
Novel Insights into the Immunomodulatory Effects of Caryophyllane Sesquiterpenes: A Systematic Review of Preclinical Studies


Abstract
:1. Introduction
2. Methodology
3. Results
3.1. Ability of Caryophyllane Sesquiterpenes to Modulate Macrophage Function

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Caryophyllane Sesquiterpene | Immune System Cells/Cell Model | Stimulation | Concentration of Test Compound [µM] | Outcome | References |
|---|---|---|---|---|---|
| β-Caryophyllene | Macrophages/ RAW 264.7 | - | 10, 50 | Lacking effects on cell viability | [30] |
| - | 50 | ↑ Ras, Akt, p21, β-catenin, caspase-3 ↓ MAPK | |||
| - | 10 | ↓ TNF-α and PGE2 Lacking effect on IL-6 | |||
| LPS a [100 ng/mL] | 10 | ↓ TNF-α and IL-6 Lacking effect on PGE2 | |||
| - | 5 | Lacking effect on IL-1β, IL-6, TNF-α | [31] | ||
| LPS a [100 ng/mL] | 5 | ↓ IL-1β, IL-6, TNF-α | |||
| Macrophages/primary splenocytes from BALB/c mice | LPS b [2.5 µg/mL] | 0.25–25 | ↓ IL-1, IL-10 ↑ Th2/Th1 ratio | [33] | |
| Macrophages/ M2-MΦs (co-cultured) | - | 5 | ↓ MCP-1, M-CSF and migration | [35] | |
| LPS c [1 µg/mL] | 0.2–50 | Cytoprotection | [40] | ||
| Microglia/C57BL/6 mice cells | LPS c [1 µg/mL] | 0.2–25 | ↓ TNF-α, IL-β, PGE2, NO ad iNOS ↑ IL-10, Arg-1, urea and GSH | ||
| Microglia/ BV2 cells | Aβ1–42 [20 µM] | 10, 25, 50 | ↓ IL-β, IL-6, TNF-α, NO, PGE2, TLR4, LDH, Nf-kB iNOS and COX-2 inhibition | [42] | |
| Monocytes/ THP-1 | TNF-α d | 5 | ↓ VCAM-1, adhesion on HUVECs | [36] | |
| Monocytes/primary monocytes CD14+ | LPS a [313 ng/mL] | 0.5 | ↓TNF-α and IL-1β Lacking effects on IL-6 and IL-10 levels ↓ Erk 1/2 and JNK 1/2 activation Lacking effects on MAPK p38 activation | [43] | |
| Lymphocytes/ isolated cells from EAE mice | MOG35–55 [30 µg/mL] | 0.2, 1 | ↓cellular proliferation ↓ IFNγ, IL-17, IFNγ/IL-4 ratio, IL17/IL10 ratio ↑ IL-4, IL-10 | [44] | |
| MOG35–55 [10 µg/mL] | 1–100 | ↑ IL-10, ↓ IFNγ, Lacking effects on IL-4 | [45] | ||
| Lymphocytes/ isolated cells from healthy donors | - | 0.5–500 | Genoprotective effects | [46] | |
| β-Caryophyllene oxide | Macrophages/ primary splenocytes from BALB/c mice | LPS b [2.5 µg/mL] | 0.25–25 | ↓ IL-2 | [33] |
| Lymphocytes/ isolated from healthy donors | - | 1–100 | Lacking genotoxic effects | [47] | |
| - | 250 and 500 | Cytotoxic effects | |||
| α-Humulene | Macrophages/ RAW 264.7 | LPS a [1 µg/mL] | 755, 1555 | ↓ NO | [32] |
| Macrophages/primary splenocytes from BALB/c mice | LPS b [2,5 µg/mL] | 0.25–25 | ↓ IL-10 | [33] | |
| Lymphocytes/MLNs | OVA [50 µg/mL] | 1, 10 | ↓ IL-5 | [48] |
| Caryophyllane Sesquiterpene | Animal Model/ Immune Cells | Test Compound Dose (Administration Route) | Outcome | References |
|---|---|---|---|---|
| β-Caryophyllene | High-fat diet and B16F10 bearing C57BL/6 mice/macrophages | 0.15–0.3% of diet (per os) | ↓ macrophages in tumor tissue ↓ macrophages in the lymphnode | [35] |
| LPS-fat-induced atherosclerosis C57BL/6 mice/macrophages | 10 mg/kg (i.p.) | ↓ macrophage infiltration to the aortic surface | [36] | |
