
Effects of Oxidized Pyrenes on the Biological Responses in the Human Bronchial Epithelial Cells
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture
2.3. Treatment with Pyrene and Its Quinone Forms
2.4. Cell Viability Assay
2.5. Detection of Cell Death via Apoptosis and Necrosis
2.6. Quantitation of Inflammatory Proteins in the Culture Supernatant
2.7. Quantification of ROS Generation
2.8. Statistical Analysis
3. Results
3.1. Effect of Pyrene and Its Quinone Forms on the Cellular Viability of Bronchial Epithelial Cells
3.2. Effects of Pyrene and Its Quinone Forms on Airway Epithelial Cell Apoptosis and Necrosis
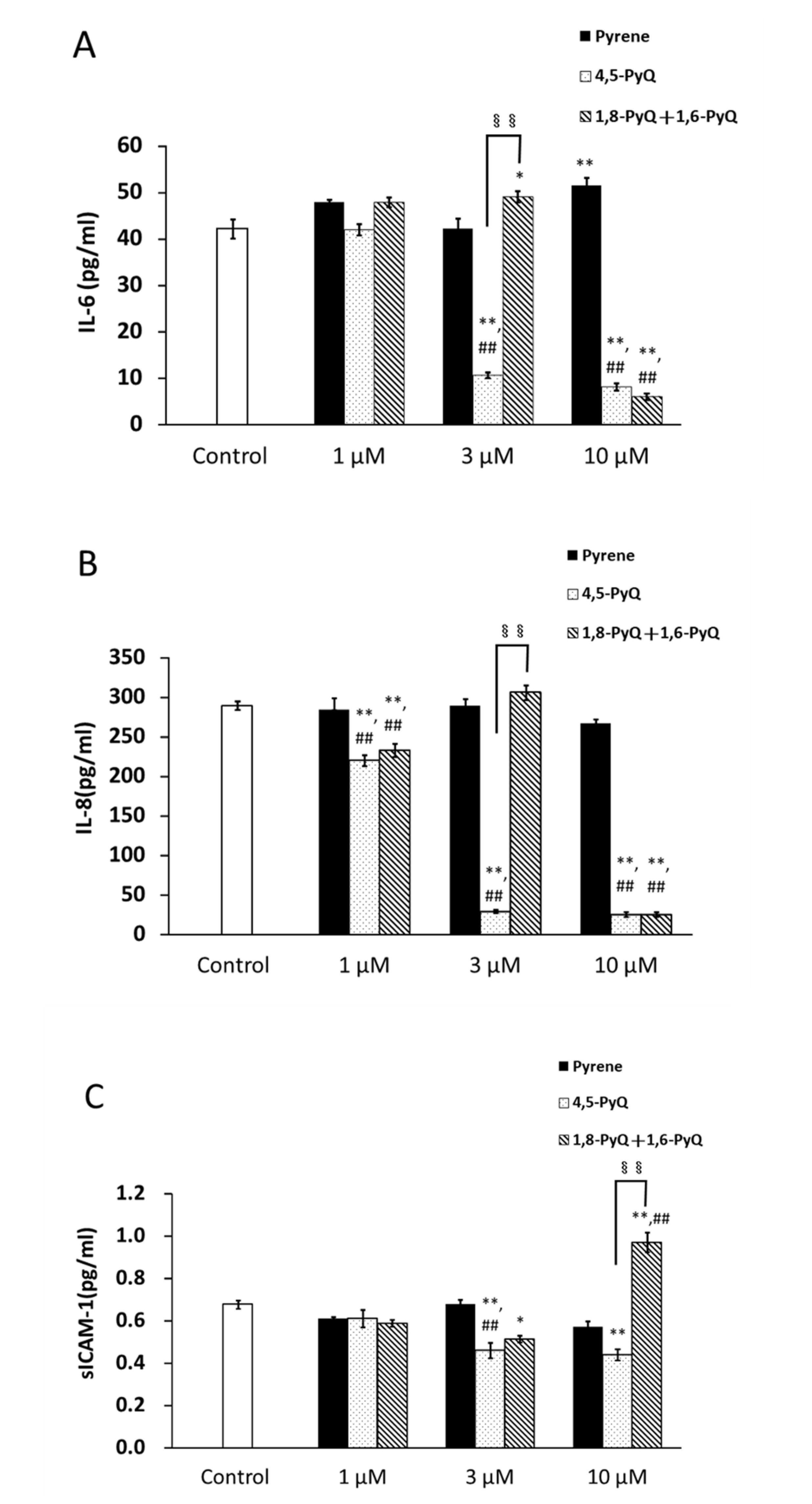
3.3. Effect of Pyrene and Its Quinone Forms on IL-6, IL-8, and sICAM-1 Production by Bronchial Epithelial Cells
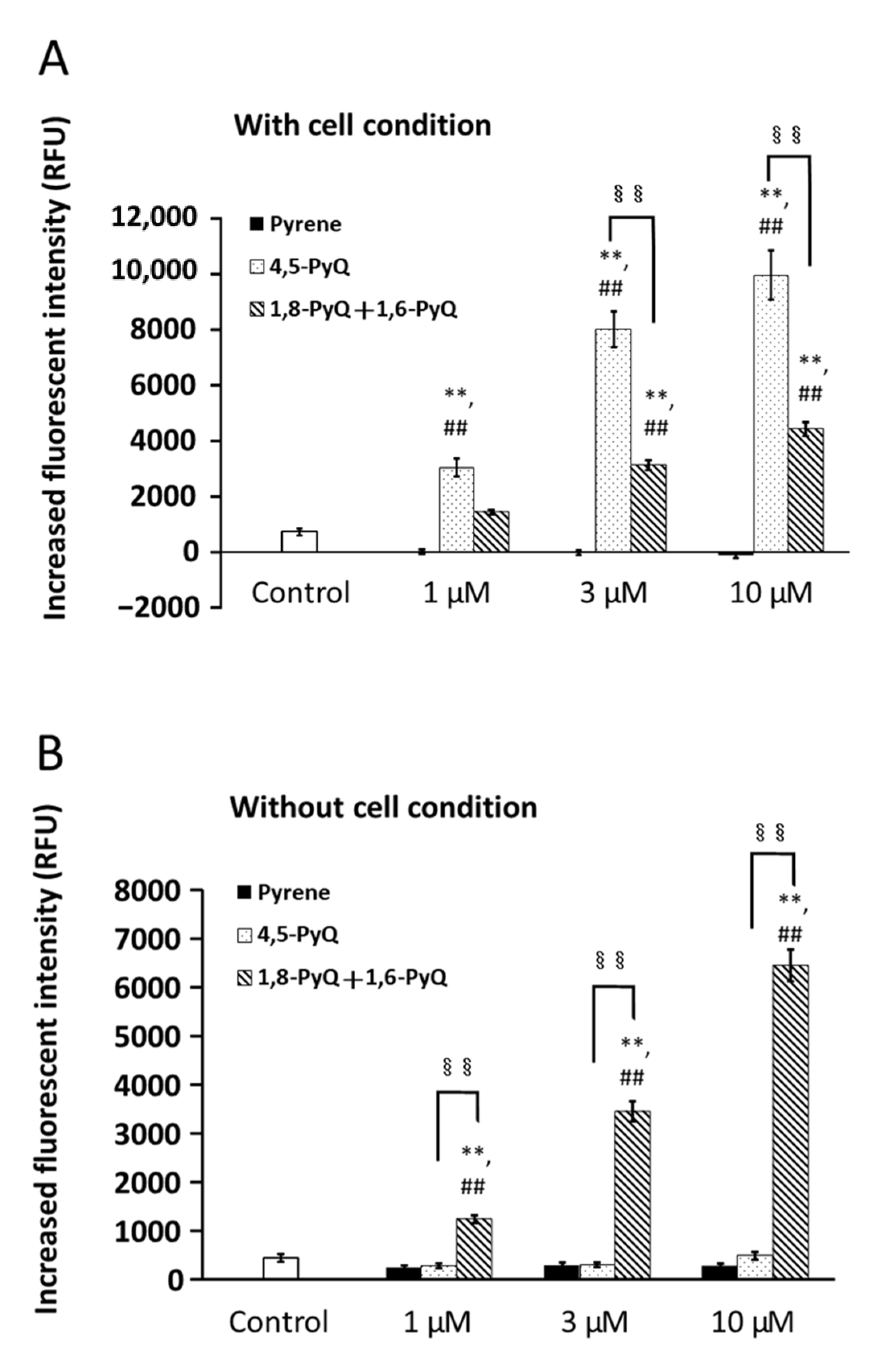
3.4. Effects of Pyrene and Its Quinone Forms on ROS Production by Bronchial Epithelial Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zanobetti, A.; Franklin, M.; Koutrakis, P.; Schwartz, J. Fine particulate air pollution and its components in association with cause-specific emergency admissions. Environ. Health 2009, 8, 58. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Chen, R.; Yang, C.; Lin, Z.; Cai, J.; Xia, Y.; Wang, C.; Li, H.; Johnson, N.; Xu, X.; et al. Association between fine particulate matter chemical constituents and airway inflammation: A panel study among healthy adults in China. Environ. Res. 2016, 150, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Schwarze, P.E.; Ovrevik, J.; Låg, M.; Refsnes, M.; Nafstad, P.; Hetland, R.B.; Dybing, E. Particulate matter properties and health effects: Consistency of epidemiological and toxicological studies. Hum. Exp. Toxicol. 2006, 25, 559–579. [Google Scholar] [CrossRef] [PubMed]
- Dockery, D.W.; Pope, C.A., 3rd; Xu, X.; Spengler, J.D.; Ware, J.H.; Fay, M.E.; Ferris, B.G., Jr.; Speizer, F.E. An association between air pollution and mortality in six U. S. cities. N. Engl. J. Med. 1993, 329, 1753–1759. [Google Scholar] [CrossRef] [PubMed]
