
Cleaning of Phototrophic Biofilms in a Show Cave: The Case of Tesoro Cave, Spain

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
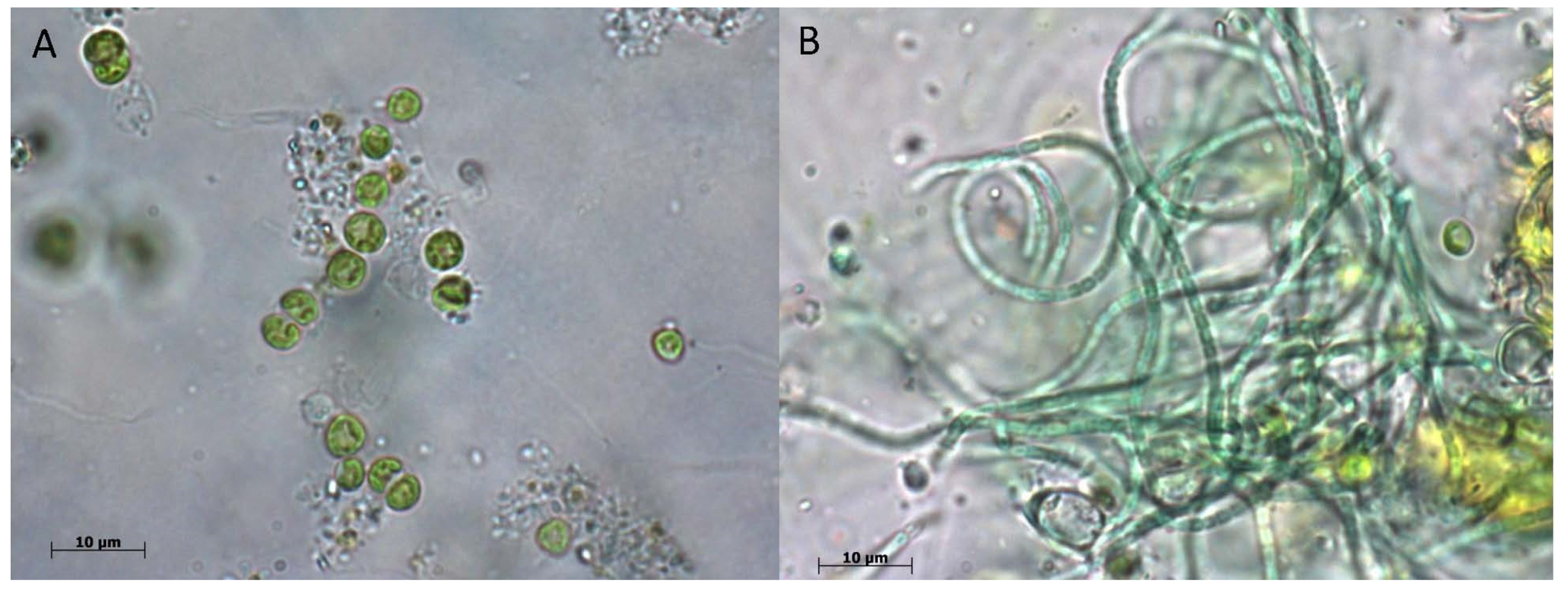
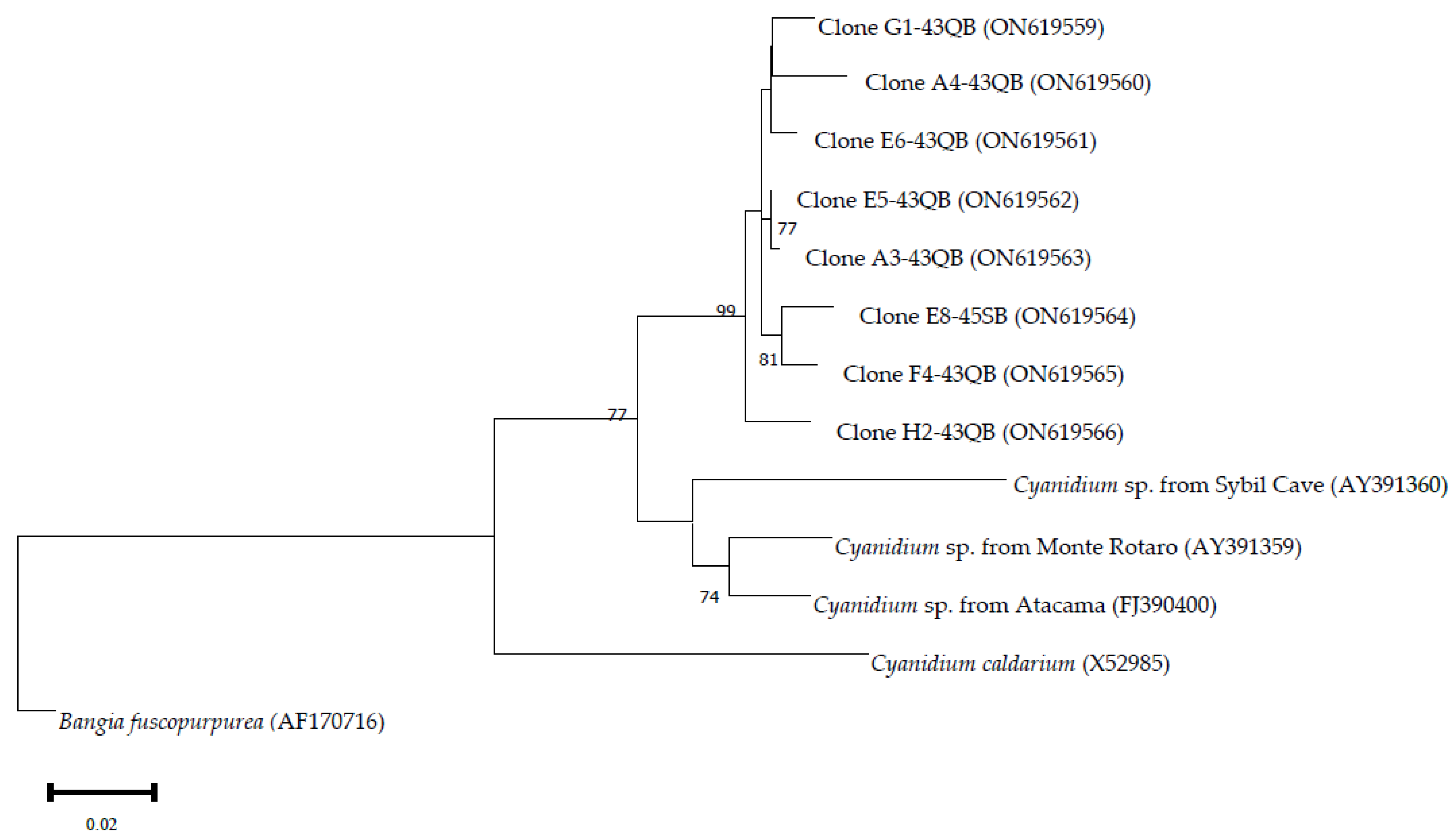
3.1. Composition of Biofilms in Different Cave Sectors
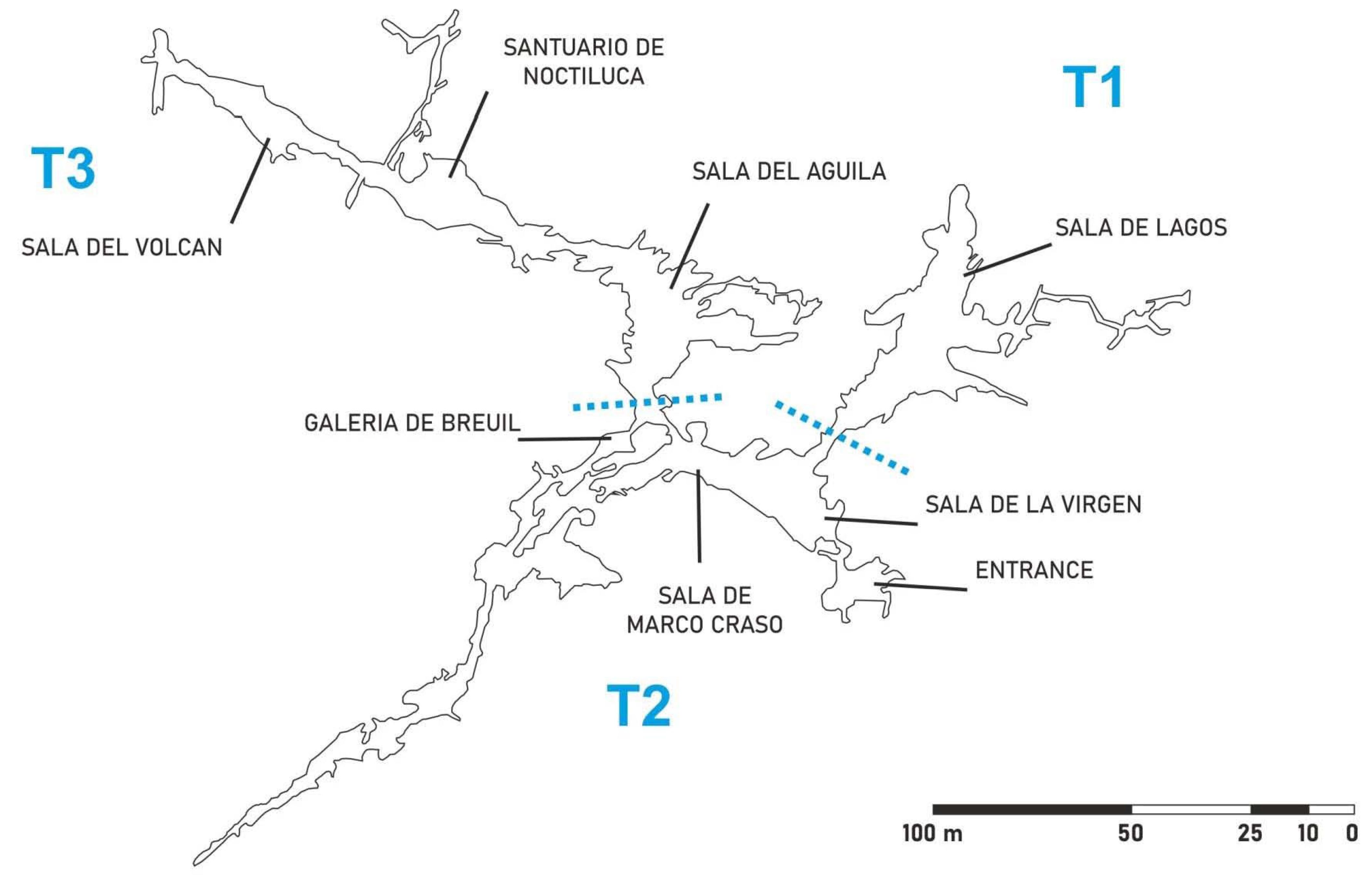
3.1.1. Sector T1
3.1.2. Sector T2
3.1.3. Sector T3
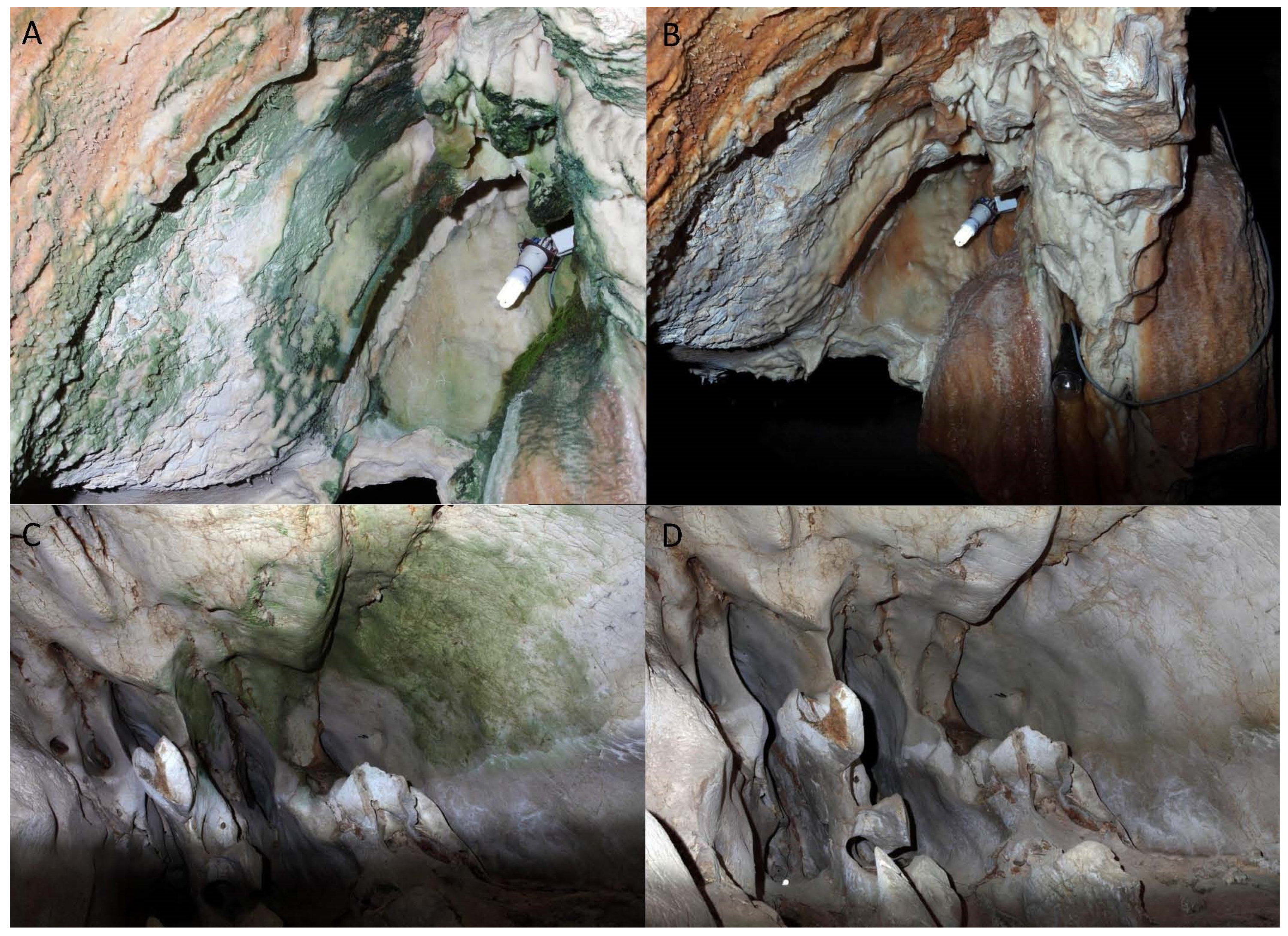
3.2. Preliminary Cleaning Testing
3.3. Recommended Cleaning Protocols
3.4. Effective Cleaning of Biofilms in Tesoro Cave by Agora S.L.
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mulec, J. Lampenflora. In Encyclopedia of Caves; White, W.B., Culver, D.C., Eds.; Academic Press: Waltham, MA, USA, 2012; pp. 451–456. [Google Scholar]
- Trinh, D.A.; Trinh, Q.H.; Tran, N.; Guinea, J.G.; Mattey, D. Eco-friendly remediation of lampenflora on speleothems in tropical karst caves. J. Cave Karst Stud. 2018, 80, 1–12. [Google Scholar] [CrossRef]
- Nikolić, N.; Zarubica, N.; Gavrilović, B.; Predojević, D.; Trbojević, I.; Simić, G.S.; Popović, S. Lampenflora and the entrance biofilm in two show caves: Comparison of microbial community, environmental, and biofilm parameters. J. Cave Karst Stud. 2020, 82, 69–81. [Google Scholar] [CrossRef]