| Alcoholic steatohepatitis C57BL/6J mice/macrophages | 10 mg/kg (i.p.) | ↓ CXCL2, TNF-α | [51] | |
| Cisplatin-induced nephrotoxicity C57BL/6J mice/macrophages | 10 mg/kg (i.p.) | ↓ macrophage infiltration | [37] | |
| DSS-induced colitis CD1 mice/macrophages | 12–50 mg/kg (per os; gavage) | ↓ infiltration in colon | [38] | |
| EAE C57BL/6 mice/microglia | 5 mg/kg (per os) | ↓ TNF-α, PGE2, iNOS, NO, iNOS/Arg1, NO/urea ↑ IL-10, Arg-1, urea | [44] | |
| EAE C57BL/6 mice/microglia | 50 mg/kg (per os; gavage) | ↓ Activation microglia | [45] | |
| Rotenone-induced Parkinson’s disease in Wistar rats/microglia | 50 mg/kg (i.p.) | ↓ Activation microglia ↓ Iba-1 | [52] | |
| EAE C57BL/6 mice/leucocytes | 5 mg/kg (per os) | ↓ leucocytes in spinal cord | [44] | |
| Cisplatin-induced nephrotoxicity C57BL/6J mice/leucocytes | 10 mg/kg (i.p.) | ↓ leucocytes infiltration | [37] | |
| High-fat diet and B16F10-bearing C57BL/6 mice/leucocytes | 0.15–0.3% of diet (per os) | ↓ lymphocytes in tumor tissue | [35] | |
| EAE C57BL/6 mice/lymphocytes | 5 mg/kg (per os) | ↓ cell proliferation ↑ IL-4, IL-10, TGF-β, GATA-3 ↓ TNF-α, IL-6, IFNγ, IL-17, T-bet, T-bet/GATA3 (Th1/Th2) | [44] | |
| EAE C57BL/6 mice/lymphocytes | 50 mg/kg (per os; gavage) | ↓ CD4+ and CD8+ T cells ↓ CD4+ and CD8+ T cells in peripheral lymphoid tissue Inhibition of Th1 myelin-specific cells in CNS | [45] | |
| OVA-induced airway allergic inflammation BALB/c mice/lymphocytes | 50 mg/kg (per os) | Lacking effects on lymphocytes infiltration | [48] | |
| Alcoholic steatohepatitis C57BL/6J mice/neutrophils | 10 mg/kg (i.p.) | ↓ ICAM-1, E-selectin, P-selectin | [51] | |
| Ehrlich ascites BALB/c mice/natural killer cells | 20 mg/kg (i.p.) | Restoration of NK cell activity (less potent than α-HUM) | [52] | |
| DSS-induced colitis CD1 mice/neutrophils | 12–50 mg/kg (per os; gavage) | ↓ infiltration in colon | [38] | |
| LPS-induced paw inflammation in Wistar rats/neutrophils | 50 mg/kg (per os) | ↓ infiltration of neutrophiles | [53] | |
| OVA-induced airway allergic inflammation BALB/c mice/eosinophils | 50 mg/kg (per os; gavage) | Lacking effects on number of eosinophils in BALF and on eosinophil peroxidase activity | [48] | |
| α-Humulene | OVA-induced airway allergic inflammation BALB/c mice/leucocytes | 1 mg/mL (aerosol) | ↓ leucocyte counts in BALF | [48] |
| LPS-induced paw inflammation in Wistar rats/neutrophils | 50 mg/kg (per os) | ↓ neutrophils infiltration | [53] | |
| OVA-induced airway allergic inflammation BALB/c mice/eosinophils | 50 mg/kg (gavage) | ↓ eosinophils infiltration ↓ eosinophils, ↓ peroxidase activity, ↓ CCL11, ↓ IL-5 in BALF | [48] | |
| 1 mg/mL (aerosol) | ↓ eosinophils, ↓ peroxidase activity, ↓ CCL11, ↓ IL-5 and ↑ IFNγ in BALF |
3.2. Effects of Caryophyllane Sesquiterpenes on Other Monocytic Cells
3.3. Modulation of Lymphocyte Function
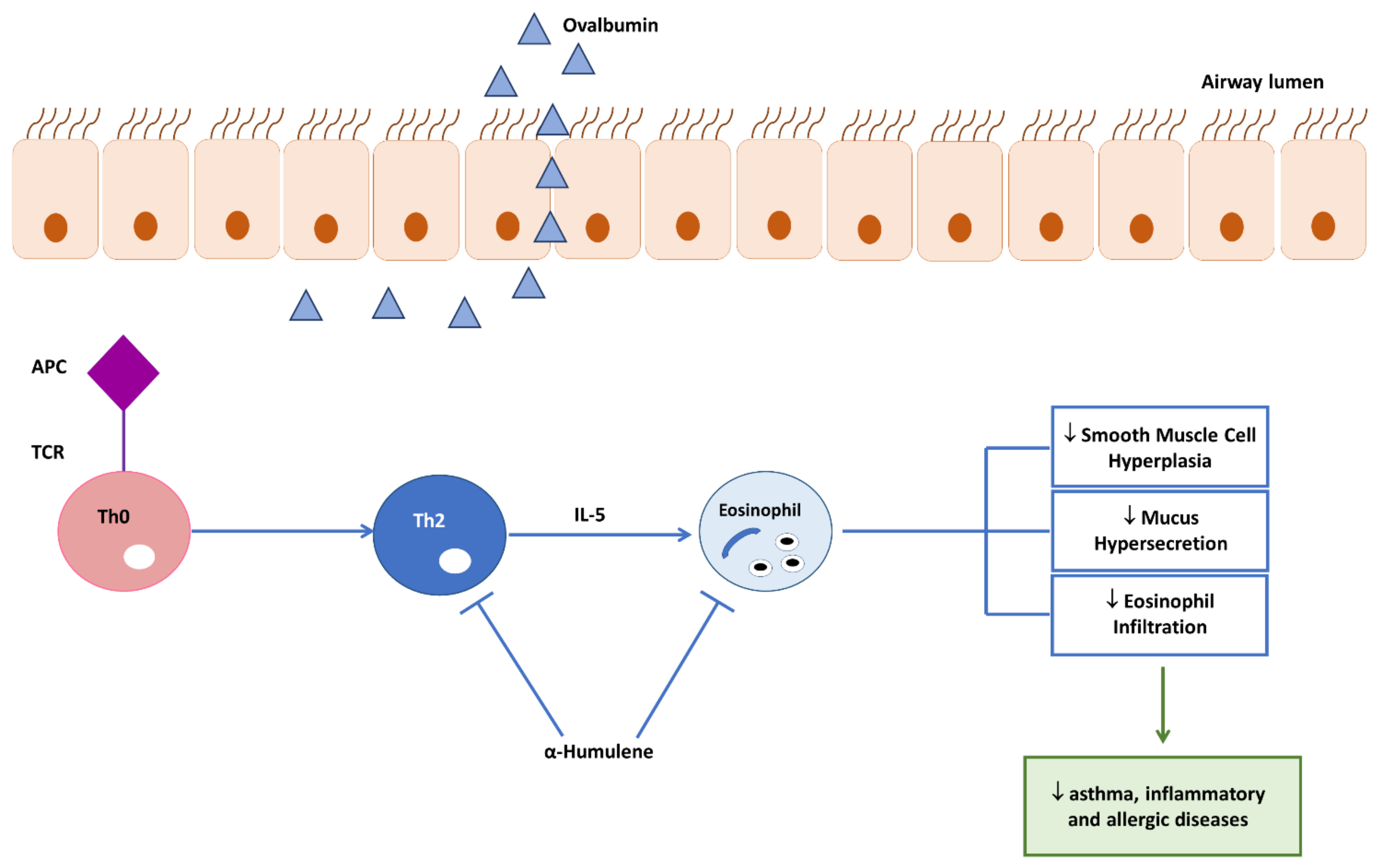
3.4. Effects of Caryophyllane Sesquiterpenes on Neutrophiles and Eosinophils
