
Effect of Soil Water Contents on Arsenic Accumulation in Phytoliths of Pteris multifida and Phragmites australis

1
OJEong Resilience Institute, Korea University, Seoul 02841, Republic of Korea
2
Division of Environmental Science and Ecological Engineering, College of Life Science and Biotechnology, Korea University, Seoul 02841, Republic of Korea
*
Author to whom correspondence should be addressed.
Appl. Sci. 2022, 12(24), 12518; https://0-doi-org.brum.beds.ac.uk/10.3390/app122412518
Submission received: 2 November 2022
/
Revised: 24 November 2022
/
Accepted: 5 December 2022
/
Published: 7 December 2022
(This article belongs to the Special Issue Potentially Toxic Trace Elements in Contaminated Sites: Fate, Risk and Remediation)
Abstract
:The accumulation of metal(loid)s in phytoliths can physically immobilize the toxic materials and reduce toxicity in plant bodies and soil. Spider brake (Pteris multifida) is a well-known As hyperaccumulator that also harbors phytolith. However, As accumulation in phytoliths has not yet been studied with P. multifida. Soil water content is considered the main factor influencing phytolith accumulation. In this study, the As concentration in the phytoliths of P. multifida was compared with that in common reed (Phragmites australis) phytoliths with various soil water content. The range of As concentration in the phytoliths of P. multifida was 414.70–1610.74 mg kg−1, and the range for P. australis phytoliths was 41.67–126.54 mg kg−1. In P. multifida, higher soil water content increased As accumulation in the phytolith but did not affect the phytolith content in the plant. In P. australis, the higher soil water content increased phytolith content in the plant but decreased As concentration in phytoliths. This study suggests that P. multifida has higher As content in phytoliths than P. australis, and this accumulation can be affected by soil water content. The current findings provide how As encapsulated in phytolith and detoxified in different plants and conditions.
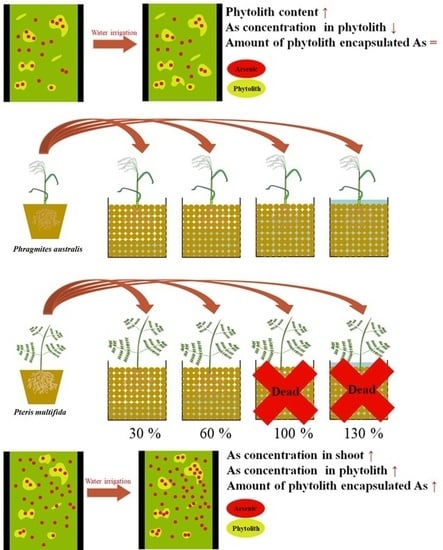
1. Introduction
Silicon (Si) is a non-essential yet helpful element in most higher plants [1]. Si enhances the resistance of plants to various biological and non-biological stresses [2]. A critical role of Si in plants is to reduce the translocation of As from roots to the shoot and to stimulate the antioxidant system [3].
Silicon deposits within cell walls of the plant endodermis and epidermis, forming amorphous opals called phytoliths [4]. Phytoliths consist mainly of Si and contain many elements and organic matters that were part of the plant, have strong resistance against weathering, and are considered to act as carbon sinks [5]. The encapsulation of metal(loid)s within phytoliths represents a new field of research, and there are two suggested roles of metal(loid)s encapsulation in phytoliths. The first is the sequestration of toxic materials from plant metabolism and the detoxification of the materials [6]. The deposited silica in the plant body also helps the dependent mechanism of the cell wall to block toxic material from the plant cell [7].
The second is the encapsulation of toxic material in soil after plant organic matter degrades. There are two opposing theories regarding phytolith-encapsulated metal(loid)s. The first one states that phytoliths can be considered stabilized metal(loid) sinks because the phytolith decomposes more slowly than plant organic matter [8]. The second considers metals stored within phytoliths as a possible leaching source, thus necessitating management for reducing the rate of runoff [9,10,11]. Another hypothesis suggests that phytoliths resistant to degradation would sequester metal(loid)s inside plants, thereby reducing the metal(loid)s-associated toxicity [6,12]. The previous studies mentioned above have been inconclusive on the matter; it is agreed that phytolith-encapsulated metal(loid)s could affect the fate of metal(loid)s and, consequently, the environment at contaminated sites.
Most previous studies on metal(loid) accumulation in phytoliths were conducted with Gramineae species [6,8,10,12]. These are well-known phytolith accumulators and are appropriate models for studying phytoliths. However, the ability of different plant species to accumulate metal(loid)s also needs to be considered, particularly for hyperaccumulators of specific metal(loid)s, to improve our understanding of phytolith-encapsulated metal(loid)s. Pteris multifida is a well-known As hyperaccumulator [13] that is grown and researched in Korea [14,15]. P. multifida phytolith accumulation has also been previously studied [16]. Further, P. multifida is expected to have the potential of storing large amounts of arsenic inside phytoliths. Higher soil water content and plant water uptake can increase Si uptake into plants, as well as its accumulation in phytoliths [17], potentially affecting metal(loid) accumulation in phytoliths. The differences in the phytolith amount and shape were considered in ancient agricultural irrigation [18]. P. multifida is a well-known As hyperaccumulator, with the relationship between soil water and As accumulation being well established [19]. Understanding the differences in As accumulation within P. multifida phytoliths with regard to different soil water contents is of interest for understanding how phytolith-encapsulated As may affect the environment. Comparison of Gramineae and Pteris species can be a guide for formerly studied species and newly suggested species. Two species that are also comparable in As contaminated the metal mine area as widespread abundant plant species and, in most of the studies, Pteris species accumulate more As than Gramineae species [20,21,22,23].
