
Nuclear Magnetic Resonance Spectroscopy Analysis of Anaerobic Microbial Metabolic Response to Benzalkonium Chloride Disinfectant


Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Microcosm Preparation
2.2. NMR Analysis
2.3. Gas Chromatography Analysis
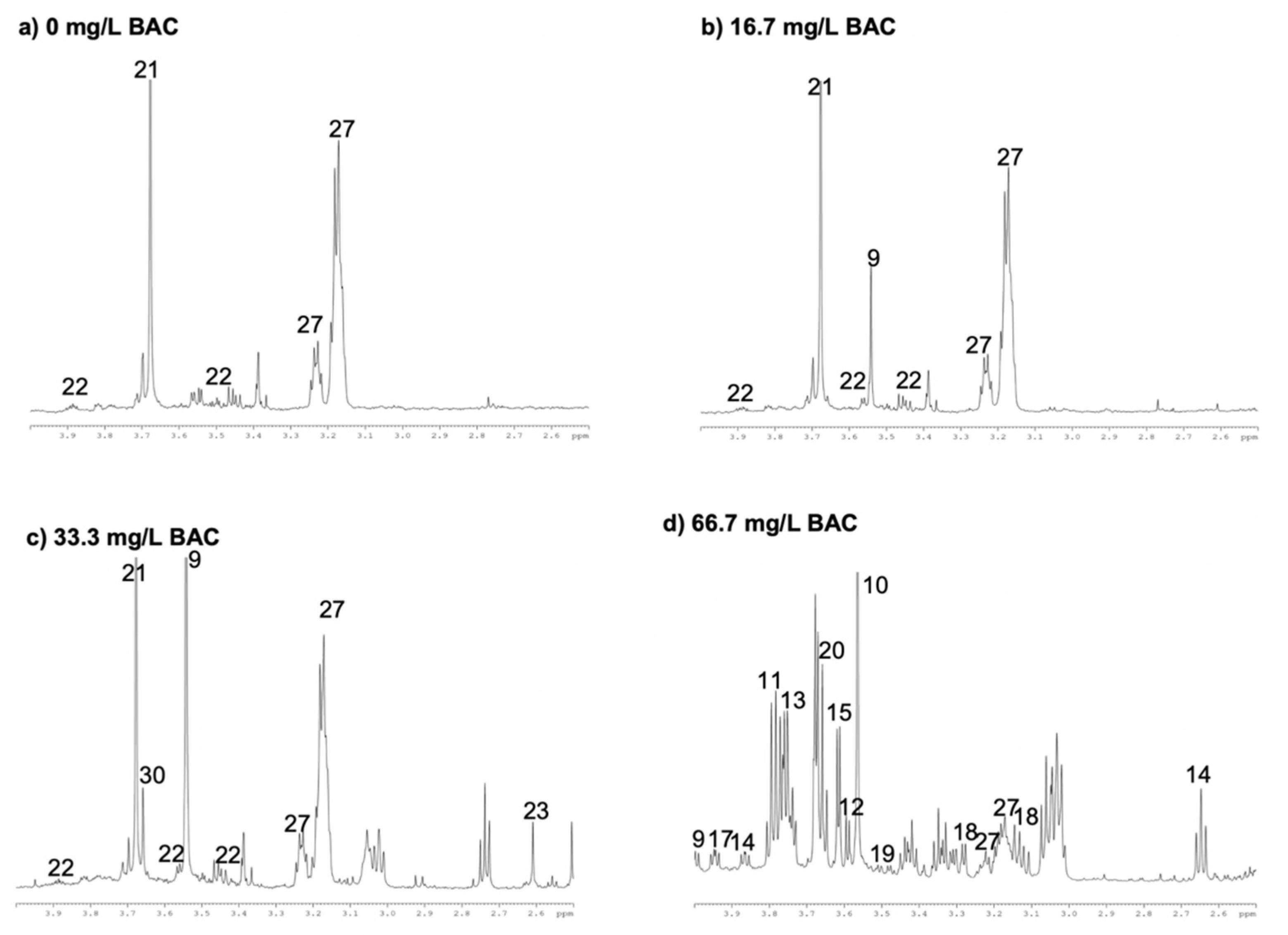
3. Results and Discussion
3.1. Identification of BAC in Wastewater Effluent
3.2. Metabolite Characterization
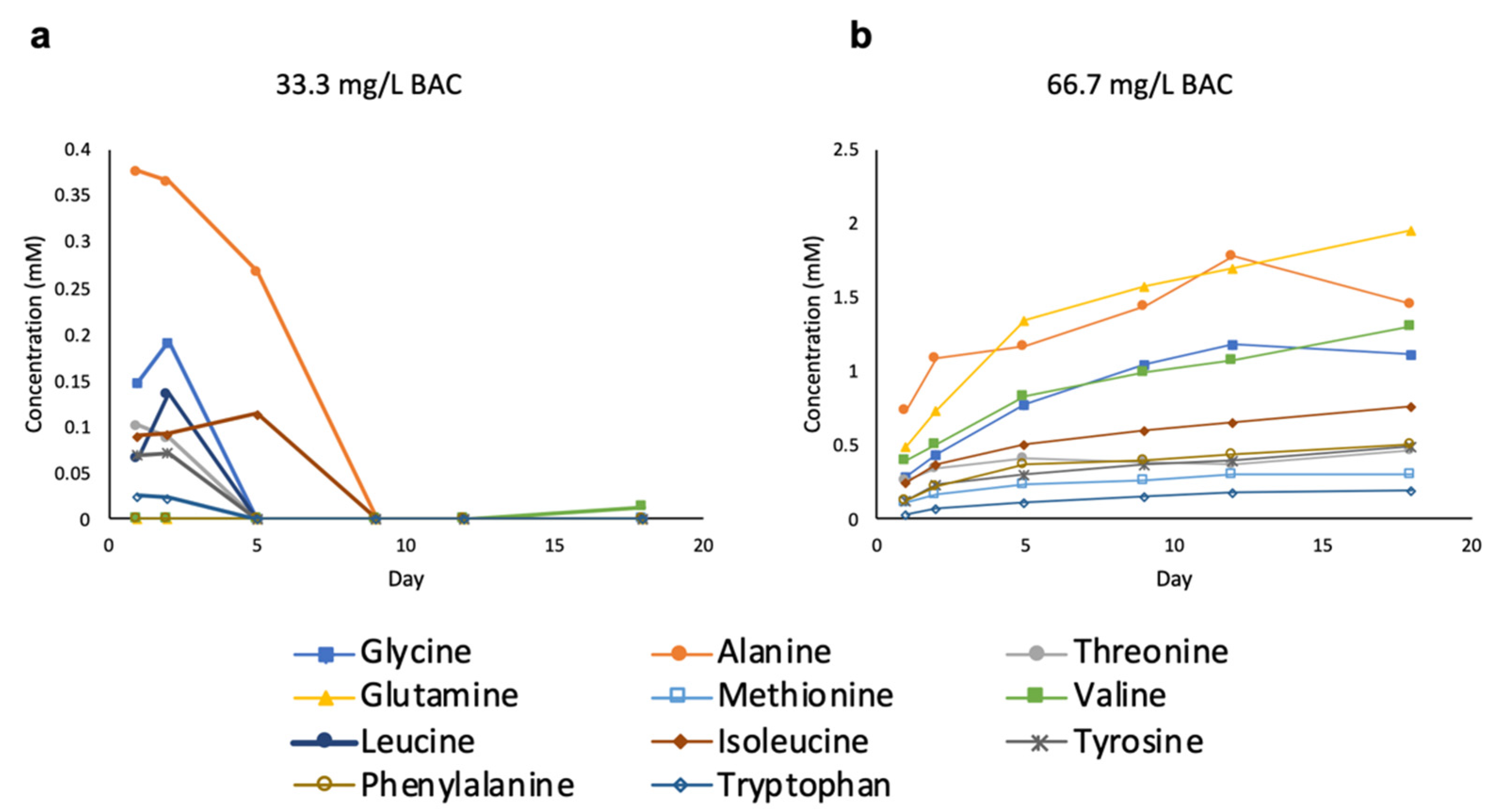
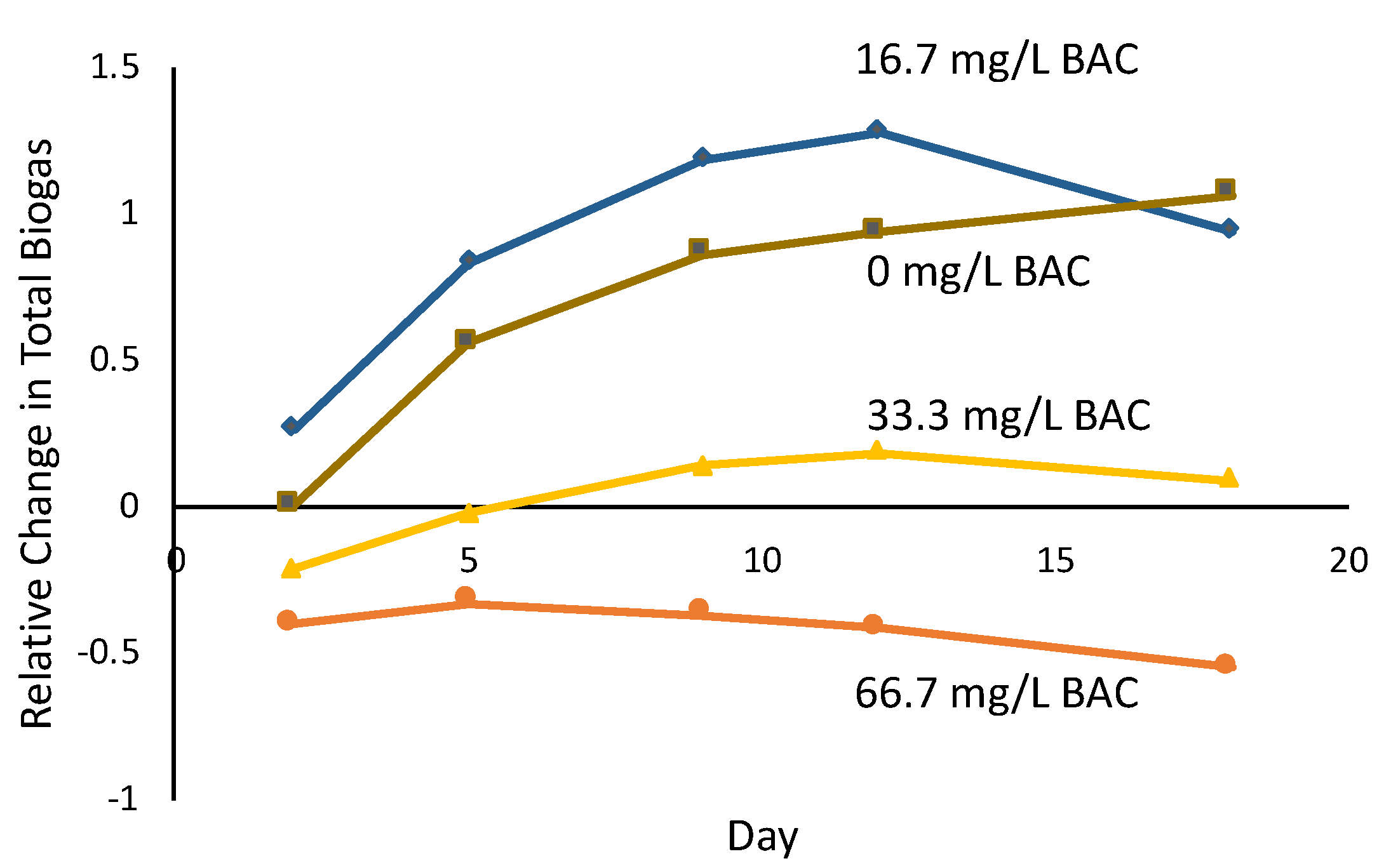
3.3. Evolution of Microbial Metabolites after Initial BAC Exposure
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oliveira, F.; Doelle, K. Anaerobic digestion of food waste to produce biogas: A comparison of bioreactors to increase methane content—A review. J. Food Process Technol. 2015, 6, 2. [Google Scholar]
- Mir, M.A.; Hussain, A.; Verma, C. Design considerations and operational performance of anaerobic digestor: A review. Cogent Eng. 2016, 3, 1181696. [Google Scholar] [CrossRef]
- Paritosh, K.; Kushwaha, S.K.; Yadav, M.; Pareek, N.; Chawade, A.; Vivekanand, V. Food Waste to Energy: An Overview of Sustainable Approaches for Food Waste Management and Nutrient Recycling. BioMed Res. Int. 2017, 2017, 2370927. [Google Scholar] [CrossRef] [PubMed]
- Franke-Whittle, I.H.; Walter, A.; Ebner, C.; Insam, H. Investigation into the effect of high concentrations of volatile fatty acids in anaerobic digestion on methanogenic communities. Waste Manag. 2014, 34, 2080–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, A.J.; Hobbs, P.J.; Holliman, P.J.; Jones, D.L. Optimisation of the anaerobic digestion of agricultural resources. Bioresour. Technol. 2008, 99, 7928–7940. [Google Scholar] [CrossRef]
- Lourenço, N.D.; Lopes, J.; Almeida, C.F.; Sarraguça, M.; Pinheiro, H. Bioreactor monitoring with spectroscopy and chemometrics: A review. Anal. Bioanal. Chem. 2012, 404, 1211–1237. [Google Scholar] [CrossRef]