- Piao, C.H.; Fan, Y.; Nguyen, T.V.; Shin, H.S.; Kim, H.T.; Song, C.H.; Chai, O.H. PM(2.5) Exacerbates Oxidative Stress and Inflammatory Response through the Nrf2/NF-κB Signaling Pathway in OVA-Induced Allergic Rhinitis Mouse Model. Int. J. Mol. Sci. 2021, 22, 8173. [Google Scholar] [CrossRef] [PubMed]
- Dishaw, L.; Yost, E.; Arzuaga, X.; Luke, A.; Kraft, A.; Walker, T.; Thayer, K. A novel study evaluation strategy in the systematic review of animal toxicology studies for human health assessments of environmental chemicals. Environ. Int. 2020, 141, 105736. [Google Scholar] [CrossRef]
- Pan, S.; Qiu, Y.; Li, M.; Yang, Z.; Liang, D. Recent Developments in the Determination of PM(2.5) Chemical Composition. Bull. Environ. Contam. Toxicol. 2022, 108, 819–823. [Google Scholar] [CrossRef]
- Kameda, T.; Azumi, E.; Fukushima, A.; Tang, N.; Matsuki, A.; Kamiya, Y.; Toriba, A.; Hayakawa, K. Mineral dust aerosols promote the formation of toxic nitropolycyclic aromatic compounds. Sci. Rep. 2016, 6, 24427. [Google Scholar] [CrossRef]
- Liao, K.; Yu, J.Z. Abundance and sources of benzo[a]pyrene and other PAHs in ambient air in Hong Kong: A review of 20-year measurements (1997–2016). Chemosphere 2020, 259, 127518. [Google Scholar] [CrossRef]
- Hiyoshi, K.; Takano, H.; Inoue, K.; Ichinose, T.; Yanagisawa, R.; Tomura, S.; Cho, A.K.; Froines, J.R.; Kumagai, Y. Effects of a single intratracheal administration of phenanthraquinone on murine lung. J. Appl. Toxicol. 2005, 25, 47–51. [Google Scholar] [CrossRef]
- Hiyoshi, K.; Takano, H.; Inoue, K.I.; Ichinose, T.; Yanagisawa, R.; Tomura, S.; Kumagai, Y. Effects of phenanthraquinone on allergic airway inflammation in mice. Clin. Exp. Allergy 2005, 35, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Takano, H.; Ichinose, T.; Tomura, S.; Yanagisawa, R.; Sakurai, M.; Sumi, D.; Cho, A.K.; Hiyoshi, K.; Kumagai, Y. Effects of naphthoquinone on airway responsiveness in the presence or absence of antigen in mice. Arch. Toxicol. 2007, 81, 575–581. [Google Scholar] [CrossRef]
- Inoue, K.; Takano, H.; Hiyoshi, K.; Ichinose, T.; Sadakane, K.; Yanagisawa, R.; Tomura, S.; Kumagai, Y. Naphthoquinone enhances antigen-related airway inflammation in mice. Eur. Respir. J. 2007, 29, 259–267. [Google Scholar] [CrossRef]