In this study, As accumulations in Phragmites australis and P. multifida were compared for different soil water contents. Comparing P. multifida with P. australis can be a helpful case study to understand possible amount of phytolith encapsulated As and characteristics of As encapsulation in phytolith of As hyper-accumulator. Soil for plant cultivation was collected nearby the Gilgok gold mine, which was mainly contaminated by arsenic. P. australis was grown in soil at 30%, 60%, 100%, and 130% water holding capacity. As P. multifida cannot survive in water-saturated soil, it was grown in soil of 30% and 60% water holding capacity. Soil characteristics (available As and Si, pH), plant characteristics (water content, water potential, and phytolith accumulation), and As accumulation from soil to phytoliths were analyzed in order to understand the effect of soil water content on As accumulation in the phytoliths of the two investigated plant species.
2. Materials and Methods
2.1. Experimental Design
Soil for the experiment was collected near the Gilgok gold mine, which is mainly contaminated with As [24]. The collected soil was 10 mm sieved to remove large size stone [25]. All shoot parts of plants were grown in a greenhouse for 30 days with uncontaminated organic seed beds and the newly grown plants were divided from the rhizome for use in the experiment. The plant root part was clearly washed and then transplanted into a pot with 1 kg of soil from the Gilgok gold mine. Soil with P. australis was irrigated at 30%, 60%, 100%, and 130% water holding capacity. Soil water holding capacity was analyzed by same method with Vengadaramana et al. [26]. In P. multifida, all plants grown under 100% and 130% of water holding capacity died within the first ten days. Thus, only those grown under 30% and 60% of water holding capacity were analyzed. Five plants were planted for each treatment. The water holding capacity of the soil was 34%. All transplanted plants were grown in a greenhouse for 90 days.
2.2. Sample Analysis
2.2.1. Soil Analysis
Control soil before plant cultivation and the soil after cultivation were air-dried and sieved through a 2 mm sieve. The soil pH was measured using a pH meter (Thermo Orion 920A, Thermo Fisher Scientific, Waltham, MA, USA). The soil was mixed with distilled water (1:5) and shaken for 1 h before measurement. Soil texture was determined using the pipette method [27]. The soil was dispersed using a 5% sodium hexametaphosphate solution and sampling aliquot solution according to Stoke’s law. Loss on ignition (LOI) was used to determine the organic matter (OM) content at 400 °C over the course of 16 h [28]. Available silicon was extracted via the method described by Imaizumi and Yoshida [29]. The soil was mixed with the extraction solution (1:10) and shaken in a water bath for 5 h at 40 °C. The extraction solution was made of 0.18 M NaOAc + 0.87 M acetic acid and adjusted to pH 4. Analysis of the pseudo total arsenic (As) was conducted using the microwave aqua-regia method [30]. Soil (0.5 g) was mixed with aqua regia (HNO3:HCl = 1:3) and digested in a microwave for 30 min at 180 °C. Arsenic in soil was also analyzed via Mehlich-3 extraction [31] as an available As. All digested and extracted soil samples were filtered and analyzed using ICP-OES (Agilent, Santa Clara, CA, USA).
2.2.2. Plant Analysis
Plant underground parts were cultivated as root parts and plant aboveground parts were cultivated and the stem and leaf were homogenized as shoot parts. The cultivated parts were rinsed with distilled water and the moisture was wiped. The wiped plant shoot parts were weighed and dried in an oven at 105 °C and weighed again to calculate the plant water content and plant dry weight. Dried plant samples were digested with 5 mL HNO3 and 3 mL H2O2 in a microwave at 180 °C for 30 min [32]. As the research focused on phytoliths, the digested solution was centrifuged and the pellet was further digested with 10 M NaOH [33] until the solution was clean. The two solutions were combined, filtered, and analyzed via ICP-MS (Agilent, Santa Clara, CA, USA).
2.2.3. Phytolith Extraction and Analysis
Both P. multifida and P. australis accumulated phytolith only in the shoot part [4]. Phytoliths were extracted from the shoot as per the dry ashing method [6,34]. The plant material was dried in an oven at 50 °C and moved to a porcelain crucible. The samples were digested in a furnace at 700 °C for 12 h and then transferred to 50 mL polyethylene tubes. HCl (10%) was added to each tube, shaken, and heated in a water bath at 70 °C for 20 min. The tubes were centrifuged at 4000 rpm for 15 min and the supernatant was discarded. The precipitated pellets were rinsed with distilled water and centrifuged three times. 15% H2O2 was put into a rinsed pellet, shaken, heated in a water bath at 70 °C for 20 min, and rinsed with distilled water. The remaining pellet was dried and weighed to determine the ratio of phytoliths in the plant. The extracted phytoliths were mixed with 10 M NaOH [33] and digested in a microwave at 180 °C until the solution was clean. The digested solution was acidified with nitric acid, filtered, and analyzed via ICP-MS (Agilent, Santa Clara, CA, USA). The shape and composition of the phytolith surfaces were analyzed using a ZEISS SUPRA 55VP SEM (Zeiss, Oberkochen, Germany) at 10 kV as well as via energy-dispersive X-ray spectroscopy (EDS).
2.2.4. Arsenic Accumulation Ratio
The accumulation ratio of As from soil to phytoliths was calculated as the ratio between As concentration in the plant shoot and soil obtained via the Mehlich-3 extraction (EFroot).
EFroot = As in plant roots/As in soil
As concentration in the plant shoot and As concentration in soil obtained via Mehlich-3 extraction (EFshoot),
EFshoot = As in plant shoot/As in soil
Concentration in the phytolith and As concentration in soil obtained via Mehlich-3 extraction (EFlith),
EFlith = As in phytolith/As in soil
Translocation factor of As from root to shoot (TF),
TF = As in shoot/As in root
As concentrations in phytoliths and As concentrations in the plant shoot (EFlith/shoot).
EFlith/shoot = As in phytolith/As in shoot
2.3. Statistical Analysis
All analyses were performed for five replicates. Significant differences were determined via Duncan’s test and a mean p < 0.05 indicated statistical significance. Plant As concentration and amount were log-transformed (log(x + 1)) due to the large-scale difference between the two species. All data were analyzed using statistical analysis software (SAS 9.4, SAS Institute Inc., Cary, NC, USA).
