
Two-Wavelength Computational Holography for Aberration-Corrected Simultaneous Optogenetic Stimulation and Inhibition of In Vitro Biological Samples

 , and
, and
Abstract
:1. Introduction
- Single-cell resolution. Using wide-field illumination in neural networks will lead to an overall, uncoordinated increase of action-potential activity, making it almost impossible to identify single signal paths. The local stimulation of a single cell can precisely define the origin of action potentials within a network and help to reduce ambiguity.
- 3D stimulation. The focal spots are placed arbitrarily in 3D space. In cardio-optogenetics, many experiments were performed on cardiomyocyte monolayers, i.e., 2D samples [4,7,8,9,21,22]. Similarly, investigations on neuronal networks and network connectivity are often conducted on multi-electrode arrays in 2D cultures [23,24]. The ability to place the stimulating light focus in an arbitrary and defined depth can help to conquer the third dimension and to address single cells in multi-layer cell systems [25].
- Patterned illumination. The concurrent activation or inhibition of a well-defined group of cells opens further possibilities in experimental studies as well as optogenetic applications. In cardio-optogenetics, a dynamic, i.e., temporally changing distributed pattern can control the propagation of the excitation wavefront in cardiac tissue and induce spiral waves of defined speed, direction and chirality [8]. Regarding implantable optogenetic defibrillators ore pacemakers, an optimized pattern can help minimize the required illumination energy and hence reduce energy consumption [10].
- Two-wavelength illumination. The ability to express activating and inhibiting opsins at the same time in one cell or cell network opens new experimental possibilities. Activating single cells or cell groups while concurrently silencing other well-defined parts in a network can help to elucidate signaling pathways in neuronal networks or to study the propagation of the excitation wavefront at hearts. For such experiments, two patterns of different wavelengths should be displayable individually and independently.
- Fast switching times. High temporal resolution in the millisecond range is necessary or advantageous in various experiments: In neural optogenetics, it should be of the same order of magnitude as the action-potential frequencies; in cardio-optogenetics, it is desired to achieve a smooth transition between successive images when displaying spatio–temporal patterns. Furthermore, high pattern update rates are a fundamental prerequisite in experiments that employ an optoelectronic closed-loop feedback control, e.g., in cardio-optogenetics, where the trajectory of the excitation wavefront is measured, processed and used to alter the illumination pattern [7].