- Lefèvre, M. La maladie verte de Lascaux. Stud. Conserv. 1974, 19, 126–156. [Google Scholar]
- Martin-Sanchez, P.; Miller, A.Z.; Saiz-Jimenez, C. Lascaux Cave: An example of fragile ecological balance in subterranean environments. In Microbial Life of Cave Systems; Engel, A.S., Ed.; DeGruiter: Berlin/Heidelberg, Germany, 2015; pp. 280–301. [Google Scholar]
- Saiz-Jimenez, C.; Cuezva, S.; Jurado, V.; Fernandez-Cortes, A.; Porca, E.; Benavente, D.; Cañaveras, J.C.; Sanchez-Moral, S. Paleolithic art in peril: Policy and science collide at Altamira Cave. Science 2011, 334, 42–43. [Google Scholar] [CrossRef] [Green Version]
- Cuezva, S.; Jurado, V.; Fernandez-Cortes, A.; Garcia-Anton, E.; Rogerio-Candelera, M.A.; Ariño, X.; Benavente, D.; Cañaveras, J.C.; Saiz-Jimenez, C.; Sanchez-Moral, S. Scientific data suggest Altamira Cave should remain closed. In Microbial Life of Cave Systems; Engel, A.S., Ed.; DeGruiter: Berlin/Heidelberg, Germany, 2015; pp. 303–320. [Google Scholar]