3. Results
3.1. Soil Characteristics
Soil characteristics are described in Table 1. Control soil was analyzed via aqua regia extraction and the As concentration was 3258.72 ± 131.53 mg kg−1. The soil texture of the control soil was sandy loam and the OM was 5.71 ± 0.07%. The range of soil As concentration via Mehlich-3 extraction was 237.45–284.58 mg kg−1. All experimental soils had significantly higher As concentrations through Mehlich-3 extraction than the control soil. Soils with 100% and 130% water holding capacity had significantly higher As concentrations through Mehlich-3 extraction than soils with lower water content. Available Si and pH levels were lowest in the control soil, but sections with no statistically significant difference were overlapping and no trend according to soil water content was observed.
3.2. Plant Water Condition and Phytoliths
SEM images and EDS spectra of phytoliths are shown in Figure 1. The shape of the phytoliths was similar to that of the previously imaged phytoliths obtained via high-temperature extraction [9]. Phytoliths were mainly composed of Si and O.
Plant water conditions and phytolith contents are described in Table 2. The phytolith content of P. australis in 30% of water holding capacity was 58,642.31 mg kg−1, which was significantly lower than all other analyzed samples. There was no significant difference in the water content of plant shoots between all samples. Higher soil water content increased the leaf water potential in P. multifida. The water potential in P. multifida with 30% water holding capacity was −11.47 bar and −7.00 bar at 60% of water holding capacity. The water potential of P. australis leaves increased with higher soil water content. The water potential of P. australis leaves at 30% water holding capacity was significantly lower than that of other P. australis leaves. The range of water content for plant roots was 70.49–85.78%. P. multifida had a significantly lower root water content than P. australis. The root water content of P. australis with 30% water holding capacity was 79.36%, the lowest root water content among P. australis samples. There was no significant difference between the other P. australis samples.
3.3. Concentration and Amount of As
The concentration and amount of As in plants and phytoliths are shown in Figure 2. P. australis roots accumulate less As under unsaturated conditions (soil water content < 100%) than saturated conditions (soil water content ≥ 100%) (Figure 2a).
In the case of P. multifida, As concentration in the shoot increased with higher soil water content (Figure 2a). When the soil water contents were 30% and 60% of the water holding capacity, the As concentrations of the P. multifida shoot were 144.54 mg kg−1 and 528.67 mg kg−1, respectively. P. australis did not exhibit a tendency in shoot As concentration based on the soil water content. All As concentrations in the shoot of P. australis were significantly lower than those determined for P. multifida.
In all treatments, As concentration in phytoliths was higher than that in plant shoots (Figure 2a). P. multifida in 60% of water holding capacity had the highest As concentration in phytoliths (1610.74 mg kg−1), while the second highest was observed in 30% of water holding capacity (414.70 mg kg−1). All phytoliths of P. australis had significantly lower As concentrations than those of P. multifida. The range of As in P. australis phytoliths was 41.67–126.54 mg kg−1. P. australis in 30% of water holding capacity had the highest As concentration in phytoliths than other P. australis.
Accumulation of As in the shoot and phytolith parts was described as the total amount in a pot (Figure 2b). P. multifida with 60% water holding capacity had the largest amount of As in the plant shoot (698.67 µg pot−1), followed by P. multifida with 30% water holding capacity (171.98 µg pot−1). The different soil water content of P. australis did not affect the amount of As in plant shoots and there was no significant difference. Arsenic amount in the plant shoot of P. australis was in the range of 9.87–17.25 µg pot−1.
P. multifida with 60% of water holding capacity had the largest amount of As in phytoliths (218.78 µg pot−1), while P. multifida with 30% of water holding capacity had the second-largest amount (57.47 µg pot−1). The soil water content of P. australis did not affect the amount of As in phytoliths and had no significant differences observed. The range of As content in the phytoliths of P. australis was 8.06–4.91 µg pot−1.
3.4. Accumulation of As from Soil to Phytolith
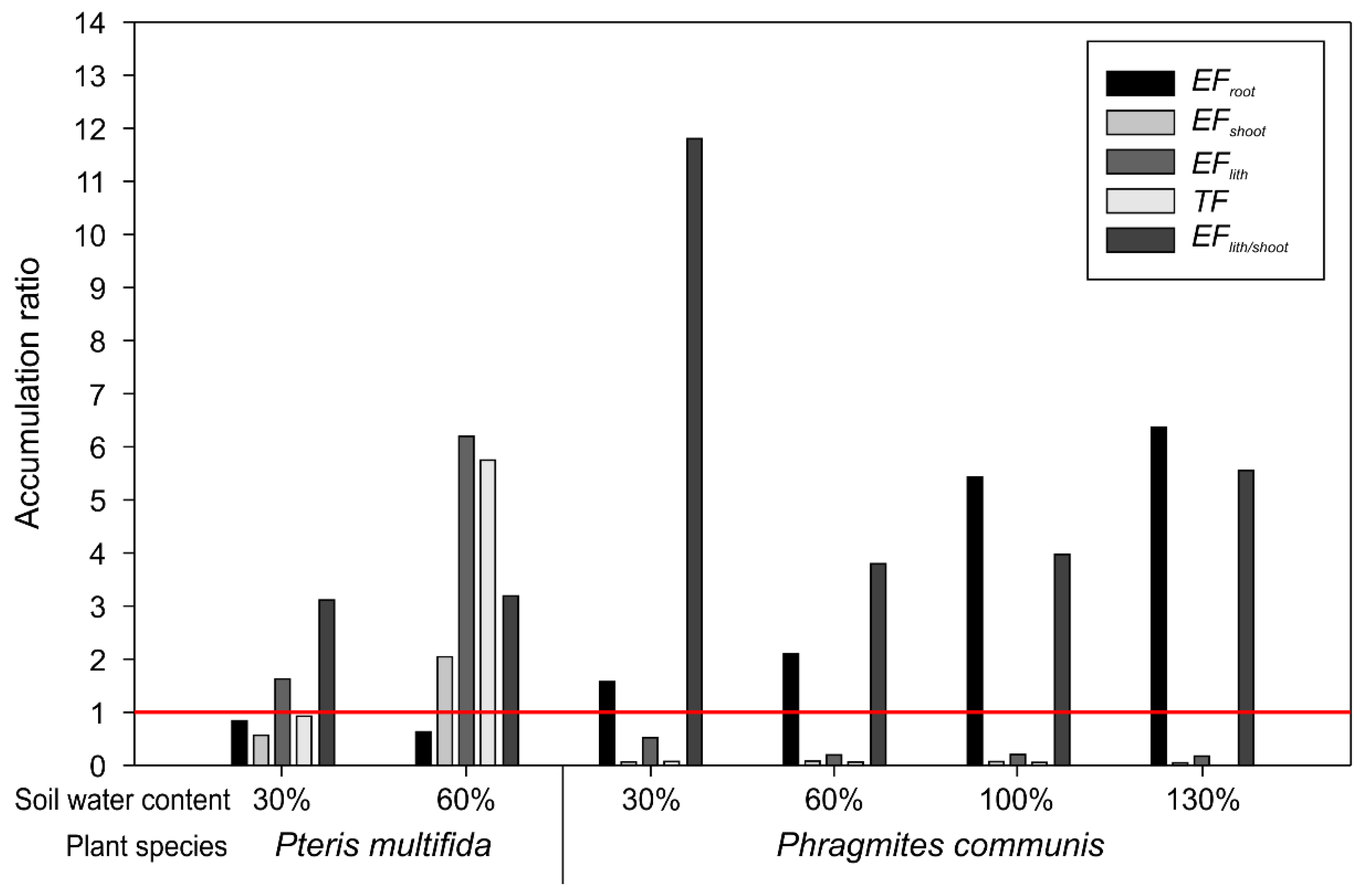