2. Experimental Setup
3. Estimation of Minimum Achievable Spot Size
4. Spatial Pattern Formation and Correction of System Aberrations
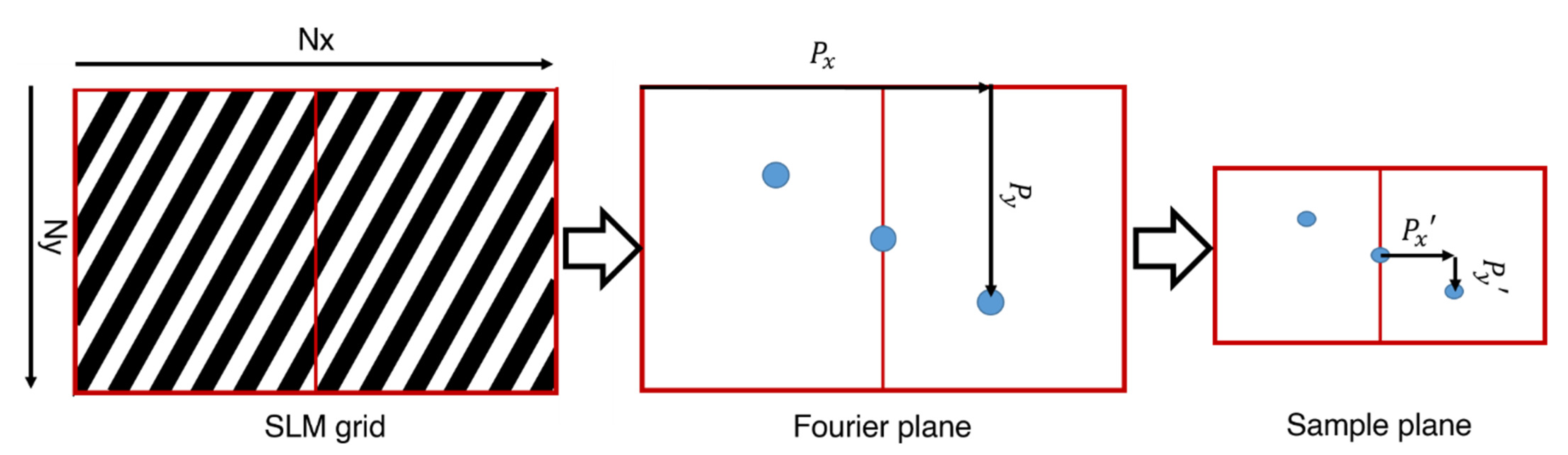
5. Generation of Multiple Spots across the Field of View
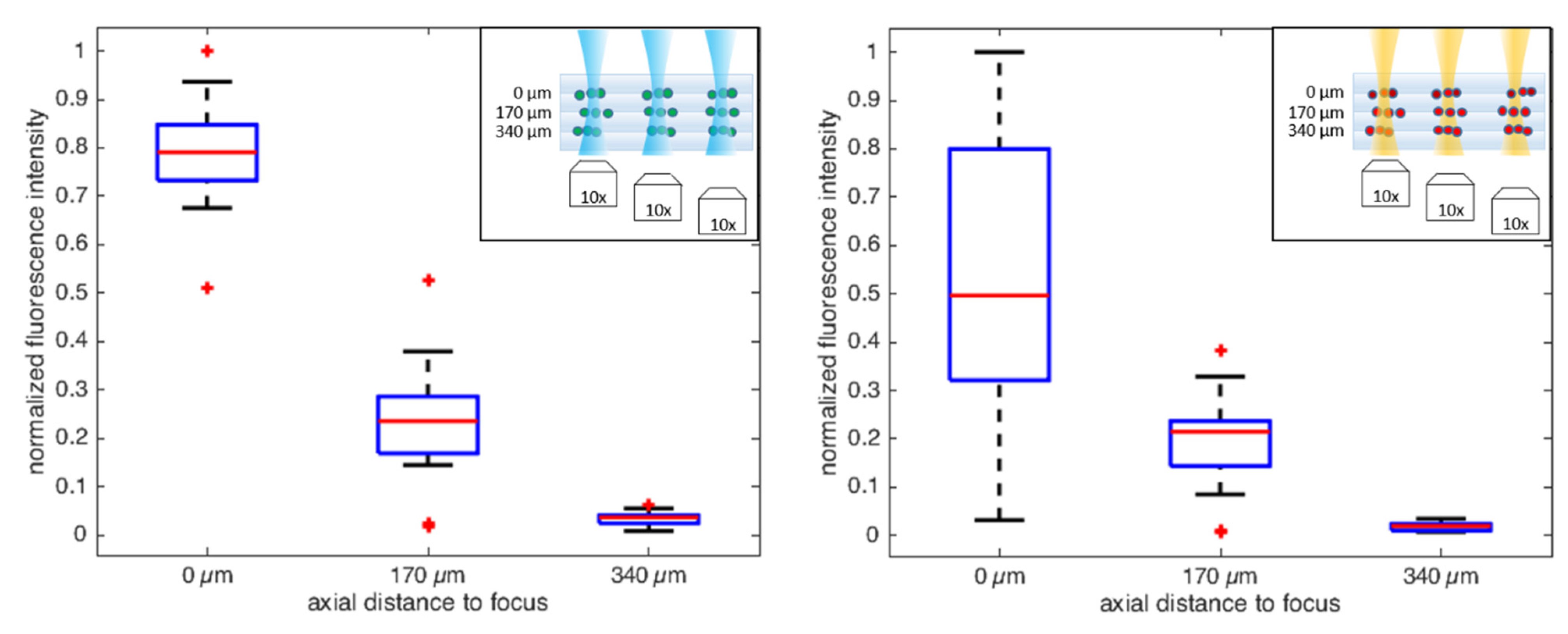
6. Excitation of Single/Multiple Fluorescent Beads in 2D/3D
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagel, G.; Ollig, D.; Fuhrmann, M.; Kateriya, S.; Musti, A.M.; Bamberg, E.; Hegemann, P. Channelrhodopsin-1: A Light-Gated Proton Channel in Green Algae. Science 2002, 296, 2395–2398. [Google Scholar] [CrossRef] [PubMed]
- Nagel, G.; Szellas, T.; Huhn, W.; Kateriya, S.; Adeishvili, N.; Berthold, P.; Ollig, D.; Hegemann, P.; Bamberg, E. Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc. Natl. Acad. Sci. USA 2003, 100, 13940–13945. [Google Scholar] [CrossRef] [Green Version]
- Boyden, E.S.; Zhang, F.; Bamberg, E.; Nagel, G.; Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 2005, 8, 1263–1268. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, T.; Kunihiro, T.; Ando, T.; Kobayashi, S.; Matsui, E.; Yada, H.; Kanda, Y.; Kurokawa, J.; Furukawa, T. Image-based evaluation of contraction–relaxation kinetics of human-induced pluripotent stem cell-derived cardiomyocytes: Correlation and complementarity with extracellular electrophysiology. J. Mol. Cell. Cardiol. 2014, 77, 178–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Shea, C.; Holmes, A.P.; Winter, J.; Correia, J.; Ou, X.; Dong, R.; He, S.; Kirchhof, P.; Fabritz, L.; Rajpoot, K.; et al. Cardiac Optogenetics and Optical Mapping—Overcoming Spectral Congestion in All-Optical Cardiac Electrophysiology. Front. Physiol. 2019, 10, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckley, C.; Carvalho, M.T.; Young, L.; Rider, S.A.; McFadden, C.; Berlage, C.; Verdon, R.F.; Taylor, J.M.; Girkin, J.M.; Mullins, J.J. Precise spatio-temporal control of rapid optogenetic cell ablation with mem-KillerRed in Zebrafish. Sci. Rep. 2017, 7, 5096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Entcheva, E.; Kay, M.W. Cardiac optogenetics: A decade of enlightenment. Nat. Rev. Cardiol. 2021, 18, 349–367. [Google Scholar] [CrossRef]
- Burton, R.A.B.; Klimas, A.; Ambrosi, C.M.; Tomek, J.; Corbett, A.; Entcheva, E.; Bub, G. Optical control of excitation waves in cardiac tissue. Nat. Photonics 2015, 9, 813–816. [Google Scholar] [CrossRef]