- Jurado, V.; Gonzalez-Pimentel, J.L.; Fernandez-Cortes, A.; Martin-Pozas, T.; Ontañon, R.; Palacio, E.; Hermosin, B.; Sanchez-Moral, S.; Saiz-Jimenez, C. Early detection of phototrophic biofilms in the Polychrome Panel, El Castillo Cave, Spain. Appl. Biosci. 2022, 1, 40–63. [Google Scholar] [CrossRef]
- Procházková, K.; Nemcová, Y.; Neustupa, J. A new species Jenufa aeroterrestrica (Chlorophyceae incertae sedis, Viridiplantae), described from Europe. Preslia 2015, 87, 403–416. [Google Scholar]
- Ariño, X.; Hernandez-Marine, M.; Saiz-Jimenez, C. Colonization of Roman tombs by calcifying cyanobacteria. Phycologia 1997, 36, 366–373. [Google Scholar] [CrossRef]
- Saiz-Jimenez, C. Biogeochemistry of weathering processes in monuments. Geomicrobiol. J. 1999, 16, 27–37. [Google Scholar] [CrossRef]
- Albertano, P. Cyanobacterial biofilms in monuments and caves. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 317–343. [Google Scholar]
- Del Rosal Padial, Y.; Jurado Lobo, V.; Hernández-Mariné, M.; Roldán Molina, M.; Sáiz-Jiménez, C. Biofilms en cuevas turísticas: La Cueva de Nerja y la Cueva del Tesoro. In El Karst y el Hombre: Las Cuevas Como Patrimonio Mundial; Andreo, B., Durán, J.J., Eds.; Asociación de Cuevas Turísticas Españolas: Madrid, Spain, 2016; pp. 103–114. [Google Scholar]