The accumulation ratio of As from soil to phytolith is shown in Figure 3. P. australis with 100% and 130% of water holding capacity had higher EFroot than the other samples. All EFroot values of P. australis were greater than 1, while those for P. multifida were lower than 1. In the case of EFshoot, only the P. multifida with 60% of water holding capacity had a value over 1. There was a large difference in EFshoot between the two species. The range of EFshoot for P. multifida was 0.54–2.04, while that for P. australis was 0.03–0.06. The range of EFlith for P. multifida was 1.61–6.23, and that for P. australis was 0.15–0.51. The range of TF for P. multifida was 0.92–5.77, and that for P. australis was 0.01–0.04. The range of EFlith/shoot, except for P. australis with 30% of water holding capacity, was 3.11–5.56. The EFlith/shoot for P. australis with 30% of water holding capacity was 11.83.
4. Discussion
4.1. Soil Characteristics
When the soil water content exceeded the water holding capacity, the available As increased (Table 1). In water-saturated conditions, the availability of As was reported to increase faster than in unsaturated conditions [35]. The increase in As availability could be the effects of anaerobic and redox conditions. Arsenic is reduced to arsenite, increasing plant availability under saturated conditions [36,37,38,39,40]. The higher As availability at 100% and 130% of water holding capacity can thus be attributed to the effect of soil saturation.
4.2. Plant Water Condition and Phytolith
Although the soil water content did not affect the water content of plant shoots, the water potential of plant leaves and the water content of plant roots indicated that soil water content affected plant water conditions, with major differences observed between 30% and 60% of the soil water holding capacity (Table 2). Soil water content is a major factor in increasing plant water uptake, but increases in the latter cease beyond the limit point and the same water potential value is maintained even when soil moisture increases [41]. In this study, only the phytolith accumulation in P. australis increased with high soil water content between 30% and 60% of soil water holding capacity, while that in P. multifida was not affected. Previous research reported that higher soil water content increases phytolith accumulation in plants, describing the change as “more water—more phytoliths” [42]. However, in most previous studies, the relationship between soil water and phytoliths was analyzed in Gramineae [17,43,44,45], with a major focus on the water management of food crops and phytolith accumulation [17,44,45]. In contrast, no previous research on the relationship between soil water content and phytolith accumulation was available for Pteridaceae species. In the present study, we determined that soil water content did not affect phytolith accumulation in P. multifida as much as in P. australis.
4.3. Concentration of As in Plant
In the soil with 100% and 130% of water holding capacity, plant roots had the highest As concentrations and EFroot values (Figure 2a and Figure 3). This difference can be explained by increased soil As availability, which was obtained via Mehlich-3 extraction (Table 1). However, the As concentration of P. australis shoots did not differ significantly between soil water content conditions (Figure 2a). In the case of P. australis, EFroot increased while TF decreased, and the As concentration in shoots did not show a significant difference. In previous research, the TF of P. australis was lower than 1, with some reported values even being lower than 0.1 [46,47,48,49]. Lee et al. [50] compared the metal accumulation of fern and Graminae, with the latter having a lower TF. Such a characteristic can explain that P. australis is an As excluder and reduces root-to-shoot translocation as a defense mechanism [51]. The result of TF in this study also reflects the characteristics of P. australis.
In the case of P. australis, phytoliths with 30% of water holding capacity had a significantly higher As concentration than other soil water contents and the EFlith/plant of P. australis with 30% of water holding capacity was the highest (Figure 2a and Figure 3). The mechanisms of phytolith formation and As accumulation in phytoliths have not yet been identified. One possible explanation for the concentration difference is that the phytolith content in P. australis decreased at 30% of water holding capacity. The As in phytoliths might have been condensed as a result of this decrease in phytolith content.
Although there was no difference in As availability in the soil, the TF of P. multifida, unlike P. australis, increased more than five times when soil water content increased from 30% to 60% of water holding capacity (Figure 3). Transpiration is one of the major factors affecting As accumulation in Pteris [19]. Higher soil water content can increase plant transpiration [52] and As accumulation differences in P. multifida may be due to differences in soil water contents.
