
Antioxidant Production in Dunaliella



1
School of Architecture, Building and Civil Engineering, Loughborough University, Epinal Way, Loughborough, Leicestershire LE11 3TU, UK
2
Algae Biotechnology Research Group, Faculty of Engineering and Science, University of Greenwich, Central Avenue, Chatham Maritime, Kent ME4 4TB, UK
*
Author to whom correspondence should be addressed.
Appl. Sci. 2021, 11(9), 3959; https://0-doi-org.brum.beds.ac.uk/10.3390/app11093959
Submission received: 7 April 2021
/
Revised: 22 April 2021
/
Accepted: 24 April 2021
/
Published: 27 April 2021
(This article belongs to the Special Issue Advances in Algal Biomass Applications)
Abstract
:Microalgae have become an attractive natural source of a diverse range of biomolecules, including enzymatic and non-enzymatic antioxidants; nevertheless, economically sustainable production of such compounds from microalgae biomass is still challenging. The main hurdles are: (a) increasing microalgae yield; (b) achieving optimal cultivation conditions; (c) energy-efficient and cost-effective downstream processing (extraction and purification); (d) optimal storage of post-processed antioxidant molecules. This review provides a detailed overview of enzymatic and non-enzymatic antioxidants in the cellular metabolism of the commercially important microalgae Dunaliella, industrial applications of antioxidant enzymes, strategies to enhanced antioxidant accumulation in cells, and the opportunities and limitations of current technologies for antioxidant enzymes production from microalgae biomass as an alternative to common microbial sources.
1. Introduction
Microalgae are singled celled micro-bio-factories capable of producing a wide variety of high-value compounds (carbohydrates, proteins, lipids, carotenoids, phycobiliproteins, phenolic, polyunsaturated fatty acids and antioxidants) used in the pharmaceutical, nutraceutical, cosmetic, and food processing industries [1,2,3,4,5,6,7,8,9,10]. Microalgal biomass can be added directly as a nutrient enhancer in animal feeds; as an enhancer for improving the quality of food; and as a stabiliser for maintaining the colour and flavour of food products [3,11,12,13,14]. The global microalgae market has gradually expanded, and worldwide sales are expected to exceed US $3.2 billion by the end of 2030 [15]. Recent studies have demonstrated the potential of microalgae to produce antioxidant molecules [16,17,18,19]. The antioxidant capacity and antioxidant activity of small molecules (β-carotene, astaxanthin, and phenolic compounds) have also been explored extensively [10,20,21]. Nevertheless, antioxidant enzymes have received little attention.
The green halotolerant microalgae Dunaliella can grow in a wide range of salt concentrations, from 0.05 M to 5.5 M NaCl [22,23]. Currently, 28 species of Dunaliella have been identified, of which 23 species live in saline environments, and 5 are rare species that inhabit freshwater [22]. Commercially, Dunaliella is cultivated in several countries, such as Australia, China, Israel, and India, with pilot-scale projects in Chile, Spain, Iran and Portugal [24,25,26] and considered one of the best sources of β carotene. Dunaliella has also been suggested as a sustainable source for industrial bioprocessing for the production of protein [27], biodiesel [4,28,29], colouring agent [30], and antioxidants [31].
Currently, antioxidant enzymes are sourced from microbial and animal sources [32,33]. Though there is no commercial production of antioxidant enzymes from Dunaliella, several reports have shown that Dunaliella can increase its levels of enzymatic (mainly catalase, peroxidase and superoxide dismutase) and non-enzymatic (carotenoids, phenolic, ascorbate and glutathione) antioxidants to mitigate stress induced by exogenous abiotic factors [31,34,35,36,37,38,39]. Its high adaptability to exogenous stressors (oxidative and osmosis) and the lack of a rigid cell wall [40] could make Dunaliella an ideal natural source for antioxidant production. Dunaliella can efficiently and sustainably produce large volumes of biomass without competing for cultivable land and fresh water [25].
2. Antioxidants
Extensive research on the effect of ROS in humans has demonstrated a substantial link between free radicals and more than sixty different health conditions, including ageing, cancer, diabetes, Alzheimer’s disease, strokes, heart attacks and atherosclerosis [41,42]. Consumption of higher levels of dietary antioxidant enzymes, as well as antioxidant molecules-enriched food or antioxidant supplements, has been found to reduce the risk of free radical-related health issues [41].
2.1. Classification of Antioxidants
Antioxidants (enzymatic or non-enzymatic) are classified depending on their mode of activity as primary antioxidants (hydrogen or electrons donors) or secondary antioxidants (oxygen scavengers or chelating agents) [43,44]. Antioxidants can also be grouped according to size, solubility, mode of action or structure, Figure 1. Abundant enzymatic antioxidants in microalgae are superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), glutathione peroxidase (GPX), glutathione reductase (GR), glutathione transferase (GST). Non-enzymatic antioxidants consist of compounds such as Vitamin C (ascorbate), glutathione, carotenoid, phenolic compounds, proline, glycine, polyamine, PUFA, and some metals (Cu, Zu) [45,46,47]. Most enzymatic antioxidants and some non-enzymatic antioxidants (glutathione, ascorbate) are hydrophilic and mainly present in the cellular fluids (cytosol or cytoplasmic matrix), whereas the hydrophobic antioxidants (carotenoid, tocopherol) are primary located in the cell membranes [48].
2.2. Antioxidant Enzymes
Antioxidant enzymes prevent or delay the oxidation of other molecules by neutralising reactive oxygen species (ROS) [48]. These enzymes eliminate ROS by reducing the energy of free radicals or by donating electrons to free radicals [42], and as such, constitute the first level of defence in the cells antioxidant network. Some molecules are not involved directly in the scavenging of free radicals but rather enhance other antioxidant molecules’ activity and may also be classified as antioxidants [41]. The majority of antioxidant enzymes are metalloenzymes and contain a metal ion in their catalytic site [45].
2.3. Commercial Applications of Antioxidant Enzymes
Antioxidants, or antioxidant-enriched extracts, are commercially used to prevent oxidative processes and to maintain the flavor, texture, and colour of food during storage. They also find uses as refining, bleaching, deodorising agents in the food processing industries [51,52,53,54,55], in extending the shelf life of lubricating oil and reducing vehicular emissions [56] and in stabilisation of synthetic fibre, rubber, thermoplastic, and adhesives by stopping autocatalytic reactions [57]. In cosmeceutical products, antioxidant compounds are used to prevent skin ageing and UV-induced skin damage, and treat the appearance of wrinkles and erythema due to inflammation [58,59,60,61]. Industrial applications of antioxidants are listed in Table 1. Global demand for antioxidants was valued at ~USD 2.25 billion in 2014 and grew at a CAGR (compounds annual growth rate) of ~5.5% between 2015 and 2020 [62]. This increasing global demand is driving the search for synthetic and natural-derived antioxidants.
Commercially, synthetic antioxidants such as butylated hydroxyl anisole (BHA), butylated hydroxyl toluene (BHT), α-tocopherol and propyl gallate are used in foods, food packaging, cosmetics and pharmaceutical products [82,84]. However, the physical properties of BHT and BHA (high volatility and instability at elevated temperature), strict legislation on the use of synthetic food additives, and the carcinogenic nature of some synthetic antioxidants [85,86,87,88] have shifted the attention to finding antioxidants from natural sources that are pharmacologically potent and have low or no toxicity.
2.4. Generation and Detoxification of ROS
ROS is a collective term for oxygen-derived products (free radicals and non-radicals reactive derivatives of oxygen). They are produced in cellular compartments either exogenously (in response to environmental stress such as UV radiation or xenobiotics) or endogenously (from the intracellular metabolic pathway, enzymatic activities, mitochondrial respiration, or photosynthesis) [66]. Accumulation of ROS leads to oxidative stress in cells and causes damage to cellular macromolecules, including proteins, lipids, carbohydrates and DNA [89]. Various sources of ROS and corresponding modes of biochemical metabolism are summarised in Table 2.
The majority of ROS are generated when electrons leak from the chloroplastic electron transport systems during photosynthesis, from the mitochondrial electron transport chain during photorespiration, and from the peroxisomal membrane electron transport chain (Figure 2) [90]. In plant cells, 1–2% of O2 consumption leads to the formation of superoxide (O2•−), and 1–5% of mitochondrial O2 consumption leads to the generation of H2O2 [45]. O2•− is generated during oxidisation of unsaturated fatty acids [91], from the activity of cytochrome P450 [92] and the cytochrome b5 family members [93,94]. Also, O2•− is produced in the peroxisome where xanthine oxidases catalyse the oxidation of xanthine and hypoxanthine to uric acid [45], and in the plasma membrane due to the reduction of O2 by NADPH oxidases [49].
Superoxide ions (O2•−) are converted into hydrogen peroxide (H2O2) by the catalytic activity of SOD. H2O2 is also produced in peroxisomes when glycolate from the photorespiration is recycled. In addition, H2O2 can be formed by D-amino acid oxidase, urate oxidase, flavin oxidase, L-α-hydroxy acid oxidase, and fatty acyl-CoA oxidase, and by cell wall peroxidases. Reactive OH• is produced from the reaction of O2•− and H2O2 at neutral pH and ambient temperature (Haber–Weiss reaction) or from H2O2 during the Fenton reaction [96]. In contrast, the addition of a proton (H+) to O2•− generates perhyroxyl radicals (HO2•). In some cases, intracellular ROS could also form during auto-oxidation of small molecules (epinephrine, flavins, and hydroquinones) [97,98]. The singlet oxygen (1O2) is generated from the reaction of oxygen (3O2) with the triplet state of chlorophyll produced by the dissipation of insufficient energy during photosynthesis [45].
Under normal physiological conditions, ROS is neutralised by the cells’ antioxidant systems where antioxidant enzymes and antioxidant molecules maintain the delicate intracellular redox balance and mitigate undesirable cellular damage caused by ROS, Table 2 [48,50].
Different isozymes of SOD exist (Mn-SOD in mitochondria and peroxisome, Fe-SOD in the chloroplast, and Cu/Zn-SOD isozyme in cytosol) but they all participate in scavenging of O2•−. In addition to SOD, some antioxidants molecules (Vitamin C, glutathione, etc.) also eliminate O2•− [45]. Further, the individual or cumulative catalytic activity of catalase or peroxidases decompose H2O2 into H2O and O2. CAT, peroxidases (GPX, APX) and SOD show a synergistic effect in the scavenging of O2•−. In addition to eliminating H2O2, GPX can protect cells by preventing intracellular lipid peroxidation [48]. APX may be more efficient compared to CAT or GPX in detoxification of H2O2 due to its higher affinity for H2O2. APX reduces H2O2 into H2O in chloroplasts, cytosol, mitochondria and peroxisomes, and in the apoplastic space using Ascorbic acid (ASc) as an electron donor [99].
Algal cells accumulate ASc with 30–40% remaining in the chloroplast. Ascorbic acid is water-soluble and acts as a potent antioxidant because of its ability to donate electrons in enzymatic and non-enzymatic reactions [100]. It protects cells by directly scavenging O2•−, HO2•− and regenerating the tocopherol from tocopheroxyl radicals [101]. All intracellular compartments generate the reduced form of glutathione, which plays a role as an excellent scavenger of many ROS such as O2•−, HO•, O3, NO2, lipid hydroperoxides [50] due to the redox-active thiol group that becomes oxidised when GSH reduces ROS [102]. Carotenoid also protects cells from light-induced oxidative stress by quenching 1O2 or dissipating excess heat (excitation energy) or scavenging peroxy radicals [45,48].
As the accumulation of enzymatic and non-enzymatic antioxidants in the cell depends on the external environment, manipulating cultivation conditions could enhance the intracellular antioxidant levels.
3. Cultivation Conditions
In addition to carbon, light energy, and water, Dunaliella requires certain mineral nutrients for growth. A suitable medium can be prepared from natural or artificial seawater enriched with nutrients (carbon, nitrogen, phosphorous, sulphur, iron, and magnesium), trace metals, and vitamins [40]. Carbon dioxide (CO2), inorganic carbon (e.g., NaHCO3, Na2CO3), and organic carbon (e.g., sodium acetate, glucose, glycerol) can be used as carbon sources. Nitrate, ammonia, and urea are commonly used as nitrogen sources for the synthesis of amino acids, nucleotides, chlorophylls and phycobilins [103]. Phosphorous is needed for several metabolic processes (ATP, DNA, RNA, and phospholipids). Iron acts as a cofactor for many enzymes (e.g., ferredoxins, catalases, nitrogenases, nitrates), and sulphur is needed for the biosynthesis of specific amino acids (cysteine, methionine). Magnesium is required for t chlorophyll biosynthesis, and other trace minerals serve as cofactors for various enzymes. Dunaliella needs light and temperature to assimilate carbon during photosynthesis and enhance biomass productivity and growth [104]. Growth conditions can be classified into three types based on energy and carbon sources (Table 3).
Photoautotrophic culturing is the most common strategy for growing Dunaliella biomass. However, in large cultures, cell-shading becomes an issue, limiting light penetration into the culture, leading to lower amounts of biomass [107]. In mixotrophic cultivation, the microalgae use CO2 and organic carbon (acetate, glucose) simultaneously along with light energy; respiratory and photosynthetic metabolism operates concurrently [105]. This increases biomass sproduction and results in a higher lipid production compared to that observed in photoautotrophic cultures [107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128], though at increased cultivation costs.
