea, M.E.; Soengas, P.; Velasco, P. Effect of Genotype and Environmental Conditions on Health-Promoting Compounds in Brassica rapa. J. Agric. Food Chem. 2011, 59, 2421–2431. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, E.H.; Brown, A.F.; Kurilich, A.C.; Keek, A.S.; Matusheski, N.; Klein, B.P.; Juvik, J.A. Variation in content of bioactive components in broccoli. Study review. J. Food Compos. Anal. 2003, 16, 323–330. [Google Scholar] [CrossRef]
- Fernandes, F.; Valentão, P.; Sousa, C.; Pereira, J.A.; Seabra, R.M.; Andrade, P.B. Chemical and antioxidative assessment of dietary turnip (Brassica rapa var. rapa L.). Food Chem. 2007, 105, 1003–1010. [Google Scholar] [CrossRef]
- Brown, P.D.; Tokuhisa, J.G.; Reichelt, M.; Gershenzon, J. Variation of glucosinolate accumulation among different organs and developmental stages of Arabidopsis thaliana. Phytochemistry 2003, 62, 471–481. [Google Scholar] [CrossRef]
- Seong, G.U.; Hwang, I.W.; Chung, S.K. Antioxidant capacities and polyphenolics of Chinese cabbage (Brassica rapa L. ssp. pekinensis) leaves. Food Chem. 2016, 199, 612–618. [Google Scholar] [CrossRef]
- Rybarczyk, A.; Wold, B.A.; Hansen, M.K.; Hagen, S.F. Vitamin C in broccoli (Brassica oleracea L. var. italica) flower buds as affected by postharvest light, UV-B irradiation and temperature. Postharvest Biol. Technol. 2014, 98, 82–89. [Google Scholar]
- Vallejo, F.; Tomas-Barberan, F.; Garcia-Viguera, C. Changes in broccoli (Brassica oleracea L. var. italica) health-promoting compounds with inflorescence development. J. Agric. Food Chem. 2003, 51, 3776–3782. [Google Scholar] [CrossRef]
- Lefsrud, M.; Kopsell, D.; Wenzel, A.; Sheehan, J. Changes in kale (Brassica oleracea L. var. acephala) carotenoid content and chlorophyll pigment concentrations during leaf ontogeny. Sci. Hort. Amst. 2007, 112, 136–141. [Google Scholar] [CrossRef]
- Vale, A.P.; Cidade, H.; Pinto, M.; Oliveira, M.B.P.P. Effect of sprouting and light cycle on antioxidant activity of Brassica oleracea varieties. Food Chem. 2014, 165, 379–387. [Google Scholar] [CrossRef]
- De Pascale, S.; Maggio, A.; Pernice, R.; Fogliano, V.; Barbieri, G. Sulphur fertilization may improve the nutritional value of Brassica rapa L. subsp. sylvestris. Eur. J. Agron. 2007, 26, 418–424. [Google Scholar] [CrossRef]
- Renaud, E.N.C.; Lammerts Van Bueren, E.T.; Myers, J.R.; Paulo, M.J.; Van Eeuwijk, F.A.; Zhu, N.; Juvik, J.A. Variation in broccoli cultivar phytochemical content under organic and conventional management systems: Implications in breeding for nutrition. PLoS ONE 2014, 9, e95683. [Google Scholar] [CrossRef] [Green Version]
- Rosa, E.A.S.; Rodrigues, A.S. Total and individual glucosinolate content in 11 broccoli cultivars grown in early and late season. HortScience 2001, 36, 56–59. [Google Scholar] [CrossRef]
- Aksouh, N.M.; Jacobs, B.C.; Stoddard, F.L.; Mailer, R.J. Response of canola to different heat stresses. Aust. J. Agric. Res. 2001, 52, 817–824. [Google Scholar] [CrossRef]
- Schonhof, I.; Klaring, H.P.; Krumbein, A.; Clauβen, W.; Schreiner, M. Effect of temperature increase under low radiation conditions on phytochemicals and ascorbic acid in greenhouse grown broccoli. Agric. Ecosyst. Environ. 2007, 119, 103–111. [Google Scholar] [CrossRef]
- Pereira, F.M.V.; Rosa, E.; Fahey, J.W.; Stephenson, K.K.; Carvalho, R.; Aires, A. Influence of temperature and ontogeny on the levels of glucosinolates in broccoli (Brassica oleracea var. italica) sprouts and their effect on the induction of mammalian phase 2 enzymes. J. Agric. Food Chem. 2002, 50, 6239–6244. [Google Scholar] [CrossRef]