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Sotto, A.; Mancinelli, R.; Gullì, M.; Eufemi, M.; Mammola, C.L.; Mazzanti, G.; Di Giacomo, S. Chemopreventive potential of caryophyllane sesquiterpenes: An overview of preliminary evidence. Cancers 2020, 12, 3034. [Google Scholar] [CrossRef] [PubMed]
- Di Giacomo, S.; Mariano, A.; Gullì, M.; Fraschetti, C.; Vitalone, A.; Filippi, A.; Mannina, L.; Scotto d’Abusco, A.; Di Sotto, A. Role of caryophyllane sesquiterpenes in the entourage Effect of Felina 32 Hemp inflorescence Phytocomplex in triple negative MDA-MB-468 breast cancer cells. Molecules 2021, 26, 6688. [Google Scholar] [CrossRef]
- Garzoli, S.; Pirolli, A.; Vavala, E.; Di Sotto, A.; Sartorelli, G.; Božović, M.; Angiolella, L.; Mazzanti, G.; Pepi, F.; Ragno, R. Multidisciplinary approach to determine the optimal time and period for extracting the essential oil from Mentha suaveolens Ehrh. Molecules 2015, 20, 9640–9655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schepetkin, I.A.; Özek, G.; Özek, T.; Kirpotina, L.N.; Khlebnikov, A.I.; Quinn, M.T. Chemical composition and immunomodulatory activity of Hypericum perforatum essential oils. Biomolecules 2020, 10, 916. [Google Scholar] [CrossRef] [PubMed]
- Ingallina, C.; Sobolev, A.P.; Circi, S.; Spano, M.; Fraschetti, C.; Filippi, A.; Di Sotto, A.; Di Giacomo, S.; Mazzoccanti, G.; Gasparrini, F.; et al. Cannabis sativa L. Inflorescences from Monoecious Cultivars Grown in Central Italy: An Untargeted Chemical Characterization from Early Flowering to Ripening. Molecules 2020, 25, 1908. [Google Scholar] [CrossRef]
- Mariano, A.; Di Sotto, A.; Leopizzi, M.; Garzoli, S.; Di Maio, V.; Gullì, M.; Dalla Vedova, P.; Ammendola, S.; Scotto d’Abusco, A. Antiarthritic effects of a root extract from Harpagophytum procumbens DC: Novel insights into the molecular mechanisms and possible bioactive phytochemicals. Nutrients 2020, 12, 2545. [Google Scholar] [CrossRef]
- Di Sotto, A.; Vitalone, A.; Di Giacomo, S. Plant-Derived Nutraceuticals and Immune System Modulation: An Evidence-Based Overview. Vaccines 2020, 8, 468. [Google Scholar] [CrossRef]
- Bain, B.J. Structure and function of red and white blood cells and platelets. Medicine 2021, 49, 183–188. [Google Scholar] [CrossRef]
- Kumar, D.; Romero, Y.; Schuck, K.N.; Smalley, H.; Subedi, B.; Fleming, S.D. Drivers and regulators of humoral innate immune responses to infection and cancer. Mol. Immunol. 2020, 121, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Day, C.L.; Kaufmann, D.E.; Kiepiela, P.; Brown, J.A.; Moodley, E.S.; Reddy, S.; Mackey, E.W.; Miller, J.D.; Leslie, A.J.; DePierres, C.; et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature 2006, 443, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Finlay, C.M.; Walsh, K.P.; Mills, K.H. Induction of regulatory cells by helminth parasites: Exploitation for the treatment of inflammatory diseases. Immunol. Rev. 2014, 259, 206–230. [Google Scholar] [CrossRef] [PubMed]