- Parawira, W.; Murto, M.; Read, J.S.; Mattiasson, B. Volatile fatty acid production during anaerobic mesophilic digestion of solid potato waste. J. Chem. Technol. Biotechnol. 2004, 79, 673–677. [Google Scholar] [CrossRef]
- Mesquita, D.; Quintelas, C.; Amaral, A.L.P.D.; Ferreira, E.C. Monitoring biological wastewater treatment processes: Recent advances in spectroscopy applications. Rev. Environ. Sci. Bio/Technol. 2017, 16, 395–424. [Google Scholar] [CrossRef] [Green Version]
- Choubert, J.M.; Rieger, L.; Shaw, A.; Copp, J.; Spérandio, M.; Sorensen, K.; Rönner-Holm, S.; Morgenroth, E.; Melcer, H.; Gillot, S. Rethinkning wastewater characterization methods for activated sludge systems. Water Sci. Technol. 2013, 67, 2363–2373. [Google Scholar] [CrossRef] [Green Version]
- Janhom, T.; Wattanachira, S.; Pavasant, P. Characterization of brewery wastewater with spectrofluorometry analysis. J. Environ. Manag. 2009, 90, 1184–1190. [Google Scholar] [CrossRef]
- Maya-Altamira, L.; Baun, A.; Angelidaki, I.; Schmidt, J. Influence of wastewater characteristics on methane potential in food-processing industry wastewaters. Water Res. 2008, 42, 2195–2203. [Google Scholar] [CrossRef] [PubMed]
- Flores, G.A.E.; Fotidis, I.A.; Karakashev, D.B.; Kjellberg, K.; Angelidaki, I. Effects of Benzalkonium Chloride, Proxel LV, P3 Hypochloran, Triton X-100 and DOWFAX 63N10 on anaerobic digestion processes. Bioresour. Technol. 2015, 193, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Cui, F.; Zeng, G.-M.; Jiang, M.; Yang, Z.-Z.; Yu, Z.-G.; Zhu, M.-Y.; Shen, L.-Q. Quaternary ammonium compounds (QACs): A review on occurrence, fate and toxicity in the environment. Sci. Total Environ. 2015, 518–519, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Tezel, U.; Li, K.; Liu, D.; Ren, R.; Du, J.; Pavlostathis, S.G. Evaluation and modeling of benzalkonium chloride inhibition and biodegradation in activated sludge. Water Res. 2011, 45, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Tezel, U.; Pavlostathis, S.G. Transformation of Benzalkonium Chloride under Nitrate Reducing Conditions. Environ. Sci. Technol. 2009, 43, 1342–1348. [Google Scholar] [CrossRef]