- Vallejo, F.; Tomas-Barberan, F.; Garcia-Viguera, C. Healthpromoting compounds in broccoli as influenced by refrigerated transport and retail sale period. J. Agric. Food Chem. 2003, 51, 3029–3034. [Google Scholar] [CrossRef] [PubMed]
- Bjorkman, M.; Klingen, I.; Birch, A.; Bones, A.; Bruce, T.; Johansen, T.J.; Meadow, R.; Molmann, J.; Seljasen, R.; Smart, L.E.; et al. Phyto-chemicals of Brassicaceae in plant protection and human health. Influences of climate; environment and agronomic practice. Phytochemistry 2011, 72, 538–556. [Google Scholar] [CrossRef]
- Francisco, M.; Cartea, M.E.; Butrón, A.M.; Sotelo, T.; Velasco, P. Environmental and genetic effects on yield and secondary metabolite production in Brassica rapa crops. J. Agric. Food Chem. 2012, 60, 5507–5514. [Google Scholar] [CrossRef] [PubMed]
- Pék, Z.; Daood, H.; Nagyné, M.G.; Neményi, A.; Helyes, L. Effect of environmental conditions and water status on the bioactive compounds of broccoli. Cent. Eur. J. Biol. 2013, 8, 777–787. [Google Scholar] [CrossRef]
- Singh, S.; Sinha, S. Accumulation of metals and its effects in Brassica juncea L. Czern. (cv. Rohini) grown on various amendments of tannery waste. Ecotox. Environ. Saf. 2005, 62, 118–127. [Google Scholar] [CrossRef]
- Xiong, Z.T.; Liu, C.; Geng, B. Phytotoxic effects of copper on nitrogen metabolism and plant growth in Brassica pekinensis Rupr. Ecotox. Environ. Saf. 2006, 64, 273–280. [Google Scholar] [CrossRef]
- Rodriguez-Hernandez, M.C.; Moreno, D.A.; Carvajal, M.; Martìnez-Ballesta, M.C. Genotype influences sulfur metabolism in broccoli (Brassica oleracea L.) under elevated CO2 and NaCl stress. Plant. Cell Physiol. 2014, 55, 2047–2059. [Google Scholar] [CrossRef] [Green Version]
- Sakr, M.Y.; Ibrahim, H.M.; ElAwady, A.E.; AboELMakarm, A.A. Growth, yield and biochemical constituents as well as post-harvest quality of water-stressed broccoli (Brassica oleraceae L. var. italica) as affected by certain biomodulators. Sci. Hortic. 2021, 275, 109605. [Google Scholar] [CrossRef]
- Ragusa, L.; Picchi, V.; Tribulato, A.; Cavallaro, C.; Lo Scalzo, R.; Branca, F. The effect of the germination temperature on the phytochemical content of broccoli and rocket sprouts. Int. J. Food. Sci. Nutr. 2016, 68, 411–420. [Google Scholar] [CrossRef]
- Sousa, C.; Pereira, D.M.; Pereira, J.A.; Bento, A.; Rodrigues, M.A.; Dopico-Garcìa, S.; Valentao, P.; Lopes, G.; Ferreres, F.; Seabra, R.M.; et al. Multivariate analysis of tronchuda cabbage (Brassica oleracea L. var. costata DC) phenolics: Influence of fertilizers. J. Agric. Food Chem. 2008, 56, 2231–2239. [Google Scholar] [CrossRef]
- Vrchoska, V.; Sousa, C.; Valentao, P.; Ferreres, F.; Pereira, J.A.; Seabra, R.M.; Andrade, P.B. Antioxidative properties of tronchuda cabbage (Brassica oleracea L. Var. costata DC) external leaves against DPPH; superoxide radical; hydroxyl radical and hypochlorous acid. Food Chem. 2006, 98, 416–425. [Google Scholar] [CrossRef]
- Dangour, A.D.; Dodhia, S.K.; Hayter, A.; Allen, E.; Lock, K.; Uauy, R. Nutritional quality of organic foods: A systematic review. Am. J. Clin. Nutr. 2009, 90, 680–685. [Google Scholar] [CrossRef] [Green Version]
- Hoefkens, C.; Sioen, I.; Baert, K.; De Meulenaer, B.; De Henauw, S.; Vandekinderen, I.; Devlieghere, F.; Opsomer, A.; Verbeke, W.; Van Camp, J. Consuming organic versus conventional vegetables: The effect on nutrient and contaminant intakes. Food Chem. Toxicol. 2010, 48, 3058–3066. [Google Scholar] [CrossRef]