- Yanaba, K.; Bouaziz, J.D.; Matsushita, T.; Magro, C.M.; St Clair, E.W.; Tedder, T.F. B-lymphocyte contributions to human autoimmune disease. Immunol. Rev. 2008, 223, 284–299. [Google Scholar] [CrossRef]
- Havnaer, A.; Han, G. Autoinflammatory disorders: A review and update on pathogenesis and treatment. Am. J. Clin. Dermatol. 2019, 20, 539–564. [Google Scholar] [CrossRef] [PubMed]
- Stegenga, M.E.; van der Crabben, S.N.; Blümer, R.M.E.; Levi, M.; Meijers, J.C.; Serlie, M.J.; Tanck, M.W.; Sauerwein, H.P.; van der Poll, T. Hyperglycemia enhances coagulation and reduces neutrophil degranulation, whereas hyperinsulinemia inhibits fibrinolysis during human endotoxemia. Blood 2008, 112, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Chao, W.C.; Yen, C.L.; Wu, Y.H.; Chen, S.Y.; Hsieh, C.Y.; Chang, T.C.; Ou, H.Y.; Shieh, C.C. Increased resistin may suppress reactive oxygen species production and inflammasome activation in type 2 diabetic patients with pulmonary tuberculosis infection. Microbes Infect. 2015, 17, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.B.; Lad, A.; Prasad, A.S.B.; Balakrishnan, A.; Ramachandra, L.; Satyamoorthy, K. High glucose modulates IL-6 mediated immune homeostasis through impeding neutrophil extracellular trap formation. FEBS Lett. 2013, 587, 2241–2246. [Google Scholar] [CrossRef] [Green Version]
- Berbudi, A.; Surendar, J.; Ajendra, J.; Gondorf, F.; Schmidt, D.; Neumann, A.L.; Wardani, A.P.; Layland, L.E.; Hoffmann, L.S.; Pfeifer, A.; et al. Filarial infection or antigen administration improves glucose tolerance in diet-induced obese mice. J. Innate Immun. 2016, 8, 601–616. [Google Scholar] [CrossRef] [PubMed]
- Chawla, A.; Nguyen, K.D.; Goh, Y.P.S. Macrophage-mediated inflammation in metabolic disease. Nat. Rev. Immunol. 2011, 11, 738–749. [Google Scholar] [CrossRef] [Green Version]
- Restrepo, B.I.; Twahirwa, M.; Rahbar, M.H.; Schlesinger, L.S. Phagocytosis via complement or Fc-gamma receptors is compromised in monocytes from type 2 diabetes patients with chronic hyperglycemia. PLoS ONE 2014, 9, e92977. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, K.; Fornaguera-Trias, J.; Sheridan, J.F. Stress-induced microglia activation and monocyte trafficking to the brain underlie the development of anxiety and depression. Curr. Top. Behav. Neurosci. 2017, 31, 155–172. [Google Scholar] [PubMed]
- Tay, T.L.; Savage, J.C.; Hui, C.W.; Bisht, K.; Tremblay, M.E. Microglia across the lifespan: From origin to function in brain development, plasticity and cognition. J. Physiol. 2017, 595, 1929–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.C.; Chang, L.Y.; Huang, C.T.; Peng, H.M.; Dutta, A.; Chen, T.C.; Yeh, C.T.; Lin, C.Y. Effector/memory but not naive regulatory T cells are responsible for the loss of concomitant tumor immunity. J. Immunol. 2009, 182, 6095–6104. [Google Scholar] [CrossRef]
- Galluzzi, L.; Chan, T.A.; Kroemer, G.; Wolchok, J.D.; López-Soto, A. The hallmarks of successful anticancer immunotherapy. Sci. Transl. Med. 2018, 10, eaat7807. [Google Scholar] [CrossRef]