- Del Rosal, Y.; Jurado, V.; Roldán, M.; Hernández-Mariné, M.; Saiz-Jiménez, C. Cyanidium sp. colonizadora de cuevas turísticas. In Estudio y Conservación del Patrimonio Cultural. Actas; Moreno Oliva, M., Rogerio Candelera, M.A., López Navarrete, J.T., Hernández Jolín, V., Eds.; Universidad de Málaga: Málaga, Spain, 2015; pp. 170–173. [Google Scholar]
- Jurado, V.; del Rosal, Y.; Gonzalez-Pimentel, J.L.; Hermosin, B.; Saiz-Jimenez, C. Biological control of phototrophic biofilms in a show cave: The case of Nerja Cave. Appl. Sci. 2020, 10, 3448. [Google Scholar] [CrossRef]
- Jurado, V.; Novakova, A.; Hernandez-Marine, M.; Saiz-Jimenez, C. Cueva del Tesoro, Rincon de la Victoria, Malaga: A treasure of biodiversity. In The Conservation of Subterranean Cultural Heritage; Saiz-Jimenez, C., Ed.; CRC Press/Balkema: Leiden, The Netherlands, 2014; pp. 207–214. [Google Scholar]
- Rippka, R.; Deruelles, J.; Waterbury, J.W.; Herdman, M.; Stanier, R.G. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Bischoff, H.W.; Bold, H.C. Phycological Studies IV. Some Soil Algae from Enchanted Rock and Related Algal Species; University of Texas Publication: Austin, TX, USA, 1963; Volume 6318, pp. 1–95. [Google Scholar]
- Ettl, H.; Gärtner, G. Syllabus der Boden, Luft und Flechtenalgen; Fischer, G., Ed.; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota I: Chroococcales. In Süsswasserflora von Mitteleuropa 19/1; Ettl, H., Gärtner, G., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer: Jena, Germany, 1998. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 2: Oscillatoriales. In Süsswasserflora von Mitteleuropa 19/2; Büdel, B., Krienitz, L., Gärtner, G., Schagerl, M., Eds.; Spektrum Akademischer Verlag: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Komárek, J. Cyanoprokaryota 3: Heterocytous Genera. In Süsswasserflora von Mitteleuropa 19/3; Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Spektrum Akademischer Verlag: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Albertano, P.; Ciniglia, C.; Pinto, G.; Pollio, A. The taxonomic position of Cyanidium, Cyanidioschyzon and Galdieria: An update. Hydrobiologia 2000, 433, 137–143. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication. Galway: National University of Ireland. Available online: https://www.algaebase.org (accessed on 1 March 2022).