This study aimed to compare the arsenic accumulation capacity of phytoliths in P. multifida and P. australis. In the plant shoot, P. multifida accumulated much more As than P. australis. P. multifida is recognized as an As hyperaccumulator [13], which can explain the observed difference in As concentration between the shoots of P. multifida and P. australis. As accumulation in phytoliths has not yet been studied in fern species and thus the mechanisms and capacity of accumulation remain unclear. However, the As concentration in P. multifida phytoliths was higher than that in shoots and the EFlith/shoot was more than 1 (Figure 2a and Figure 3). It can thus be assumed that P. multifida accumulates As inside phytoliths, similarly to P. australis.
The concentration of As in P. multifida shoots was much higher than that in P. australis shoots (Figure 2). Even though the accumulation ratio from shoot to phytolith was similar, the As concentration of the P. multifida phytolith was higher than that of P. australis. The soil water content did not affect the EFlith/plant of P. multifida (Figure 3), and the increase in As concentration in P. multifida shoots at higher soil water contents directly affected the As concentration in phytoliths. Finally, P. multifida with higher soil water content was in better condition for accumulating As in phytoliths.
4.4. Amount of As in Plant
Both P. multifida and P. australis are perennial plants that grow in rhizomes. In this study, separated rhizomes were newly grown for 90 days. Rhizomes store carbohydrates from old shoots and supply them to newly grown ones [53]. Because plant shoots could not obtain nutrients from the old shoots and the growing time was not enough to represent total As accumulation in plants, it is difficult to determine the actual As accumulation. Nevertheless, we can compare the relative values.
Both P. multifida with 30% and 60% water holding capacity had a greater amount of As in the shoots and phytoliths than P. australis (Figure 2b). EFlith and EFlith/shoot of P. multifida were higher than 1 for all soil water contents. In a previous study, the biomass of Pteris was 13.02 t ha−1, and that of Phragmites was 19.26 t ha−1 [36]. Although the biomass of Phragmites is usually larger than that of fern species, the difference is not large enough to influence As accumulation and P. multifida may act as a better As sink through its phytoliths.
Phytolith encapsulated As has different availability and fate with As in plant organic matter because of the unique characteristics of silicate [6]. The amount of As in the phytoliths of P. multifida increased with soil water content. A large amount of As accumulation suggests that P. multifida may play a role as a hyperaccumulator in the phytolith and P. multifida might affect As fate in soil with phytolith-occluded As. Moreover, the effect on As fate may be greater with higher soil water content. However, since the change in the amount of As in the phytolith changed in proportion to the total concentration of As in the plant, it was difficult to say that it had an effect on the role of arsenic sequestration and lowering toxicity in the plant. The soil water content of the P. australis pot affected the As concentration in phytoliths but did not affect that in shoots (Figure 2a). However, the amount of As in the phytoliths and shoots was not significantly different (Figure 2b). This is because the phytolith increase was offset by the decrease in phytolith content inside the plant, thus eliminating the change in total amount.
5. Conclusions
The effect of soil water content on As accumulation in phytoliths varied depending on the plant species. Higher soil water content increased the As concentration in the shoots of P. multifida. As the ratio of As in shoots and phytoliths did not change with soil water content, we concluded that As accumulation in the phytoliths of P. multifida increased parallel to the soil water content. In the case of P. australis, phytolith As was highest under low water content conditions. However, since phytolith content decreased with higher phytolith As concentrations, the total amount of As stored in phytoliths was not affected by soil water content. P. multifida is an As hyperaccumulator and accumulated As in phytoliths to a greater extent than in plant shoots. The amount of phytolith-encapsulated As was largest for P. multifida under high soil water content. It is not yet clear whether phytolith-encapsulated As can act as a sink of As in soil or a releasing fraction. However, some studies have suggested that trace element accumulation in phytoliths may occur for a significant fraction of soil trace elements. The results of this study suggest that As is highly accumulated in phytoliths under certain conditions, thus contributing to our understanding of As fate in the environment.
Author Contributions
All authors contributed to the study conception and design. Material preparation, data collection, and analysis were performed by H.-G.M., M.-S.K. and J.-G.K. The first draft of the manuscript was written by H.-G.M. and all authors commented on previous versions of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2021R1F1A1051997) and partly by Korea university.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Epstein, E. The anomaly of silicon in plant biology. Proc. Natl. Acad. Sci. USA 1994, 91, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. J. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Tripathi, P.; Tripathi, R.D.; Singh, R.P.; Dwivedi, S.; Goutam, D.; Shri, M.; Trivedi, P.K.; Chakrabarty, D. Silicon mediates arsenic tolerance in rice (Oryza sativa L.) through lowering of arsenic uptake and improved antioxidant defence system. Ecol. Eng. 2013, 52, 96–103. [Google Scholar] [CrossRef]