Although heterotrophic cultivation of algae [106] eliminates the requirements for light and facilitates biomass production in the dark, attempts to grow D. para heterotrophically were unsuccessful [110]. However, some Dunaliella strains can grow heterotrophically using organic carbon (such as tryptone, yeast extract, and urea, etc.) as an energy source, together with nutrients [128,129,130,131,132,133]. No studies have been published supporting the heterotrophic growth of Dunaliella strains at a commercial scale.
Dunaliella can grow phototrophically at temperatures ranging between 10 °C and 30 °C and in a wide variety of media, where the maximum cells density range between 0.3 × 106 cells mL−1 and 24 × 106 cells mL−1 [25]. In addition, these species are halotolerant and can grow in a high saline medium (0.5–4.0 M NaCl) [111]. Dunaliella strains can adapt to grow under quite different nutrients conditions, which demonstrates the diversity of these strains [112,113,114,115]. The essential photoautotrophic growth parameters of microalgae (Dunaliella) are shown in Figure 3.
Using available technologies, Dunaliella biomass production costs are approximately US $4.92 kg−1, which, for biodiesel production, is exceptionally high. Production costs could be reduced (US $3.05 kg−1) if biomass was produced using waste water as a nutrient source [116]. A biomass production costs of US$ 4.92 kg−1 would be acceptable if biomass was used for the production of high-value products (processed biomass value, US $123 kg−1) [117]; β-carotene (from Dunaliella spp.) and astaxanthin (from Haematococcus spp.) command a market value of $100–1000 kg−1 [117]. Moreover, the demand for astaxanthin, β-carotene and lutein is now emerging in the market. The global carotenoid market was valued at €1.3 billion in 2017 and is projected to reach €1.8 billion by 2022 [7]. Other high-value compounds such as antioxidant enzymes could make Dunaliella biotechnologies economically profitable. The current biomass productivity of D. salina lies between 0.75 g m−2 d−1 and 3.0 g m−2 d−1 (ash-free dry weight, AFDW) [118], depending on seasonality and other cultivation factors.
Variation of growth parameters disturbs intracellular metabolism via photosynthesis and photorespiration, which results in an imbalance between the generation and detoxification of ROS, ultimately stimulating enzymatic and non-enzymatic antioxidant responses. Therefore, modifications of cultivation factors may enhance antioxidants levels, making Dunaliella a competitive candidate for antioxidant (enzyme and non-enzyme) production.
4. Tuning Antioxidant Enzyme Activity in Dunaliella
There are two ways of inducing stress in Dunaliella during growth: biotic and abiotic manipulation [119]. Due to a lack of control over biotic stress, abiotic stress strategies are more sustainable. Abiotic stress conditions are classified as the nutrimental and physical conditions [105,120,121], which are introduced either during the inoculation of cells or during cell growth. Nutrimental factors include:
- i.
- Concentration and source nutrients (e.g., carbon, nitrogen, phosphorus, iron)
- ii.
- Concentration of trace elements (e.g., Zn2+, Cu2+)
- iii.
- Chemicals (e.g., phenol, H2O2)
- iv.
- Heavy metals (e.g., Hg2+, Cd2+)
Physical factors include:
- i.
- Irradiation level
- ii.
- Types of light (direct sunlight, artificial light sources (white, red, blue))
- iii.
- Temperature
- iv.
- pH
- v.
- Salinity
- vi.
- Size of inoculation
4.1. Salinity
Salinity induces osmotic stress, which has been found to affect antioxidant enzyme activity in Dunaliella species. Osmotic stress is divided into two types based on the salinity of the medium: hyposaline (salinity < 0.5 M) and hypersaline (salinity > 0.5 M).
CAT and APX activities were found to be highest when the cells were grown at a low salinity level (>0.5 M), with the activity decreasing with increased salinity. In contrast, SOD activity increased twofold at 1 M and threefold at 2 M compared to the activity at 0.5 M NaCl [122]. However, above 2 M NaCl, a decrease in the SOD activity was observed [122]. Above 2 M NaCl, accumulation of glycerol acts as an osmolyte for balancing high salinity-induced osmotic stress [123], which may be the reason for reduced enzymatic antioxidant activity. Similarly, CAT, APX, and GPX activity were found to decrease in D. tertiolecta concurrently with an increase in salt concentrations [124], perhaps due to the participation of glutathione (GSH) and ascorbate in the scavenging of H2O2 [124,125]. A higher GR activity in Dunaliella species (salina and tertiolecta) was observed at a low salt concentration (0.1 M NaCl) [124] which would increase the ratio of NADP +/NADPH, thereby ensuring the availability of NADP + to accept electrons resulting in a reduced flow of electrons to O2 for generation of ROS [126].
On the contrary, a study using D. tertiolecta demonstrated that CAT, SOD, DHAR, and GTX activity were unaltered under a wide range (0.05–3.00 M NaCl) of salinities, whilst APX activity increased by 200%, and MDHAR activity by 300% relative to the optimal growth salinity (0.5 M NaCl) [125]. Increased level of enzymatic activity in hypersaline condition may be due to the increase in the formation of ROS (peroxides) or increases in ATP synthesis via the Mehler-peroxidase reaction (which generates ATP by substituting dioxygen for carbon as the electron acceptor) [127]. Also, Dunaliella can retain a higher level of Mehler-peroxidase reaction at higher saline conditions [125].
The data above indicates that high salinity-induced osmotic stress is not an effective way to activate all antioxidant enzymes in Dunaliella strains. The above investigations also suggest that different Dunaliella strains, when exposed to external salinity, show different antioxidant enzyme activity due to differences in resistance and response. Hence, consideration of strain selection must be given if this approach is chosen.
4.2. Light Irradiation
D. salina exposed to UV-B irradiation enhanced CAT, SOD, POD activities, reaching maximum levels within three days after they levelled off [128], suggesting that CAT, SOD, POD activity could be part of the cell’s short-term adaptation against UV-B irradiation. Micosporine-like amino acids (shinorine, porphyra, and palythinol) could act as part of a supportive antioxidants network to protect cells against UV-B irradiation or ROS [129]. The synthesis of these compounds could be the reason for the levelling off enzymatic antioxidant response after three days of growth [128]. D. tertiolecta exposed to static and fluctuating natural UV radiation (400–700 nm) for short-term (1–3 days) or long-term (4–7 days) showed that only short-term irradiation significantly increased SOD and GR activity compared to non-UV irradiated cells. Glutathione content was reduced under both short and long-term irradiation, whilst APX activity did not significantly change, indicating that natural UV radiation-induced O2.− was mitigated by SOD, and that H2O2 was detoxified by the consumption of glutathione [130]. In contrast, D. salina treated with UV-B irradiation (irradiated cultures for 4 h) and cultured for 15 days increased its CAT, POD, and SOD activity compared to untreated cells [34]. Moreover, APX activity (140 µmol ascorbate mg−1 protein h−1) was found to increase in D. bardawil cells exposed to UV-A radiation as an addition to cultivation under high light energy (150 µmol photons m−2 s−1) compared to cells cultivated at low light intensities (35 µmol photons m−2 s−1). Increased APX activation under high light with UV-A radiation may be due to the direct effect of UV-A irradiance on the photosynthetic reactions that produce increased ROS within the chloroplast and leads to the increased enzymatic activity [131].
Exposing D. salina to extremely high light intensities (1000 µmol photons m−2 s−1) causes excess electron flow in electron transport chains of the photosystem, which leak electrons onto O2, thereby generating superoxide ion (O2•−) and results in enhancing SOD enzyme activity as the expression level of the antioxidant genes (SOD gene, Fe-SOD) increases [132]. No study has yet shown the effect of high light (>920 µmol photons m−2 s−1) on CAT activity in Dunaliella strains. Perhaps CAT is not involved in mitigating oxidative stress generated during acclimation of Dunaliella under high light intensities. Rather, peroxidases or other small antioxidant molecules (carotenoid) have been suggested to participate in the detoxification of ROS, as β-carotene content was found to increase when the algal was cultivated under increasing light intensities [133]. Therefore, CAT, POD, SOD and APX activity in Dunaliella can be improved by growing cells under UV radiation combined with a wide range of light irradiation (150–920 µmol photons m−2 s−1). However, further studies are needed to evaluate the effect of extreme high light-induced (e.g., 1500 µmol photons m−2 s−1) oxidative stress on antioxidant enzyme response.
4.3. Temperature
Temperature affects the microalgal growth rate and the biochemical compositions as the cells’ ability to assimilate nutrients are reduced [134], which can cause intracellular stress.
Cultivation at low temperature (13 °C) together with low light irradiation (20 µmol photons m−2 s−1) was found to enhance SOD, APX, MDHAR, DHAR activity in D. salina compared to the unstressed controls [135], whilst cultivation at a very low temperature (5 °C) decreased CAT activity [136]. Less carotenoid and ascorbate contents was also observed when the cells were cultivated at extremely low temperatures [135]. D. salina exposed to low temperature and cultivated under a high light intensity (100–1200 µmol photons m−2 s−1) was found to increase SOD, MDHAR, GR, APX and POD activity relative to unstressed cells. High light intensities provide saturated CO2, absorption and the low temperature decreased the rate of CO2 absorption, which induced photo-oxidative stress and resulted in increased enzymatic activity [35]. SOD, APX, DHAR, activity were increased in D. salina (IR-1) when cells were grown for 2 days under high temperature (28 °C) under light irradiation ranging between 100–1200 µmol photons m−2 s−1 compared to a different strain (Gh-U) [35]. The different oxidative stress responses between two strains could result from variations in resistance to low temperature and high light. Only a few studies describe the effect of temperature on antioxidant enzyme responses in Dunaliella, and further investigations are required to explore the effect of temperature on antioxidant enzyme activity.
4.4. Nutrients
Deprivation of nitrogen, sulphur, or phosphorous in the culture medium was found to enhance CAT, SOD, and APX activity compared to unstressed Dunaliella cells [37,137]. Deprivation of nitrogen reduces chlorophyll content and the synthesis of chloroplastic proteins, whereas deprivation of sulphur decreased the generation of carotenoid and increased ROS levels, which further increased antioxidant enzyme activity. Deprivation of Mn, Zn, Fe was also found to produce an oxidative stress response in D. salina with overexpressed SOD isoenzymes [36]. Nitrogen limitation in the growth medium elevated CAT, SOD, and APX levels in D. salina [138], indicating that depletion (limitation) of nitrogen also can play a pivotal role as an oxidative stressor for improving antioxidant enzyme activity. Research has also demonstrated that the deprivation of any single nutrient was more effective in enhancing antioxidant enzyme activity relative to combined nutrients deprivation conditions [137].
4.5. Metals
Heavy metals can induce oxidative stress in microalgae, by generating ROS by auto-oxidation, blocking essential functional groups in biomolecules, or substituting essential metal ions [139]. When D. tertiolecta was cultured with silver nanoparticle (AgNPs), CAT, SOD, and POD activity increased initially and then decreased over time [140], which indicate that antioxidant enzymes act as a first line of defence to protect the cells by mitigating metal-induced oxidative stress. Nonetheless, antioxidant enzyme activity could not eliminate the metal toxicity during prolonged exposure of cells [140].
Short-term exposer of Hg2+ increased APX activity in D. tertiolecta compared to unstressed cells. Hg2+ binds to a sulfhydryl group and disturbs the protein functions leading to stress conditions [141]. In contrast, long-term exposer to Hg2+ increased β-carotenoid content in Dunaliella cells. β-carotenoid acts as a supportive antioxidant. Pre-treatment of Dunaliella with Zn followed by cultivation in the presence of ROS generating agents (e.g., H2O2, paraquat) was found to decrease carotenoids level, CAT and APX activities [142], as cells were unable to enhance antioxidants levels. Moreover, the enhanced growth rate indicated that heavy metal toxicity in Zn-treated cells was detoxified by other antioxidants systems (either chelation reactions (metal to PC) or by displacements of Zn from a PC-Zn complex).
Small quantities of Cu2+ are essential for growing cells. During cultivation with Cu2+ (>5 µM CuCl2) oxidative stress was induced in Dunaliella cells [143]. D. salina and D. tertiolecta exposed to Cu2+ in the growth medium had enhanced APX activity. A higher enzymatic activity was measured in D. tertiolecta compared to D. salina, which could be due to the generation of lower amounts of carotenoid in D. tertiolecta [143]. Metals such as cobalt (Co), copper (Cu), chromium (Cr), iron (Fe), magnesium (Mg), manganese (Mn), molybdenum (Mo), nickel (Ni), selenium (Se), and zinc (Zn) are also essential micronutrients required for various biochemical and physiological functions during microalgae growth [144]. However, non-essential heavy metals such as arsenic (As), cadmium (Cd), chromium (Cr), lead (Pb), and mercury (Hg) are systematic toxicants [145]. Epidemiological and experimental studies have shown that these metals have adverse effects on health in humans, including cardiovascular diseases, developmental abnormalities, neurologic and neurobehavioral disorders, diabetes, hearing loss, hematologic and immunologic disorders, and various types of cancer, an association between exposure and cancer incidence in humans and animals [146]. Using heavy metals enriched biomass in the food industry or as a feed in the aquaculture industry is not a viable strategy to induce stress in algae cultures.