- He, M.; Ichinose, T.; Yoshida, S.; Ito, T.; He, C.; Yoshida, Y.; Arashidani, K.; Takano, H.; Sun, G.; Shibamoto, T. PM2.5-induced lung inflammation in mice: Differences of inflammatory response in macrophages and type II alveolar cells. J. Appl. Toxicol. 2017, 37, 1203–1218. [Google Scholar] [CrossRef] [PubMed]
- Laskin, D.L.; Mainelis, G.; Turpin, B.J.; Patel, K.J.; Sunil, V.R. Pulmonary effects of inhaled diesel exhaust in young and old mice: A pilot project. Res. Rep. Health Eff. Inst. 2010, 151, 3–31. [Google Scholar]
- Sun, B.; Shi, Y.; Li, Y.; Jiang, J.; Liang, S.; Duan, J.; Sun, Z. Short-term PM(2.5) exposure induces sustained pulmonary fibrosis development during post-exposure period in rats. J. Hazard. Mater. 2020, 385, 121566. [Google Scholar] [CrossRef]
- Ichinose, T.; Furuyama, A.; Sagai, M. Biological effects of diesel exhaust particles (DEP). II. Acute toxicity of DEP introduced into lung by intratracheal instillation. Toxicology 1995, 99, 153–167. [Google Scholar] [CrossRef]
- Ichinose, T.; Yamanushi, T.; Seto, H.; Sagai, M. Oxygen radicals in lung carcinogenesis accompanying phagocytosis of diesel exhaust particles. Int. J. Oncol. 1997, 11, 571–575. [Google Scholar] [CrossRef]
- Sagai, M.; Furuyama, A.; Ichinose, T. Biological effects of diesel exhaust particles (DEP). III. Pathogenesis of asthma like symptoms in mice. Free Radic. Biol. Med. 1996, 21, 199–209. [Google Scholar] [CrossRef]
- Takano, H.; Yoshikawa, T.; Ichinose, T.; Miyabara, Y.; Imaoka, K.; Sagai, M. Diesel exhaust particles enhance antigen-induced airway inflammation and local cytokine expression in mice. Am. J. Respir. Crit. Care Med. 1997, 156, 36–42. [Google Scholar] [CrossRef]
- Takano, H.; Ichinose, T.; Miyabara, Y.; Shibuya, T.; Lim, H.B.; Yoshikawa, T.; Sagai, M. Inhalation of diesel exhaust enhances allergen-related eosinophil recruitment and airway hyperresponsiveness in mice. Toxicol. Appl. Pharmacol. 