- Hajaya, M.G.; Pavlostathis, S.G. Fate and effect of benzalkonium chloride in continuous-flow biological nitrogen removal system treating poultry processing water. Bioresour. Technol. 2012, 118, 73–81. [Google Scholar] [CrossRef]
- Tezel, U.; Pierson, J.A.; Pavlostathis, S.G. Fate and effect of quaternary ammonium compounds on a mixed methanogenic culture. Water Res. 2006, 40, 3660–3668. [Google Scholar] [CrossRef]
- Tezel, U.; Pierson, J.A.; Pavlostathis, S.G. Effect of dodecyl dimethyl ammonium chloride on nitrate reduction in a mixed methanogenic culture. Water Sci. Technol. 2008, 57, 541–546. [Google Scholar] [CrossRef] [Green Version]
- Sütterlin, H.; Alexy, R.; Kümmerer, K. The toxicity of the quaternary ammonium compound benzalkonium chloride alone and in mixtures with other anionic compounds to bacteria in test systems with Vibrio fischeri and Pseudomonas putida. Ecotoxicol. Environ. Saf. 2008, 71, 498–505. [Google Scholar] [CrossRef]
- Garcı́a, M.; Campos, E.; Sánchez-Leal, J.; Ribosa, I. Anaerobic degradation and toxicity of commercial cationic surfactants in anaerobic screening tests. Chemosphere 2000, 41, 705–710. [Google Scholar] [CrossRef]
- Li, S.; Zhao, S. Spectrophotometric determination of cationic surfactants with benzothiaxolyldiazoaminoazobenzene. Anal. Chim. Acta 2004, 501, 99–102. [Google Scholar] [CrossRef]
- Yamamoto, K.; Oka, M.; Murakami, H. Spectrophotometric determination of trace ionic and non-ionic surfactants based on a collection on a membrane filter as the ion associate of the surfactant with Erythrosine B. Anal. Chim. Acta 2002, 455, 83–92. [Google Scholar] [CrossRef]
