
Effects of Benzo[a]pyrene, Cortisol, and 17ß-Estradiol on Liver Microsomal EROD Activity of Anguilla anguilla: An In Vitro Approach


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Test Organisms
2.3. Preparation of Biological Material for In Vitro Assays
2.4. Microsomal In Vitro Exposure Conditions
2.4.1. In Vitro Effects of Cortisol, 17β-Estradiol, and B[a]P
2.4.2. In Vitro Effects of B[a]P after Microsomal Pre-Exposure to Steroid Hormones
2.5. EROD Activity Determination
2.6. Statistical Analysis
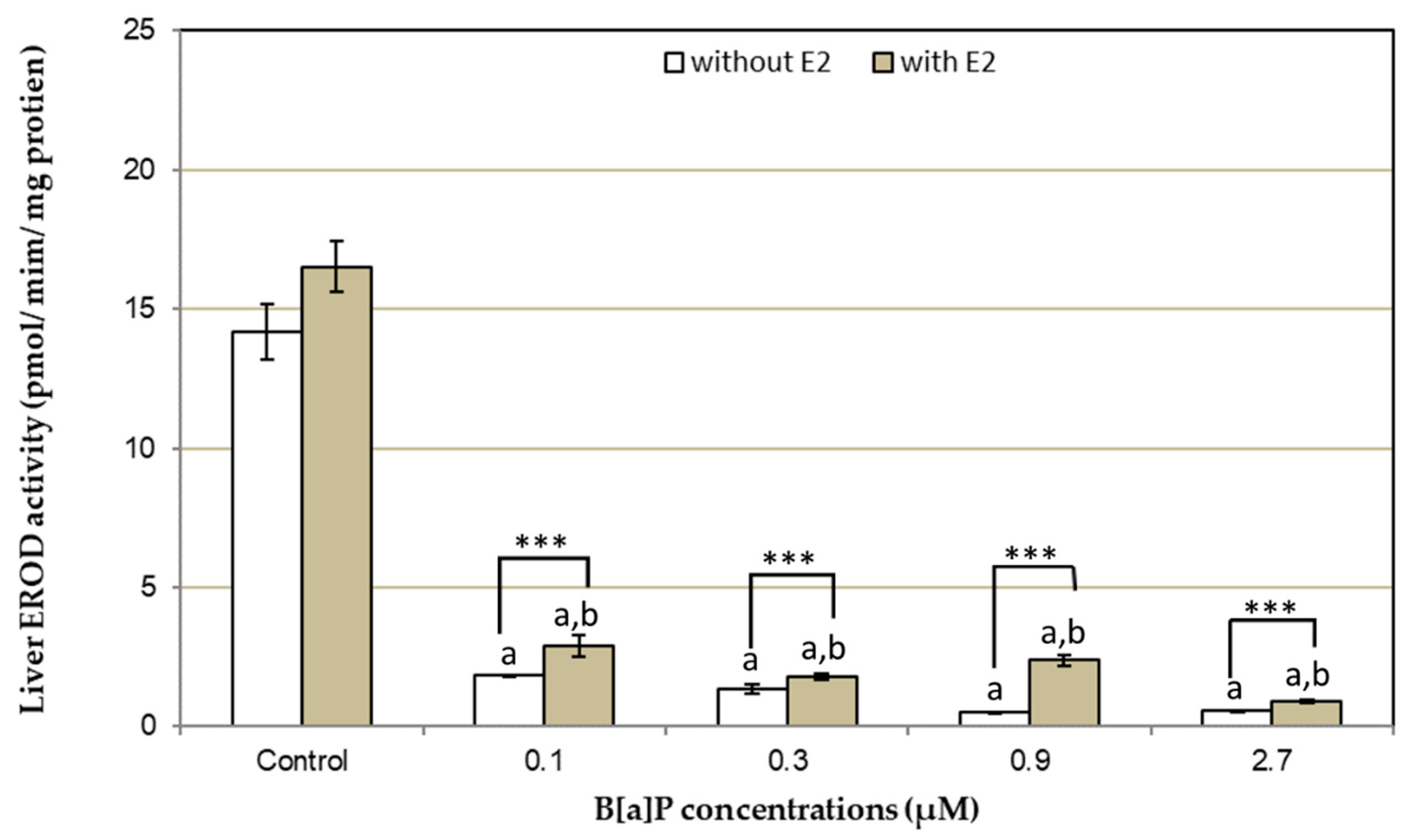
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bo, J.; Gopalakrishnan, J.; Chen, F.; Wang, K. Benzo[a]pyrene modulates the biotransformation, DNA damage and cortisol level of red sea bream challenged with lipopolysaccharide. Mar. Pollut. Bull. 2014, 85, 463–470. [Google Scholar] [CrossRef]
- Wolska, L.; Mechlińska, A.; Rogowska, J.; Namieśnik, J. Sources and fate of PAHs and PCBs in the marine environment. Crit. Rev. Environ. Sci. Technol. 2012, 42, 1172–1189. [Google Scholar] [CrossRef]
- Liao, K.; Yu, J.Z. Abundance and sources of benzo[a]pyrene and other PAHs in ambient air in Hong Kong: A review of 20-year measurements (1997–2016). Chemosphere 2020, 259, 127518. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.; Souza, M.; Junior, A.; Soares, L.; Frena, M.; Alexandre, M. Polycyclic aromatic hydrocarbons (PAH) in superficial water from a tropical estuarine system: Distribution, seasonal variations, sources and ecological risk assessment. Mar. Pollut. Bull. 2018, 127, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Knecht, A.; Truong, L.; Simonich, M.; Tanguay, R. Developmental benzo[a]pyrene (B[a]P) exposure impacts larval behavior and impairs adult learning in zebrafish. Neurotoxicol. Teratol. 2017, 59, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Logan, D.T. Perspective on ecotoxicology of PAHs to fish. Hum. Ecol. Risk Assess 2007, 13, 302–316. [Google Scholar] [CrossRef]
- Boehler, S.; Lörracher, A.; Schubert, J.; Braunbeck, T. Comparative live-imaging of in vivo EROD (ethoxyresorufin-O-deethylase) induction in zebrafish (Danio rerio) and fathead minnow (Pimephales promelas) embryos after exposure to PAHs and river sediment extracts. Sci. Total Environ. 2018, 621, 827–838. [Google Scholar] [CrossRef]
- Gagnon, M.; Rawson, C. Bioindicator species for EROD activity measurements: A review with Australian fish as a case study. Ecol. Indic. 2017, 73, 166–180. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.; Pacheco, M.; Santos, M.A. Organ specific antioxidant responses in golden grey mullet (Liza aurata) following a short-term exposure to phenanthrene. Sci. Total Environ. 2008, 396, 70–78. [Google Scholar] [CrossRef]