- Taefehshokr, S.; Parhizkar, A.; Hayati, S.; Mousapour, M.; Mahmoudpour, A.; Eleid, L.; Rahmanpour, D.; Fattahi, S.; Shabani, H.; Taefehshokr, N. Cancer immunotherapy: Challenges and limitations. Pathol. Res. Pract. 2021, 229, 153723. [Google Scholar] [CrossRef] [PubMed]
- Welch, V.; Petticrew, M.; Petkovic, J.; Moher, D.; Waters, E.; White, H.; Tugwell, P. Extending the PRISMA statement to equity-focused systematic reviews (PRISMA-E 2012): Explanation and elaboration. Int. J. Equity Health 2015, 14, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, O.; Donnelly, C.R.; Ji, R.R. Regulation of pain by neuro-immune interactions between macrophages and nociceptor sensory neurons. Curr. Opin. Neurobiol. 2020, 62, 17–25. [Google Scholar] [CrossRef]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef]
- Domoto, R.; Sekiguchi, F.; Tsubota, M.; Kawabata, A. Macrophage as a peripheral pain regulator. Cells 2021, 10, 1881. [Google Scholar] [CrossRef]
- Li, C.; Xu, M.M.; Wang, K.; Adler, A.J.; Vella, A.T.; Zhou, B. Macrophage polarization and meta- inflammation. Transl. Res. 2018, 191, 29–44. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Levy, R.M. Metaxalone suppresses production of inflammatory cytokines associated with painful conditions in mouse macrophages RAW264.7 cells in Vitro: Synergistic effect with β-caryophyllene. Curr. Mol. Med. 2020, 20, 643–652. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Levy, R.M. The combination of catechin, baicalin and β-caryophyllene potentially suppresses the production of inflammatory cytokines in mouse macrophages in vitro. Exp. Ther. Med. 2019, 17, 4312–4318. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, S.M.M.; Costa, C.R.R.; Gelfuso, G.M.; Guerra, E.N.S.; de Medeiros Nóbrega, Y.K.; Gomes, S.M.; Pic-Taylor, A.; Fonseca-Bazzo, Y.M.; Silveira, D.; Magalhães, P.O. Wound healing effect of essential oil extracted from Eugenia dysenterica DC (Myrtaceae) Leaves. Molecules 2018, 24, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ku, C.M.; Lin, J.Y. Anti-inflammatory effects of 27 selected terpenoid compounds tested through modulating Th1/Th2 cytokine secretion profiles using murine primary splenocytes. Food Chem. 2013, 141, 1104–1113. [Google Scholar] [CrossRef]
- Guilliams, M.; Mildner, A.; Yona, S. Developmental and functional heterogeneity of monocytes. Immunity 2018, 49, 595–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.I.; Kim, E.J.; Kwon, G.T.; Jung, Y.J.; Park, T.; Kim, Y.; Park, J.H.Y. β-Caryophyllene potently inhibits solid tumor growth and lymph node metastasis of B16F10 melanoma cells in high-fat diet–induced obese C57BL/6N mice. Carcinogenesis 2015, 36, 1028–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Yang, C.; Dai, X.; Ao, Y.; Li, Y. Inhibitory effect of trans-caryophyllene (TC) on leukocyte- endothelial attachment. Toxicol. Appl. Pharmacol. 