4.6. Chemicals
Chemicals modulate cellular metabolism and produce oxidative stress resulting in higher antioxidant levels in cells [147]. Culturing D. bardawil with a mixture of two surfactants (sodium dodecylbenzene sulfonate (SDBS) and cetyl trimethyl ammonium chloride (CTAC)) induced oxidative stress, increasing CAT and SOD activity compared to untreated cells [148].
The exposure of D. salina to 2-chlorophenol increased CAT, SOD, and GST activities to mitigate oxidative stress, which was produced by protein carbonyls formation through metal catalyzed oxidation reaction [149]. However, APX activity decreased when D. salina was cultured with 2-chlorophenol. Increased level of GST activity may be due to the ability of GST to catalyze the reduced glutathione in the presence of phenol [149]. Moreover, GST can also reduce lipid hydroperoxides to the corresponding alcohols [150]. Investigation of the effect of two pesticides on the antioxidant response in the culture of Dunaliella demonstrated that both pollutants (trichlorfon and dimehypo) at a low concentration induced oxidative stress that stimulated the activity of CAT compared to untreated cells [151]. CAT activity in D. salina was improved when cells were grown with herbicides (methylene blue and norflurazon) under low-temperature conditions [152]. Herbicides suppressed the synthesis of chlorophyll and decreased the carotenoid biosynthesis leading to an increase in CAT activity. Therefore, phenolic compounds or herbicides as stressors during Dunaliella culturing could be an effective strategy for enhancing enzyme activity.
4.7. Combination of Abiotic Factors on Antioxidant Enzyme Activity
Most research on antioxidant enzyme activity in Dunaliella focuses on only one factor, with a few studies assessing the combined effect of two factors. The studies indicate that a combination of factors is more effective in producing enzymatic antioxidant responses in Dunaliella compared to exposure to a single stressor [37,103,137]. Factors that can be combined to produce an antioxidant response include UV-B radiation + high salinity + nitrogen deficiency (enhanced CAT, SOD, POD activities).
UV-B radiation combined with high salinity and nitrogen deficiency enhanced CAT, SOD, POD activity relative to the non-radiated cells grown under normal growth conditions [34]. The combined effect of UV-B irradiation and nitrogen starvation can produce oxidative stress, which was eliminated by enhanced SOD, APX, and GPX activities relative to untreated D. salina cells, whilst GR activity did not significantly change. However, the combined effect on antioxidant enzyme activity was significantly lower relative to the effect of either UV-B radiation or nitrogen deprivation [37]. Therefore, the combination of UV-B irradiation and nitrogen deprivation may not be effective in enhancing antioxidant enzyme activity in Dunaliella. Salinity (2–3 M NaCl) associated with propyl gallate can enhance CAT and APX activity, whilst SOD activity was unaltered in D. salina compared to unstressed cells [152] suggests that salinity associated with other chemical-induced stress may increase in antioxidant enzyme activity in Dunaliella. There is a need for further research into this are to develop a deeper understanding of the ‘tuning parameters’ for enhanced enzymatic activity. The effects of abiotic factors on antioxidant enzyme activity are summarised in Table 4.
5. Culturing Systems
Commercially, large-scale microalgae biomass is produced in open ponds, raceway ponds, enclosed photobioreactors (PBRs), or a combination of hybrid systems [157]. The cultivation of Dunaliella also employs these various production systems [40,106,158,159,160,161] depending on the location and the desired end-products.
The PBR is considered a suitable production method for microalgae (Dunaliella) cultivation to enhance antioxidant production with biomass productivity in a ‘monoculture’ and to maintain culturing reproducibility. This method is flexible, controllable, and able to provide sustainable biomass production [105,162]. In open pond systems, the cell concentration is lower compared to that obtained during PBR cultivation as external factors (light irradiation, temperature, evaporation, contamination, aeration) are difficult to control. However, PBRs are not yet outperforming open pond systems in production volume and costs [40]. To the best of knowledge, no studies so far have reported the use of specific PBRs to culture Dunaliella strains to enhance enzymatic antioxidants and non-enzymatic antioxidants. The effect of the production system on antioxidants needs to be studied further.
6. Post-Culture Downstream Processing
High-cost downstream processing is one of the major challenges to making microalgae bio-refinery economically profitable [10,163]. Technological developments in harvesting, post-harvest storage, cell disruption, extraction, and possibly fractionation are needed. Harvesting the cells from the growth medium at the end of the culturing process is one of the most critical and challenging steps in mass Dunaliella culture [164].
6.1. Harvesting
The selection of appropriate harvesting techniques is dependent on the characteristics of the microalgae. Considerable challenges in harvesting Dunaliella cells are the lack of a rigid cell wall, its small size (length 5–25 µm), the low cell density (0.1%) in the culture medium, and the high salinity (2–3 M NaCl) of growth medium. Traditionally, harvesting procedures to concentrate microalgae cells are filtration, sedimentation, flocculation, flotation, centrifugation [163,164,165]. Several authors have extensively reviewed the harvesting of microalgae, and in the following section, a few key points are made with particular reference to Dunaliella [164,166,167,168].
6.2. Post-Harvesting Storage Processing
For Dunaliella to be a viable and sustainable natural antioxidant enzyme source, suitable drying and storage of biomass must be established. Optimized post-harvest storage conditions can minimise the loss of antioxidant enzyme activity and increase these valuable compounds’ shelf life. Cold storage is commonly used to preserve chemical, nutritional, and sensory properties of post-harvest processed living cells as respiration and other metabolic reactions are reduced at lower temperatures [169]. Biomass can be prepared for cold storage via a dry or a wet route. In the dry route, Dunaliella biomass is freeze-dried or air-dried, then stored at cold temperature. In contrast, in a wet route, biomass is preserved at cold temperature straightway [25]. Several reports demonstrate that antioxidant stability depends on the type of biomass, type of chemical components, length of storage time, storage temperature, and drying method [170,171,172,173]. Roy et al., (2020) compared the storages of both wet and freeze-dried algal biomass and suggested that CAT and SOD activity in freeze-dried D. tertiolecta biomass can be retained for eight months when stored −20 °C, whereas activities in wet biomass or crude extract remain unchanged for four months when stored at −80 °C [173,174,175]. Therefore, using wet biomass, either fresh or frozen, as feedstock may be an economically feasible method for antioxidant enzyme storage compared to dried cell storage. Extraction procedure of antioxidants from algae biomass depends on the desired products. For example, lipid-soluble antioxidants (e.g., carotenoids, tocopherol, flavonoids, ascorbate) are extracted by organic solvents (e.g., hexane, heptane, methyl tertiary burate, methanol, ethanol) with a mechanical treatment (e.g., ultrasonication, microwave, high pressure) [2,10,16,174]. Water-soluble antioxidants or antioxidant enzymes are recovered from the algae biomass by aqueous solvent or buffer systems together with mechanical assistance [37,128,138,175,176]. Recently, green solvents (e.g., cyclopentyl methyl ether (CPME), dimethyl carbonate (DMC)), ionic liquids (e.g., ethanol containing 1-n-butyl-3-methylimidazolium) and supercritical CO2—based extraction have been used for the recovery of carotenoids (e.g., β-carotene, astraxanthin) [7,177].
7. Challenges and Future Opportunities
Despite the potential of Dunaliella as a source for high-value compounds including antioxidants, the main challenges are as follows:
- i.
- ii.
- Transferring lab-scale optimised abiotic stress strategies to a commercial scale;
- iii.
- Seasonal and environmental variation in biomass production, and the control of competitors in open ponds cultivation systems;
- iv.
- Engineering developments of sustainable large-scale technologies (open raceways ponds and PBR) for Dunaliella and microalgae in general for efficient biomass and enzymatic antioxidant production;
- v.
- Developing sustainable downstream technologies (harvesting, extraction, and purification) for antioxidant enzymes production at commercial scale;
- vi.
- Establishing tools for proteomics, genomics, and metabolomics, and implement them to develop strains for the accumulation of intracellular antioxidant enzymes;
- vii.
- Stabilisation of antioxidant enzymes in food ingredients and extension of their shelf life;
- viii.
- Algae extracts can be used as taste enhances in foods; however, some strains are associated with a bitter or astringent taste;
- ix.
- Introducing a general platform to solve regulatory and labelling issues, as this varies from country to country.
Based on the reviewed literature, emphasis has been on carotenoid production from Dunaliella, limiting the exploration of Dunaliella biomass to produce other high-value compounds such as antioxidant enzymes. The tolerance of Dunaliella to extreme environments (such as high salinity and high light levels) makes these strains versatile. It enables them to survive in stressful environments, providing opportunities to explore these strains as natural antioxidant enzyme sources either as a main product or by-products. In addition, there is an opportunity to improve technological developments in harvesting biomass by employing cost-effective processes (such as flocculation or microbubbles techniques). Due to the lack of a rigid cell wall, the extraction process is cheaper relative to other algae cells or plants, which will reduce downstream processing costs. As antioxidant enzymes in wet Dunaliella biomass can be retained for one month when stored at −20 °C [175]. The development of an extraction procedure for antioxidant enzymes from wet biomass can save the energy costs required for drying the biomass.
Techno-economic assessment can be conducted to assure the profitability of developed cultivation and downstream processing systems to produce specific antioxidants enzymes or biorefinery strategies. There is also an opportunity to explore novel applications of Dunaliella-derived antioxidant enzymes in the pharmaceuticals, nutraceuticals, agricultural and cosmetic industries. Growing end-user applications for antioxidant enzymes and increasing end-user awareness may enhance the demand in the global market.
8. Conclusions
Dunaliella can accumulate a wide range of molecules, with a high value in the global food, aquaculture, animal feed, pharmaceutical, and nutraceutical industry. Currently, β-carotene production from D. salina is commercially cost-effective. High biomass production costs; lack of optimal cultivation conditions and culturing systems; technical difficulties in scaling–up; lack of cost-effective downstream processing technologies; and instability of antioxidant enzymes at room temperature are the major constraints to the commercialisation of Dunaliella-based antioxidant enzyme production. Tuning abiotic stress factors during cultivation can activate different antioxidant systems in Dunaliella. A multiple factors-based stress strategy is more effective relative to a single inducing stress factor during growth in a photobioreactor (considered monoculture) system, giving design flexibility and controllability.
Author Contributions
Conceptualisation U.K.R., B.V.N., J.J.M.; Writing—original manuscript preparation U.K.R.; Writing—review and editing U.K.R., B.V.N. and J.J.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hadi, M.R.; Shariati, M.; Afsharzadeh, S. Microalgal biotechnology: Carotenoid and glycerol production by the green algae Dunaliella isolated from the Gave-Khooni salt marsh, Iran. Biotechnol. Bioprocess Eng. 2008, 13, 540–544. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algae as production systems of bioactive compounds. Eng. Life Sci. 2015, 15, 160–176. [Google Scholar] [CrossRef]
- Camacho, F.; Macedo, A.; Malcata, F. Potential industrial applications and commercialization of microalgae in the functional food and feed industries: A short review. Mar. Drugs 2019, 17, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, R.A.; He, M.; Aftab, R.A.; Zheng, S.; Nagi, M.; Bakri, R.; Wang, C. Bioenergy application of Dunaliella salina SA 134 grown at various salinity levels for lipid production. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Gujar, A.; Cui, H.; Ji, C.; Kubar, S.; Li, R. Development, Production and Market Value of Microalgae Products. Appl. Microbiol. 2019, 5, 1–9. [Google Scholar] [CrossRef]
- Goiris, K.; Muylaert, K.; De Cooman, L. Microalgae as a Novel Source of Antioxidants for Nutritional Applications. Handb. Mar. Microalgae Biotechnol. Adv. 2015, 10, 160–180. [Google Scholar] [CrossRef]
- Marino, T.; Casella, P.; Sangiorgio, P.; Verardi, A.; Ferraro, A.; Hristoforou, E.; Molino, A.; Musmarra, D. Natural beta-carotene: A microalgae derivate for nutraceutical applications. Chem. Eng. Trans. 2020, 79, 103–108. [Google Scholar] [CrossRef]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for high-value products towards human health and nutrition. Mar. Drugs. 2019, 17, 304. [Google Scholar] [CrossRef] [Green Version]
- Raja, R.; Coelho, A.; Hemaiswarya, S.; Kumar, P.; Carvalho, I.S.; Alagarsamy, A. Applications of microalgal paste and powder as food and feed: An update using text mining tool. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 740–747. [Google Scholar] [CrossRef]
- Khanra, S.; Mondal, M.; Halder, G.; Tiwari, O.N.; Gayen, K.; Bhowmick, T.K. Downstream processing of microalgae for pigments, protein and carbohydrate in industrial application: A review. Food Bioprod. Process. 2018, 110, 60–84. [Google Scholar] [CrossRef]
- Figueira, F.D.S.; Crizel, T.D.M.; Silva, C.R.; Salas-Mellado, M.d.l.M. Elaboration of gluten-free bread enriched with the microalgae Spirulina platensis. Braz. J. Food Technol. 2011, 14, 308–316. [Google Scholar]
- Gouveia, L.; Oliveira, A.C. Microalgae as a raw material for biofuels production. J. Ind. Microbiol. Biotechnol. 2009, 36, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Fradique, M.; Batista, A.P.; Nunes, M.C.; Gouveia, L.; Bandarra, N.M.; Raymundo, A. Incorporation of Chlorella vulgaris and Spirulina maxima biomass in pasta products. Part 1: Preparation and evaluation. J. Sci. Food Agric. 2010, 90, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Rumin, J.; Nicolau, E.; de Oliveira, R.G.; Fuentes-Grünewald, C.; Picot, L. Analysis of Scientific Research Driving Microalgae Market Opportunities in Europe. Mar. Drugs 2020, 18, 264. [Google Scholar] [CrossRef] [PubMed]
- Transparency Market Research. Microalgae—Based Products Market (Product: Spirulina, Chlorella, Astaxanthin, β-Carotene, PUFA Fatty Acid, Lutein, and Others; Food and Beverages, Personal Care, Nutraceuticals and Dietary Supplements, Pharmaceuticals, Chemicals, Fuel, Animal Feed)—Global Industry Analysis, Sixe, Share, Growth, Trends, and Forecast, 2020–2030. 2020. Available online: https://www.transparencymarketresearch.com/about-us.html (accessed on 26 April 2021).