1998, 150, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Takano, H.; Yanagisawa, R.; Ichinose, T.; Sadakane, K.; Yoshino, S.; Yoshikawa, T.; Morita, M. Diesel exhaust particles enhance lung injury related to bacterial endotoxin through expression of proinflammatory cytokines, chemokines, and intercellular adhesion molecule-1. Am. J. Respir. Crit. Care Med. 2002, 165, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.M.; Kim, H.R.; Park, Y.J.; Lee, S.Y.; Chung, K.H. Organic extracts of urban air pollution particulate matter (PM2.5)-induced genotoxicity and oxidative stress in human lung bronchial epithelial cells (BEAS-2B cells). Mutat. Res. 2011, 723, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Dergham, M.; Lepers, C.; Verdin, A.; Billet, S.; Cazier, F.; Courcot, D.; Shirali, P.; Garçon, G. Prooxidant and proinflammatory potency of air pollution particulate matter (PM2.5–0.3) produced in rural, urban, or industrial surroundings in human bronchial epithelial cells (BEAS-2B). Chem. Res. Toxicol. 2012, 25, 904–919. [Google Scholar] [CrossRef] [PubMed]
- Fraser, M.P.; Buzcu, B.; Yue, Z.W.; McGaughey, G.R.; Desai, N.R.; Allen, D.T.; Seila, R.L.; Lonneman, W.A.; Harley, R.A. Separation of fine particulate matter emitted from gasoline and diesel vehicles using chemical mass balancing techniques. Environ. Sci. Technol. 2003, 37, 3904–3909. [Google Scholar] [CrossRef] [PubMed]
- Cho, A.K.; di Stefano, E.; You, Y.; Rodriguez, C.E.; Schmitz, D.A.; Kumagai, Y.; Miguel, A.H.; Eiguren-Fernandez, A.; Kobayashi, T.; Avol, E.; et al. Determination of four quinones in diesel exhaust particles, SRM 1649a, and atmospheric PM2.5 special issue of aerosol science and technology on findings from the fine particulate matter supersites program. Aerosol Sci. Technol. 2004, 38, 68–81. [Google Scholar] [CrossRef]
- Schuetzle, D.; Lee, F.S.; Prater, T.J. The identification of polynuclear aromatic hydrocarbon (PAH) derivatives in mutagenic fractions of diesel particulate extracts. Int. J. Environ. Anal. Chem. 1981, 9, 93–144. [Google Scholar] [CrossRef]
- Fine, P.M.; Cass, G.R.; Simoneit, B.R. Chemical characterization of fine particle emissions from fireplace combustion of woods grown in the northeastern United States. Environ. Sci.Technol. 2001, 35, 2665–2675. [Google Scholar] [CrossRef]