2017, 329, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Horváth, B.; Mukhopadhyay, P.; Kechrid, M.; Patel, V.; Tanchian, G.; Wink, D.A.; Pacher, P. β-Caryophyllene ameliorates cisplatin-induced nephrotoxicity in a cannabinoid 2 receptor-dependent manner. Free Radic. Biol. Med. 2012, 52, 1325–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bento, A.F.; Marcon, R.; Dutra, R.C.; Claudino, R.F.; Cola, M.; Leite, D.F.P.; Calixto, J.B. β- Caryophyllene inhibits dextran sulfate sodium-induced colitis in mice through CB2 receptor activation and PPARγ pathway. Am. J. Clin. Pathol. 2011, 178, 1153–1166. [Google Scholar] [CrossRef]
- Ojha, S.; Javed, H.; Azimullah, S.; Haque, M.E. β-Caryophyllene, a phytocannabinoid attenuates oxidative stress, neuroinflammation, glial activation, and salvages dopaminergic neurons in a rat model of Parkinson disease. Mol. Cell. Biochem. 2016, 418, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Askari, V.R.; Shafiee-Nick, R. The protective effects of β-caryophyllene on LPS-induced primary microglia M1/M2 imbalance: Mechanistic evaluation. Life Sci. 2019, 219, 40–73. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Liu, H.; Xiang, F.; Xu, L.; Dong, Z. β-Caryophyllene protects against ischemic stroke by promoting polarization of microglia toward M2 phenotype via the TLR4 pathway. Life Sci. 2019, 237, 116915. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zeng, Z.; Wang, B.; Guo, S. Trans-caryophyllene inhibits amyloid β (Aβ) oligomer-induced neuroinflammation in BV-2 microglial cells. Int. Immunopharmacol. 2017, 51, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Gertsch, J.; Leonti, M.; Raduner, S.; Racz, I.; Chen, J.Z.; Xie, X.Q.; Zimmer, A. Beta-caryophyllene is a dietary cannabinoid. Proc. Natl. Acad. Sci. USA 2008, 105, 9099–9104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askari, V.R.; Rahimi, V.B.; Tabatabaee, S.A.; Shafiee-Nick, R. Combination of Imipramine, a sphingomyelinase inhibitor, and β-caryophyllene improve their therapeutic effects on experimental autoimmune encephalomyelitis (EAE). Int. Immunopharmacol. 2019, 77, 105923. [Google Scholar] [CrossRef] [PubMed]
- Alberti, T.B.; Barbosa, W.L.R.; Vieira, J.L.F.; Raposo, N.R.B.; Dutra, R.C. (−)-β-Caryophyllene, a CB2 receptor-selective phytocannabinoid, suppresses motor paralysis and neuroinflammation in a murine model of multiple sclerosis. Int. J. Mol. Sci. 2017, 18, 691. [Google Scholar] [CrossRef] [PubMed]
- Di Sotto, A.; Mazzanti, G.; Carbone, F.; Hrelia, P.; Maffei, F. Inhibition by β-caryophyllene of ethyl methanesulfonate-induced clastogenicity in cultured human lymphocytes. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2010, 699, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Di Sotto, A.; Maffei, F.; Hrelia, P.; Castelli, F.; Sarpietro, M.G.; Mazzanti, G. Genotoxicity assessment of β-caryophyllene oxide. Regul. Toxicol. Pharmacol. 2013, 66, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Rogerio, A.P.; Andrade, E.L.; Leite, D.F.; Figueiredo, C.P.; Calixto, J.B. Preventive and therapeutic anti-inflammatory properties of the sesquiterpene α-humulene in experimental airways allergic inflammation. Br. J. Pharmacol. 2009, 158, 1074–1087. [Google Scholar] [CrossRef] [Green Version]