- Bingen, B.O.; Engels, M.C.; Schalij, M.J.; Jangsangthong, W.; Neshati, Z.; Feola, I.; Ypey, D.L.; Askar, S.F.; Panfilov, A.; Pijnappels, D.A.; et al. Light-induced termination of spiral wave arrhythmias by optogenetic engineering of atrial cardiomyocytes. Cardiovasc. Res. 2014, 104, 194–205. [Google Scholar] [CrossRef] [Green Version]
- Crocini, C.; Ferrantini, C.; Coppini, R.; Scardigli, M.; Yan, P.; Loew, L.M.; Smith, G.; Cerbai, E.; Poggesi, C.; Pavone, F.S.; et al. Optogenetics design of mechanistically-based stimulation patterns for cardiac defibrillation. Sci. Rep. 2016, 6, 35628. [Google Scholar] [CrossRef]
- Nyns, E.C.; Kip, A.; Bart, C.I.; Plomp, J.J.; Zeppenfeld, K.; Schalij, M.J.; de Vries, A.A.; Pijnappels, D.A. Optogenetic termination of ventricular arrhythmias in the whole heart: Towards biological cardiac rhythm management. Eur. Hear. J. 2016, 38, 2132–2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nussinovitch, U.; Gepstein, L. Optogenetics for in vivo cardiac pacing and resynchronization therapies. Nat. Biotechnol. 2015, 33, 750–754. [Google Scholar] [CrossRef] [PubMed]
- Schmieder, F.; Habibey, R.; Busskamp, V.; Büttner, L.; Czarske, J.W. Analysis of human iPSC-derived neuronal networks (hiPSCNN) using holographic single cell and full field optogenetic stimulation. In Biophotonics Congress 2021; Paper BM1B.4; Optical Society of America: Washington DC, USA, 2021. [Google Scholar]
- Nabavi, S.; Fox, R.; Proulx, C.D.; Lin, J.; Tsien, R.Y.; Malinow, R. Engineering a memory with LTD and LTP. Nature 2014, 511, 348–352. [Google Scholar] [CrossRef]
- Schmieder, F.; Habibey, R.; Busskamp, V.; Büttner, L.; Czarske, J.W. Investigation of in vitro human iPSC-derived neuronal networks using holographic stimulation (Conference Presentation). In Optogenetics and Optical Manipulation; Proc. SPIE 11227, 112270D; SPIE: Bellingham, WA, USA, 2020. [Google Scholar]
- Reutsky-Gefen, I.; Golan, L.; Farah, N.; Schejter, A.; Tsur, L.; Brosh, I.; Shoham, S. Holographic optogenetic stimulation of patterned neuronal activity for vision restoration. Nat. Commun. 2013, 4, 1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, L.; Dahmann, C. Increased lateral tension is sufficient for epithelial folding in Drosophila. Development 2020, 147, 194316. [Google Scholar] [CrossRef] [PubMed]
- Osaki, T.; Uzel SG, M.; Kamm, R.D. Microphysiological 3D model of amyotrophic lateral sclerosis (ALS) from human iPS-derived muscle cells and optogenetic motor neurons. Sci. Adv. 2018, 4, eaat5847. [Google Scholar] [CrossRef] [Green Version]
- Maschio, M.D.; Donovan, J.C.; Helmbrecht, T.O.; Baier, H. Linking Neurons to Network Function and Behavior by Two-Photon Holographic Optogenetics and Volumetric Imaging. Neuron 2017, 94, 774–789.e5. [Google Scholar] [CrossRef]