- Maadane, A.; Merghoub, N.; Ainane, T.; El Arroussi, H.; Benhima, R.; Amzazi, S.; Bakri, Y.; Wahby, I. Antioxidant activity of some Moroccan marine microalgae: Pufa profiles, carotenoids and phenolic content. J. Biotechnol. 2015, 215, 13–19. [Google Scholar] [CrossRef]
- Gangadhar, K.N.; Pereira, H.; Rodrigues, M.J.; Custódio, L.; Barreira, L.; Malcata, F.X.; Varela, J. Microalgae-based unsaponifiable matter as source of natural antioxidants and metal chelators to enhance the value of wet Tetraselmis chuii biomass. Open Chem. 2016, 14, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Assuncao, M.F.G.; Amaral, R.; Martins, C.B.; Ferreira, J.D.; Ressurreicao, S.; Santos, S.D.; Varejão, J.M.T.B.; Santos, L.M.A. Screening microalgae as potential sources of antioxidants. J. Appl. Phycol. 2017, 29, 865–877. [Google Scholar] [CrossRef]
- García, J.L.; de Vicente, M.; Galán, B. Microalgae, old sustainable food and fashion nutraceuticals. Microb. Biotechnol. 2017, 10, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangl, D.; Zedler, J.A.Z.; Rajakumar, P.D.; Martinez, E.M.R.; Riseley, A.; Włodarczyk, A.; Purton, S.; Sakuragi, Y.; Howe, C.J.; Jensen, P.E.; et al. Biotechnological exploitation of microalgae. J. Exp. Bot. 2015, 66, 6975–6990. [Google Scholar] [CrossRef] [Green Version]
- Villarruel-López, A.; Ascencio, F.; Nuño, K. Microalgae, a Potential Natural Functional Food Source—A Review. Polish J. Food Nutr. Sci. 2017, 67, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Jiang, J.G. Osmotic responses of Dunaliella to the changes of salinity. J. Cell. Physiol. 2009, 219, 251–258. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Avron, M. The biotechnology of cultivating the halotolerant alga Dunaliella. Trends Biotechnol. 1990, 8, 121–126. [Google Scholar] [CrossRef]
- Del Campo, J.A.; García-González, M.; Guerrero, M.G. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Shariati, M.; Hadi, M.R. Microalgal Biotechnology and Bioenergy in Dunaliella. In Progress in Molecular and Environmental Bioengineering—From Analysis and Modeling to Technology Applications; Carpi, A., Ed.; Intechopen: Rijeka, Croatia, 2011; Available online: https://www.intechopen.com/books/progress-in-molecular-and-environmental-bioengineering-from-analysis-and-modeling-to-technology-applications/microalgal-biotechnology-and-bioenergy-in-dunaliella (accessed on 26 April 2021). [CrossRef] [Green Version]
- Chen, J.; Wang, Y.; Benemann, J.R.; Zhang, X.; Hu, H.; Qin, S. Microalgal industry in China: Challenges and prospects. J. Appl. Phycol. 2016, 28, 715–725. [Google Scholar] [CrossRef]
- Sui, Y.; Vlaeminck, S.E. Dunaliella Microalgae for Nutritional Protein: An Undervalued Asset. Trends Biotechnol. 2020, 38, 10–12. [Google Scholar] [CrossRef]
- Liang, M.H.; Xue, L.L.; Jiang, J.G. Two-stage cultivation of Dunaliella tertiolecta with glycerol and triethylamine for lipid accumulation: A viable way to alleviate the inhibitory effect of triethylamine on biomass. Appl. Environ. Microbiol. 2019, 85, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhalamurugan, G.L.; Valerie, O.; Mark, L. Valuable bioproducts obtained from microalgal biomass and their commercial applications: A review. Environ. Eng. Res. 2018, 23, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Church, J.; Hwang, J.H.; Kim, K.T.; McLean, R.; Oh, Y.K.; Nam, B.; Joo, J.C.; Lee, W.H. Effect of salt type and concentration on the growth and lipid content of Chlorella vulgaris in synthetic saline wastewater for biofuel production. Bioresour. Technol. 2017, 243, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Widowati, I.; Zainuri, M.; Kusumaningrum, H.P.; Susilowati, R.; Hardivillier, Y.; Leignel, V.; Bourgougnon, N.; Mouget, J.L. Antioxidant activity of three microalgae Dunaliella salina, Tetraselmis chuii and Isochrysis galbana clone Tahiti. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bali, Indonesia, 2017; Volume 55, p. 012067. [Google Scholar]
- Singh, R.; Kumar, M.; Mittal, A.; Kumar, P. Microbial enzymes: Industrial progress in 21st century. 3 Biotech 2016, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Raveendran, S.; Kuruvilla, A.; Rebello, S. Applications of Microbial Enzymes in Food Industry. Food Technol. Biotechnol. 2018, 56, 16–30. [Google Scholar] [CrossRef]
- Hanaa, H.; El Baz, F.K.; El-Baroty, G.S. Production of Antioxidant by the Green Alga Dunaliella salina. Int. J. Agric. Biol. 2004, 6, 49–57. [Google Scholar]
- Haghjou, M.M.; Shariati, M.; Smirnoff, N. The effect of acute high light and low temperature stresses on the ascorbate-glutathione cycle and superoxide dismutase activity in two Dunaliella salina strains. Physiol. Plant. 2009, 135, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.K.; Moane, S.; Murray, P. Effect of macro- and micro-nutrient limitation on superoxide dismutase activities and carotenoid levels in microalga Dunaliella salina CCAP 19/18. Bioresour. Technol. 2013, 147, 23–28. [Google Scholar] [CrossRef]
- Al-Rashed, S.A.; Ibrahim, M.M.; El-Gaaly, G.A.; Al-Shehri, S.; Mostafa, A. Evaluation of radical scavenging system in two microalgae in response to interactive stresses of UV-B radiation and nitrogen starvation. Saudi J. Biol. Sci. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Baranwal, M.; Reddy, S.M. Antioxidant and cytotoxic activity of carotenes produced by Dunaliella salina under stress. Pharm. Biol. 2016, 54, 2269–2275. [Google Scholar] [CrossRef] [Green Version]
- Mirshekari, M.; Einali, A.; Valizadeh, J. Metabolic changes and activity pattern of antioxidant enzymes induced by salicylic acid treatment in green microalga Dunaliella salina under nitrogen deficiency. J. Appl. Phycol. 2019, 31, 1709–1719. [Google Scholar] [CrossRef]
- Hosseini Tafreshi, A.; Shariati, M. Dunaliella biotechnology: Methods and applications. J. Appl. Microbiol. 2009, 107, 14–35. [Google Scholar] [CrossRef]
- Rahman, K. Studies on free radicals, antioxidants, and co-factors. Clin. Interv. Aging 2007, 2, 219–236. [Google Scholar]
- Krishnamurthy, P.; Wadhwani, A. Antioxidant Enzymes and Human Health. In Antioxidant Enzyme; Intech: Rijeka, Croatia, 2012; Volume 1, pp. 3–18. Available online: https://www.intechopen.com/books/antioxidant-enzyme/antioxidant-enzymes-and-human-health (accessed on 26 April 2021). [CrossRef] [Green Version]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. Biomed. Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.K. Assay Guided Comparison for Enzymatic and Non-Enzymatic Antioxidant Activities with Special Reference to Medicinal Plants. In Antioxidant Enzyme; Intech: Rijeka, Croatia, 2012; Volume 14, pp. 381–400. Available online: https://www.intechopen.com/books/antioxidant-enzyme/assay-guided-comparison-for-enzymatic-and-non-enzymatic-antioxidant-activities-with-special-referenc (accessed on 26 April 2021). [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Mtaki, K.; Kyewalyanga, M.S.; Mtolera, M.S.P. Assessment of antioxidant contents and free radical-scavenging capacity of chlorella vulgaris cultivated in low cost media. Appl. Sci. 2020, 10, 8611. [Google Scholar] [CrossRef]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Oxidative damage and antioxidative system in algae. Toxicol. Rep. 2019, 6, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef] [Green Version]
- Kaushal, J.; Mehandia, S.; Singh, G.; Raina, A.; Arya, S.K. Catalase enzyme: Application in bioremediation and food industry. Biocatal. Agric. Biotechnol. 2018, 16, 192–199. [Google Scholar] [CrossRef]
- Gopal, R.K.; Elumalai, S. Industrial Production of Superoxide Dismutase (SOD): A Mini Review. J. Probiotics Health 2017, 5. [Google Scholar] [CrossRef]
- Pandey, V.P.; Awasthi, M.; Singh, S.; Tiwari, S.; Dwivedi, U.N. A Comprehensive Review on Function and Application of Plant Peroxidases. Biochem. Anal. Biochem. 2017, 6, 1–16. [Google Scholar] [CrossRef]
- Keokamnerd, T.; Acton, J.C.; Han, I.Y.; Dawson, P.L. Effect of commercial rosemary oleoresin preparations on ground chicken thigh meat quality packaged in a high-oxygen atmosphere. Poult. Sci. 2008, 87, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Hamid, M. Khalil-ur-Rehman Potential applications of peroxidases. Food Chem. 2009, 115, 1177–1186. [Google Scholar] [CrossRef]
- Canter, N. Use of antioxidants in automotive lubricants. Tribol. Lubr. Technol. 2008, 64, 12. [Google Scholar]
- Jegannathan, K.R.; Nielsen, P.H. Environmental assessment of enzyme use in industrial production-a literature review. J. Clean. Prod. 2013, 42, 228–240. [Google Scholar] [CrossRef] [Green Version]
- Godic, A.; Poljšak, B.; Adamic, M.; Dahmane, R. The role of antioxidants in skin cancer prevention and treatment. Oxid. Med. Cell. Longev. 2014, 2014, 860479. [Google Scholar] [CrossRef]
- Allemann, I.B.; Baumann, L. Antioxidants used in skin care formulations. Ski. Ther. Lett 2008, 13, 5–9. [Google Scholar]
- Stephenie, S.; Chang, Y.P.; Gnanasekaran, A.; Esa, N.M.; Gnanaraj, C. An insight on superoxide dismutase (SOD) from plants for mammalian health enhancement. J. Funct. Foods 2020, 68, 103917. [Google Scholar] [CrossRef]
- Bafana, A.; Dutt, S.; Kumar, S.; Ahuja, P.S. Superoxide dismutase: An industrial perspective. Crit. Rev. Biotechnol. 2011, 31, 65–76. [Google Scholar] [CrossRef]
- Beach, D. Antioxidants (Natural, and Synthetic) Market for Pharmaceuticals, Food & Beverages Sector, Feed Additives, Cosmetics Industry, and Other Applications: Global Industry Perspective, Comprehensive Analysis, and Forecast, 2014–2020, Marrket Research Store. 2016. Available online: http://www.marketresearchstore.com/report/antioxidants-market-for-pharmaceuticals-food-z38119 (accessed on 26 April 2021).
- Sasse, A.; Colindres, P.; Brewer, M.S. Effect of natural and synthetic antioxidants on the oxidative stability of cooked, frozen pork patties. J. Food Sci. 2009, 74, 1–6. [Google Scholar] [CrossRef]
- Schillaci, C.; Nepravishta, R.; Bellomaria, A. Antioxidants in food and pharmaceutical research. Albanian J. Pharm. Sci. 2014, 1, 15–25. [Google Scholar]
- Ito, N.; Hirose, M.; Fukushima, S.; Tsuda, H.; Shirai, T.; Tatematsu, M. Studies on antioxidants: Their carcinogenic and modifying effects on chemical carcinogenesis. Food Chem. Toxicol. 1986, 24, 1071–1082. [Google Scholar] [CrossRef]
- Thompson, D.C.; Trush, M.A. Studies on the mechanism of enhancement of butylated hydroxytoluene-induced mouse lung toxicity by butylated hydroxyanisole. Toxicol. Appl. Pharmacol. 1988, 96, 122–131. [Google Scholar] [CrossRef]
- Silaghi-dumitrescu, L.; Loghin, F. Evaluation of the possible endocrine disruptive effect of butylated hydroxyanisole, butylated hydroxytoluene and propyl gallate in immature female rats. Farmacia 2013, 61, 202–211. [Google Scholar]
- Thorat, I.D.; Jagtap, D.D.; Mohapatra, D.; Joshi, D.C.; Sutar, R.F.; Kapdi, S.S. Antioxidants, their properties, uses in food products and their legal implications. Int. J. Food Stud. 2013, 2, 81–104. [Google Scholar] [CrossRef]
- El Shafey, H.M.; Bahashwan, S.A.; Alghaithy, A.A.; Ghanem, S. Microbial superoxide dismutase enzyme as therapeutic agent and future gene therapy. Curr. Res. Technol. Educ. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 1, 435–443. [Google Scholar]
- Surai, P.; Surai, P.F. Antioxidant Systems in Poultry Biology: Superoxide Dismutase. Anim. Nutr. 2015, 1, 1–17. [Google Scholar] [CrossRef]
- Gridley, D.S.; Green, L.M.; Nelson, G.A.; Pecaut, M.J.; Slater, J.M. Therapeutic Utilities of SOD Mimetics: Cancer, Radiotherapy and SOD Mimetics. In Madame Curie Bioscience Database [Internet]; Landes Bioscience: Austin, TX, USA, 2013. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK6099/ (accessed on 26 April 2021).