- Monks, T.J.; Lau, S.S. Toxicology of quinone-thioethers. Crit. Rev. Toxicol. 1992, 22, 243–270. [Google Scholar] [CrossRef]
- Henry, T.R.; Wallace, K.B. Differential mechanisms of cell killing by redox cycling and arylating quinones. Arch. Toxicol. 1996, 70, 482–489. [Google Scholar] [CrossRef]
- Bolton, J.L.; Trush, M.A.; Penning, T.M.; Dryhurst, G.; Monks, T.J. Role of quinones in toxicology. Chem. Res. Toxicol. 2000, 13, 135–160. [Google Scholar] [CrossRef] [PubMed]
- Schuetzle, D. Sampling of vehicle emissions for chemical analysis and biological testing. Environ. Health Perspect. 1983, 47, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Traub, S.; Nikonova, A.; Carruthers, A.; Dunmore, R.; Vousden, K.A.; Gogsadze, L.; Hao, W.; Zhu, Q.; Bernard, K.; Zhu, J.; et al. An anti-human ICAM-1 antibody inhibits rhinovirus-induced exacerbations of lung inflammation. PLoS Pathog. 2013, 9, e1003520. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Paik, S.H.; Lim, C.M.; Lee, S.D.; Koh, Y.; Kim, W.S.; Kim, W.D. Value of ICAM-1 expression and soluble ICAM-1 level as a marker of activity in sarcoidosis. Chest 1999, 115, 1059–1065. [Google Scholar] [CrossRef]
- Marguet, C.; Dean, T.P.; Warner, J.O. Soluble intercellular adhesion molecule-1 (sICAM-1) and interferon-gamma in bronchoalveolar lavage fluid from children with airway diseases. Am. J. Respir. Crit. Care Med. 2000, 162, 1016–1022. [Google Scholar] [CrossRef]
- Takizawa, H.; Abe, S.; Ohtoshi, T.; Kawasaki, S.; Takami, K.; Desaki, M.; Sugawara, I.; Hashimoto, S.; Azuma, A.; Nakahara, K.; et al. Diesel exhaust particles up-regulate expression of intercellular adhesion molecule-1 (ICAM-1) in human bronchial epithelial cells. Clin. Exp. Immunol. 2000, 120, 356–362. [Google Scholar] [CrossRef]
- Rui, W.; Guan, L.; Zhang, F.; Zhang, W.; Ding, W. PM2.5-induced oxidative stress increases adhesion molecules expression in human endothelial cells through the ERK/AKT/NF-κB-dependent pathway. J. Appl. Toxicol. 2016, 36, 48–59. [Google Scholar] [CrossRef]