- Ding, W.-H.; Liao, Y.-H. Determination of Alkylbenzyldimethylammonium Chlorides in River Water and Sewage Effluent by Solid-Phase Extraction and Gas Chromatography/Mass Spectrometry. Anal. Chem. 2000, 73, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Norberg, J.; Thordarson, E.; Mathiasson, L.; Jonsson, J.A. Microporous membrane liquid–liquid extraction coupled on-line with normal-phase liquid chromatography for the determination of cationic surfactants in river and wastewater. J. Chromatogr. A 2000, 869, 523–529. [Google Scholar] [CrossRef]
- Ding, W.-H.; Tsai, P.-C. Determination of Alkyltrimethylammonium Chlorides in River Water by Gas Chromatography/Ion Trap Mass Spectrometry with Electron Impact and Chemical Ionization. Anal. Chem. 2003, 75, 1792–1797. [Google Scholar] [CrossRef]
- Longstaffe, J.; Konzuk, J. NMR in the environmental industry. Magn. Reson. Chem. 2015, 53, 691–693. [Google Scholar] [CrossRef]
- Kostidis, S.; Addie, R.D.; Morreau, H.; Mayboroda, O.; Giera, M. Quantitative NMR analysis of intra- and extracellular metabolism of mammalian cells: A tutorial. Anal. Chim. Acta 2017, 980, 1–24. [Google Scholar] [CrossRef]
- Xue, J.; Isern, N.G.; Ewing, R.J.; Liyu, A.V.; Sears, J.A.; Knapp, H.; Iversen, J.; Sisk, D.R.; Ahring, B.K.; Majors, P.D. New generation NMR bioreactor coupled with high-resolution NMR spectroscopy leads to novel discoveries in Moorella thermo-acetica metabolic profile. Appl. Microbiol. Biotechnol. 2014, 98, 8367–8375. [Google Scholar] [CrossRef]
- Bai, L.; Carlton, D.D.; Schug, K.A. Complex mixture quantification without calibration using gas chromatography and a comprehensive carbon reactor in conjunction with flame ionization detection. J. Sep. Sci. 2018, 41, 4031–4037. [Google Scholar] [CrossRef]
- Pavia, D.L.; Lampman, G.M.; Kriz, G.S. Introduction to Spectroscopy, 3rd ed.; Thomson Learning Inc.: Bellingham, WA, USA, 2001. [Google Scholar]