- Miłek, J.; Wójcik, M.; Verschelde, W. Thermal stability for the effective use of commercial catalase. Polish J. Chem. Technol. 2014, 16, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Morón, Ú.M.; Castilla-Cortázar, I. Protection Against Oxidative Stress and “IGF-I Deficiency Conditions”. In Antioxidant Enzyme; Intechopen: Rijeka, Croatia, 2012; Available online: https://www.intechopen.com/books/antioxidant-enzyme/protection-against-oxidative-stress-and-igf-i-deficiency-conditions- (accessed on 26 April 2021). [CrossRef] [Green Version]
- Chenevix Trench, G.; Young, J.; Coggan, M.; Board, P. Glutathione S-transferase M1 and T1 polymorphisms: Susceptibility to colon cancer and age of onset. Carcinogenesis 1995, 16, 1655–1657. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, E.; Chan, G.P. Skin-whitening and skin-condition-improving effects of topical oxidized glutathione: A double- blind and placebo-controlled clinical trial in healthy women. Clin. Cosmet. Investig. Dermatol. 2014, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Barrita, J.; Sánchez, M. Antioxidant Role of Ascorbic Acid and His Protective Effects on Chronic Diseases. Oxidative Stress Chronic Degener. Dis. A Role Antioxid. 2013, 450–484. [Google Scholar] [CrossRef] [Green Version]
- Borek, C. Antioxidants and the prevention of hormonally regulated cancer. J. Men’s Health Gend. 2005, 2, 346–352. [Google Scholar] [CrossRef]
- Constantinou, C.; Papas, A.; Constantinou, A.I. Vitamin E and cancer: An insight into the anticancer activities of vitamin E isomers and analogs. Int. J. Cancer 2008, 123, 739–752. [Google Scholar] [CrossRef]
- Petrulea, M.; Muresan, A.; Duncea, I. Oxidative Stress and Antioxidant Status in Hypo- and Hyperthyroidism. In Antioxidant Enzyme; Intechopen: Rijeka, Croatia, 2012; Available online: https://www.intechopen.com/books/antioxidant-enzyme/oxidative-stress-and-antioxidant-status-in-hypo-and-hyperthyroidism (accessed on 26 April 2021). [CrossRef] [Green Version]
- Kaliora, A.C.; Dedoussis, G.V.Z.; Schmidt, H. Dietary antioxidants in preventing atherogenesis. Atherosclerosis 2006, 187, 1–17. [Google Scholar] [CrossRef]
- Sak, K. Cytotoxicity of dietary flavonoids on different human cancer types. Pharmacogn. Rev. 2014, 8, 122–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinayagam, R.; Xu, B. Antidiabetic properties of dietary flavonoids: A cellular mechanism review. Nutr. Metab. 2015, 12, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Wei, H.; He, B. Dietary flavonoids intake and the risk of coronary heart disease: A dose-response meta-analysis of 15 prospective studies. Thromb. Res. 2015, 135, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Hertog, M.G.L.; Feskens, E.J.M.; Kromhout, D.; Hertog, M.G.L.; Hollman, P.C.H.; Hertog, M.G.L.; Katan, M.B. Dietary antioxidant flavonoids and risk of coronary heart disease: The Zutphen Elderly Study. Lancet 1993, 342, 1007–1011. [Google Scholar] [CrossRef]
- Yaakob, Z.; Ali, E.; Zainal, A.; Mohamad, M.; Takriff, M.S. An overview: Biomolecules from microalgae for animal feed and aquaculture. J. Biol. Res. 2014, 21, 6. [Google Scholar] [CrossRef] [Green Version]
- Martins, D.A.; Custódio, L.; Barreira, L.; Pereira, H.; Ben-Hamadou, R.; Varela, J.; Abu-Salah, K.M. Alternative sources of n-3 long-chain polyunsaturated fatty acids in marine microalgae. Mar. Drugs 2013, 11, 2259–2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, W.S.; Mozaffarian, D.; Rimm, E.; Kris-Etherton, P.; Rudel, L.L.; Appel, L.J.; Engler, M.M.; Engler, M.B.; Sacks, F. Omega-6 fatty acids and risk for cardiovascular disease: A science advisory from the American Heart Association nutrition subcommittee of the council on nutrition, physical activity, and metabolism; council on cardiovascular nursing; and council on epidem. Circulation 2009, 119, 902–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundebye, A.-K.; Hove, H.; Måge, A.; Bohne, V.J.B.; Hamre, K. Levels of synthetic antioxidants (ethoxyquin, butylated hydroxytoluene and butylated hydroxyanisole) in fish feed and commercially farmed fish. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2010, 27, 1652–1657. [Google Scholar] [CrossRef]
- Dawidowicz, A.L.; Olszowy, M.; Jóźwik-Dolęba, M. Antagonistic Antioxidant Effect in Butylated Hydroxytoluene/Butylated Hydroxyanisole Mixture. J. Food Process. Preserv. 2015, 39, 2240–2248. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Cirulis, J.T.; Scott, J.A.; Ross, G.M. Management of oxidative stress by microalgae. Can. J. Physiol. Pharmacol. 2013, 91, 15–21. [Google Scholar] [CrossRef]
- Kaushik, D.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Bak, S.; Beisson, F.; Bishop, G.; Hamberger, B.; Höfer, R.; Paquette, S.; Werck-Reichhart, D. Cytochromes P450. Arab. B 2011, 9, e0144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Montalvo, A.; Sun, Y.; Diaz-Ruiz, A.; Ali, A.; Gutierrez, V.; Palacios, H.H.; Curtis, J.; Siendones, E.; Ariza, J.; Abulwerdi, G.A. Cytochrome b5 reductase and the control of lipid metabolism and healthspan. NPJ Aging Mech. Dis. 2016, 2, 16006. [Google Scholar] [CrossRef] [PubMed]
- Pruchniak, M.P.; Araźna, M.; Demkc, U. Biochemistry of oxidative stress. Adv. Exp. Med. Biol. 2016, 878, 9–19. [Google Scholar] [CrossRef]
- Barbusiński, K. Controversy Over Fenton Mechanism. Ecol. Chem. Eng. S 2009, 16, 347–358. [Google Scholar]
- Tatsumi, H.; Nakase, H.; Kano, K.; Ikeda, T. Mechanistic study of the autoxidation of reduced flavin and quinone compounds. J. Electroanal. Chem. 1998, 443, 236–242. [Google Scholar] [CrossRef]
- Vašková, J.; Vaško, L.; Kron, I. Oxidative Processes and Antioxidative Metaloenzymes. In Antioxidant Enzyme; Intechopen: Rijeka, Croatia, 2012; Volume 2, pp. 19–58. Available online: https://www.intechopen.com/books/antioxidant-enzyme/oxidative-processes-and-antioxidative-metaloenzymes (accessed on 26 April 2021). [CrossRef] [Green Version]
- Gupta, V.K.; Sharma, S.K. Plants as Natural Antioxidants. Indian J. Nat. Prod. Resour. 2006, 5, 326–334. [Google Scholar]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate peroxidase and catalase activities and their genetic regulation in plants subjected to drought and salinity stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Mallick, N.; Mohn, F.H. Reactive oxygen species: Response of algal cells. J. Plant Physiol. 2000, 157, 183–193. [Google Scholar] [CrossRef]
- Marí, M.; Morales, A.; Colell, A.; García-Ruiz, C.; Fernández-Checa, J.C. Mitochondrial glutathione, a key survival antioxidant. Antioxid. Redox Signal. 2009, 11, 2685–2700. [Google Scholar] [CrossRef] [Green Version]
- Jajic, I.; Sarna, T.; Strzalka, K. Senescence, Stress, and Reactive Oxygen Species. Plants 2015, 4, 393–411. [Google Scholar] [CrossRef] [Green Version]
- Juneja, A.; Ceballos, R.M.; Murthy, G.S. Effects of environmental factors and nutrient availability on the biochemical composition of algae for biofuels production: A review. Energies 2013, 6, 4607–4638. [Google Scholar] [CrossRef] [Green Version]
- Ben-Amotz, A.; Polle, J.E.W.; Rao, D.V.S. The Alga Dunaliella; CRC Press: Boca Raton, FL, USA, 2009; ISBN 1578085454. [Google Scholar]
- Perez-Garcia, O.; Escalante, F.M.E.; de-Bashan, L.E.; Bashan, Y. Heterotrophic cultures of microalgae: Metabolism and potential products. Water Res. 2011, 45, 11–36. [Google Scholar] [CrossRef] [PubMed]
- Benavente-Valdes, J.R.; Aguilar, C.; Contreras-Esquivel, J.C.; Méndez-Zavala, A.; Montañez, J. Strategies to enhance the production of photosynthetic pigments and lipids in chlorophycae species. Biotechnol. Rep. 2016, 10, 117–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.L.; Lee, S.M.; Choi, H.J. Integration of microalgal cultivation system for wastewater remediation and sustainable biomass production. World J. Microbiol. Biotechnol. 2016, 32. [Google Scholar] [CrossRef] [PubMed]
- Kharati-Koupaei, M.; Moradshasi, A. Effects of sodium nitrate and mixotrophic culture on biomass and lipid production in hypersaline microalgae Dunaliella viridis Teod. Braz. Arch. Biol. Technol. 2016, 59, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.Y.; Yeh, K.L.; Aisyah, R.; Lee, D.J.; Chang, J.S. Cultivation, photobioreactor design and harvesting of microalgae for biodiesel production: A critical review. Bioresour. Technol. 2011, 102, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Gibor, A. The culture of brine algae. Biol. Bull. 1956, 111, 223–229. [Google Scholar] [CrossRef]
- Gladue, R.M.; Maxey, J.E. Microalgal feeds for aquaculture. J. Appl. Phycol. 1994, 6, 131–141. [Google Scholar] [CrossRef]
- Ramos, A.A.; Polle, J.; Tran, D.; Cushman, J.C.; Jin, E.; Varela, J.C. The unicellular green alga Dunaliella salina Teod. as a model for abiotic stress tolerance: Genetic advances and future perspectives. Algae 2011, 26, 3–20. [Google Scholar] [CrossRef]
- Borowitzka, M.A.; Borowitzka, L.J. Micro-Algal Biotechnology; Cambridge University Press: Cambridge, UK, 1988; ISBN 0521323495. [Google Scholar]
- Herrero, C.; Cid, A.; Fabregas, J.; Abalde, J. Yields in biomass and chemical constituents of four commercially important marine microalgae with different culture media. Aquac. Eng. 1991, 10, 99–110. [Google Scholar] [CrossRef]
- Cifuentes, A.S.; Gonzalez, M.A.; Parra, O.O. The effect of salinity on the growth and carotenogenesis in two Chilean strains of Dunaliella salina Teodoresco. Biol. Res. 1996, 29, 227–236. [Google Scholar] [PubMed]
- Ibañez, E.; Cifuentes, A. Benefits of using algae as natural sources of functional ingredients. J. Sci. Food Agric. 2013, 93, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Tredici, M.R.; Rodolfi, L.; Biondi, N.; Bassi, N.; Sampietro, G. Techno-economic analysis of microalgal biomass production in a 1-ha Green Wall Panel (GWP®) plant. Algal Res. 2016, 19, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Singh, B. Algal biorefinery: An integrated approach for sustainable biodiesel production. Biomass Bioenergy 2019, 131, 105398. [Google Scholar] [CrossRef]
- Harvey, P.J.; Ben-Amotz, A. Towards a sustainable Dunaliella salina microalgal biorefinery for 9-cis β-carotene production. Algal Res. 2020, 50, 102002. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Paglia, G.; Magnúsdóttir, M.; Steinarsdóttir, E.A.; Gudmundsson, S.; Palsson, B.Ø.; Andrésson, Ó.S.; Brynjólfsson, S. Effects of abiotic stressors on lutein production in the green microalga Dunaliella salina. Microb. Cell Fact. 2014, 13, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minhas, A.K.; Hodgson, P.; Barrow, C.J.; Adholeya, A. A review on the assessment of stress conditions for simultaneous production of microalgal lipids and carotenoids. Front. Microbiol. 2016, 7, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Jha, B. Antioxidant response of the microalga Dunaliella salina under salt stress. Bot. Mar. 2011, 54, 195–199. [Google Scholar] [CrossRef]
- Mishra, A.; Mandoli, A.; Jha, B. Physiological characterization and stress-induced metabolic responses of Dunaliella salina isolated from salt pan. J. Ind. Microbiol. Biotechnol. 2008, 35, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Tammam, A.A.; Fakhry, E.M.; El-sheekh, M. Effect of salt stress on antioxidant system and the metabolism of the reactive oxygen species in Dunaliella salina and Dunaliella tertiolecta. J. Biotechnol. 2011, 10, 3795–3808. [Google Scholar] [CrossRef]
- Jahnke, L.S.; White, A.L. Long-term hyposaline and hypersaline stresses produce distinct antioxidant responses in the marine alga Dunaliella tertiolecta. J. Plant Physiol. 2003, 160, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]
- Tian, J.; Yu, J. Changes in ultrastructure and responses of antioxidant systems of algae (Dunaliella salina) during acclimation to enhanced ultraviolet-B radiation. J. Photochem. Photobiol. B Biol. 2009, 97, 152–160. [Google Scholar] [CrossRef]
- Shick, J.M.; Dunlap, W.C. Mycosporine-like amino acids and related gadusols: Biosynthesis, acumulation, and UV-protective functions in aquatic organisms. Annu. Rev. Physiol. 2002, 64, 223–262. [Google Scholar] [CrossRef] [Green Version]
- Janknegt, P.J.; De Graaff, C.M.; Van De Poll, W.H.; Visser, R.J.W.; Helbling, E.W.; Buma, A.G.J. Antioxidative responses of two marine microalgae during acclimation to static and fluctuating natural uv radiation. Photochem. Photobiol. 2009, 85, 1336–1345. [Google Scholar] [CrossRef]
- White, A.L.; Jahnke, L.S. Contrasting effects of UV-A and UV-B on photosynthesis and photoprotection of beta-carotene in two Dunaliella spp. Plant Cell Physiol. 2002, 43, 877–884. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Polle, J.E.W.; Melis, A.; Lee, T.K.; Jin, E. Up-regulation of photoprotection and PSII-repair gene expression by irradiance in the unicellular green alga Dunaliella salina. Mar. Biotechnol. 2006, 8, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Lamers, P.P.; Van De Laak, C.C.W.; Kaasenbrood, P.S.; Lorier, J.; Janssen, M.; De Vos, R.C.H.; Bino, R.J.; Wijffels, R.H. Carotenoid and fatty acid metabolism in light-stressed Dunaliella salina. Biotechnol. Bioeng. 2010, 106, 638–648. [Google Scholar] [CrossRef] [PubMed]
- Ras, M.; Steyer, J.P.; Bernard, O. Temperature effect on microalgae: A crucial factor for outdoor production. Rev. Environ. Sci. Biotechnol. 2013, 12, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Haghjou, M.M.; Shariati, M.; Pozveh, M.H. The effect of low light intensities on oxidative stress induced by short-term chilling in Dunaliella salina teod. Pak. J. Biol. Sci. 2006, 9, 2048–2054. [Google Scholar] [CrossRef]
- Alizadeh, G.I.; Aliev, I.I.; Magerramova, K.K.; Galandarli, I.Z.; Dibirova, G.H.; Jalilova, A.R. The response reaction of Dunaliella cells against the influence of Methylene blue and Norflurazon under the low temperature stress conditions. Eur. J. Biotechnol. Biosci. 2015, 3, 7–10. [Google Scholar]
- Lv, H.; Cui, X.; Wahid, F.; Xia, F.; Zhong, C.; Jia, S. Analysis of the physiological and molecular responses of Dunaliella salina to macronutrient deprivation. PLoS ONE 2016, 11, e0152226. [Google Scholar] [CrossRef] [PubMed]
- Yilancioglu, K.; Cokol, M.; Pastirmaci, I.; Erman, B.; Cetiner, S. Oxidative Stress Is a Mediator for Increased Lipid Accumulation in a Newly Isolated Dunaliella salina Strain. PLoS ONE 2014, 9, e91957. [Google Scholar] [CrossRef]
- Michalak, a Phenolic Compounds and Their Antioxidant Activity in Plants Growing under Heavy Metal Stress. Plant Cell 2006, 15, 523–530.