- Conversa, G.; Bonasia, A.; Lazzizera, C.; Elia, A. Bio-physical; physiologiacal; and nutritional aspects of ready-to-use cima di rapa (Brassica rapa L. subsp. sylvestris L. Janch. var. esculenta Hort.) as affected by conventional and organic growing systems and storage time. Sci. Hortic. 2016, 213, 76–86. [Google Scholar] [CrossRef]
- Khan, M.A.M.; Ulrichs, C.; Mewis, I. Drought stress—Impact on glucosinolate profile and performance of phloem feeding cruciferous insects. Acta Hortic. 2011, 917, 111–117. [Google Scholar] [CrossRef]
- Cogo, S.L.P.; Chaves, F.C.; Schirmer, M.A.; Zambiazi, R.C.; Nora, L.; Silva, J.A.; Rombaldi, C.V. Low soil water content during growth contributes to preservation of green colour and bioactive compounds of cold-stored broccoli (Brassica oleracea L.) florets. Postharvest Biol. Technol. 2011, 60, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Hooks, C.R.R.; Johnson, M.W. Impact of agricultural diversification on the insect community of cruciferous crops. Crop. Prot. 2003, 22, 223–238. [Google Scholar] [CrossRef]
- Kim, S.J.; Matsuo, T.; Watanabe, M.; Watanabe, Y. Effect of nitrogen and sulphur application on the glucosinolate content in vegetable turnip rape (Brassica rapa L.). Soil Sci. Plant. Nutr. 2002, 48, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Kopsell, D.E.; Kopsell, D.A.; Randle, W.A.; Coolong, T.W.; Sams, C.E.; Curran-Celentano, J. Kale carotenoids remain stable while flavor compounds respond to changes in sulfur fertility. J. Agric. Food Chem. 2003, 51, 5319–5325. [Google Scholar] [CrossRef]
- Li, S.; Schonhof, I.; Krumbein, A.; Li, L.; Stutzel, H.; Schreiner, M. Glucosinolate concentration in turnip (Brassica rapa ssp. rapifera L.) roots as affected by nitrogen and sulfur supply. J. Agric. Food Chem. 2007, 55, 8452–8457. [Google Scholar] [CrossRef]
- Schreiner, M.; Huyskens-Keil, S.; Peters, P.; Schonhof, I.; Krumbein, A.; Widell, S. Seasonal climate effects on root colour and compounds of red radish. J. Sci. Food Agric. 2002, 82, 1325–1333. [Google Scholar] [CrossRef]
- Barbieri, G.; Bottino, A.; Orsini, F.; De Pascale, S. Sulfur fertilization and light exposure during storage are critical determinants of the nutritional value of ready-to-eat friariello campano (Brassica rapa L. subsp. sylvestris). J. Sci. Food Agric. 2009, 89, 2261–2266. [Google Scholar] [CrossRef]
- Dubuis, P.H.; Marazzi, C.; Städler, E.; Mauch, F. Sulphur deficiency causes a reduction in antimicrobial potential and leads to increased disease susceptibility of oilseed rape. J. Phytopathol. 2005, 153, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Kopsell, D.A.; Kopsell, D.E.; Curran-Celentano, J. Carotenoid pigments in kale are influenced by nitrogen concentration and form. J. Sci. Food Agric. 2007, 87, 900–907. [Google Scholar] [CrossRef]
- Zhao, F.; Evans, E.J.; Bilsborrow, P.E.; Syers, J.K. Influence of nitrogen and sulphur on the glucosinolate profile of rapeseed (Brassica napus). J. Sci. Food Agric. 1994, 64, 295–304. [Google Scholar] [CrossRef]
- Robbins, R.J.; Keck, A.S.; Banuelos, G.; Finley, J.W. Cultivation conditions and selenium fertilization alter the phenolic profile; glucosinolate; and sulforaphane content of broccoli. J. Med. Food 2005, 8, 204–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabek, S.; Toth, N.; Redovnikovic, I.R.; Custic, M.H.; Benko, B.; Žutic, I. The effect of nitrogen fertilization on nitrate accumulation; and the content of minerals and glucosinolates in broccoli cultivars. Food Technol. Biotechnol. 2012, 50, 9. [Google Scholar]