- Kim, N.Y.; Lee, S.; Yu, J.; Kim, N.; Won, S.S.; Park, H.; Heo, W.D. Optogenetic control of mRNA localization and translation in live cells. Nat. Cell Biol. 2020, 22, 341–352. [Google Scholar] [CrossRef]
- Majumder, R.; Feola, I.; Teplenin, A.S.; Vries, A.A.F.D.; Panfilov, A.V.; A Pijnappels, D. Optogenetics enables real-time spatiotemporal control over spiral wave dynamics in an excitable cardiac system. eLife 2018, 7, 41076. [Google Scholar] [CrossRef]
- McNamara, H.M.; Dodson, S.; Huang, Y.-L.; Miller, E.W.; Sandstede, B.; Cohen, A.E. Geometry-Dependent Arrhythmias in Electrically Excitable Tissues. Cell Syst. 2018, 7, 359–370.e6. [Google Scholar] [CrossRef] [Green Version]
- Klapper, S.D.; Sauter, E.J.; Swiersy, A.; Hyman, M.A.E.; Bamann, C.; Bamberg, E.; Busskamp, V. On-demand optogenetic activation of human stem-cell-derived neurons. Sci. Rep. 2017, 7, 14450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Ryu, J.R.; Lee, B.; Chae, U.; Son, J.W.; Park, B.H.; Cho, I.-J.; Sun, W. Interpreting the Entire Connectivity of Individual Neurons in Micropatterned Neural Culture With an Integrated Connectome Analyzer of a Neuronal Network (iCANN). Front. Neuroanat. 2021, 15, 746057. [Google Scholar] [CrossRef] [PubMed]
- Pelkonen, A.; Pistono, C.; Klecki, P.; Gómez-Budia, M.; Dougalis, A.; Konttinen, H.; Stanová, I.; Fagerlund, I.; Leinonen, V.; Korhonen, P.; et al. Functional Characterization of Human Pluripotent Stem Cell-Derived Models of the Brain with Microelectrode Arrays. Cells 2021, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Papagiakoumou, E. Optical developments for optogenetics. Biol. Cell 2013, 105, 443–464. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Fajardo, O.; Shum, J.; Schärer, Y.-P.Z.; Friedrich, R.W. High-resolution optical control of spatiotemporal neuronal activity patterns in zebrafish using a digital micromirror device. Nat. Protoc. 2012, 7, 1410–1425. [Google Scholar] [CrossRef]
- Ciccone, G.; Meloni, I.; Lahore, R.G.F.; Vierock, J.; Reineke, S.; Kleemann, H.; Hegemann, P.; Leo, K.; Murawski, C. Tailoring Organic LEDs for Bidirectional Optogenetic Control via Dual-Color Switching. Adv. Funct. Mater. 2021, 2110590, 2110590. [Google Scholar] [CrossRef]
- Murawski, C.; Pulver, S.R.; Gather, M.C. Segment-specific optogenetic stimulation in Drosophila melanogaster with linear arrays of organic light-emitting diodes. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Ferraro, P.; Grilli, S.; Miccio, L.; Alfieri, D.; De Nicola, S.; Finizio, A.; Javidi, B. Full Color 3-D Imaging by Digital Holography and Removal of Chromatic Aberrations. J. Disp. Technol. 2008, 4, 97–100. [Google Scholar] [CrossRef]