- Hazani, A.A.; Ibrahim, M.M.; Shehata, A.I.; El-Gaaly, G.A.; Daoud, M.; Fouad, D.; Rizwana, H.; Moubayed, N. Ecotoxicity of Ag-nanoparticles on two microalgae, Chlorella vulgaris and Dunaliella tertiolecta. Arch. Biol. Sci. 2013, 65, 1447–1457. [Google Scholar] [CrossRef]
- Zamani, N.; Rasekh, F.; Moradshahi, A.; Kholdebarin, B. Physiological responses of Dunaliella tertiolecata to Hg2+-induced oxidative stress. Iran. J. Sci. Technol. 2009, 33, 65–74. [Google Scholar]
- Tsuji, N.; Hirayanagi, N.; Okada, M.; Miyasaka, H.; Hirata, K.; Zenk, M.H.; Miyamoto, K. Enhancement of tolerance to heavy metals and oxidative stress in Dunaliella tertiolecta by Zn-induced phytochelatin synthesis. Biochem. Biophys. Res. Commun. 2002, 293, 653–659. [Google Scholar] [CrossRef]
- Nikookar, K.; Moradshahi, A.; Hosseini, L. Physiological responses of Dunaliella salina and Dunaliella tertiolecta to copper toxicity. Biomol. Eng. 2005, 22, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Sarkany, N.; Cui, Y. Biomass and lipid productivities of Chlorella vulgaris under autotrophic, heterotrophic and mixotrophic growth conditions. Biotechnol. Lett. 2009, 31, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.S.; Dietz, K.J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2009, 14, 43–50. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. Mol. Clin. Environ. Toxicol. 2012, 101, 133–164. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Chen, L.; Zhang, W. Chemicals to enhance microalgal growth and accumulation of high-value bioproducts. Front. Microbiol. 2015, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Qv, X.Y.; Jiang, J.G. Toxicity evaluation of two typical surfactants to Dunaliella bardawil, an environmentally tolerant alga. Environ. Toxicol. Chem. 2013, 32, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Mofeed, J.; Abdel-aal, E.I. Effect of Phenol on Some Antioxidant Enzymes in the Marine Microalga Dunaliella salina. J. Environ. Sci. (JOESE 5) 2015, 44, 185–196. [Google Scholar]
- Tappel, A.L. Glutathione peroxidase and hydroperoxides. Methods Enzymol. 1978, 52, 506–513. [Google Scholar]
- Chen, H.; Jiang, J.G. Toxic effects of chemical pesticides (trichlorfon and dimehypo) on Dunaliella salina. Chemosphere 2011, 84, 664–670. [Google Scholar] [CrossRef]
- Einali, A.; Valizadeh, J. Propyl gallate promotes salt stress tolerance in green microalga Dunaliella salina by reducing free radical oxidants and enhancing β-carotene production. Acta Physiol. Plant. 2015, 37, 1–11. [Google Scholar] [CrossRef]
- Nikookar, K.; Moradshahi, A.; Kharati, M. Influence of Salinity on the Growth, Pigmentation and Ascorbate Peroxidase Activity of Dunaliella Lake in Shiraz. Iran. J. Sci. Technol. 2004, 28, 117–125. [Google Scholar]
- Marcano, L.B.C.; Carruyo, I.M.; Montiel, X.M.; Morales, C.B.; De Soto, P.M. Effect of cadmium on cellular viability in two species of microalgae (Scenedesmus sp. and Dunaliella viridis). Biol. Trace Elem. Res. 2009, 130, 86–93. [Google Scholar] [CrossRef]
- Arun, N.; Vidyalaxmi, V.; Singh, D.P. Chromium (VI) induced oxidative stress in halotolerant alga Dunaliella salina and D. tertiolecta isolated from sambhar salt lake of Rajasthan (India). Cell. Mol. Biol. 2014, 60, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Tsiaka, P.; Tsarpali, V.; Ntaikou, I.; Kostopoulou, M.N.; Lyberatos, G.; Dailianis, S. Carbamazepine-mediated pro-oxidant effects on the unicellular marine algal species Dunaliella tertiolecta and the hemocytes of mussel Mytilus galloprovincialis. Ecotoxicology 2013, 22, 1208–1220. [Google Scholar] [CrossRef] [PubMed]
- Novoveská, L.; Ross, M.E.; Stanley, M.S.; Pradelles, R.; Wasiolek, V.; Sassi, J.F. Microalgal carotenoids: A review of production, current markets, regulations, and future direction. Mar. Drugs 2019, 17, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sierra, E.; Acién, F.G.; Fernández, J.M.; García, J.L.; González, C.; Molina, E. Characterization of a flat plate photobioreactor for the production of microalgae. Chem. Eng. J. 2008, 138, 136–147. [Google Scholar] [CrossRef]
- García-González, M.; Moreno, J.; Manzano, J.C.; Florencio, F.J.; Guerrero, M.G. Production of Dunaliella salina biomass rich in 9-cis-??-carotene and lutein in a closed tubular photobioreactor. J. Biotechnol. 2005, 115, 81–90. [Google Scholar] [CrossRef]
- Zhu, Y.H.; Jiang, J.G. Continuous cultivation of Dunaliella salina in photobioreactor for the production of β-carotene. Eur. Food Res. Technol. 2008, 227, 953–959. [Google Scholar] [CrossRef] [Green Version]
- Kleinegris, D.M.M.; Janssen, M.; Brandenburg, W.A.; Wijffels, R.H. Continuous production of carotenoids from Dunaliella salina. Enzym. Microb. Technol. 2011, 48, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Ndukwe, G.I.; Mustapha, K.B.; Ayo, R.I. Constraints to large scale algae biomass production and utilization. J. Algal Biomass Util. 2012, 3, 14–32. [Google Scholar]
- Kim, J.; Yoo, G.; Lee, H.; Lim, J.; Kim, K.; Kim, C.W.; Park, M.S.; Yang, J.W. Methods of downstream processing for the production of biodiesel from microalgae. Biotechnol. Adv. 2013, 31, 862–876. [Google Scholar] [CrossRef] [PubMed]
- Milledge, J.J.; Heaven, S. A review of the harvesting of micro-algae for biofuel production. Rev. Environ. Sci. Biotechnol. 2013, 12, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.K.; Garg, S.; Li, Y.; Malekizadeh, A.; Schenk, P.M. Critical analysis of current Microalgae dewatering techniques. Biofuels 2013, 4, 397–407. [Google Scholar] [CrossRef]
- Japar, A.S.; Takriff, M.S.; Yasin, N.H.M. Harvesting microalgal biomass and lipid extraction for potential biofuel production: A review. J. Environ. Chem. Eng. 2017, 5, 555–563. [Google Scholar] [CrossRef]
- Roy, M.; Mohanty, K. A comprehensive review on microalgal harvesting strategies: Current status and future prospects. Algal Res. 2019, 44, 101683. [Google Scholar] [CrossRef]
- Najjar, Y.S.; Abu-Shamleh, A. Harvesting of microalgae by centrifugation for biodiesel production: A review. Algal Res. 2020, 51, 102046. [Google Scholar] [CrossRef]
- Gao, S.; Yan, R.; Chen, F. Effects of Different Storage Temperatures and Times on Germination and Antioxidant Responses of Jatropha curcas L. Seeds. J. Agric. Sci. Technol. 2015, 17, 1619–1628. [Google Scholar]
- Ahmed, F.; Li, Y.; Fanning, K.; Netzel, M.; Schenk, P.M. Effect of drying, storage temperature and air exposure on astaxanthin stability from Haematococcus pluvialis. Food Res. Int. 2015, 74, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Trung, T.S.; Huyen, N.T.K.; Minh, N.C.; Le Trang, T.T.; Han, N.T. Optimization of harvesting of microalgal Thalassiosira pseudonana biomass using chitosan prepared from shrimp shell waste. Asian J. Agric. Res. 2016, 10, 162–174. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.S.; Rufino, M.S.; Moura, C.F.; Cavalcanti, F.R.; Alves, R.E.; Miranda, M.R. The influence of processing and long-term storage on the antioxidant metabolism of acerola (Malpighia emarginata) purée. Rev. Bras. Frutic. 2011, 30, 398–408. [Google Scholar] [CrossRef]
- Mediani, A.; Abas, F.; Tan, C.; Khatib, A. Effects of Different Drying Methods and Storage Time on Free Radical Scavenging Activity and Total Phenolic Content of Cosmos Caudatus. Antioxidants 2014, 3, 358–370. [Google Scholar] [CrossRef] [Green Version]
- Roy, U.K.; Nielsen, B.V.; Milledge, J.J. Effect of post-harvest conditions on antioxidant enzyme activity in Dunaliella tertiolecta biomass. Biocatal. Agric. Biotechnol. 2020, 27, 101661. [Google Scholar] [CrossRef]
- Xu, Y.; Ibrahim, I.M.; Wosu, C.I.; Ben-Amotz, A.; Harvey, P.J. Potential of new isolates of Dunaliella salina for natural β-carotene production. Biology 2018, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.R.; Rao, P.H.; Subramanian, V.V.; Sivasubramanian, V. Enzymatic and non-enzymatic antioxidant potentials of Chlorella vulgaris grown in effluent of a confectionery industry. J. Food Sci. Technol. 2014, 51, 322–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miȩkus, N.; Iqbal, A.; Marszałek, K.; Puchalski, C.; Świergiel, A. Green chemistry extractions of carotenoids from daucus carota L.-Supercritical carbon dioxide and enzyme-assisted methods. Molecules 2019, 24, 4339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavka, A.; Jornsson, L.J. Comparison of the growth of filamentous fungi and yeasts in lignocellulose-derived media. Biocatal. Agric. Biotechnol. 2014, 3, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Gomez, J.A.; Höffner, K.; Barton, P.I. From sugars to biodiesel using microalgae and yeast. Green Chem. 2016, 18, 461–475. [Google Scholar] [CrossRef] [Green Version]
- Munir, N.; Sharif, N.; Naz, S.; Farkhanda, M. Algae: A potent antioxidant source. Sky J. Microbiol. Res. 2013, 1, 22–31. [Google Scholar]

Figure 2.