- Hait, N.C.; Maiti, A. The role of sphingosine-1-phosphate and ceramide-1-phosphate in inflammation and cancer. Mediat. Inflamm. 2017, 2017, 4806541. [Google Scholar] [CrossRef] [PubMed]
- Dhopeshwarkar, A.; Mackie, K. CB2 Cannabinoid receptors as a therapeutic target-what does the future hold? Mol. Pharmacol. 2014, 86, 430–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varga, Z.V.; Matyas, C.; Erdelyi, K.; Cinar, R.; Nieri, D.; Chicca, A.; Pacher, P. β-Caryophyllene protects against alcoholic steatohepatitis by attenuating inflammation and metabolic dysregulation in mice. Br. J. Pharmacol. 2018, 175, 320–334. [Google Scholar] [CrossRef] [PubMed]
- Javed, H.; Azimullah, S.; Haque, M.E.; Ojha, S.K. Cannabinoid type 2 (CB2) receptors activation protects against oxidative stress and neuroinflammation associated dopaminergic neurodegeneration in rotenone model of Parkinson’s disease. Front. Neurosci. 2016, 10, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medeiros, R.; Passos, G.F.; Vitor, C.E.; Koepp, J.; Mazzuco, T.L.; Pianowski, L.F.; Calixto, J.B. Effect of two active compounds obtained from the essential oil of Cordia verbenacea on the acute inflammatory responses elicited by LPS in the rat paw. Br. J. Pharmacol. 2007, 151, 618–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, G.; Tacke, R.; Hedrick, C.C.; Hanna, R.N. Nonclassical patrolling monocyte function in the vasculature. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1306–1316. [Google Scholar] [CrossRef] [Green Version]
- Larosa, D.F.; Orange, J.S. 1. Lymphocytes. J. Allergy Clin. Immunol. 2008, 121, S364–S412. [Google Scholar] [CrossRef]
- Cosmi, L.; Maggi, L.; Santarlasci, V.; Liotta, F.; Annunziato, F. T helper cells plasticity in inflammation. Cytom. A 2014, 85, 36–42. [Google Scholar] [CrossRef]
- Da Silva, S.L.; Figueiredo, P.M.; Yano, T. Chemotherapeutic potential of the volatile oils from Zanthoxylum rhoifolium Lam leaves. Eur. J. Pharmacol. 2007, 576, 180–188. [Google Scholar] [CrossRef]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Hart, O.M.; Athie-Morales, V.; O’Connor, G.M.; Gardiner, C.M. TLR7/8-mediated activation of human NK cells results in accessory cell-dependent IFN-γ production. J. Immunol. 2005, 175, 1636–1642. [Google Scholar] [CrossRef]
- Long, E.O. Ready for prime time: NK cell priming by dendritic cells. Immunity 2007, 26, 385–387. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S. Elie Metchnikoff: Father of natural immunity. Eur. J. Immunol. 2008, 38, 3257–3264. [Google Scholar] [CrossRef]