- Sharma, R.; Kumar, V.; Kumar, R. Distribution of phytoliths in plants: A review. Geol. Ecol. Landsc. 2019, 3, 123–148. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Liu, H.; Strömberg, C.A.; Yang, X.; Zhang, X. Phytolith carbon sequestration in global terrestrial biomes. Sci. Total Environ. 2017, 603, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Delplace, G.; Schreck, E.; Pokrovsky, O.S.; Zouiten, C.; Blondet, I.; Darrozes, J.; Viers, J. Accumulation of heavy metals in phytoliths from reeds growing on mining environments in Southern Europe. Sci. Total Environ. 2020, 712, 135595. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Li, Y.; Jin, Q.; Li, F. Silica nanoparticles inhibit arsenic uptake into rice suspension cells via improving pectin synthesis and the mechanical force of the cell wall. Environ. Sci. Nano 2020, 7, 162–171. [Google Scholar] [CrossRef]
- Fernandes-Horn, H.M.; Sampaio, R.A.; Horn, A.H.; de Oliveira, E.S.A.; Lepsch, I.F.; Bilal, E. Use of Si-Phytoliths in depollution of mining areas in the Cerrado-Caatinga region, MG, Brazil. Int. J. Geomate 2016, 11, 2216–2221. [Google Scholar]
- Nguyen, T.N.; Nguyen, M.N.; McNamara, M.; Dultz, S.; Meharg, A.; Nguyen, V.T. Encapsulation of lead in rice phytoliths as a possible pollutant source in paddy soils. Environ. Exp. Bot. 2019, 162, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.N.; Dam, T.T.N.; Nguyen, A.T.Q.; Nguyen, A.M.; Nguyen, L.N.; Duong, L.T.; Dang, Q.T.; Tran, T.T. Arsenic in rice straw phytoliths: Encapsulation and release properties. Appl. Geochem. 2021, 127, 104907. [Google Scholar] [CrossRef]
- Nguyen, M.N.; Nguyen, A.T.; Dultz, S.; Tsubota, T.; Duong, L.T.; Nguyen, A.M.; Pham, N.T. Thermal induced changes of rice straw phytolith in relation to arsenic release: A perspective of rice straw arsenic under open burning. J. Environ. Manag. 2022, 304, 114294. [Google Scholar] [CrossRef] [PubMed]
- de Melo Farnezi, M.M.; de Barros Silva, E.; Lopes Dos Santos, L.; Silva, A.C.; Grazziotti, P.H.; Prochnow, J.T.; Pereira, I.M.; Fontan, I.D.C.I. Potential of grasses in phytolith production in soils contaminated with cadmium. Plants 2020, 9, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, W.; Li, Z.; Zou, B.; Peng, S. Pteris multifida Poir., a new arsenic hyperaccumulator: Characteristics and potential. Int. J. Environ. Pollut. 2005, 23, 388–396. [Google Scholar] [CrossRef]
- Han, J.H.; Kwon, H.J.; Lee, C.H. Effect of Arsenic Types in Soil on Growth and Arsenic Accumulation of Pteris multifida. Korean J. Plant Resour. 2014, 27, 344–353. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.W.; Seo, J.Y.; Oh, W.K.; Sung, S.H. Anti-neuroinflammatory ent-kaurane diterpenoids from Pteris multifida roots. Molecules 2017, 22, 27. [Google Scholar] [CrossRef] [Green Version]
- Sundue, M. Silica bodies and their systematic implications in Pteridaceae (Pteridophyta). Bot. J. Linn. Soc. 2009, 161, 422–435. [Google Scholar] [CrossRef]
- Madella, M.; Jones, M.K.; Echlin, P.; Powers-Jones, A.; Moore, M. Plant water availability and analytical microscopy of phytoliths: Implications for ancient irrigation in arid zones. Quat. Int. 2009, 193, 32–40. [Google Scholar] [CrossRef]
- Rosen, A.M.; Weiner, S. Identifying ancient irrigation: A new method using opaline phytoliths from emmer wheat. J. Archaeol. Sci. 1994, 21, 125–132. [Google Scholar] [CrossRef]
- Wan, X.M.; Lei, M.; Chen, T.B.; Yang, J.X.; Liu, H.T.; Chen, Y. Role of transpiration in arsenic accumulation of hyperaccumulator Pteris vittata L. Environ. Sci. Pollut. Res. 2015, 22, 16631–16639. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Maah, M.J.; Yusoff, I. Evaluation of natural phytoremediation process occurring at ex-tin mining catchment. Chiang Mai J. Sci. 2013, 40, 198–213. [Google Scholar]
- Praveen, A.; Mehrotra, S.; Singh, N. Rice planted along with accumulators in arsenic amended plots reduced arsenic uptake in grains and shoots. Chemosphere 2017, 184, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Praveen, A.; Mehrotra, S.; Singh, N. Mixed plantation of wheat and accumulators in arsenic contaminated plots: A novel way to reduce the uptake of arsenic in wheat and load on antioxidative defence of plant. Ecotoxicol. Environ. Saf. 2019, 182, 109462. [Google Scholar] [CrossRef] [PubMed]
- Raj, A.; Jamil, S.; Srivastava, P.K.; Tripathi, R.D.; Sharma, Y.K.; Singh, N. Feasibility Study of Phragmites karka and Christella dentata Grown in West Bengal as Arsenic Accumulator. Int. J. Phytoremediat. 2015, 17, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Lee, S.H.; Kim, J.G. Assessment of fraction and mobility of arsenic in soil near the mine Waste Dam. Sustainability 2020, 12, 1480. [Google Scholar] [CrossRef] [Green Version]
- Madyiwa, S.; Chimbari, M.J.; Schutte, F. Lead and cadmium interactions in Cynodon nlemfuensis and sandy soil subjected to treated wastewater application under greenhouse conditions. Phys. Chem. Earth 2004, 29, 1043–1048. [Google Scholar] [CrossRef]
- Vengadaramana, A.; Jashothan, P.T.J. Effect of organic fertilizers on the water holding capacity of soil in different terrains of Jaffna peninsula in Sri Lanka. J. Nat. Prod. Plant Resour. 2012, 2, 500–503. [Google Scholar]
- NIAST. Method of SOIL and Plant Analysis; National Institute of Agricultural Science and Technology, Rural Development Administration: Suwon, Korea, 2000.