Generation of ROS by energy transfer and during intracellular metabolic pathways in microalgae (FD; ferredoxin, FNR; ferredoxin-NADP+ oxidoreductase; PSI-Photosystem I; PSII-Photosystem II, PC-plastocyanin [90,95]).

Figure 3.
Phototrophic growth parameters essential for Dunaliella.


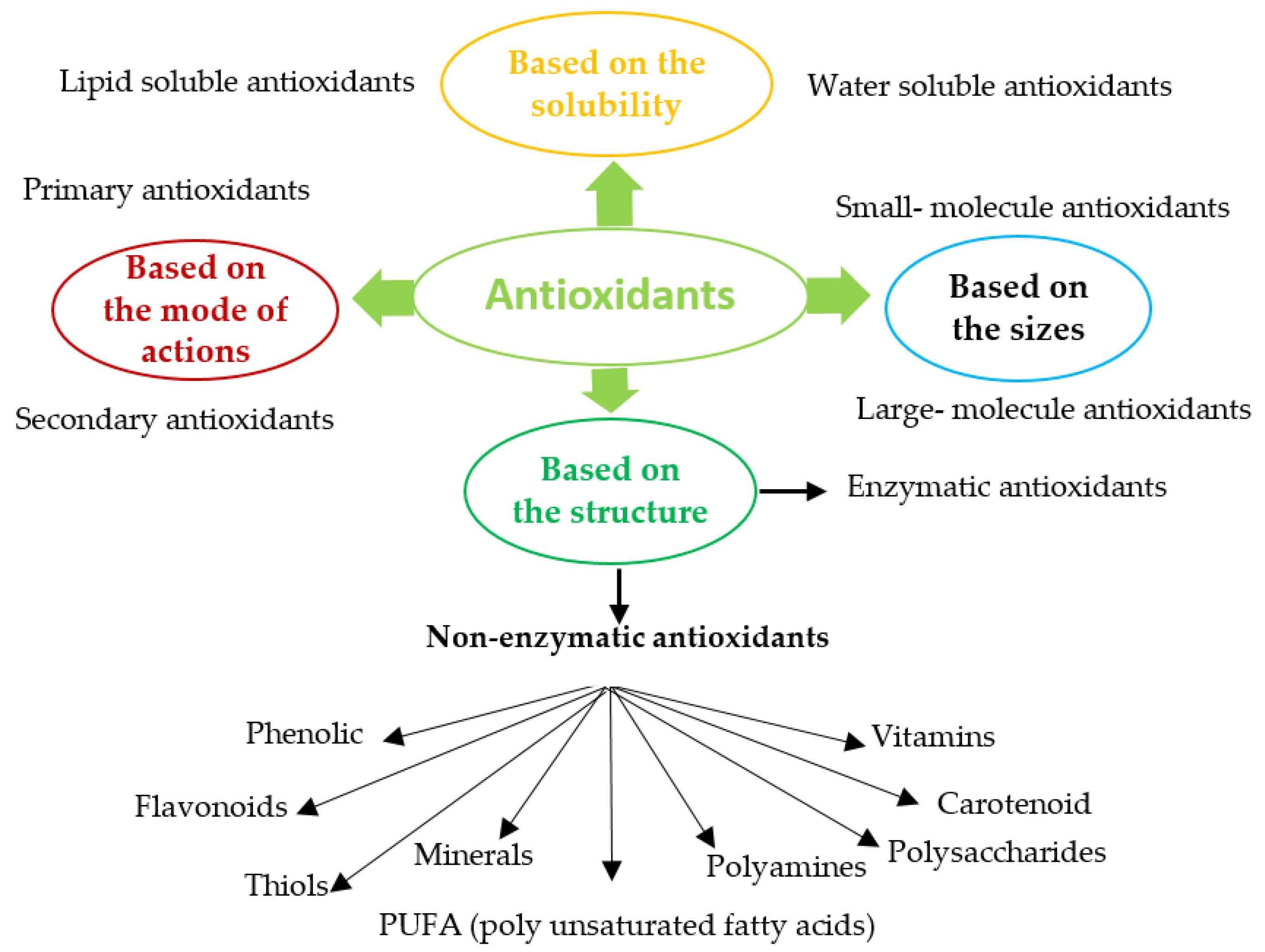{kind=link}
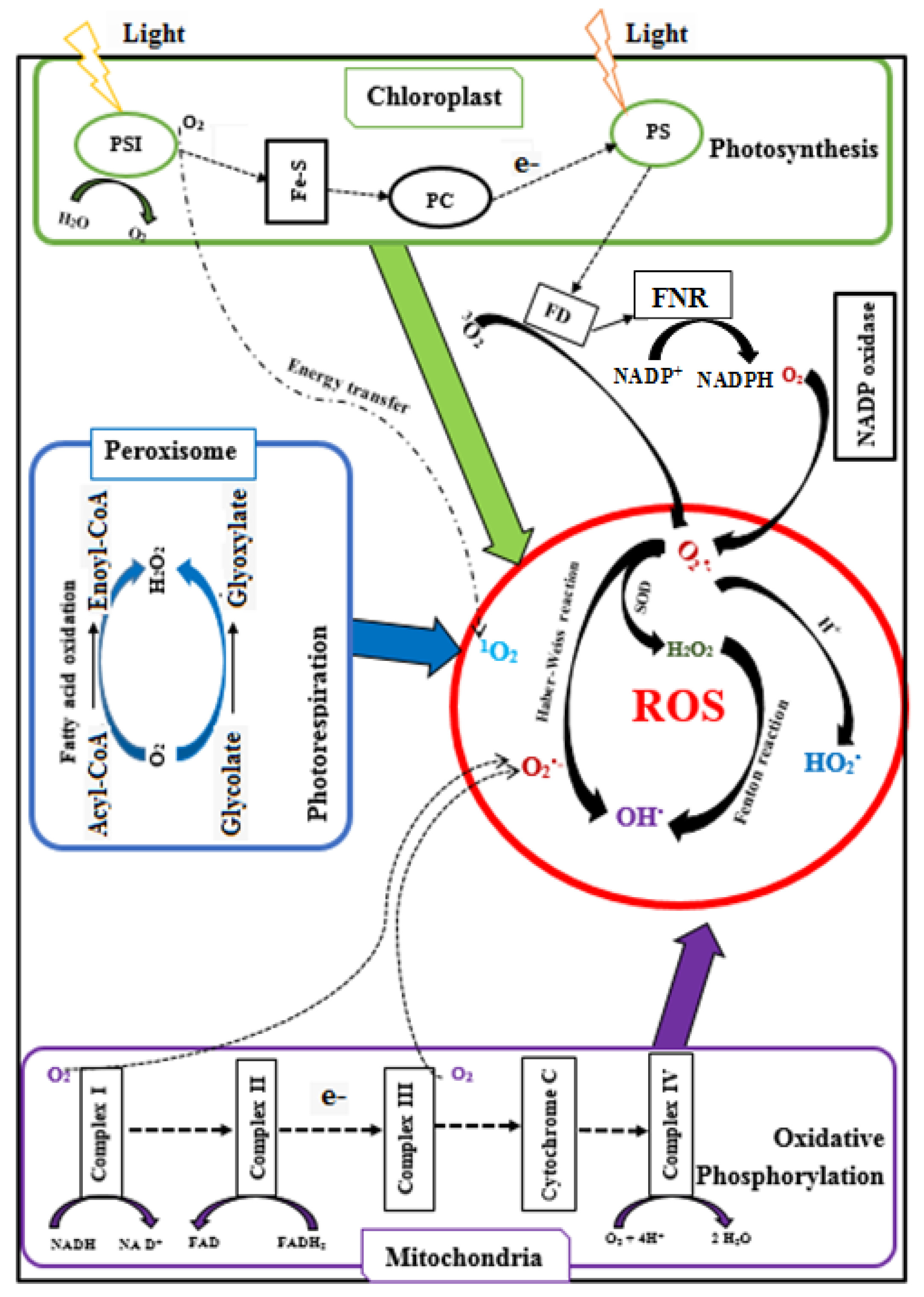{kind=link}
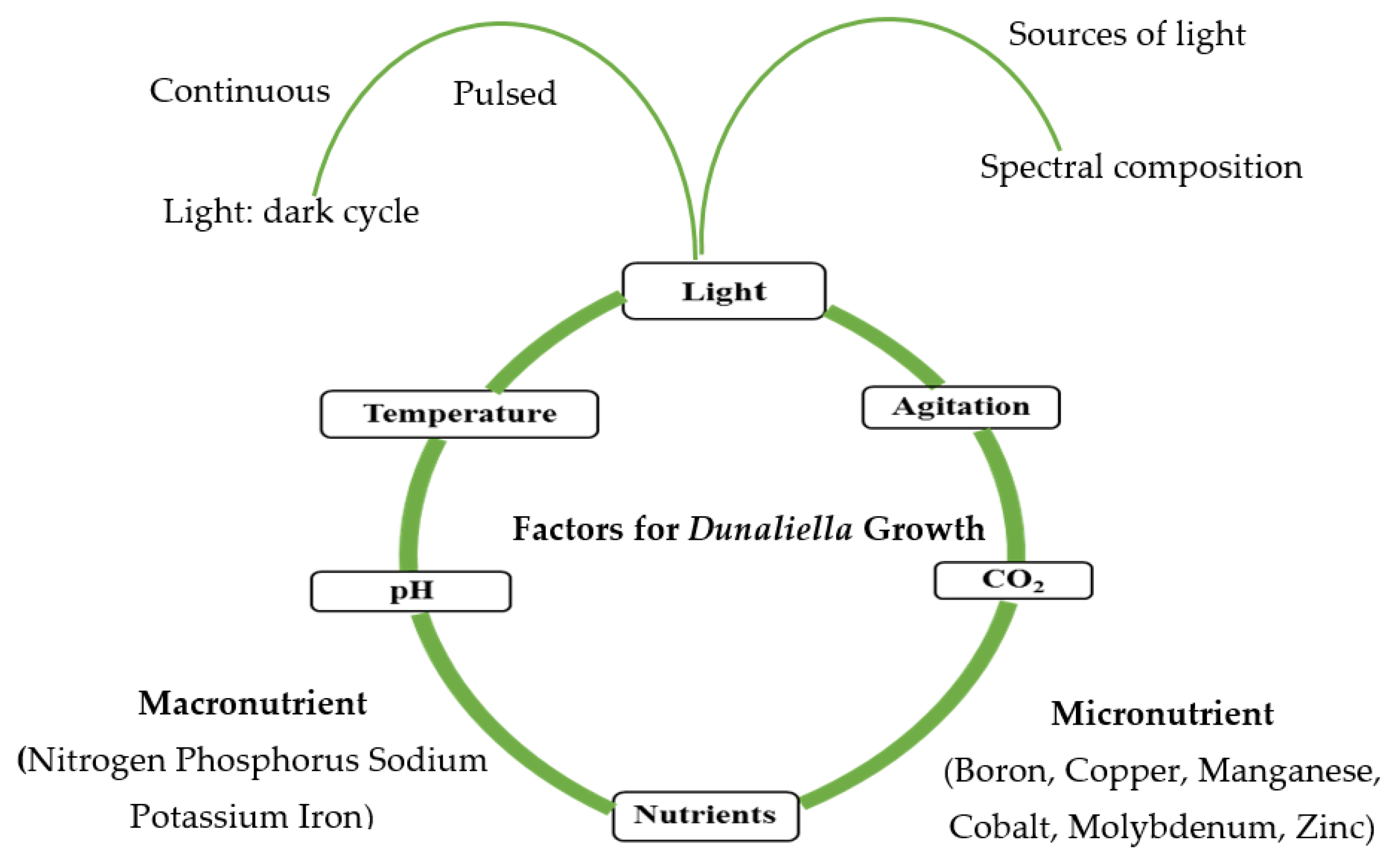{kind=link}
Table 1.