- Bucheli, T.D.; Fent, K. Induction of cytocrome P450 as a biomarker for environmental contamination in aquatic ecosystems. Crit. Rev. Environ. Sci. Technol. 1995, 25, 201–268. [Google Scholar] [CrossRef]
- Wunderlich, A.; Silva, R.; Zica, E.; Rebelo, M.; Parente, T.; Martínez, V. The influence of seasonality, fish size and reproductive status on EROD activity in Plagioscion squamosissimus: Implications for biomonitoring of tropical/subtropical reservoirs. Ecol. Indic. 2015, 58, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.; Pacheco, M.; Santos, M.A. Fish thyroidal and stress responses in contamination monitoring—An integrated biomarker approach. Ecotoxicol. Environ. Saf. 2011, 74, 1265–1270. [Google Scholar] [CrossRef] [PubMed]
- Reddam, A.; Magera, E.; Grosella, M.; McDonald, M. The impact of acute PAH exposure on the toadfish glucocorticoid stress response. Aquat. Toxicol. 2017, 192, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Randall, D.; Burggren, W.W.; French, K. Glands and Hormones—Physiological effects of hormones. In Animal Physiology—Mechanisms and Adaptations, 5th ed.; W. H. Freeman and Company: New York, NY, USA, 2001; Chapter 9; pp. 338–347. [Google Scholar]
- Oliveira, M.; Pacheco, M.; Santos, M.A. Cytochrome P4501A, genotoxic and stress responses in golden grey mullet (Liza aurata) following short-term exposure to phenanthrene. Chemosphere 2007, 66, 1284–1291. [Google Scholar] [CrossRef] [PubMed]
- Teles, M.; Santos, M.A.; Pacheco, M. Responses of European eel (Anguilla anguilla L.) in two polluted environments: In situ experiments. Ecotoxicol. Environ. Saf. 2004, 58, 373–378. [Google Scholar] [CrossRef]
- Teles, M.; Pacheco, M.; Santos, M.A. Sparus aurata L. liver EROD and GST activities, plasma cortisol, lactate, glucose and erythrocytic nuclear anomalies after short-term exposure either to 17ß-estradiol (E2) or E2 combined with 4-nonylphenol. Sci. Total Environ. 2005, 336, 57–69. [Google Scholar] [CrossRef]
- Teles, M.; Santos, M.A.; Pacheco, M. Biotransformation, stress and genotoxic effects of 17β-estradiol in juvenile sea bass (Dicentrarchus labrax L.). Environ. Int. 2006, 32, 470–477. [Google Scholar] [CrossRef]
- Barreto, A.; Carvalho, A.; Silva, D.; Pinto, E.; Almeida, A.; Paíga, P.; Correia-Sá, L.; Delerue-Matos, C.; Trindade, T.; Soares, A.M.V.M.; et al. Effects of single and combined exposures to gold (nano versus ionic form) and gemfibrozil ina liver organ culture of Sparus aurata. Mar. Pollut. Bull. 2020, 160, 111665. [Google Scholar] [CrossRef]