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; Part 3 Chemical Methods, 5.3.; Wiley: New York, NY, USA, 1996; pp. 961–1010. [Google Scholar]
- Imaizumi, K.; Yoshida, S. Edaphological studies on silicon supplying power of paddy fields. Bull. Natl. Inst. Agric. Sci. 1958, 8, 261–304. [Google Scholar]
- Tighe, M.; Lockwood, P.; Wilson, S.; Lisle, L. Comparison of digestion methods for ICP-OES analysis of a wide range of analytes in heavy metal contaminated soil samples with specific reference to arsenic and antimony. Commun. Soil Sci. Plant Anal. 2004, 35, 1369–1385. [Google Scholar] [CrossRef]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Gulz, P.A.; Gupta, S.K.; Schulin, R. Arsenic accumulation of common plants from contaminated soils. Plant Soil 2005, 272, 337–347. [Google Scholar] [CrossRef]
- Li, Z.; Song, Z.; Cornelis, J.T. Impact of rice cultivar and organ on elemental composition of phytoliths and the release of bio-available silicon. Front. Plant Sci. 2014, 5, 529. [Google Scholar] [CrossRef] [PubMed]
- Parr, J.F.; Lentfer, C.J.; Boyd, W.E. A comparative analysis of wet and dry ashing techniques for the extraction of phytoliths from plant material. J. Archaeol. Sci. 2001, 28, 875–886. [Google Scholar] [CrossRef]
- Onken, B.M.; Adriano, D.C. Arsenic Availability in Soil with Time under Saturated and Subsaturated Conditions. Soil Sci. Soc. Am. J. 1997, 61, 746–752. [Google Scholar] [CrossRef]
- Ascar, L.; Ahumada, I.; Richter, P. Influence of redox potential (Eh) on the availability of arsenic species in soils and soils amended with biosolid. Chemosphere 2008, 72, 1548–1552. [Google Scholar] [CrossRef]
- Meharg, A.A. Arsenic in rice–understanding a new disaster for South-East Asia. Trends Plant Sci. 2004, 9, 415–417. [Google Scholar] [CrossRef]
- Meharg, A.A.; Jardine, L. Arsenite transport into paddy rice (Oryza sativa) roots. New Phytol. 2003, 157, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syu, C.H.; Huang, C.C.; Jiang, P.Y.; Lee, C.H.; Lee, D.Y. Arsenic accumulation and speciation in rice grains influenced by arsenic phytotoxicity and rice genotypes grown in arsenic-elevated paddy soils. J. Hazard. Mater. 2015, 286, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.J.; Ma, J.F.; Meharg, A.A.; McGrath, S.P. Arsenic uptake and metabolism in plants. New Phytol. 2009, 181, 777–794. [Google Scholar] [CrossRef]
- Teuling, A.J.; Uijlenhoet, R.; Hupert, F.; Troch, P.A. Impact of plant water uptake strategy on soil moisture and evapotranspiration dynamics during drydown. Geophys. Res. Lett. 2006, 33, 3–7. [Google Scholar] [CrossRef] [Green Version]
- Katz, O.; Lev-Yadun, S.; Pua Bar, K. Plasticity and variability in the patterns of phytolith formation in Asteraceae species along a large rainfall gradient in Israel. Flora Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 438–444. [Google Scholar] [CrossRef]
- Webb, E.A.; Longstaffe, F.J. The relationship between phytolith- and plant-water δ 18O values in grasses. Geochim. Cosmochim. Acta 2003, 67, 1437–1449. [Google Scholar] [CrossRef]
- Jenkins, E.; Jamjoum, K.; Nuimat, S.; Stafford, R.; Nortcliff, S.; Mithen, S. Identifying ancient water availability through phytolith analysis: An experimental approach. J. Archaeol. Sci. 2016, 73, 82–93. [Google Scholar] [CrossRef] [Green Version]
- Meunier, J.D.; Barboni, D.; Anwar-ul-Haq, M.; Levard, C.; Chaurand, P.; Vidal, V.; Grauby, O.; Huc, R.; Laffont-Schwob, I.; Rabier, J.; et al. Effect of phytoliths for mitigating water stress in durum wheat. New Phytol. 2017, 215, 229–239. [Google Scholar] [CrossRef] [Green Version]
- Ghassemzadeh, F.; Yousefzadeh, H.; Arbab-Zavar, M.H. Arsenic phytoremediation by Phragmites australis: Green technology. Int. J. Environ. Stud. 2008, 65, 587–594. [Google Scholar] [CrossRef]
- Ghassemzadeh, F.; Yousefzadeh, H.; Arbab-Zavar, M.H. Removing arsenic and antimony by Phragmites australis: Rhizofiltration technology. J. Appl. Sci. 2008, 8, 1668–1675. [Google Scholar] [CrossRef] [Green Version]
- Štrbac, S.; Šajnović, A.; Kašanin, G.M.; Vasić, N.; Dojčinović, B.; Simonović, P.; Jovančićević, B. Metals in sediment and phragmites Australis (common reed) from tisza river, Serbia. Appl. Ecol. Environ. Res. 2014, 12, 105–122. [Google Scholar] [CrossRef]
- Castaldi, P.; Silvetti, M.; Manzano, R.; Brundu, G.; Roggero, P.P.; Garau, G. Mutual effect of Phragmites australis, Arundo donax and immobilization agents on arsenic and trace metals phytostabilization in polluted soils. Geoderma 2018, 314, 63–72. [Google Scholar] [CrossRef]
- Lee, S.H.; Ji, W.; Lee, W.S.; Koo, N.; Koh, I.H.; Kim, M.S.; Park, J.S. Influence of amendments and aided phytostabilization on metal availability and mobility in Pb, Zn mine tailings. J. Environ. Manag. 2014, 139, 15–21. [Google Scholar] [CrossRef]
- Prica, M.; Andrejić, G.; Šinžar-Sekulić, J.; Rakić, T.; Dželetović, Ž. Bioaccumulation of heavy metals in common reed (Phragmites australis) growing spontaneously on highly contaminated mine tailing ponds in Serbia and potential use of this species in phytoremediation. Bot. Serb. 2019, 43, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Gardner, W.R.; Ehlig, C.F. The influence of soil water on transpiration by plants. J. Geophys. Res. 1963, 68, 5719–5724. [Google Scholar] [CrossRef]
- Sakamaki, Y.; Ino, Y. Tubers and rhizome fragments as propagules: Competence for vegetative reproduction in Equisetum arvense. J. Plant Res. 2006, 119, 677–683. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
SEM images and EDS spectra of phytoliths from (a) P. multifida at 60% water holding capacity soil and (b) P. australis at 60% water holding capacity soil.