- Botheju, D.; Bakke, R. Oxygen effects in anaerobic digestion—A review. Open Waste Manag. J. 2011, 4, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Theuerl, S.; Klang, J.; Prochnow, A. Process disturbances in agricultural biogas production-cause, mechanisms and effects in the biogas microbiome: A review. Energies 2019, 12, 365. [Google Scholar] [CrossRef] [Green Version]
- McCarty, P.L. One hundred years of anaerobic treatment. In Anaerobic Digestion; Hughes, D.E., Ed.; Elsevier Biomedical Press: Amsterdam, The Netherlands, 1982; pp. 3–22. [Google Scholar]
- Mittal, K.M. Biogas Systems: Policies, Progress and Prospects; New Age International: New Delhi, India, 1997. [Google Scholar]
- Bryant, M.P. Microbial Methane Production—Theoretical Aspects2. J. Anim. Sci. 1979, 48, 193–201. [Google Scholar] [CrossRef]
- Schink, B. Energetics of syntrophic cooperation in methanogenic degradation. Microbiol. Mol. Biol. Rev. 1997, 61, 262–280. [Google Scholar] [PubMed]
- Tretter, L.; Patocs, A.; Chinopoulos, C. Succinate, an intermediate in metabolism, signal transduction, ROS, hypoxia, and tumorigenesis. Biochim. Biophys. Acta. 2016, 1857, 1086–1101. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Lee, S.Y. Production of succinic acid by bacterial fermentation. Enzyme Microb. Technol. 2006, 39, 352–361. [Google Scholar] [CrossRef]
- Garcia, M.T.; Campos, E.; Sanchez-Leal, J.; Ribosa, I. Effect of the alkyl chain length on the anaerobic biodegradability and toxicity of quaternary ammonium based surfactants. Chemosphere 1999, 38, 3473–3483. [Google Scholar] [CrossRef]
- Durham, J.; Young, J.C. Toxic Impact of Commercial Biocides on Industrial Wastewater Treatment Systems. Proc. Water Environ. Fed. 2009, 2009, 5670–5689. [Google Scholar] [CrossRef]
- Tezel, U.; Pierson, J.A.; Pavlostathis, S.G. Effect of polyelec-trolytes and quaternary ammonium compounds on the anaerobic biological treatment of poultry processing wastewater. Water Res. 2007, 41, 1334–1342. [Google Scholar] [CrossRef]