- Billiard, S.M.; Bols, N.C.; Hodson, P.V. In vitro and in vivo comparisons of fish-specific CYP1A induction relative potency factors for selected polycyclic aromatic hydrocarbons. Ecotoxicol. Environ. Saf. 2004, 59, 292–299. [Google Scholar] [CrossRef]
- Jain, A.; Singh, D.; Dubey, K.; Maurya, R.; Mittal, S.; Pandey, A. Chapter 3—Models and Methods for In Vitro Toxicity. In In Vitro Toxicology; Academic Press: Cambridge, MA, USA, 2018; pp. 45–65. [Google Scholar]
- Lange, U.; Danischewski, D.; Siebers, D. Regional variability and sexual differences in ethoxyresorufin-O-deethylase activities and cytochrome P450 concentration in the liver of mature dab (Limanda limanda L.) in the German Bight. In Variability of EROD and Cytochrome P450 in North Sea Dab; Braumbeck, T., Hanke, W., Segner, H., Eds.; VCH, Verlag Chemie: New York, NY, USA, 1993. [Google Scholar]
- Monod, G.; Vindimian, E. Effect of storage conditions and subcellular fractionation of fish and cytochrome P-450-dependent enzymatic activities used for the monitoring of water pollution. Water Res. 1991, 25, 173–177. [Google Scholar] [CrossRef]
- Pacheco, M.; Santos, M.A. Induction of liver EROD and erythrocytic nuclear abnormalities by cyclophosphamide and PAH’s in Anguilla anguilla L. Ecotoxicol. Environ. Saf. 1998, 40, 71–76. [Google Scholar] [CrossRef]
- Hyeon, J.; Hur, S.; Kim, B.; Byun, J.; Kim, E.; Lim, B.; Lee, B.; Kim, S.; Takemura, A.; Kim, S. Involvement of Estrogen and Its Receptors in Morphological Changes in the Eyes of the Japanese Eel, Anguilla japonica, in the Process of Artificially-Induced Maturation. Cells 2019, 8, 310. [Google Scholar] [CrossRef] [Green Version]
- Tirado, J.; Valladares, L.; Muñoz, D.; Caza, J.; Manjunatha, B.; Kundapur, R. Levels of 17β-estradiol, vitellogenin, and prostaglandins during the reproductive cycle of Oreochromis niloticus. Lat. Am. J. Aquat. Res. 2017, 45, 930–936. [Google Scholar] [CrossRef]
- Gornall, A.C.; Bardawill, C.J.; David, M.M. Determination of serum proteins by means of biuret reaction. J. Biol. Chem. 1949, 177, 751–766. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Oliveira, M.; Santos, M.A.; Pacheco, M. Heavy metal inhibition of Anguilla anguilla L. liver microsomal EROD activity and thiol protection. Fresen. Environ. Bull. 2005, 14, 59–64. [Google Scholar]
- Santos, M.A.; Maria, V.L. Abietic, Dehydroabietic acids and retene in vitro effets on Anguilla anguilla L. liver microsomal EROD activity. Fresen. Environ. Bull. 2005, 14, 698–702. [Google Scholar]
- Oliveira, M.; Santos, M.A.; Gravato, C.; Pacheco, M. Chromium effects on Anguilla anguilla liver organ culture. Fresen. Environ. Bull. 2003, 12, 349–352. [Google Scholar]