- Ibrahim, D. Simultaneous dual-wavelength digital holographic microscopy with compensation of chromatic aberration for accurate surface characterization. Appl. Opt. 2019, 58, 6388–6395. [Google Scholar] [CrossRef]
- Khodadad, D.; Bergström, P.; Hällstig, E.; Sjödahl, M. Fast and robust automatic calibration for single-shot dual-wavelength digital holography based on speckle displacements. Appl. Opt. 2015, 54, 5003–5010. [Google Scholar] [CrossRef]
- A Matkivsky, V.; A Moiseev, A.; Gelikonov, G.V.; Shabanov, D.V.; A Shilyagin, P.; Gelikonov, V.M. Correction of aberrations in digital holography using the phase gradient autofocus technique. Laser Phys. Lett. 2016, 13, 35601. [Google Scholar] [CrossRef]
- Kontenis, G.; Gailevičius, D.; Jonušauskas, L.; Jonušauskas, L.; Purlys, V. Dynamic aberration correction via spatial light modulator (SLM) for femtosecond direct laser writing: Towards spherical voxels. Opt. Express 2020, 28, 27850–27864. [Google Scholar] [CrossRef] [PubMed]
- Radner, H.; Büttner, L.; Czarske, J. Interferometric velocity measurements through a fluctuating phase boundary using two Fresnel guide stars. Opt. Lett. 2015, 40, 3766–3769. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Lee, M.; Lee, K.; Kim, N.; Kim, J.M.; Park, J.; Yu, H.; Choi, C.; Heo, W.D.; Park, Y. Optogenetic control of cell signaling pathway through scattering skull using wavefront shaping. Sci. Rep. 2015, 5, 13289. [Google Scholar] [CrossRef] [Green Version]
- Vellekoop, I.M. Feedback-based wavefront shaping. Opt. Express 2015, 23, 12189–12206. [Google Scholar] [CrossRef]
- Schmieder, F.; Klapper, S.D.; Koukourakis, N.; Busskamp, V.; Czarske, J.W. Optogenetic Stimulation of Human Neural Networks Using Fast Ferroelectric Spatial Light Modulator—Based Holographic Illumination. Appl. Sci. 2018, 8, 1180. [Google Scholar] [CrossRef] [Green Version]
- Rothe, S.; Radner, H.; Koukourakis, N.; Czarske, J.W. Transmission Matrix Measurement of Multimode Optical Fibers by Mode-Selective Excitation Using One Spatial Light Modulator. Appl. Sci. 2019, 9, 195. [Google Scholar] [CrossRef] [Green Version]
- Haufe, D.; Koukourakis, N.; Büttner, L.; Czarske, J.W. Transmission of Multiple Signals through an Optical Fiber Using Wavefront Shaping. J. Vis. Exp. 2017, 55407. [Google Scholar] [CrossRef]
- Schmieder, F.; Czarske, J.; Büttner, L. Adaptive laser-induced ultrasound generation using a micro-mirror array spatial light modulator. Opt. Express 2016, 24, 22536. [Google Scholar] [CrossRef]