- Kaufmann, S.H. Immunology’s foundation: The 100-year anniversary of the Nobel Prize to Paul Ehrlich and Elie Metchnikoff. Nat. Immunol. 2008, 9, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Borregaard, N.; Sørensen, O.E.; Theilgaard-Mönch, K. Neutrophil granules: A library of innate immunity proteins. Trends Immunol. 2007, 28, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Megiovanni, A.M.; Sanchez, F.; Robledo-Sarmiento, M.; Morel, C.; Gluckman, J.C.; Boudaly, S. Polymorphonuclear neutrophils deliver activation signals and antigenic molecules to dendritic cells: A new link between leukocytes upstream of T lymphocytes. J. Leukoc. Biol. 2006, 79, 977–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, B.; Ahmad, M.F.; Nagy, L.E.; Tsukamoto, H. Inflammatory pathways in alcoholic steatohepatitis. J. Hepatol. 2019, 70, 249–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izzo, A.A.; Teixeira, M.; Alexander, S.P.; Cirino, G.; Docherty, J.R.; George, C.H.; Insel, P.A.; Ji, Y.; Kendall, D.A.; Panattieri, R.A. A practical guide for transparent reporting of research on natural products in British Journal of Pharmacology: Reproducibility of natural product research. Br. J. Pharmacol. 2020, 177, 2169–2178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Sotto, A.; Paolicelli, P.; Nardoni, M.; Abete, L.; Božović, M.; Di Giacomo, S.; Mazzanti, G.; Casadei, M.A.; Petralito, S. SPC Liposomes as possible delivery systems for improving bioavailability of the natural sesquiterpene β-caryophyllene: Lamellarity and drug-loading as key features for a rational drug delivery design. Pharmaceutics 2018, 10, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, J.; Teng, Z.; Zhang, L.; Yang, J.; Ma, L.; Wang, F.; Tian, X.; An, R.; Yang, M.; Zhang, Q. β-caryophyllene/hydroxypropyl-β-cyclodextrin inclusion complex improves cognitive deficits in rats with vascular dementia through the cannabinoid receptor type 2 -mediated pathway. Front. Pharmacol. 2017, 8, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neves, J.K.D.O.; Apolinário, A.C.; Saraiva, K.L.A.; Da Silva, D.T.C.; Reis, M.Y.D.F.A.; Damasceno, B.P.G.L.; Pessoa, A., Jr.; Galvão, M.A.M.; Soares, L.A.L.; Júnior, V.F.D.V. Microemulsions containing Copaifera multijuga Hayne oil-resin: Challenges to achieve an efficient system for β-caryophyllene delivery. Ind. Crops Prod. 2018, 111, 185–192. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gullì, M.; Percaccio, E.; Di Giacomo, S.; Di Sotto, A. Novel Insights into the Immunomodulatory Effects of Caryophyllane Sesquiterpenes: A Systematic Review of Preclinical Studies. Appl. Sci. 2022, 12, 2292. https://0-doi-org.brum.beds.ac.uk/10.3390/app12052292
Gullì M, Percaccio E, Di Giacomo S, Di Sotto A. Novel Insights into the Immunomodulatory Effects of Caryophyllane Sesquiterpenes: A Systematic Review of Preclinical Studies. Applied Sciences. 2022; 12(5):2292. https://0-doi-org.brum.beds.ac.uk/10.3390/app12052292
Chicago/Turabian StyleGullì, Marco, Ester Percaccio, Silvia Di Giacomo, and Antonella Di Sotto. 2022. "Novel Insights into the Immunomodulatory Effects of Caryophyllane Sesquiterpenes: A Systematic Review of Preclinical Studies" Applied Sciences 12, no. 5: 2292. https://0-doi-org.brum.beds.ac.uk/10.3390/app12052292