- Gravato, C.; Santos, M.A. Juvenile sea bass liver P450 EROD induction and erythrocytic genotoxic responses to PAH and PAH-like compounds. Ecotoxicol. Environ. Saf. 2002, 51, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Maria, V.L.; Gravato, C.; Correia, A.C.; Santos, M.A. Biotransformation and genotoxicity responses to PAHs in two teleost species. Fresen. Environ. Bull. 2002, 11, 609–615. [Google Scholar]
- Anyakora, C.; Ogbeche, A.; Palmer, P.; Coker, H. Determination of polynuclear aromatic hydrocarbons in marine samples of Siokolo Fishing Settlement. J. Chromatogr. A 2005, 1073, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Teles, M.; Pacheco, M.; Santos, M.A. Endocrine and metabolic responses of A. anguilla L. caged in a freshwater-wetland (Pateira de Fermentelos—Portugal). Sci. Total Environ. 2007, 372, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.A.; Pacheco, M.; Magalhães, I. Anguilla anguilla L. liver organ culture as a toxicological model: In vivo and in vitro EROD induction by β-naphthoflavone. Fresen. Environ. Bull. 2000, 9, 527–534. [Google Scholar]
- Teles, M.; Gravato, C.; Pacheco, M.; Santos, M.A. Juvenile sea bass biotransformation, genotoxic and endocrine responses to β-naphthoflanove, 4-nonylphenol and 17ß-estradiol individual and combined exposures. Chemosphere 2004, 57, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Vaccaro, E.; Meucci, V.; Intorre, L.; Soldani, G.; Di Bello, D.; Longo, V.; Gervasi, P.G.; Pretti, C. Effects of 17ß-estradiol, 4-nonylphenol and PCB 126 on the estrogenic activity and phase I and 2 biotransformation enzymes in male sea bass (Dicentrarchus labrax). Aquat. Toxicol. 2005, 75, 293–305. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, C.S.S.; Oliveira, M.; Santos, M.A.; Pacheco, M. Effects of Benzo[a]pyrene, Cortisol, and 17ß-Estradiol on Liver Microsomal EROD Activity of Anguilla anguilla: An In Vitro Approach. Appl. Sci. 2021, 11, 2533. https://0-doi-org.brum.beds.ac.uk/10.3390/app11062533
Ferreira CSS, Oliveira M, Santos MA, Pacheco M. Effects of Benzo[a]pyrene, Cortisol, and 17ß-Estradiol on Liver Microsomal EROD Activity of Anguilla anguilla: An In Vitro Approach. Applied Sciences. 2021; 11(6):2533. https://0-doi-org.brum.beds.ac.uk/10.3390/app11062533
Chicago/Turabian StyleFerreira, C.S.S., Miguel Oliveira, Maria Ana Santos, and Mário Pacheco. 2021. "Effects of Benzo[a]pyrene, Cortisol, and 17ß-Estradiol on Liver Microsomal EROD Activity of Anguilla anguilla: An In Vitro Approach" Applied Sciences 11, no. 6: 2533. https://0-doi-org.brum.beds.ac.uk/10.3390/app11062533