- Buettner, L.; Thümmler, M.; Czarske, J. Velocity measurements with structured light transmitted through a multimode optical fiber using digital optical phase conjugation. Opt. Express 2020, 28, 8064–8075. [Google Scholar] [CrossRef]
- Kuschmierz, R.; Scharf, E.; Ortegón-González, D.F.; Glosemeyer, T.; Czarske, J.W. Ultra-thin 3D lensless fiber endoscopy using diffractive optical elements and deep neural networks. Light. Adv. Manuf. 2021, 2, 1–10. [Google Scholar] [CrossRef]
- Urmann, D.; Lorenz, C.; Linker, S.M.; Braun, M.; Wachtveitl, J.; Bamann, C. Photochemical Properties of the Red-shifted Channelrhodopsin Chrimson. Photochem. Photobiol. 2017, 93, 782–795. [Google Scholar] [CrossRef] [PubMed]
- Gradinaru, V.; Thompson, K.R.; Deisseroth, K. eNpHR: A Natronomonas halorhodopsin enhanced for optogenetic applications. Brain Cell Biol. 2008, 36, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Chuong, A.S.; Miri, M.L.; Busskamp, V.; Matthews, G.A.C.; Acker, L.C.; Sorensen, A.; Young, A.; Klapoetke, N.; Henninger, M.A.; Kodandaramaiah, S.; et al. Noninvasive optical inhibition with a red-shifted microbial rhodopsin. Nat. Neurosci. 2014, 17, 1123–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vierock, J.; Rodriguez-Rozada, S.; Dieter, A.; Pieper, F.; Sims, R.; Tenedini, F.; Bergs, A.C.F.; Bendifallah, I.; Zhou, F.; Zeitzschel, N.; et al. BiPOLES is an optogenetic tool developed for bidirectional dual-color control of neurons. Nat. Commun. 2021, 12, 1–20. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 488 nm | 589 nm | ||
|---|---|---|---|
| Before optimization | x | ||
| y | |||
| After optimization | x | ||
| y | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmieder, F.; Büttner, L.; Hanitzsch, T.; Busskamp, V.; Czarske, J.W. Two-Wavelength Computational Holography for Aberration-Corrected Simultaneous Optogenetic Stimulation and Inhibition of In Vitro Biological Samples. Appl. Sci. 2022, 12, 2283. https://0-doi-org.brum.beds.ac.uk/10.3390/app12052283
Schmieder F, Büttner L, Hanitzsch T, Busskamp V, Czarske JW. Two-Wavelength Computational Holography for Aberration-Corrected Simultaneous Optogenetic Stimulation and Inhibition of In Vitro Biological Samples. Applied Sciences. 2022; 12(5):2283. https://0-doi-org.brum.beds.ac.uk/10.3390/app12052283
Chicago/Turabian StyleSchmieder, Felix, Lars Büttner, Tony Hanitzsch, Volker Busskamp, and Jürgen W. Czarske. 2022. "Two-Wavelength Computational Holography for Aberration-Corrected Simultaneous Optogenetic Stimulation and Inhibition of In Vitro Biological Samples" Applied Sciences 12, no. 5: 2283. https://0-doi-org.brum.beds.ac.uk/10.3390/app12052283