- Nasreen, N.; Khodayari, N.; Sriram, P.S.; Patel, J.; Mohammed, K.A. Tobacco smoke induces epithelial barrier dysfunction via receptor EphA2 signaling. Am. J. Physiol. Cell Physiol. 2014, 306, C1154–C1166. [Google Scholar] [CrossRef]
- Chen, W.Y.; Wang, M.; Zhang, J.; Barve, S.S.; McClain, C.J.; Joshi-Barve, S. Acrolein Disrupts Tight Junction Proteins and Causes Endoplasmic Reticulum Stress-Mediated Epithelial Cell Death Leading to Intestinal Barrier Dysfunction and Permeability. Am. J. Pathol. 2017, 187, 2686–2697. [Google Scholar] [CrossRef]
- Fischer, A.; Gluth, M.; Pape, U.F.; Wiedenmann, B.; Theuring, F.; Baumgart, D.C. Adalimumab prevents barrier dysfunction and antagonizes distinct effects of TNF-α on tight junction proteins and signaling pathways in intestinal epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G970–G979. [Google Scholar] [CrossRef]
- Long, J.; Manchandia, T.; Ban, K.; Gao, S.; Miller, C.; Chandra, J. Adaphostin cytoxicity in glioblastoma cells is ROS-dependent and is accompanied by upregulation of heme oxygenase-1. Cancer Chemother. Pharmacol. 2007, 59, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhong, Y.; Liao, J.; Wang, G. PM2.5-related cell death patterns. Int. J. Med. Sci. 2021, 18, 1024–1029. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Honda, A.; Inoue, K.-i.; Takai, S.; Kameda, T.; Ueda, K.; Takano, H. Effects of Oxidized Pyrenes on the Biological Responses in the Human Bronchial Epithelial Cells. Appl. Sci. 2022, 12, 9664. https://0-doi-org.brum.beds.ac.uk/10.3390/app12199664
Honda A, Inoue K-i, Takai S, Kameda T, Ueda K, Takano H. Effects of Oxidized Pyrenes on the Biological Responses in the Human Bronchial Epithelial Cells. Applied Sciences. 2022; 12(19):9664. https://0-doi-org.brum.beds.ac.uk/10.3390/app12199664
Chicago/Turabian StyleHonda, Akiko, Ken-ichiro Inoue, Satsuki Takai, Takayuki Kameda, Kayo Ueda, and Hirohisa Takano. 2022. "Effects of Oxidized Pyrenes on the Biological Responses in the Human Bronchial Epithelial Cells" Applied Sciences 12, no. 19: 9664. https://0-doi-org.brum.beds.ac.uk/10.3390/app12199664