- Tolrà, R.P.; Poschenrieder, C.; Alonso, R.; Barceló, D.; Barceló, J. Influence of zinc hyperaccumulation on glucosinolates in Thlaspi caerulescens. New Phytol. 2001, 151, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Aghajanzadeh, T.A.; Prajapati, D.H.; Burow, M. Copper toxicity affects indolic glucosinolates and gene expression of key enzymes for their biosynthesis in Chinese cabbage. Arch. Agron. Soil Sci. 2020, 66, 1288–1301. [Google Scholar] [CrossRef]
- Pandey, C.; Augustine, R.; Panthri, M.; Zia, I.; Bisht, N.C.; Gupta, M. Arsenic affects the production of glucosinolate, thiol and phytochemical compounds: A comparison of two Brassica cultivars. Plant Physiol. Biochem. 2017, 111, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Bodnaryk, R.P. Potent effect of jasmonates on indole glucosinolates in oilseed rape and mustard. Phytochemistry 1994, 35, 301–305. [Google Scholar] [CrossRef]
- Mollers, C.; Nehlin, L.; Glimelius, K.; Iqbal, M.C.M. Influence of in vitro culture conditions on glucosinolate composition of microspore-derived embryos of Brassica napus. Physiol. Plant. 1999, 107, 441–446. [Google Scholar] [CrossRef]
- Guo, R.; Shen, W.; Qian, H.; Zhang, M.; Liu, L.; Wang, Q. Jasmonic acid and glucose synergistically modulate the accumulation of glucosinolates in Arabidopsis thaliana. J. Exp. Bot. 2013, 64, 5707–5719. [Google Scholar] [CrossRef] [Green Version]
- Gallo, M.; Esposito, G.; Ferracane, R.; Vinale, F.; Naviglio, D. Beneficial effects of Trichoderma genus microbes on qualitative parameters of Brassica rapa L. subsp. sylvestris L. Janch. Var. esculenta Hort. Eur. Food Res. Technol. 2013, 236, 1063–1071. [Google Scholar] [CrossRef]
- Muelchen, A.M.; Rand, R.E.; Parke, J.L. Evaluation of crucifer manures for controlling aphanomyces root rot of peas. Plant Dis. 1990, 74, 651–654. [Google Scholar] [CrossRef]
- Pant, A.P.; Radovich, T.J.K.; Hue, N.V.; Talcott, S.T.; Krenek, K.A. Vermicompost extracts influence growth; mineral nutrients; phytonutrients and antioxidant activity in pak choi (Brassica rapa cv. bonsai; Chinensis group) grown under vermicompost and chemical fertiliser. J. Sci. Food Agric. 2009, 89, 2383–2392. [Google Scholar] [CrossRef]
- Lola-Luz, T.; Hennequart, F.; Gaffney, M. Enhancement of phenolic and flavonoid compounds in cabbage (Brassica oleraceae) following application of commercial seaweed extracts of the brown seaweed; (Ascophyllum nodosum). J. Agric. Food Sci. 2013, 22, 288–295. [Google Scholar] [CrossRef] [Green Version]
- Argento, S.; Raccuia, S.A.; Melilli, M.G.; Toscano, V.; Branca, F. Brassicas and their glucosinolate content for the biological control of root-knot nematodes in protected cultivation. Acta Hortic. 2013, 1005, 539–544. [Google Scholar] [CrossRef]


{kind=link}
| Species | ssp./var. | Common Name/Italian Name |
|---|---|---|
| Brassica oleracea L. | italica | Broccoli |
| capitata capitata f. rubra capitata f. alba | Cabbage Red cabbage White cabbage | |
| botrytis | Cauliflower | |
| acephala sabellica acephala laciniata acephala | Curly Kale, Red, Green and Russian curly kale Black Cabbage, Italian or Tuscan cabbage Collards | |
| gemmifera | Brussels sprouts | |
| gongylodes | Kohlrabi | |
| sabauda | Savoy cabbage | |
| alboglabra | Chinese kale | |
| costata | Tronchuda cabbage | |
| Brassica rapa L. | rapa | Turnip broccoli |
| sylvestris | Turnip top, broccoli raab/cima di rapa, friarielli | |
| peckinensis | Chinese cabbage | |
| chinensis | Pak-choi, Chinese mustard | |
| japonica | Mizuna, curled mustard, Japanese greens | |
| perviridis | Tendergreen, Spinach mustard | |
| rapifera | Turnip | |
| Brassica napus L. | napus | Rapeseed |