- Chen, J.L.; Ortiz, R.; Steele, T.W.; Stuckey, D.C. Toxicants inhibiting anaerobic digestion: A review. Biotechnol. Adv. 2014, 32, 1523–1534. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Compound | ID | Compound | ID | Compound | ID | Compound |
|---|---|---|---|---|---|---|---|
| Fatty Acids | Amino Acids | Alcohols | Auxins | ||||
| 1 | Formic Acid | 10 | Glycine | 20 | Ethanol | 28 | Indole-3-Acetate |
| 2 | Acetic Acid | 11 | Alanine | 21 | Ethylene Glycol | Disinfecting Agents | |
| 3 | Propionic Acid | 12 | Threonine | 22 | Propylene Glycol | 29 | Benzalkonium Chloride |
| 4 | Butyric Acid | 13 | Glutamine | Amines | |||
| 5 | Isobutryic Acid | 14 | Methionine | 23 | Methyl Amine | ||
| 6 | Valeric Acid | 15 | Valine | Amides | |||
| 7 | Isovaleric Acid | 16 | Isoleucine | 24 | Acetamide | ||
| 8 | Succinic Acid | 17 | Tyrosine | Pyrimidines | |||
| Fatty Acid Metabolites | 18 | Phenylalanine | 25 | Uracil | |||
| 9 | Phenylacetate | 19 | Tryptophan |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freemantle, R.; Butson, N.; Goodfellow, J.; Konzuk, J.; Longstaffe, J.G. Nuclear Magnetic Resonance Spectroscopy Analysis of Anaerobic Microbial Metabolic Response to Benzalkonium Chloride Disinfectant. Appl. Sci. 2022, 12, 4620. https://0-doi-org.brum.beds.ac.uk/10.3390/app12094620
Freemantle R, Butson N, Goodfellow J, Konzuk J, Longstaffe JG. Nuclear Magnetic Resonance Spectroscopy Analysis of Anaerobic Microbial Metabolic Response to Benzalkonium Chloride Disinfectant. Applied Sciences. 2022; 12(9):4620. https://0-doi-org.brum.beds.ac.uk/10.3390/app12094620
Chicago/Turabian StyleFreemantle, Ryan, Nick Butson, Janet Goodfellow, Julie Konzuk, and James G. Longstaffe. 2022. "Nuclear Magnetic Resonance Spectroscopy Analysis of Anaerobic Microbial Metabolic Response to Benzalkonium Chloride Disinfectant" Applied Sciences 12, no. 9: 4620. https://0-doi-org.brum.beds.ac.uk/10.3390/app12094620