- Nübel, U.; Garcia-Pichel, F.; Muyzer, G. PCR primers to amplify 16S rRNA genes from cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 3327–3332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Gross, W.; Heilmann, I.; Lenze, D.; Schnarrenberger, C. Biogeography of the Cyanidiaceae (Rhodophyta) based on 18S ribosomal RNA sequence data. Eur. J. Phycol. 2001, 36, 275–280. [Google Scholar] [CrossRef]
- Walker, J.J.; Spear, J.R.; Pace, N.R. Geobiology of a microbial endolithic community in the Yellowstone geothermal environment. Nature 2005, 434, 1011–1014. [Google Scholar] [CrossRef]
- Iovinella, N.; Eren, A.; Pinto, G.; Pollio, A.; Davis, S.J.; Cennamo, P.; Ciniglia, C. Cryptic dispersal of Cyanidiophytina (Rhodophyta) in non-acidic environments from Turkey. Extremophiles 2018, 22, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Ciniglia, C.; Pinto, G.; Pollio, A. Cyanidium from caves: A reinstatement of Cyanidium chilense Schwabe (Cyanidiophytina, Rhodophyta). Phytotaxa 2017, 295, 86–88. [Google Scholar] [CrossRef]
- Ciniglia, C.; Yoon, H.S.; Pollio, A.; Pinto, G.; Bhattacharya, D. Hidden biodiversity of the extremophilic Cyanidiales red algae. Mol. Ecol. 2004, 13, 1827–1838. [Google Scholar] [CrossRef] [PubMed]
- Ciniglia, C.; Cennamo, P.; De Natale, A.; De Stefano, M.; Sirakov, M.; Iovinella, M.; Yoon, H.S.; Pollio, A. Cyanidium chilense (Cyanidiophyceae, Rhodophyta) from tuff rocks of the archeological site of Cuma, Italy. Phycol. Res. 2019, 67, 311–319. [Google Scholar] [CrossRef]
- Azúa-Bustos, A.; González-Silva, C.; Mancilla, R.A.; Salas, L.; Palma, R.E.; Wynne, J.J.; McKay, C.P.; Vicuña, R. Ancient photosynthetic eukaryote biofilms in an Atacama Desert coastal cave. Microb. Ecol. 2009, 58, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Mulec, J.; Kubesová, S. Diversity of bryophytes in show caves in Slovenia and relation to light intensities. Acta Carsologica 2010, 39, 587–596. [Google Scholar] [CrossRef]
- Glime, J.M. Caves—Overall bryophyte flora. In Bryophyte Ecology. Habitat and Role; Glime, J.M., Ed.; Michigan Technological University: Houghton, MI, USA, 2021; Chapter 18-2; Volume 4, pp. 1–30. [Google Scholar]
- Whitehouse, H.L.K. The production of protonemal gemmae by mosses growing in deep shade. J. Bryol. 1980, 11, 133–138. [Google Scholar] [CrossRef]
- Saiz-Jimenez, C.; Hermosin, B. Thermally assisted hydrolysis and methylation of the black deposit coating the ceiling and walls of Cueva del Encajero, Quesada, Spain. J. Anal. Appl. Pyrol. 1999, 49, 349–357. [Google Scholar] [CrossRef]
- Kol, E. Algal growth experiments in the Baradla cave at Aggletek. Int. J. Speleol. 1967, 2, 457–474. [Google Scholar] [CrossRef] [Green Version]
- Mulec, J.; Kosi, G.G.; Vrhovsek, D. Characterization of cave aerophytic algal communities and effects of irradiance levels on production of pigments. J. Cave Karst Stud. 2008, 70, 3–12. [Google Scholar]
- Czerwik-Marcinkowska, J.; Mrozińska, T. Algae and cyanobacteria in caves of the Polish Jura. Pol. Bot. J. 2011, 56, 203–243. [Google Scholar]