Figure 1.
SEM images and EDS spectra of phytoliths from (a) P. multifida at 60% water holding capacity soil and (b) P. australis at 60% water holding capacity soil.

Figure 2.
Arsenic in Pteris multifida and Phragmites australis as a function of the water content. (a) As concentration in plant shoot, phytolith, and roots. As in the shoot and roots was calculated based on plant dry weight, and As in phytoliths was calculated based on phytolith weight. (b) The amount of As in plant shoots and phytoliths for each pot. Water content means the percentage of water holding capacity. Different letters in the same color bar indicate significant differences at the 5% level determined via Duncan’s test after log-scale transformation.
Figure 2.
Arsenic in Pteris multifida and Phragmites australis as a function of the water content. (a) As concentration in plant shoot, phytolith, and roots. As in the shoot and roots was calculated based on plant dry weight, and As in phytoliths was calculated based on phytolith weight. (b) The amount of As in plant shoots and phytoliths for each pot. Water content means the percentage of water holding capacity. Different letters in the same color bar indicate significant differences at the 5% level determined via Duncan’s test after log-scale transformation.

Figure 3.
Arsenic accumulation ratio from soil to plant in Pteris multifida and Phragmites australis as a function of the water content. Soil water content means the percentage of water holding capacity. Accumulation ratio of 1 is marked with a red horizontal line. The EFroot is As concentration in root per As concentration in soil determined via Mehlich-3 extraction. EFshoot is As concentration in shoot per As concentration in soil determined via Mehlich-3 extraction. EFlith is As concentration in phytolith per As concentration in soil determined via Mehlich-3 extraction. TF is As concentration in plant shoot per As concentration in plant root. EFlith/shoot is As concentration in phytolith per As concentration in soil determined via Mehlich-3 extraction.
Figure 3.
Arsenic accumulation ratio from soil to plant in Pteris multifida and Phragmites australis as a function of the water content. Soil water content means the percentage of water holding capacity. Accumulation ratio of 1 is marked with a red horizontal line. The EFroot is As concentration in root per As concentration in soil determined via Mehlich-3 extraction. EFshoot is As concentration in shoot per As concentration in soil determined via Mehlich-3 extraction. EFlith is As concentration in phytolith per As concentration in soil determined via Mehlich-3 extraction. TF is As concentration in plant shoot per As concentration in plant root. EFlith/shoot is As concentration in phytolith per As concentration in soil determined via Mehlich-3 extraction.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Soil As, Si concentration and pH *, **.
| Treatments | Results | ||||||
|---|---|---|---|---|---|---|---|
| Plant Species | Soil Water Content a | Soil Available As b (mg kg−1) | Soil Available Si c (mg kg−1) | Soil pH | |||
| Control d | 237.45 | d | 203.43 | c | 8.51 | c | |
| Pteris multifida | 30% | 256.03 | bc | 220.50 | abc | 8.63 | abc |
| 60% | 258.77 | b | 213.64 | bc | 8.58 | bc | |
| Phragmites australis | 30% | 249.86 | c | 228.76 | ab | 8.71 | a |
| 60% | 255.88 | bc | 233.74 | a | 8.63 | abc | |
| 100% | 284.58 | a | 223.59 | ab | 8.67 | ab | |
| 130% | 280.67 | a | 216.93 | abc | 8.58 | bc | |
* Pseudo-total As concentration in control soil with aqua regia extraction was 3258.72 ± 131.53 mg kg−1. ** A different letter in the same column indicates significant differences at the 5% level by Duncan’s test. a—Soil water content indicates the percentage of water holding capacity. b—Soil available As is As concentration with Mehlich-3 extraction. c—Soil available Si was extracted with acetic acid buffer. d—Control is composite soil before plant growth experiments.
Table 2.
Plant water content water potential and phytolith content *.
| Treatments | Results | ||||||
|---|---|---|---|---|---|---|---|
| Plant Species | Soil Water Content a | Phytolith Content (mg kg−1) | Water Content in Plant Shoot (%) | Water Potential in Plant Leaf (bar) | Water Content in Plant Root (%) | ||
| Pteris multifida | 30% | 114,157.51 a | 64.41 a | −11.47 | cd | 70.49 | c |
| 60% | 104,516.49 a | 69.78 a | −7.00 | a | 72.06 | c | |
| Phragmites australis | 30% | 58,642.31 b | 69.61 a | −13.60 | d | 79.36 | b |
| 60% | 137,370.92 a | 67.78 a | −10.73 | c | 84.32 | a | |
| 100% | 116,530.55 a | 68.28 a | −10.40 | bc | 85.08 | a | |
| 130% | 103,573.27 a | 69.15 a | −8.87 | ab | 85.78 | a | |
* Different letter in same column indicates significant differences at the 5% level determined via Duncan’s test. a Soil water content indicates percentage of water holding capacity.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Min, H.-G.; Kim, M.-S.; Kim, J.-G. Effect of Soil Water Contents on Arsenic Accumulation in Phytoliths of Pteris multifida and Phragmites australis. Appl. Sci. 2022, 12, 12518. https://0-doi-org.brum.beds.ac.uk/10.3390/app122412518
AMA Style
Min H-G, Kim M-S, Kim J-G. Effect of Soil Water Contents on Arsenic Accumulation in Phytoliths of Pteris multifida and Phragmites australis. Applied Sciences. 2022; 12(24):12518. https://0-doi-org.brum.beds.ac.uk/10.3390/app122412518
Chicago/Turabian StyleMin, Hyun-Gi, Min-Suk Kim, and Jeong-Gyu Kim. 2022. "Effect of Soil Water Contents on Arsenic Accumulation in Phytoliths of Pteris multifida and Phragmites australis" Applied Sciences 12, no. 24: 12518. https://0-doi-org.brum.beds.ac.uk/10.3390/app122412518
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.