| napobrassica | Swede/rutabaga | |
| Brassica kaber/Brassica arvensis/Sinapis arvensis | Charlock/kaber | |
| Brassica alba/Sinapis alba, Brassica hirta | White or yellow mustard | |
| Brassica nigra/Sinapis nigra | Black mustard | |
| Brassica campestris | Field mustard | |
| Brassica carinata | Ethiopian mustard, Abyssinian mustard, Texsel greens | |
| Brassica juncea | Brown mustards | |
| Raphanus sativus L. | Radish | |
| Raphanus raphanistrum L. | Wild radish | |
| Nasturtium officinale R. BR. | Watercress/Crescione d’acqua | |
| Eruca sativa Mill. | Rocket/Rucola | |
| Eruca vesicaria L. | Rocket Ruca, ruchetta | |
| Diplotaxis tenuifolia L. | Wild rocket/Rughetta selvatica | |
| Diplotaxis muralis L. | Wall rocket |
| Crop | Aliphatic Glucosinolates | Indole Glucosinolates | Arylaliphatic Glucosinolates | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GIB | PRO | SIN | GAL | GRA | GNA | GBN | GIV | GER | GNL | GBS | NGBS | 4HGBS | 4MGBS | GST | |
| Brassica oleracea | |||||||||||||||
| White cabbage [58,59,60,61] | + | + | + | + | + | + | + | + | + | - | + | + | + | + | + |
| Savoy cabbage [58,59,60,61] | + | + | + | - | + | + | + | + | - | - | + | + | - | + | + |
| Red cabbage [58,60,61] | + | + | + | - | + | + | - | + | - | - | + | + | - | - | - |
| Kale [58,60,61,62] | + | + | + | - | + | + | - | + | - | - | + | + | + | + | + |
| Collard [62] | + | + | + | - | - | - | - | + | + | - | + | - | - | - | - |
| Tronchuda cabbage [60,63] | + | + | + | + | + | + | + | + | - | - | + | + | + | + | + |
| Broccoli [62,64] | + | + | + | + | + | + | + | - | + | + | + | + | + | + | + |
| Brussel sprouts [61,62,64] | + | + | + | - | + | + | - | + | - | - | + | + | - | - | - |
| Cauliflower [61,64] | + | + | + | - | + | - | - | + | - | - | + | + | - | - | - |
| Kohlrabi [61] | + | + | + | + | + | + | - | + | - | - | + | + | + | + | - |
| Brassica rapa | |||||||||||||||
| Turnip [59] | + | + | - | - | - | + | + | - | + | + | + | + | + | + | + |
| Turnip greens [65] | + | + | - | + | + | + | + | + | - | + | + | + | + | - | + |
| Turnip tops [66] | + | + | - | - | - | + | + | + | - | - | + | + | + | - | + |
| Chinese cabbage [59] | + | + | - | - | - | + | + | - | - | + | + | + | - | + | + |
| Brassica napus | |||||||||||||||
| Swede [59] | - | + | - | - | + | - | + | - | - | + | + | + | + | + | + |
| Leaf rape [67] | - | + | - | + | - | + | + | + | - | + | + | + | - | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biondi, F.; Balducci, F.; Capocasa, F.; Visciglio, M.; Mei, E.; Vagnoni, M.; Mezzetti, B.; Mazzoni, L. Environmental Conditions and Agronomical Factors Influencing the Levels of Phytochemicals in Brassica Vegetables Responsible for Nutritional and Sensorial Properties. Appl. Sci. 2021, 11, 1927. https://0-doi-org.brum.beds.ac.uk/10.3390/app11041927
Biondi F, Balducci F, Capocasa F, Visciglio M, Mei E, Vagnoni M, Mezzetti B, Mazzoni L. Environmental Conditions and Agronomical Factors Influencing the Levels of Phytochemicals in Brassica Vegetables Responsible for Nutritional and Sensorial Properties. Applied Sciences. 2021; 11(4):1927. https://0-doi-org.brum.beds.ac.uk/10.3390/app11041927
Chicago/Turabian StyleBiondi, Francesca, Francesca Balducci, Franco Capocasa, Marino Visciglio, Elena Mei, Massimo Vagnoni, Bruno Mezzetti, and Luca Mazzoni. 2021. "Environmental Conditions and Agronomical Factors Influencing the Levels of Phytochemicals in Brassica Vegetables Responsible for Nutritional and Sensorial Properties" Applied Sciences 11, no. 4: 1927. https://0-doi-org.brum.beds.ac.uk/10.3390/app11041927