- Cennamo, P.; Marzano, C.; Ciniglia, C.; Pinto, G.; Cappelletti, P.; Caputo, P.; Pollio, A. A survey of the algal flora of anthropogenic caves of Campi Flegrei (Naples, Italy) archeological district. J. Cave Karst Stud. 2012, 74, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Czerwik-Marcinkowska, J. Observations on aerophytic cyanobacteria and algae from ten caves in the Ojców National Park. Acta Agrobot. 2013, 66, 39–52. [Google Scholar] [CrossRef]
- Czerwik-Marcinkowska, J.; Wojciechowska, A.; Massalski, A. Biodiversity of limestone caves: Aggregations of aerophytic algae and cyanobacteria in relation to site factors. Pol. J. Ecol. 2015, 63, 481–499. [Google Scholar] [CrossRef]
- Cennamo, P.; Montuori, N.; Trojsi, G.; Fatigati, G.; Moretti, A. Biofilms in churches built in grottoes. Sci. Total Environ. 2016, 543, 727–738. [Google Scholar] [CrossRef]
- Popkova, A.; Mazina, S. Microbiota of hypogean habitats in Otap Head Cave. J. Environ. Res. Eng. Manag. 2019, 75, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Popović, S.; Nikolić, N.; Jovanović, J.; Predojević, D.; Trbojević, I.; Manić, L.; Simić, G.S. Cyanobacterial and algal abundance and biomass in cave biofilms and relation to environmental and biofilm parameters. Int. J. Speleol. 2019, 48, 49–61. [Google Scholar] [CrossRef]
- Mulec, J.; Kosi, G. Lampenflora algae and methods of growth control. J. Cave Karst Stud. 2009, 71, 109–115. [Google Scholar]
- Faimon, J.; Stelcl, J.; Kubesová, S.; Zimák, J. Environmentally acceptable effect of hydrogen peroxide on cave “lamp-flora”, calcite speleothems and limestones. Environ. Pollut. 2003, 122, 417–422. [Google Scholar] [CrossRef]
- Esteban Pérez, E. Study and remediation of environmental problems caused due to the growth of algae in speleothems of calcareous caves adapted for tourism—A case of success in Spain. J. Environ. Geol. 2018, 2, 20–28. [Google Scholar]
- Baquedano Estévez, C.; Moreno Merino, L.; de la Losa Román, A.; Durán Valsero, J.J. The lampenflora in show caves and its treatment: An emerging ecological problem. Int. J. Speleol. 2019, 48, 249–277. [Google Scholar] [CrossRef] [Green Version]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurado, V.; Hernandez-Marine, M.; Rogerio-Candelera, M.A.; Ruano, F.; Aguilar, C.; Aguilar, J.; Saiz-Jimenez, C. Cleaning of Phototrophic Biofilms in a Show Cave: The Case of Tesoro Cave, Spain. Appl. Sci. 2022, 12, 7357. https://0-doi-org.brum.beds.ac.uk/10.3390/app12157357
Jurado V, Hernandez-Marine M, Rogerio-Candelera MA, Ruano F, Aguilar C, Aguilar J, Saiz-Jimenez C. Cleaning of Phototrophic Biofilms in a Show Cave: The Case of Tesoro Cave, Spain. Applied Sciences. 2022; 12(15):7357. https://0-doi-org.brum.beds.ac.uk/10.3390/app12157357
Chicago/Turabian StyleJurado, Valme, Mariona Hernandez-Marine, Miguel Angel Rogerio-Candelera, Francisco Ruano, Clara Aguilar, Juan Aguilar, and Cesareo Saiz-Jimenez. 2022. "Cleaning of Phototrophic Biofilms in a Show Cave: The Case of Tesoro Cave, Spain" Applied Sciences 12, no. 15: 7357. https://0-doi-org.brum.beds.ac.uk/10.3390/app12157357