Commercial uses of enzymatic and non-enzymatic antioxidant.
| Antioxidants | Applications | Ref | |
|---|---|---|---|
| Natural antioxidants | SOD | Added to cosmetic products to protect against skin damage | [49,52,60,63,64] |
| Protect against lipid peroxidation, heat, and cold stress in poultry production | |||
| As a therapeutic agent for treatment of inflammatory disorders | |||
| Normal cells protector during radiotherapy for cancer patients | |||
| CAT | Eliminate excessive H2O2 in the textile industry, pulp, and paper industry used for bleaching fibres and pulp, and as a bactericidal disinfectant in food processing and in the pharmaceutical industry | [51,57,65] | |
| In aesthetics (mask treatment) to increase cellular oxygenation in the upper layers of facial epidermis | |||
| Reducing the risk of diabetes mellitus | |||
| GPX | Immune system booster | [42,66] | |
| GST | Protective role against neurogenerative diseases | [66,67] | |
| Decreases the risk of tumours of the head and neck, oral, cavity and colon | |||
| Glutathione | Anti-wrinkle formation, and as a modifier of skin smoothness | [68] | |
| Vitamins | As a food preservative and bread improver, protective activity against heart diseases, reduced the risk of colorectal adenomas and prostate cancer, reduction of thyroid hormone levels | [69,70,71,72,73] | |
| Flavonoids | As cancer preventive agents, protection against type 2 diabetes Functional food additive | [74,75,76,77] | |
| Carotenoid | Anticancer agents, additive to cosmetics and multivitamin preparation Food colouring agent, pro-vitamin A in food and animal feed | [29,38,78] | |
| PUFA | Prevention of heart and inflammatory diseases | [79,80] | |
| Synthetic Antioxidants | BHA | Extending the shelf life of vegetable oil, frying oil, animal feed, cereals, chewing gum, potato flakes and cosmetic products | [81,82,83] |
| BHT | Increasing the shelf life of animal fats, chewing gum, animal feed, vegetable oils | [82,83] | |
| TBHQ | Used as preservative for enhancing storage stability of vegetable oils, margarine, fish oil, fried foods, essential oils, nuts, edible animal fats, butterfat, and packed fried foods | [82] | |
| Propyl gallate | As an antioxidant agent in foods and vegetable oil | [82] | |
| BHA = butylated hydroxyl anisole; TBHQ = tert-butylhydroquinone; BHT = butylated hydroxyl toluene | |||
Table 2.
Production of ROS in cells during biological metabolism and their corresponding neutralising antioxidants.
Table 2.
Production of ROS in cells during biological metabolism and their corresponding neutralising antioxidants.
| ROS | Reaction | Life Span | Function | Sources of ROS | Scavenging Antioxidants |
|---|---|---|---|---|---|
| Singlet (1O2) First excited electronic state of O2 | Chlorophyll triplet state (Chl) is generated sue to insufficient energy dissipation during photosynthesis Chl + 3O2 → 1O2 Reduction of transition metal (Fe3+) O2•− + Fe3+ → 1O2 + Fe2+ | 3 µs (appx.) 4 µs in H2O 100 µs in polar solvents | Gene up-regulation, molecular defense against photo-oxidative stress | Chloroplast | β-carotene, lycopene, tocopherol, ASc, plastoquinone, and proline |
| Superoxide radical (O2•−) One electron reduction of 3O2 | Reduction of oxygen (3O2) during electron transport during the photosynthesis process in chloroplasts or during oxidative phosphorylation in the mitochondria 3O2 + e− → O2•− 3O2 + Xanthine (Xanthine oxidase) → O2•− + uric acid 3O2 + NADPH (NADPH oxidase) → O2•− + NADP+ + H+ | 2–4 µs | Triggering the formation of more ROS which further participates in membrane lipid peroxidation | PSI in Chloroplast oxidative phosphorylation in mitochondria Peroxisomes Plasma membrane | SOD, ASc, glutathione (GSH), flavonoids, Cu |
| H2O2 Two electron reduction of 3O2 or univalent reduction of O2•− | Protonation reaction (acidic conditions) O2•− + H+ + HO2• →H2O2 + 2O2 Glycolate + O2 (Glycolate oxidase) → H2O2 + Glyoxylate Fatty acids (β−oxidation) → Acetyl coenzyme + H2O2 | 1 ms | Act as a signaling molecule (low concentration of H2O2), at high concentration of H2O2 triggers tolerance to various stress, as a regulator of physiological processes (photorespiration and photosynthesis) | Chloroplast, Mitochondria, and Peroxisome | CAT, POD (GPX and APX), peroxiredoxin, ASc, tocopherol, GSH, β-carotene, Se flavonoids, lipoic acid, |
| HO2• | Protonation of superoxide ions O2•− + H+ → HO2• | Attacks PUFA in the negatively charged membrane surface | Mitochondria, microsomes and peroxisomes | ||
| HO• (Three electron reduction of 3O2) | Fenton reaction: H2O2 + Fe2+ → HO• + HO− + Fe3+ Haber-Weiss reactions: O2•− + H2O2 → HO• + HO− + O2 | 1 ps | Attack unsaturated fatty acids in membranes | Mitochondria | ASc, GSH, flavonoids, lipoic acid, proline |
| Nitric oxide (NO•) | L-arginine + O2 Nitric oxide synthase → NO• + Citrulline | Intercellular messenger, the quencher of O2•−, defense against various protozoa, fungi, and mycobacteria | Peroxisome, cytosol | GSH | |
| Peroxynitrite (ONOO−) | NO• + O2•− → ONOO− | React with amino acids residues in enzymes causing inactivation | Peroxiredoxin, Uric acid | ||
| Lipid hydroperoxide | Oxidation of PUFA | Tissue injuries and diseases | Mitochondrial membrane PUFA |
| Characteristics | Photo-Autotrophic | Heterotrophic | Mixotrophic |
|---|---|---|---|
| Carbon assimilation process | Photosynthesis | Aerobic respiration | Photosynthesis and aerobic respiration |
| Energy sources | Light (solar or artificial) | Organic carbon (e.g., glucose, acetic acid glycerol, wastewaters) | Light and organic carbon |
| Carbon sources | Inorganic carbon (CO2, NaHCO3) | Organic carbon | Inorganic and organic |
| Light energy | Mandatory | Not required | Not essential |
| Photo-inhibition effect | High | No effect | Low |
| Growth-limiting factor | Light | Oxygen | Light and oxygen |
| Photo-oxidative damage risk | High | Very low | Protected by accumulating oxygen |
| Types of bioreactors | Photo-bioreactors and open ponds | Fermenters | Photo-bioreactors, open ponds |
| Availability of bioreactor’s vessels | Photo-bioreactor | Commercially available | Bioreactor |
| Bioreactor’s surface tovolume ratio (m2/m3) | High | Low | High |
| Sterility | Usually sanitised | Sterility required | Not required |
| Contamination risk | Low | High | Medium |
| Harvesting cost | High due to the low density of biomass | Low due to high biomass concentration | Low due to high biomass concentration |
| Bioreactor set up cost (per unit of volume) | High | Low | High |
| Bioreactor operation cost (per kg of biomass) | Low | Medium | High |
| Scale-up | Design optimisation required | Easy | Design optimisation required |
| Efficiency in wastewater treatment | Low | High | High |
| Growth rate | Low | Medium | High |
| Biomass density (g L−1) | Low | Medium | High |
| Lipids (%) | Low | High | High |
| Proteins (%) | High | Low | Medium |
| Carbohydrates (%) | Low | High | Low |
| Chlorophyll and β-carotene (%) | High | Low | Medium |
| Lutein (%) | Medium | High | High |
Table 4.
Antioxidant enzymatic response of Dunaliella exposed to different abiotic exogenous stress.
Table 4.
Antioxidant enzymatic response of Dunaliella exposed to different abiotic exogenous stress.
| Stress Conditions | Strains | Optimum Light (µmol Photon m−2 s−1) | Salinity (NaCl) (M) | Temperature (°C) | Antioxidant Enzymatic Response | References | |
|---|---|---|---|---|---|---|---|
| Salinity | 0.05–3.00 M NaCl | D. tertiolecta (UTEX999) | 150 Continuous light | 0.1–0.5 | 26 | No effect on SOD and CAT, GTR activities APX increased (2-fold) at high salinities (0.2–3 M) | [125] |
| 0.5–5.5 M NaCl | D. salina | 150 Light: dark cycle (12:12 h) | 1.5 | (25 ± 2) | CAT activity decreases with salinity SOD increased (0.5–2 M) and then decrease APX decreases (0.5–2 M) and then constant | [122] | |
| 0.05–4.0 M NaCl | D. salina and D. tertiolecta | 150 Light: dark cycle (12:12 h) | 1.25 | (25 ± 2) | CAT and SOD activities increase at hypo saline condition in D. tertiolecta APX activity in D. tertiolecta higher at hypo saline and lower at hyper saline condition GPX activity decreased in D. tertiolecta at higher saline condition (>1.25 M) | [124] | |
| 1–4.0 M NaCl | D. salina | 54 (Continuous) | 2 | 22 | APX increase (171%) at 4 M compared to 2 M | [153] | |
| Light | Outdoor with natural UVR | D. tertiolecta | 250 Light: dark cycle (12:12 h) | - | 20 | SOD activity slightly changed in short and unaltered for long term exposure but no change in APX and GTX activity | [130] |
| UV-B for 6 days | D. salina | 60–80 Light: dark cycle (12:12 h) | - | 20 | CAT activity increased on 3rd day SOD activity increased on 4th day POD activity increased on 3rd day | [128] | |
| UV-B and UV-C for 24 h | D. bardawil and D. salina | 150 UV-A | - | 26 | APX increased by UV -A | [131] | |
| UV-B for 4 h | D. salina | 920 | - | 25 | CAT, SOD and POD activity increased | [34] | |
| Nutrients | Depletion of N, P, S, NP, NS, NPS | D. salina | 60 (Continuous) | - | 30 | SOD and CAT activity increased | [137] |
| KNO3 (0.05, 0.5, 5 mM) | D. salina | 150 Continuous light | - | 25 | CAT, SOD and APX activity decreased with increased nitrogen concentration | [138] | |
| Light and nutrients | Light intensity and depletion of Mn, Zn, Fe, N | D. salina | 70 Light: dark cycle (16:8 h) | 20 | Mn-SOD overexpressed | [36] | |
| UV-B 2 h day−1 for 3 days and deprivation of nitrogen | D. salina | 85 Continuous light | 25±5 | POD and APX activity increased | [37] | ||
| Light and temperature | Low temperature and low light for 24 h | D. salina (Gh-U) | 100 Light: dark (16:8 h) | 28 ± 0.5 | SOD activity increased APX activity increased at low light level GR, MDHAR increased at low temperature | [135] | |
| Low temperature and medium light for 24 h | D. salina (IR-1 and Gh-U) | 100 Light: dark (16:8 h) | 28 ± 0.5 | POD activity increased, GTX increased in strain Gh-U but not in IR-1 at low temperature GR, DHAR activity increased in Gh-U than IR-1 | [35] | ||
| Metals | Cu2+ (1–20 µM) for 24–72 h | D. salina and D. tertiolecta | 54 Continuous light | 22 | APX activity increased | [143] | |
| Hg2+ (0–40 µM) for 48 h and 28 days | D. tertiolecta | 54 (Continuous light) | (22 ± 2) | APX activity increased | [141] | ||
| Cd2+ (0–20 µg L−1) for 24, 48,72, and 96 h | D. viridis | 117 (Light: dark) (12:12 h) | (28 ± 2) | Pigment production decreased | [154] | ||
| Cr6+ (40 ppm) for 24 h | D. salina and D. tertiolecta | 46 White fluorescent light | 25 | SOD activity decreased Chlorophyll ‘a’ content decreased with increase in the exposure time | [155] | ||
| Silver nanoparticles (10, 50, 100 and 200 mg L−1 Ag-NPs) for 8 days | (D. tertiolecta) | 100 Continuous light | 25 | CAT activity increased until 6 days then decreased, SOD activity increased up to 4 days, POD activity increased until 6 days | [140] | ||
| Chemicals | Carbamazepine (CBZ) (0–200 mg L−1) 24, 48,72, and 96 h | D. tertiolecta (CCAP19/6B) | 58 Continuous | (24 ± 1) | Increased carotenoid | [156] | |
| SDBS and CTAC for 48 h | D. bardawil | 144 (Light: dark) (14:10 h) | 26 | CAT and SOD activity increased | [148] | ||
| Trichlorfon and dimehypo | D. salina | 54 (Light: dark) (14:10 h) | 26 | CAT activity increased | [151] | ||
| 2-chlorophenol for 24, 48, and 96 h | D. salina | 100 (Continuous daylight) | (25 ± 1) °C | SOD, CAT, GTX activity increased APX activity decreased | [149] | ||
| Chemical and salinity | (1, 2, 3 M NaCl) and PG (propyl gallate) for 48 h | D. salina (UTEX 200) | 70 Light: dark (16:8 h) | 25 | Maximum CAT activity at 2 M NaCl and no change in SOD activity at 1 and 3 M NaCl Minor increase in APX activity at 3 M NaCl | [152] | |
| Chemical and temperature | 10 °C and 5 °C, Methylene blue and norflurazon | D. salina (IPASS D-294) | 73.6 White fluorescent light | 27 | CAT activity increased Chlorophyll and carotenoid content decreased with temperature | [136] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Roy, U.K.; Nielsen, B.V.; Milledge, J.J. Antioxidant Production in Dunaliella. Appl. Sci. 2021, 11, 3959. https://0-doi-org.brum.beds.ac.uk/10.3390/app11093959
AMA Style
Roy UK, Nielsen BV, Milledge JJ. Antioxidant Production in Dunaliella. Applied Sciences. 2021; 11(9):3959. https://0-doi-org.brum.beds.ac.uk/10.3390/app11093959
Chicago/Turabian StyleRoy, Uttam Kumer, Birthe Vejby Nielsen, and John James Milledge. 2021. "Antioxidant Production in Dunaliella" Applied Sciences 11, no. 9: 3959. https://0-doi-org.brum.beds.ac.uk/10.3390/app11093959
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.